Heme Oxygenase-1 Inhibitors Induce Cell Cycle Arrest and Suppress Tumor Growth in Thyroid Cancer Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
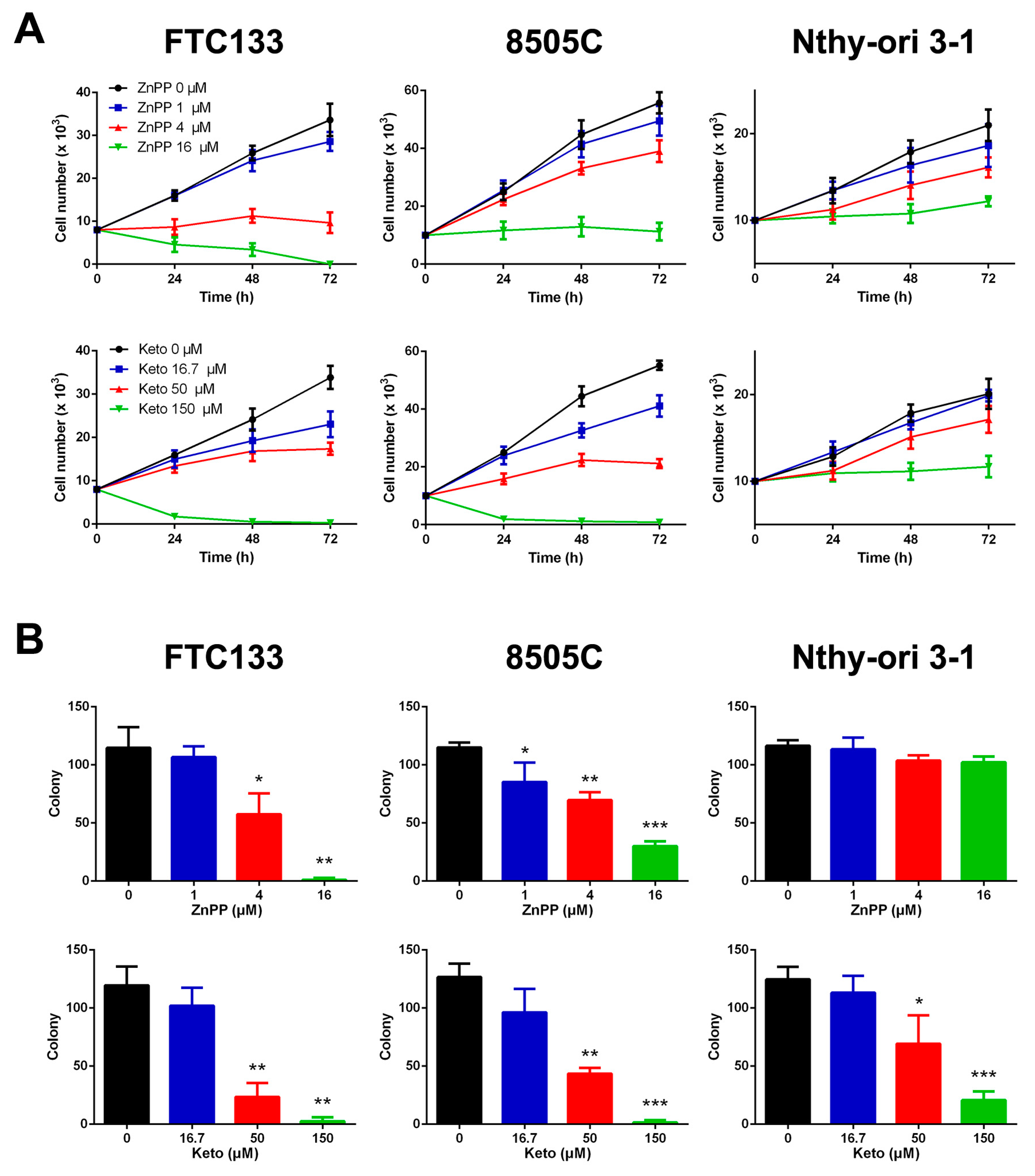
2.1. Suppression of Cell Viability and Clonogenic Ability by HO-1 Inhibitors
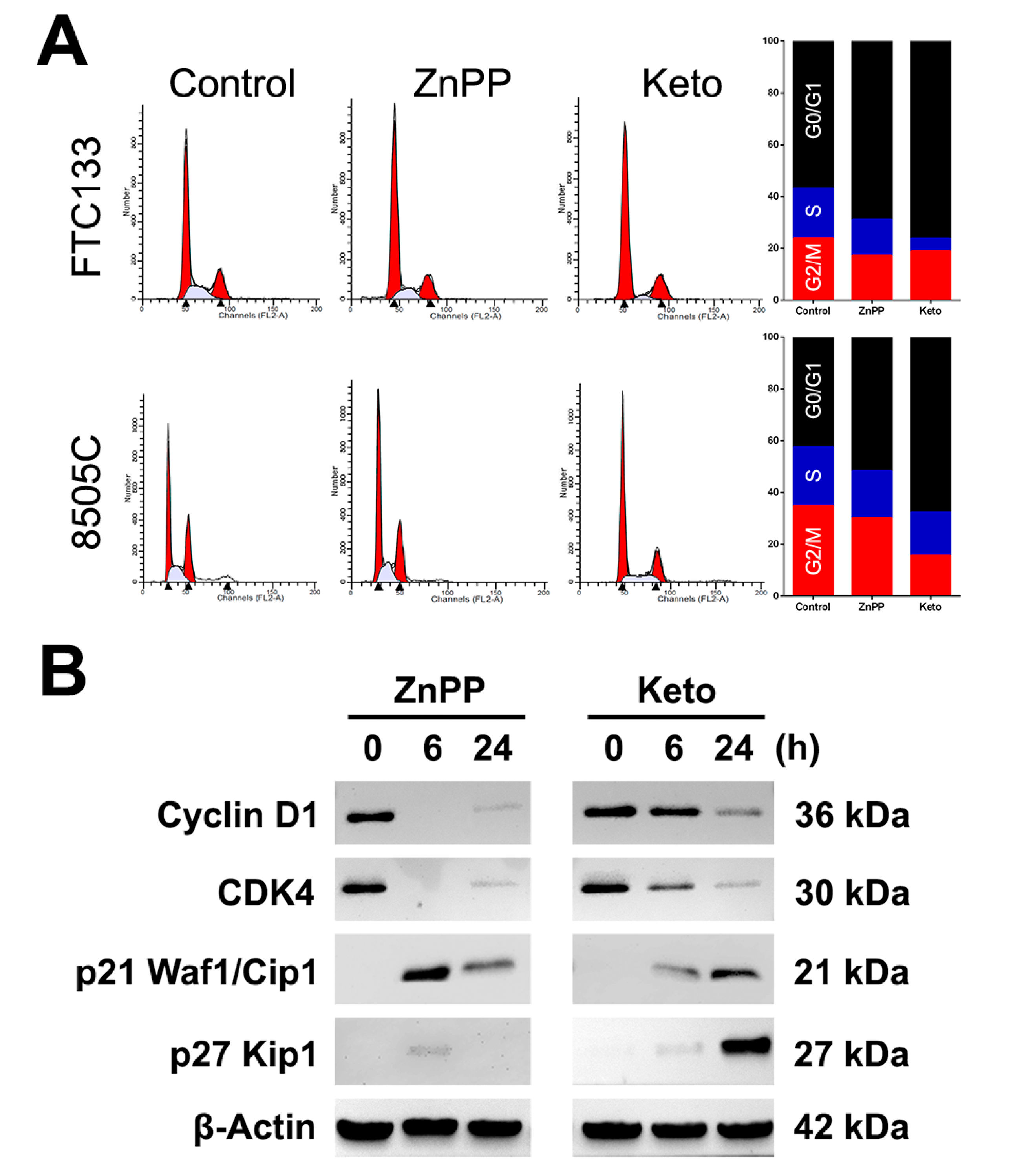
2.2. Cell Cycle Arrest Induced by HO-1 Inhibitors
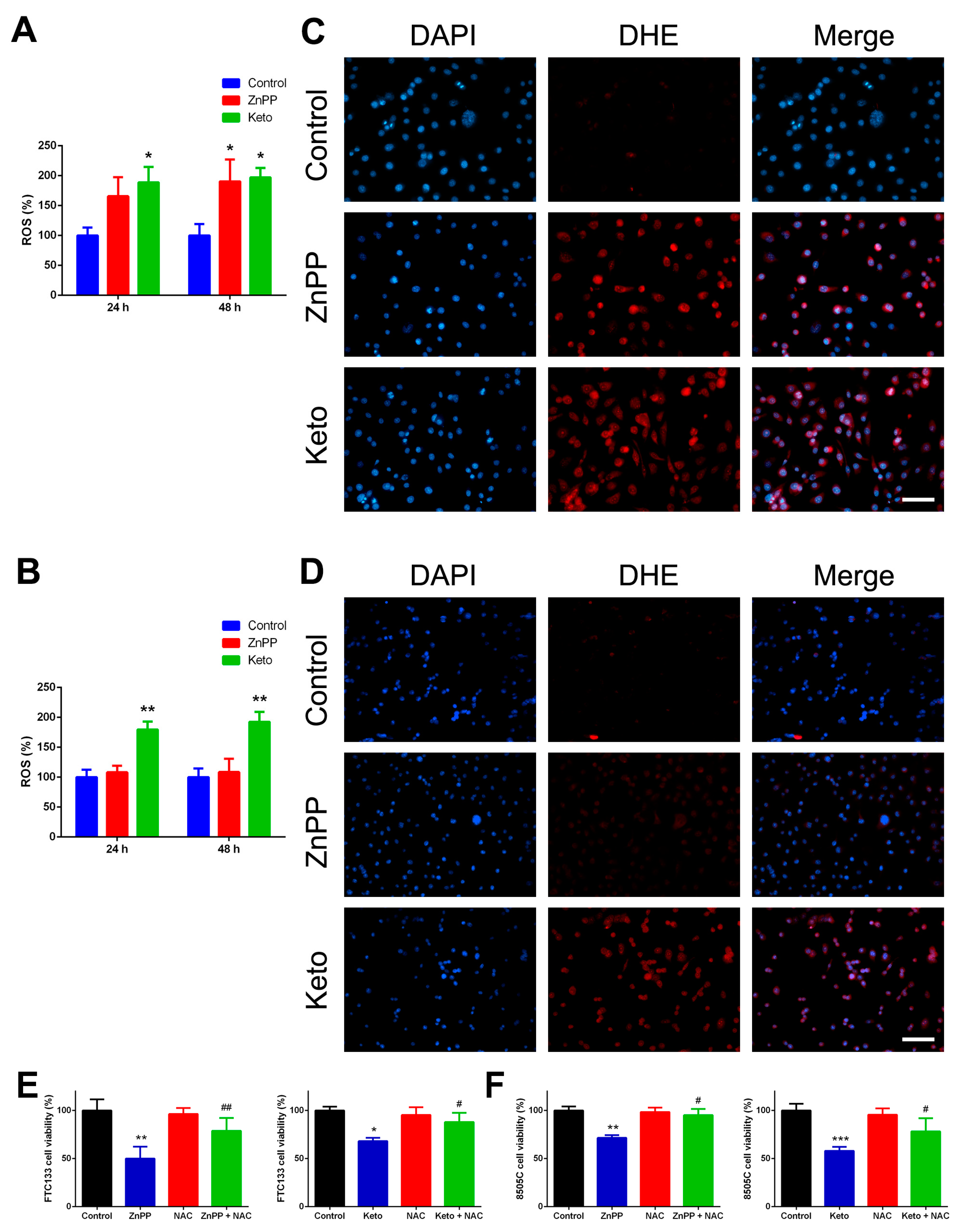
2.3. ROS Induction by HO-1 Inhibitors
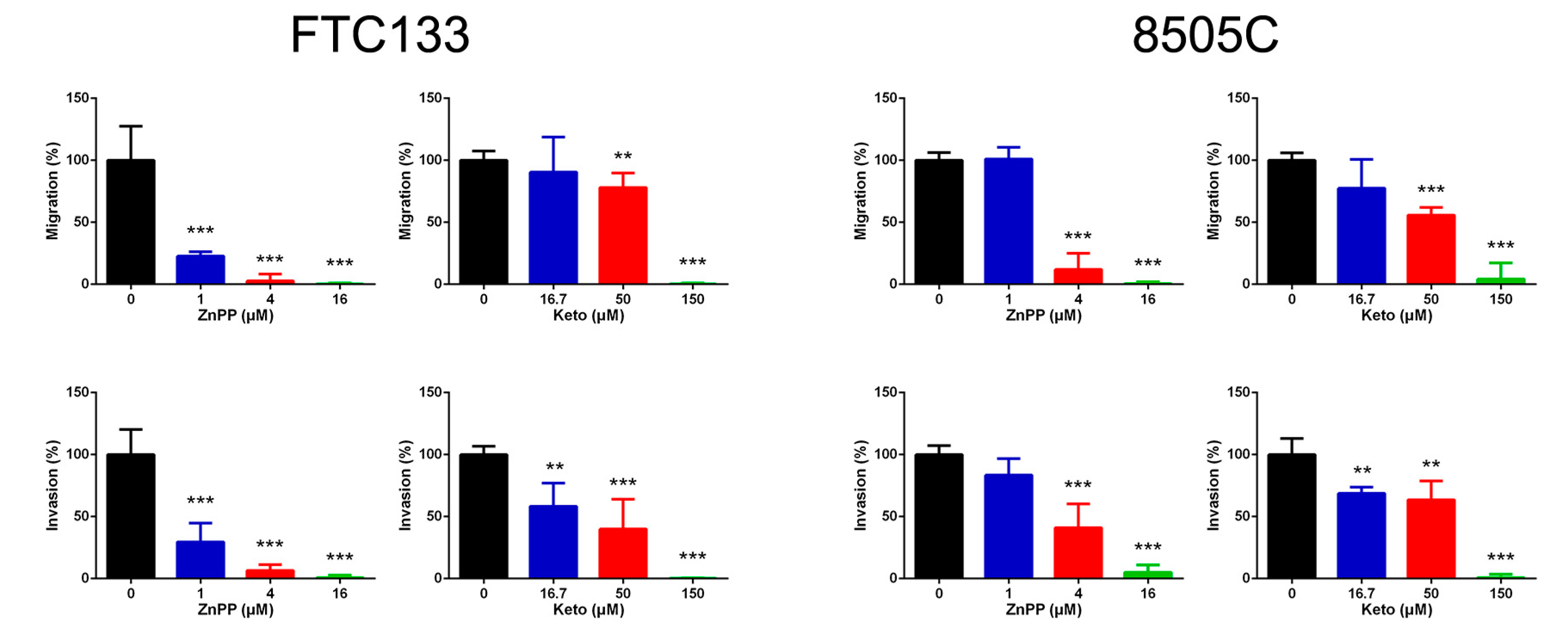
2.4. Suppression of Cell Migration and Invasion by HO-1 Inhibitors
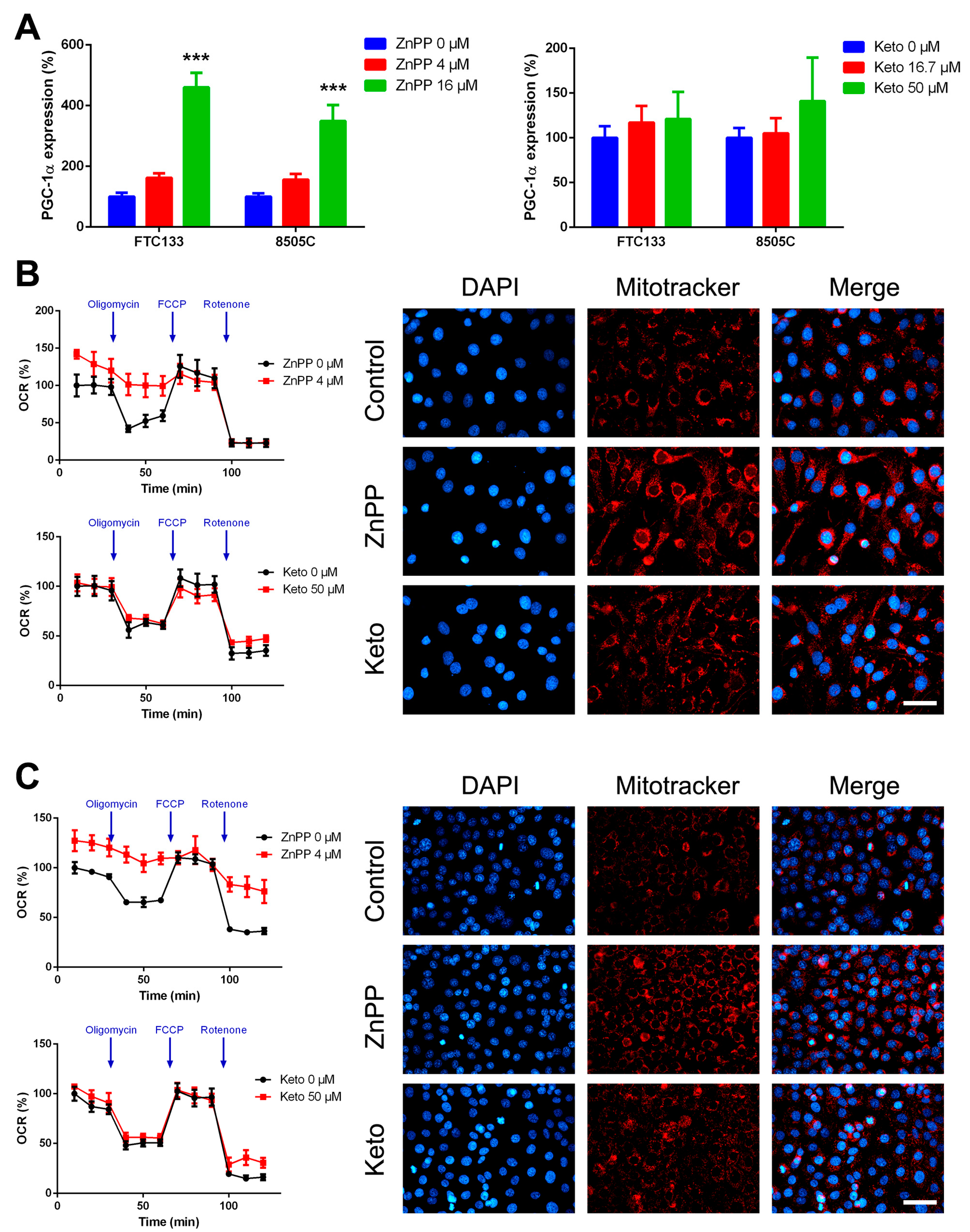
2.5. Increased Mitochondrial Mass and Oxygen Consumption by HO-1 Inhibitors
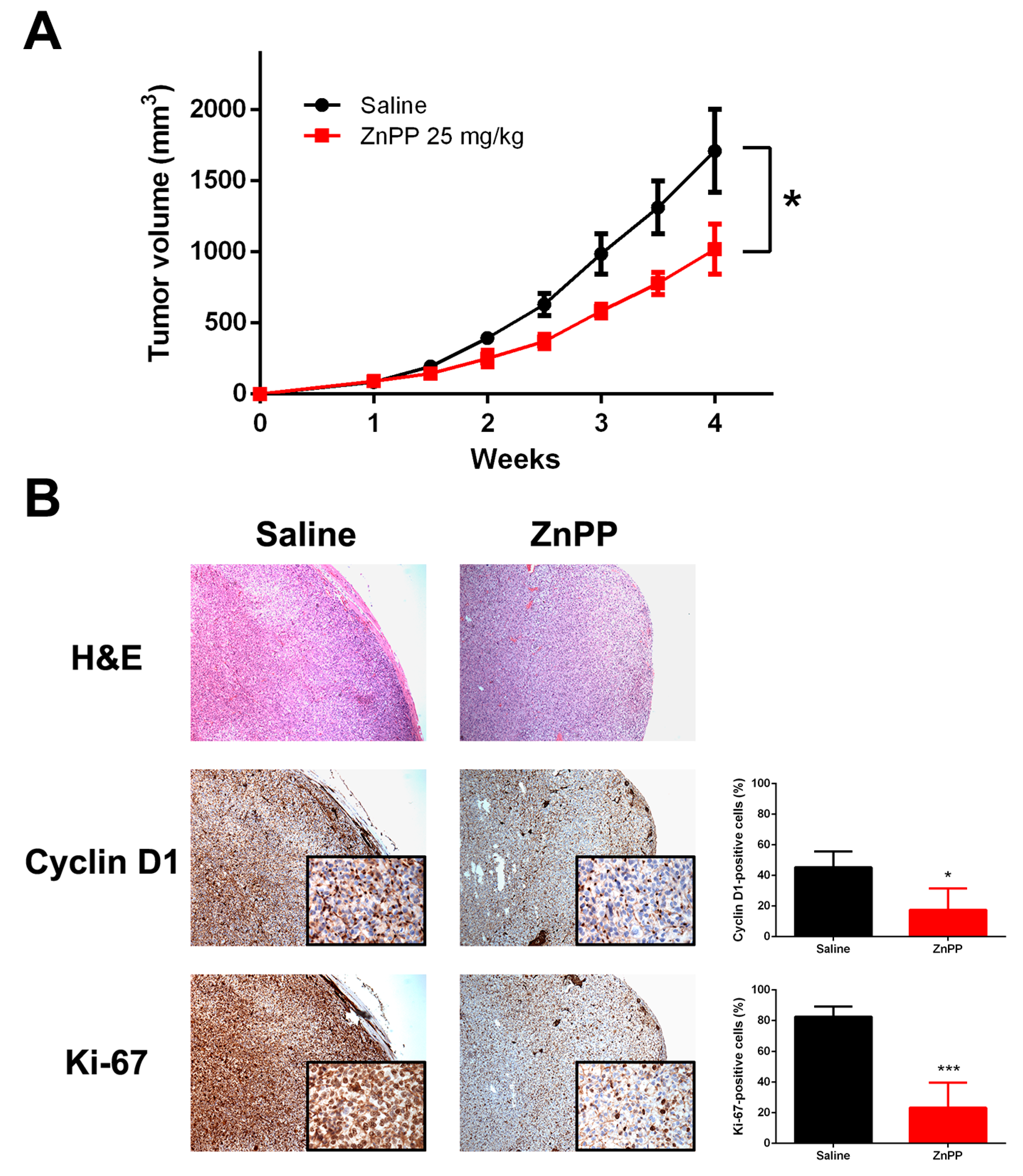
2.6. Delayed Xenograft Tumor Growth with in Vivo ZnPP Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Reagents
4.2. Cell Growth and Colony Formation
4.3. Reverse-Transcription Polymerase Chain Reaction
4.4. Cell Cycle Analysis
4.5. Western Blotting
4.6. ROS Detection Assays
4.7. Transwell Migration and Invasion Assays
4.8. Oxygen Consumption
4.9. MitoTracker Red Staining
4.10. Xenograft Implantation
4.11. Immunohistochemistry
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HO-1 | heme oxygenase-1 |
| ROS | reactive oxygen species |
| ZnPP | zinc protoporphyrin-IX |
| IC50 | The concentration causes 50% inhibition |
| DHE | dihydroethidium |
| NAC | N-Acetyl-l-cysteine |
| PGC-1α | peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| OCR | oxygen consumption rate |
| ECAR | extracellular acidification rate |
| FBS | fetal bovine serum |
| DCFDA | 2′,7′-dichlorofluorescein diacetate |
| DAPI | 4′,6-diamidino-2-phenylindole |
| H&E | hematoxylin and eosin |
References
- Motterlini, R.; Foresti, R. Heme oxygenase-1 as a target for drug discovery. Antioxid. Redox Signal. 2014, 20, 1810–1826. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HO-1/CO system in tumor growth, angiogenesis and metabolism—Targeting HO-1 as an anti-tumor therapy. Vascul. Pharmacol. 2015, 74, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Li Volti, G.; Tibullo, D.; Vanella, L.; Giallongo, C.; Di Raimondo, F.; Forte, S.; Di Rosa, M.; Signorelli, S.S.; Barbagallo, I. The heme oxygenase system in hematological malignancies. Antioxid. Redox Signal. 2017, 27, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.W.; Park, J.Y.; Sung, J.Y.; Kwak, S.H.; Yu, J.; Chang, J.H.; Kim, J.H.; Ha, S.Y.; Paik, E.K.; Lee, W.S.; et al. Genomic evidence of reactive oxygen species elevation in papillary thyroid carcinoma with Hashimoto thyroiditis. Endocr. J. 2015, 62, 857–877. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.P.; Chen, M.J.; Chien, M.N.; Lin, C.H.; Lee, J.J.; Liu, C.L. Overexpression of teneurin transmembrane protein 1 is a potential marker of disease progression in papillary thyroid carcinoma. Clin. Exp. Med. 2017, 17, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Y.; Liu, C.L.; Chen, M.J.; Lee, J.J.; Pun, P.C.; Cheng, S.P. Expression of haem oxygenase-1 correlates with tumour aggressiveness and BRAF V600E expression in thyroid cancer. Histopathology 2015, 66, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Vreman, H.J.; Ekstrand, B.C.; Stevenson, D.K. Selection of metalloporphyrin heme oxygenase inhibitors based on potency and photoreactivity. Pediatr. Res. 1993, 33, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Kongpetch, S.; Puapairoj, A.; Ong, C.K.; Senggunprai, L.; Prawan, A.; Kukongviriyapan, U.; Chan-On, W.; Siew, E.Y.; Khuntikeo, N.; Teh, B.T.; et al. Haem oxygenase 1 expression is associated with prognosis in cholangiocarcinoma patients and with drug sensitivity in xenografted mice. Cell Prolif. 2016, 49, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Kappas, A.; Drummond, G.S. Control of heme metabolism with synthetic metalloporphyrins. J. Clin. Investig. 1986, 77, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.J.; Vreman, H.J.; Schulz, S.; Kalish, F.S.; Pierce, N.W.; Stevenson, D.K. In vitro inhibition of heme oxygenase isoenzymes by metalloporphyrins. J. Perinatol. 2011, 31 (Suppl. 1), S35–S41. [Google Scholar] [CrossRef] [PubMed]
- Kinobe, R.T.; Dercho, R.A.; Vlahakis, J.Z.; Brien, J.F.; Szarek, W.A.; Nakatsu, K. Inhibition of the enzymatic activity of heme oxygenases by azole-based antifungal drugs. J. Pharmacol. Exp. Ther. 2006, 319, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Salerno, L.; Pittala, V.; Romeo, G.; Modica, M.N.; Siracusa, M.A.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Tibullo, D.; Sorrenti, V. Evaluation of novel aryloxyalkyl derivatives of imidazole and 1,2,4-triazole as heme oxygenase-1 (HO-1) inhibitors and their antitumor properties. Bioorg. Med. Chem. 2013, 21, 5145–5153. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Lan, W.M.; Chau, L.Y. TRC8 suppresses tumorigenesis through targeting heme oxygenase-1 for ubiquitination and degradation. Oncogene 2013, 32, 2325–2334. [Google Scholar] [CrossRef] [PubMed]
- Hinds, T.D., Jr.; Sodhi, K.; Meadows, C.; Fedorova, L.; Puri, N.; Kim, D.H.; Peterson, S.J.; Shapiro, J.; Abraham, N.G.; Kappas, A. Increased HO-1 levels ameliorate fatty liver development through a reduction of heme and recruitment of FGF21. Obesity 2014, 22, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Joe, Y.; Zheng, M.; Kim, H.J.; Uddin, M.J.; Kim, S.K.; Chen, Y.; Park, J.; Cho, G.J.; Ryter, S.W.; Chung, H.T. Cilostazol attenuates murine hepatic ischemia and reperfusion injury via heme oxygenase-dependent activation of mitochondrial biogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G21–G29. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Su, Y.; Lui, W.Y.; Chau, G.Y.; Yin, P.H.; Lee, H.C.; Chi, C.W. Peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α) upregulated E-cadherin expression in HepG2 cells. FEBS Lett. 2008, 582, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Su, D.H.; Chang, S.H.; Chang, T.C. The impact of locoregional recurrences and distant metastases on the survival of patients with papillary thyroid carcinoma. Clin. Endocrinol. 2015, 82, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Chien, M.N.; Yang, P.S.; Lee, J.J.; Wang, T.Y.; Hsu, Y.C.; Cheng, S.P. Recurrence-associated genes in papillary thyroid cancer: An analysis of data from The Cancer Genome Atlas. Surgery 2017, 161, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [PubMed]
- Ziros, P.G.; Manolakou, S.D.; Habeos, I.G.; Lilis, I.; Chartoumpekis, D.V.; Koika, V.; Soares, P.; Kyriazopoulou, V.E.; Scopa, C.D.; Papachristou, D.J.; et al. Nrf2 is commonly activated in papillary thyroid carcinoma, and it controls antioxidant transcriptional responses and viability of cancer cells. J. Clin. Endocrinol. Metab. 2013, 98, E1422–E1427. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, H.E.; Martinez, J.R.; Vargas-Salas, S.; Solar, A.; Veliz, L.; Cruz, F.; Arias, T.; Loyola, S.; Horvath, E.; Tala, H.; et al. A 10-gene classifier for indeterminate thyroid nodules: Development and multicenter accuracy study. Thyroid 2017, 27, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.G.; Liu, Z.M.; Vlantis, A.C.; Tse, G.M.; Leung, B.C.; van Hasselt, C.A. Heme oxygenase-1 protects against apoptosis induced by tumor necrosis factor-alpha and cycloheximide in papillary thyroid carcinoma cells. J. Cell. Biochem. 2004, 92, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Nowis, D.; Bugajski, M.; Winiarska, M.; Bil, J.; Szokalska, A.; Salwa, P.; Issat, T.; Was, H.; Jozkowicz, A.; Dulak, J.; et al. Zinc protoporphyrin IX, a heme oxygenase-1 inhibitor, demonstrates potent antitumor effects but is unable to potentiate antitumor effects of chemotherapeutics in mice. BMC Cancer 2008, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Outschoorn, U.E.; Peiris-Pages, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Jais, A.; Einwallner, E.; Sharif, O.; Gossens, K.; Lu, T.T.; Soyal, S.M.; Medgyesi, D.; Neureiter, D.; Paier-Pourani, J.; Dalgaard, K.; et al. Heme oxygenase-1 drives metaflammation and insulin resistance in mouse and man. Cell 2014, 158, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Anastasiou, D.; Poulogiannis, G.; Asara, J.M.; Boxer, M.B.; Jiang, J.K.; Shen, M.; Bellinger, G.; Sasaki, A.T.; Locasale, J.W.; Auld, D.S.; et al. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 2011, 334, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Salvatori, M.; Biondi, B.; Rufini, V. Imaging in endocrinology: 2-[18F]-fluoro-2-deoxy-d-glucose positron emission tomography/computed tomography in differentiated thyroid carcinoma: Clinical indications and controversies in diagnosis and follow-up. Eur. J. Endocrinol. 2015, 173, R115–R130. [Google Scholar] [PubMed]
- Hsu, Y.C.; Liu, C.L.; Yang, P.S.; Tsai, C.H.; Lee, J.J.; Cheng, S.P. Interaction of age at diagnosis with transcriptional profiling in papillary thyroid cancer. World J. Surg. 2016, 40, 2922–2929. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.F.; Yeh, C.T.; Sun, Y.J.; Chiang, M.T.; Lan, W.M.; Li, F.A.; Lee, W.H.; Chau, L.Y. Signal peptide peptidase-mediated nuclear localization of heme oxygenase-1 promotes cancer cell proliferation and invasion independent of its enzymatic activity. Oncogene 2015, 34, 2360–2370. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, R.E.; Klopper, J.P.; Korch, C.; Pugazhenthi, U.; Benezra, M.; Knauf, J.A.; Fagin, J.A.; Marlow, L.A.; Copland, J.A.; Smallridge, R.C.; et al. Deoxyribonucleic acid profiling analysis of 40 human thyroid cancer cell lines reveals cross-contamination resulting in cell line redundancy and misidentification. J. Clin. Endocrinol. Metab. 2008, 93, 4331–4341. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Chen, C.K.; Chen, M.J.; Lin, J.C.; Lin, C.H.; Huang, W.C.; Cheng, S.P.; Chen, S.N.; Liu, C.L. Expression of 3β-hydroxysteroid dehydrogenase type 1 in breast cancer is associated with poor prognosis independent of estrogen receptor status. Ann. Surg. Oncol. 2017, 24, 4033–4041. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Asakura, M.; Min, K.D.; Ito, S.; Fu, H.Y.; Yamazaki, S.; Takahashi, A.; Imazu, M.; Fukuda, H.; Nakajima, Y.; et al. Cartilage intermediate layer protein 1 suppresses TGF-β signaling in cardiac fibroblasts. Int. J. Gerontol. 2017, 11, 67–74. [Google Scholar] [CrossRef]
- Chang, Y.C.; Hsu, Y.C.; Liu, C.L.; Huang, S.Y.; Hu, M.C.; Cheng, S.P. Local anesthetics induce apoptosis in human thyroid cancer cells through the mitogen-activated protein kinase pathway. PLoS ONE 2014, 9, e89563. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Yang, P.S.; Chien, M.N.; Chang, Y.C.; Lin, C.H.; Cheng, S.P. Expression of serine peptidase inhibitor Kunitz type 1 in differentiated thyroid cancer. Histochem. Cell Biol. 2018, 149, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.P.; Liu, C.L.; Chen, M.J.; Chien, M.N.; Leung, C.H.; Lin, C.H.; Hsu, Y.C.; Lee, J.J. CD74 expression and its therapeutic potential in thyroid carcinoma. Endocr. Relat. Cancer 2015, 22, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Wang, T.Y.; Liu, C.L.; Chien, M.N.; Chen, M.J.; Hsu, Y.C.; Leung, C.H.; Cheng, S.P. Dipeptidyl peptidase IV as a prognostic marker and therapeutic target in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 2017, 102, 2930–2940. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.P.; Yang, P.S.; Chien, M.N.; Chen, M.J.; Lee, J.J.; Liu, C.L. Aberrant expression of tumor-associated carbohydrate antigen Globo H in thyroid carcinoma. J. Surg. Oncol. 2016, 114, 853–858. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.-S.; Hsu, Y.-C.; Lee, J.-J.; Chen, M.-J.; Huang, S.-Y.; Cheng, S.-P. Heme Oxygenase-1 Inhibitors Induce Cell Cycle Arrest and Suppress Tumor Growth in Thyroid Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2502. https://doi.org/10.3390/ijms19092502
Yang P-S, Hsu Y-C, Lee J-J, Chen M-J, Huang S-Y, Cheng S-P. Heme Oxygenase-1 Inhibitors Induce Cell Cycle Arrest and Suppress Tumor Growth in Thyroid Cancer Cells. International Journal of Molecular Sciences. 2018; 19(9):2502. https://doi.org/10.3390/ijms19092502
Chicago/Turabian StyleYang, Po-Sheng, Yi-Chiung Hsu, Jie-Jen Lee, Ming-Jen Chen, Shih-Yuan Huang, and Shih-Ping Cheng. 2018. "Heme Oxygenase-1 Inhibitors Induce Cell Cycle Arrest and Suppress Tumor Growth in Thyroid Cancer Cells" International Journal of Molecular Sciences 19, no. 9: 2502. https://doi.org/10.3390/ijms19092502
APA StyleYang, P.-S., Hsu, Y.-C., Lee, J.-J., Chen, M.-J., Huang, S.-Y., & Cheng, S.-P. (2018). Heme Oxygenase-1 Inhibitors Induce Cell Cycle Arrest and Suppress Tumor Growth in Thyroid Cancer Cells. International Journal of Molecular Sciences, 19(9), 2502. https://doi.org/10.3390/ijms19092502