Development of an Image-Guided Orthotopic Xenograft Mouse Model of Endometrial Cancer with Controllable Estrogen Exposure

 , ,
, ,  , ,
, , 
Abstract
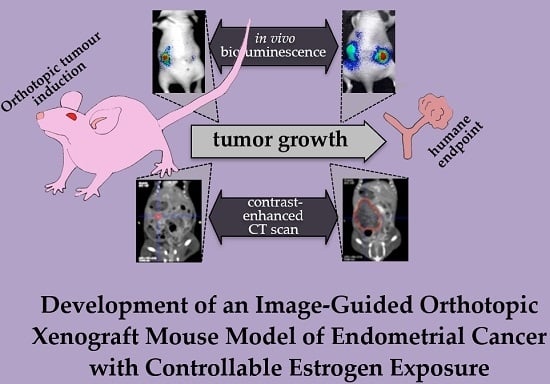
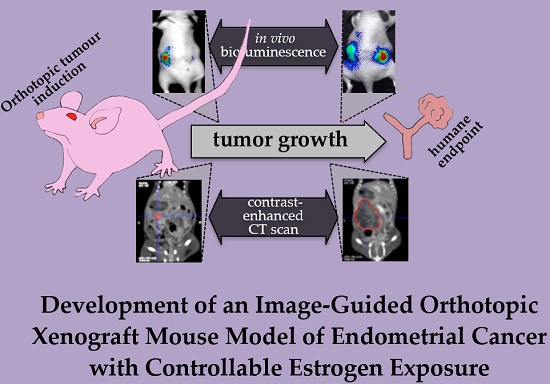
1. Introduction
2. Results
2.1. Orthotopic EC Xenograft: Cell Titre Optimization
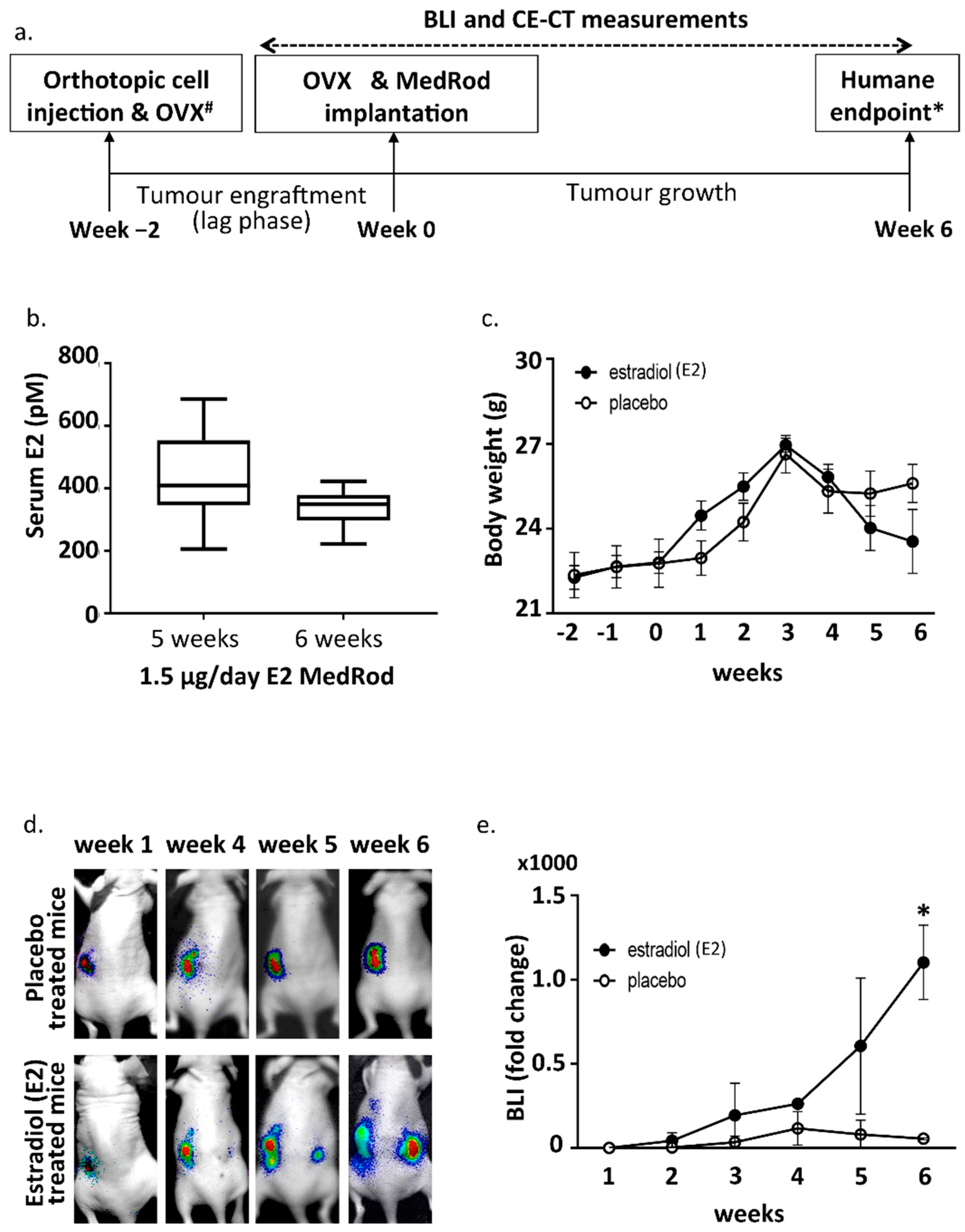
2.2. Ovariectomy and Estrogen Supplementation (MedRod Implants)
2.3. Estrogen Responsiveness of the Orthotopic EC Xenograft
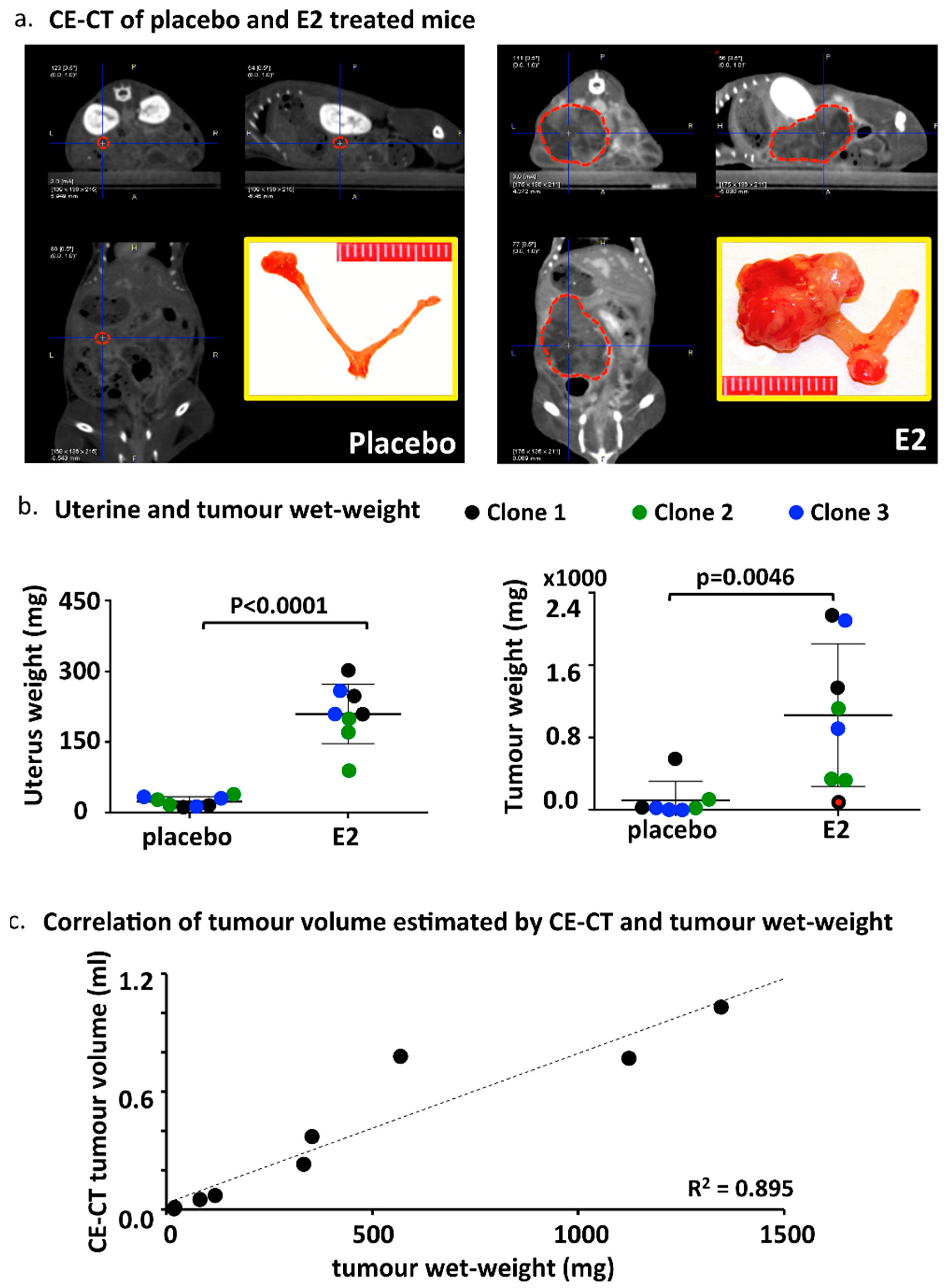
2.4. CE-CT, Endpoint and Tumor Characteristics
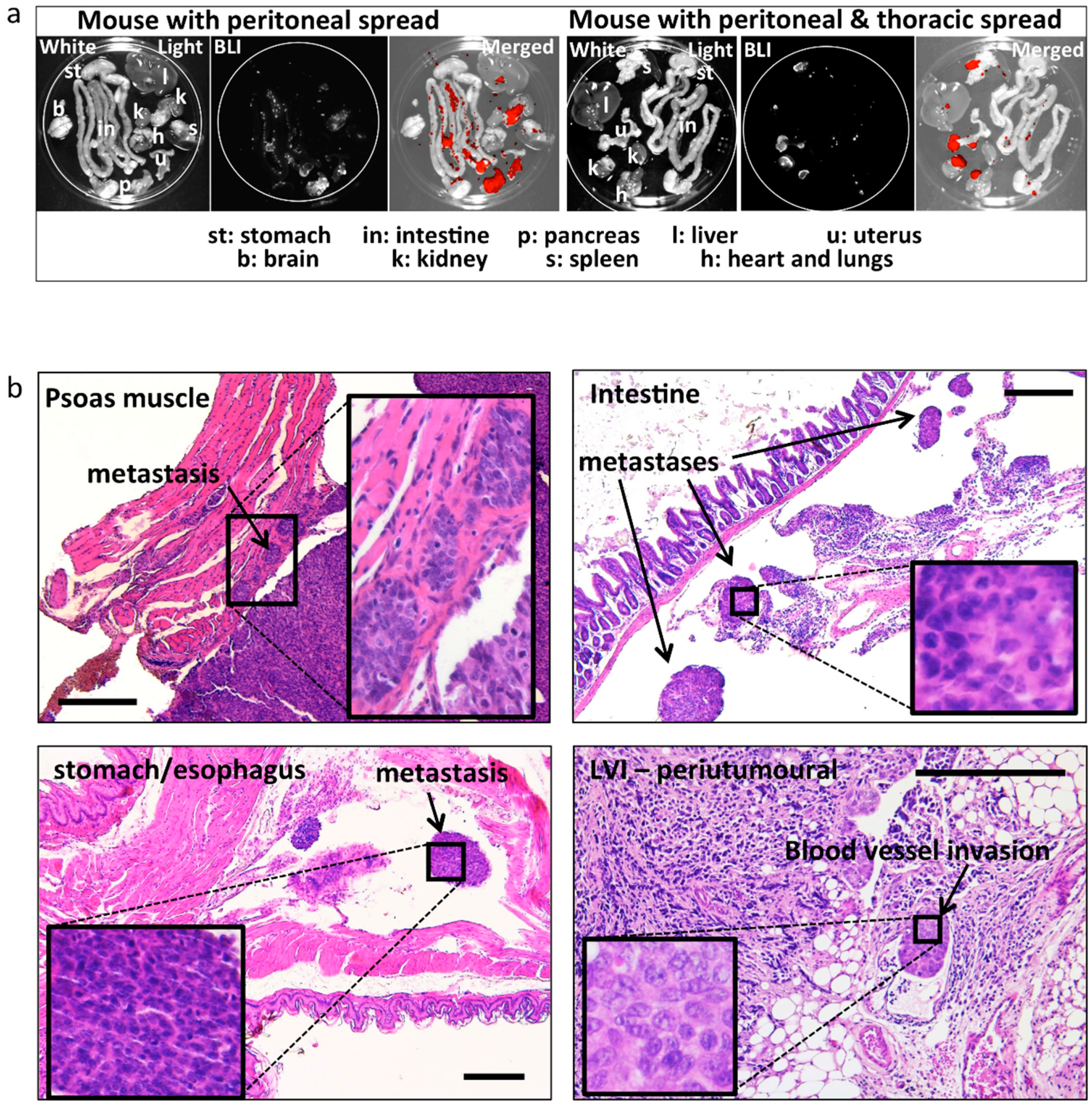
2.5. Metastases
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cell Lines and Tumor Graft Preparation
4.3. Optimization of the MedRod Steroid Delivery System
4.4. Orthotopic Estrogen Dependent Endometrial Cancer Mouse Model
4.5. Imaging by BLI
4.6. Micro Contrast Enhanced-Computed Tomography (CE-CT)
4.7. Histological Examination and Immunohistochemistry
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BLI | Bioluminescence imaging |
| CAM | Chorioallantoic membrane assay |
| CE-CT | Contrast-enhanced computerised axial tomography |
| E2 | 17β-Estradiol |
| EC | Endometrial cancer |
| GFP | Green fluorescent protein |
| LC-MS/MS | Liquid chromatography tandem mass spectrometry |
| LVI | Lymphovascular invasion |
| OVX | Ovariectomy |
| ROI | Region of Interest |
References
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial cancer. Lancet 2015, 387, 1094–1108. [Google Scholar] [CrossRef]
- Vale, C.L.; Tierney, J.; Bull, S.J.; Symonds, P.R. Chemotherapy for advanced, recurrent or metastatic endometrial carcinoma. Cochrane Database Syst. Rev. 2012, 8, CD003915. [Google Scholar] [CrossRef] [PubMed]
- Decruze, S.B.; Green, J.A. Hormone therapy in advanced and recurrent endometrial cancer: A systematic review. Int. J. Gynecol. Cancer 2007, 17, 964–978. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, K.; Malander, S.; Christensen, R.D.; Mirza, M.R.; Kristensen, G.B.; Aavall-Lundqvist, E.; Vergote, I.; Rosenberg, P.; Boman, K.; Nordstrom, B. Examestane in advanced or recurrent endometrial carcinoma: A prospective phase II study by the Nordic Society of Gynecologic Oncology (NSGO). BMC Cancer 2014, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.G.; Brunetto, V.L.; VanLe, L.; Bell, J.; Walker, J.L.; Lee, R.B. A phase II trial of anastrozole in advanced recurrent or persistent endometrial carcinoma: A Gynecologic Oncology Group study. Gynecol. Oncol. 2000, 78, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Vandenput, I.; Van Eygen, K.; Moerman, P.; Vergote, I.; Amant, F. Ineffective attempt to preserve fertility with a levonorgestrel-releasing intrauterine device in a young woman with endometrioid endometrial carcinoma: A case report and review of the literature. Eur. J. Gynaecol. Oncol. 2009, 30, 313–316. [Google Scholar] [PubMed]
- Slomovitz, B.M.; Jiang, Y.; Yates, M.S.; Soliman, P.T.; Johnston, T.; Nowakowski, M.; Levenback, C.; Zhang, Q.; Ring, K.; Munsell, M.F.; et al. Phase II study of everolimus and letrozole in patients with recurrent endometrial carcinoma. J. Clin. Oncol. 2015, 33, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Tangen, I.L.; Onyango, T.B.; Kopperud, R.; Berg, A.; Halle, M.K.; Oyan, A.M.; Werner, H.M.; Trovik, J.; Kalland, K.H.; Salvesen, H.B.; et al. Androgen receptor as potential therapeutic target in metastatic endometrial cancer. Oncotarget 2016, 7, 49289–49298. [Google Scholar] [CrossRef] [PubMed]
- Sinreih, M.; Knific, T.; Anko, M.; Hevir, N.; Vouk, K.; Jerin, A.; Frkovic Grazio, S.; Rizner, T.L. The Significance of the Sulfatase Pathway for Local Estrogen Formation in Endometrial Cancer. Front. Pharmacol. 2017, 8, 368. [Google Scholar] [CrossRef] [PubMed]
- Konings, G.F.; Cornel, K.M.; Xanthoulea, S.; Delvoux, B.; Skowron, M.A.; Kooreman, L.; Koskimies, P.; Krakstad, C.; Salvesen, H.B.; van Kuijk, K.; et al. Blocking 17β-hydroxysteroid dehydrogenase type 1 in endometrial cancer: A potential novel endocrine therapeutic approach. J. Pathol. 2018, 244, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Gizzo, S.; Noventa, M.; Di Gangi, S.; Litta, P.; Saccardi, C.; D’Antona, D.; Nardelli, G.B. Could in-vitro studies on Ishikawa cell lines explain the endometrial safety of raloxifene? Systematic literature review and starting points for future oncological research. Eur. J. Cancer Prev. 2015, 24, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.H.; O’Donnell, A.L.; Mohamed, S.; Mousa, S.; Dandona, P. Overexpression of estrogen receptor-alpha in the endometrial carcinoma cell line Ishikawa: Inhibition of growth and angiogenic factors. Gynecol. Oncol. 2004, 95, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, S.; Llaurado, M.; Castellvi, J.; Fernandez, Y.; Alameda, F.; Colas, E.; Ruiz, A.; Doll, A.; Schwartz, S., Jr.; Carreras, R.; et al. Generation and characterization of orthotopic murine models for endometrial cancer. Clin. Exp. Metastasia 2012, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Haldorsen, I.S.; Popa, M.; Fonnes, T.; Brekke, N.; Kopperud, R.; Visser, N.C.; Rygh, C.B.; Pavlin, T.; Salvesen, H.B.; McCormack, E.; et al. Multimodal Imaging of Orthotopic Mouse Model of Endometrial Carcinoma. PLoS ONE 2015, 10, e0135220. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Ogawa, H.; Izumi, K.; Uehara, H. The soluble EP2 receptor FuEP2/Ex2 suppresses endometrial cancer cell growth in an orthotopic xenograft model in nude mice. Cancer Lett. 2011, 306, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Liu, B.Y.; Liao, Y.; Zhang, H.J.; Yang, T.T.; He, Y.Y.; Xia, Y.H.; Lu, W.; He, X.Y.; Chen, Z.; et al. Activation of a positive feedback loop involving IL-6 and aromatase promotes intratumoral 17beta-estradiol biosynthesis in endometrial carcinoma microenvironment. Int. J. Cancer 2014, 135, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Foster, P.A.; Woo, L.W.; Potter, B.V.; Reed, M.J.; Purohit, A. The use of steroid sulfatase inhibitors as a novel therapeutic strategy against hormone-dependent endometrial cancer. Endocrinology 2008, 149, 4035–4042. [Google Scholar] [CrossRef] [PubMed]
- Husen, B.; Huhtinen, K.; Poutanen, M.; Kangas, L.; Messinger, J.; Thole, H. Evaluation of inhibitors for 17β-hydroxysteroid dehydrogenase type 1 in vivo in immunodeficient mice inoculated with MCF-7 cells stably expressing the recombinant human enzyme. Mol. Cell. Endocrinol. 2006, 248, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Husen, B.; Huhtinen, K.; Saloniemi, T.; Messinger, J.; Thole, H.H.; Poutanen, M. Human hydroxysteroid (17-β) dehydrogenase 1 expression enhances estrogen sensitivity of MCF-7 breast cancer cell xenografts. Endocrinology 2006, 147, 5333–5339. [Google Scholar] [CrossRef] [PubMed]
- Chitcholtan, K.; Sykes, P.H.; Evans, J.J. The resistance of intracellular mediators to doxorubicin and cisplatin are distinct in 3D and 2D endometrial cancer. J. Transl. Med. 2012, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, G. Endometrial cancer: Experimental models useful for studies on molecular aspects of endometrial cancer and carcinogenesis. Endocr. Relat. Cancer 2003, 10, 23–42. [Google Scholar] [CrossRef] [PubMed]
- Skowron, M.A.; Sathe, A.; Romano, A.; Hoffmann, M.J.; Schulz, W.A.; van Koeveringe, G.A.; Albers, P.; Nawroth, R.; Niegisch, G. Applying the chicken embryo chorioallantoic membrane assay to study treatment approaches in urothelial carcinoma. Urol. Oncol. 2017, 35, 544.e11–544.e23. [Google Scholar] [CrossRef] [PubMed]
- Ingberg, E.; Theodorsson, A.; Theodorsson, E.; Strom, J.O. Methods for long-term 17β-estradiol administration to mice. Gen. Comp. Endocrinol. 2012, 175, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Saloniemi, T.; Jarvensivu, P.; Koskimies, P.; Jokela, H.; Lamminen, T.; Ghaem-Maghami, S.; Dina, R.; Damdimopoulou, P.; Makela, S.; Perheentupa, A.; et al. Novel hydroxysteroid (17β) dehydrogenase 1 inhibitors reverse estrogen-induced endometrial hyperplasia in transgenic mice. Am. J. Pathol. 2010, 176, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, L.; Shepherd, L.; Torrado, A.I.; Torres-Diaz, Y.M.; Miranda, J.D.; Segarra, A.C. Comparison of Two Methods of Estradiol Replacement: Their Physiological and Behavioral Outcomes. J. Vet. Sci. Technol. 2015, 6, 276. [Google Scholar] [CrossRef] [PubMed]
- Dall, G.; Vieusseux, J.; Unsworth, A.; Anderson, R.; Britt, K. Low Dose, Low Cost Estradiol Pellets Can Support MCF-7 Tumour Growth in Nude Mice without Bladder Symptoms. J. Cancer 2015, 6, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, A.E.; Ryan, N.; Crosbie, E.J. Biomarkers needed to predict progestin response in endometrial cancer. BJOG 2017, 124, 1584. [Google Scholar] [CrossRef] [PubMed]
- Rizner, T.L. The Important Roles of Steroid Sulfatase and Sulfotransferases in Gynecological Diseases. Front. Pharmacol. 2016, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Cornel, K.M.; Kruitwagen, R.F.; Delvoux, B.; Visconti, L.; Van de Vijver, K.K.; Day, J.M.; Van Gorp, T.; Hermans, R.J.; Dunselman, G.A.; Romano, A. Overexpression of 17β-Hydroxysteroid Dehydrogenase Type 1 Increases the Exposure of Endometrial Cancer to 17β-Estradiol. J. Clin. Endocrinol. Metab. 2012, 97, E591–E601. [Google Scholar] [CrossRef] [PubMed]
- Depreeuw, J.; Hermans, E.; Schrauwen, S.; Annibali, D.; Coenegrachts, L.; Thomas, D.; Luyckx, M.; Gutierrez-Roelens, I.; Debruyne, D.; Konings, K.; et al. Characterization of patient-derived tumor xenograft models of endometrial cancer for preclinical evaluation of targeted therapies. Gynecol. Oncol. 2015, 139, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hakkinen, M.R.; Heinosalo, T.; Saarinen, N.; Linnanen, T.; Voutilainen, R.; Lakka, T.; Jaaskelainen, J.; Poutanen, M.; Auriola, S. Analysis by LC-MS/MS of endogenous steroids from human serum, plasma, endometrium and endometriotic tissue. J. Pharm. Biomed. Anal. 2018, 152, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, R.; Lindsay, P.E.; Ansell, S.; Wilson, G.; Jelveh, S.; Hill, R.P.; Jaffray, D.A. Characterization of image quality and image-guidance performance of a preclinical microirradiator. Med. Phys. 2011, 38, 845–856. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Injected Cells (Number) | Number of Mice | Mice with Tumor (%) | Peritoneal Metastases * | Distant Metastases # |
|---|---|---|---|---|
| 1 × 106 | 3 | 2 (67%) | I | 0 |
| 3 × 106 | 3 | 3 (100%) | I, L, K, S, St | 1 (T + LVI) |
| 5 × 106 | 3 | 3 (100%) | I, L, K, S, St | 0 |
| Ishikawa (3 × 103 Cells) | OVX | Placebo/E2 | Number of Mice | Mice with Tumor | CE-CT | Peritoneal Metastases * | LVI # (No.) |
|---|---|---|---|---|---|---|---|
| Clone 1 | + | Placebo | 3 | 3 (100%) | 3 | I, L, K, S (P, A) | 0 |
| + | E2 | 3 | 3 (100%) | 2 | I, L, K, S, St (P, A) | 3 | |
| Clone 2 | + | Placebo | 2 | 2 (100%) | 2 | I, L, K, St (A) | 1 |
| + | E2 | 3 | 3 (100%) | 3 | I, L, K, S, St (P, A) | 2 | |
| Clone 3 | + | Placebo | 3 | 3 (100%) | 0 | (A, P) | 0 |
| + | E2 | 2 | 2 (100%) | 0 | (A) | 2 |
| Ishikawa | Placebo/E2 | x (cm) 1 | y (cm) 2 | z (cm) 3 | Tumor Volume (mL) 4 | Tumor Density ± SD 5 | Tumor Weight * |
|---|---|---|---|---|---|---|---|
| Clone 1 | placebo | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Clone 1 | placebo | 0.27 | 0.23 | 0.27 | 0.01 | 131.7 ± 15.6 | 20 |
| Clone 1 | placebo | 0.63 | 0.53 | 0.41 | 0.07 | −9.3 ± 4.9 | 118 |
| Clone 2 | placebo | 0.14 | 0.14 | 0.23 | 0.002 | 137.3 ± 25.9 | 18 |
| Clone 2 | placebo | 1.18 | 0.9 | 1.47 | 0.78 | 107.8 ± 4.5 | 568 |
| Clone 2 | E2 | 0.68 | 0.74 | 0.93 | 0.23 | 24.8 ± 4.7 | 333 |
| Clone 1 | E2 | 0.57 + 0.80 + 1.0 | 0.51 + 0.94 + 0.76 | 0.63 + 0.95 + 0.85 | 0.77 | 57.3 ± 8.0 | 1123 |
| Clone 1 | E2 | 0.78 | 0.98 | 0.97 | 0.37 | 31.2 ± 3.2 | 353 |
| Clone 2 | E2 | n.d. | n.d. | n.d. | n.d. | n.d. | 2147 |
| Clone 2 | E2 | 1.25 | 1.04 | 1.58 | 1.03 | 47.8 ± 15.2 | 1347 |
| Clone 2 | E2 | 0.49 | 0.45 | 0.43 | 0.05 | −6.3 ± 10.4 | 81 6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konings, G.F.; Saarinen, N.; Delvoux, B.; Kooreman, L.; Koskimies, P.; Krakstad, C.; Fasmer, K.E.; Haldorsen, I.S.; Zaffagnini, A.; Häkkinen, M.R.; et al. Development of an Image-Guided Orthotopic Xenograft Mouse Model of Endometrial Cancer with Controllable Estrogen Exposure. Int. J. Mol. Sci. 2018, 19, 2547. https://doi.org/10.3390/ijms19092547
Konings GF, Saarinen N, Delvoux B, Kooreman L, Koskimies P, Krakstad C, Fasmer KE, Haldorsen IS, Zaffagnini A, Häkkinen MR, et al. Development of an Image-Guided Orthotopic Xenograft Mouse Model of Endometrial Cancer with Controllable Estrogen Exposure. International Journal of Molecular Sciences. 2018; 19(9):2547. https://doi.org/10.3390/ijms19092547
Chicago/Turabian StyleKonings, Gonda FJ, Niina Saarinen, Bert Delvoux, Loes Kooreman, Pasi Koskimies, Camilla Krakstad, Kristine E. Fasmer, Ingfrid S. Haldorsen, Amina Zaffagnini, Merja R. Häkkinen, and et al. 2018. "Development of an Image-Guided Orthotopic Xenograft Mouse Model of Endometrial Cancer with Controllable Estrogen Exposure" International Journal of Molecular Sciences 19, no. 9: 2547. https://doi.org/10.3390/ijms19092547
APA StyleKonings, G. F., Saarinen, N., Delvoux, B., Kooreman, L., Koskimies, P., Krakstad, C., Fasmer, K. E., Haldorsen, I. S., Zaffagnini, A., Häkkinen, M. R., Auriola, S., Dubois, L., Lieuwes, N., Verhaegen, F., Schyns, L. E., Kruitwagen, R. F., ENITEC Consortium, Xanthoulea, S., & Romano, A. (2018). Development of an Image-Guided Orthotopic Xenograft Mouse Model of Endometrial Cancer with Controllable Estrogen Exposure. International Journal of Molecular Sciences, 19(9), 2547. https://doi.org/10.3390/ijms19092547