CTLA-4 Mediates Inhibitory Function of Mesenchymal Stem/Stromal Cells

 , ,
, ,
Abstract
:
1. Introduction
2. Results
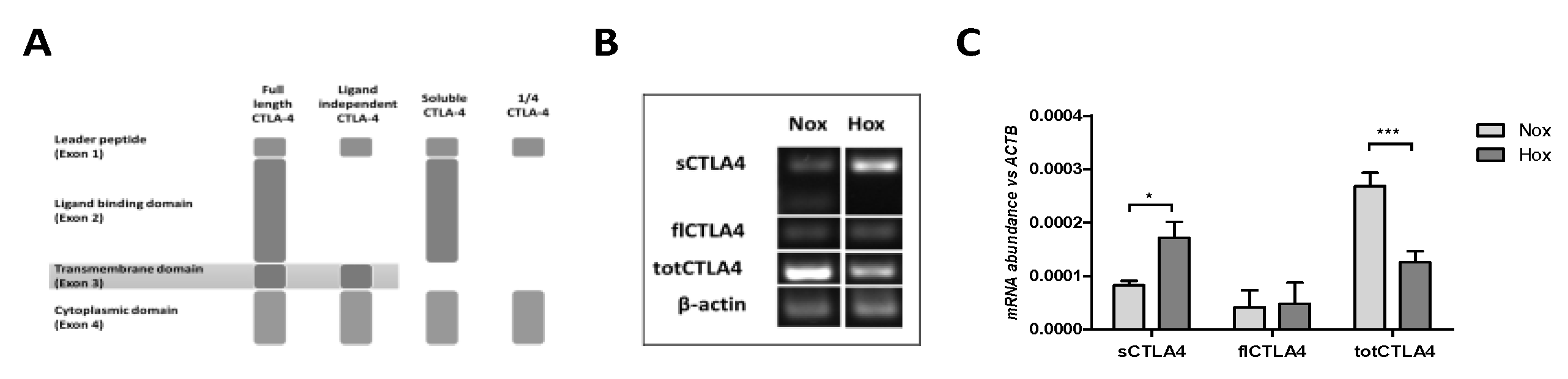
2.1. MSCs Express Different Splice Variants of CTLA-4
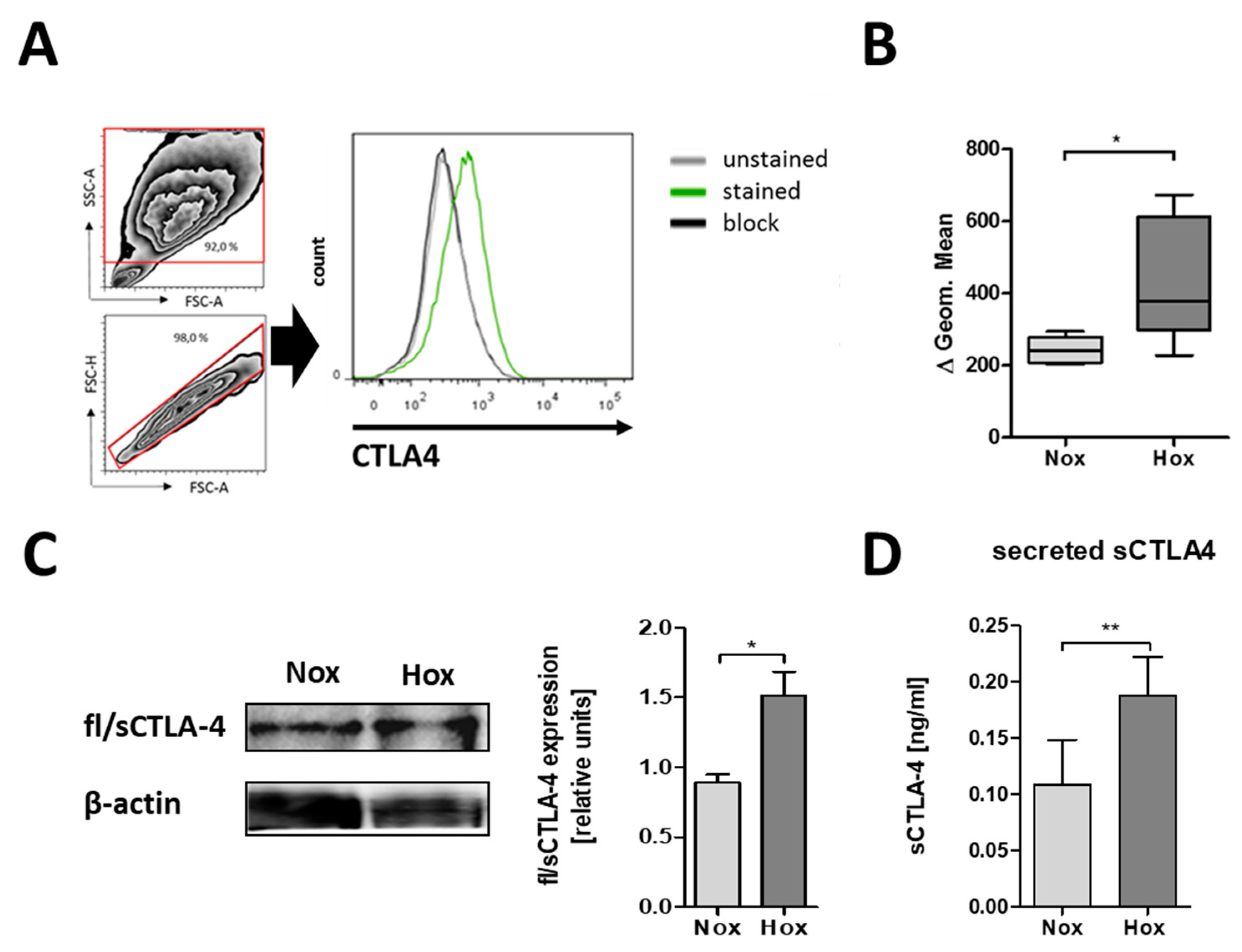
2.2. CTLA-4 is Expressed on the Cell Surface of MSCs
2.3. MSCs are Able to Secrete CTLA-4
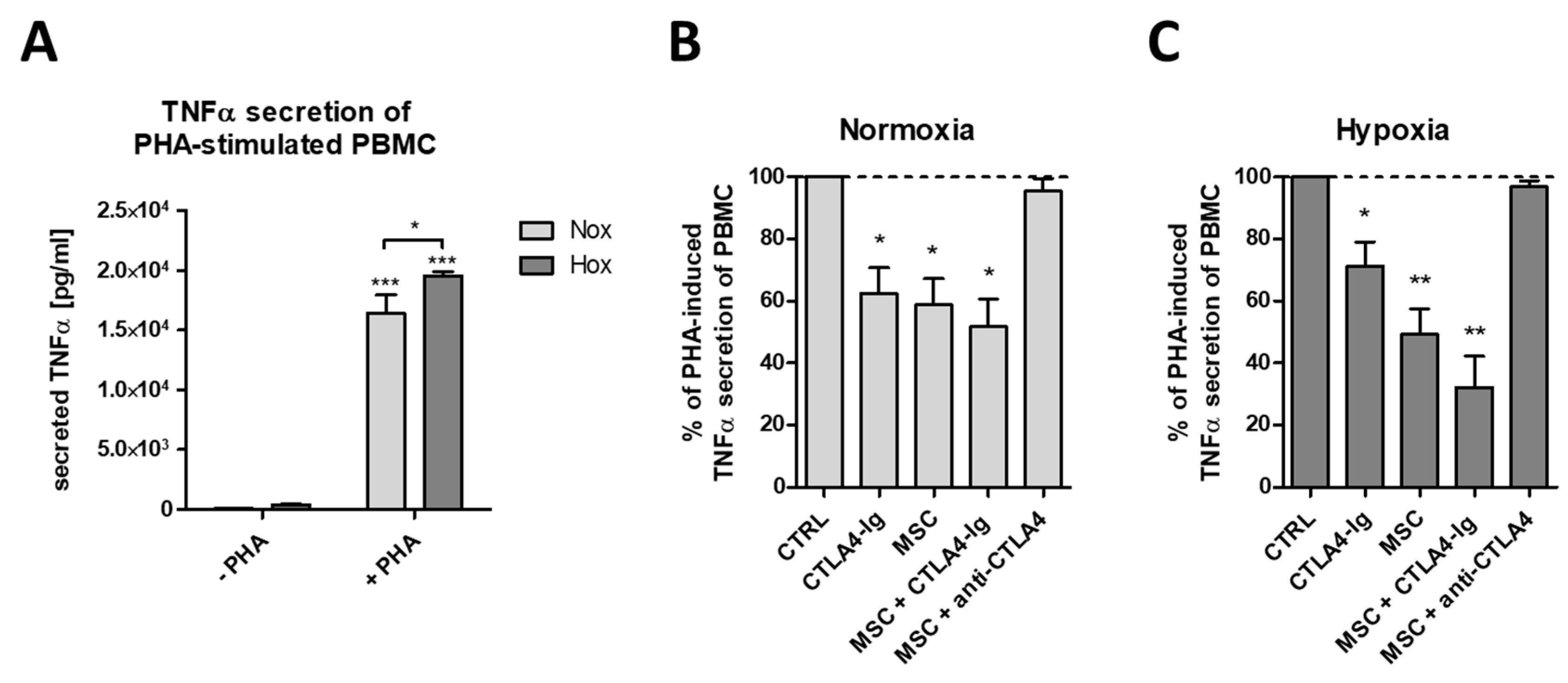
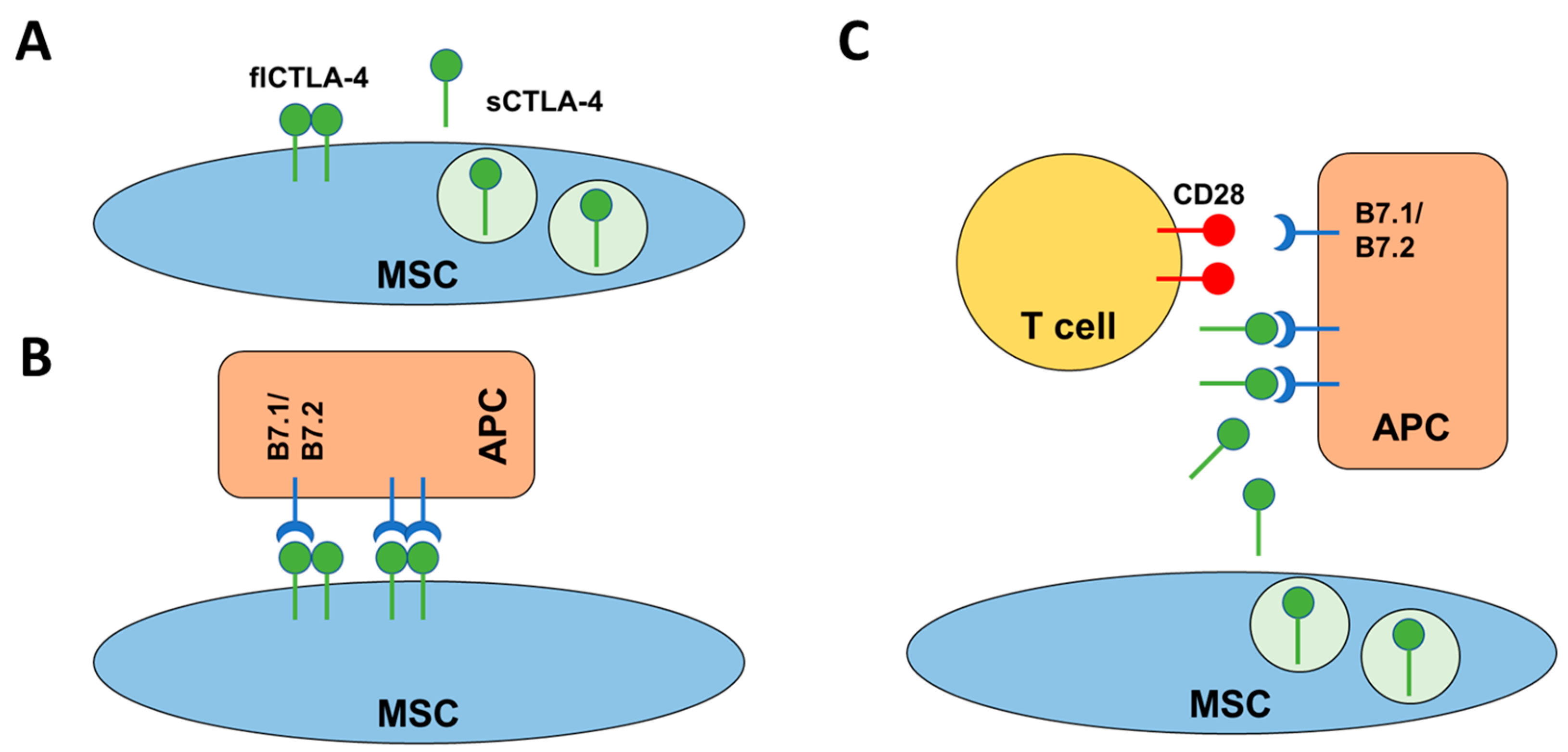
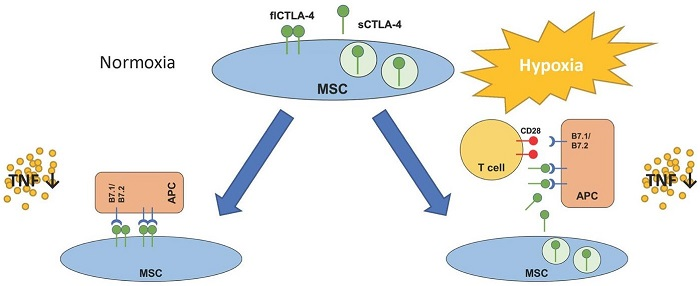
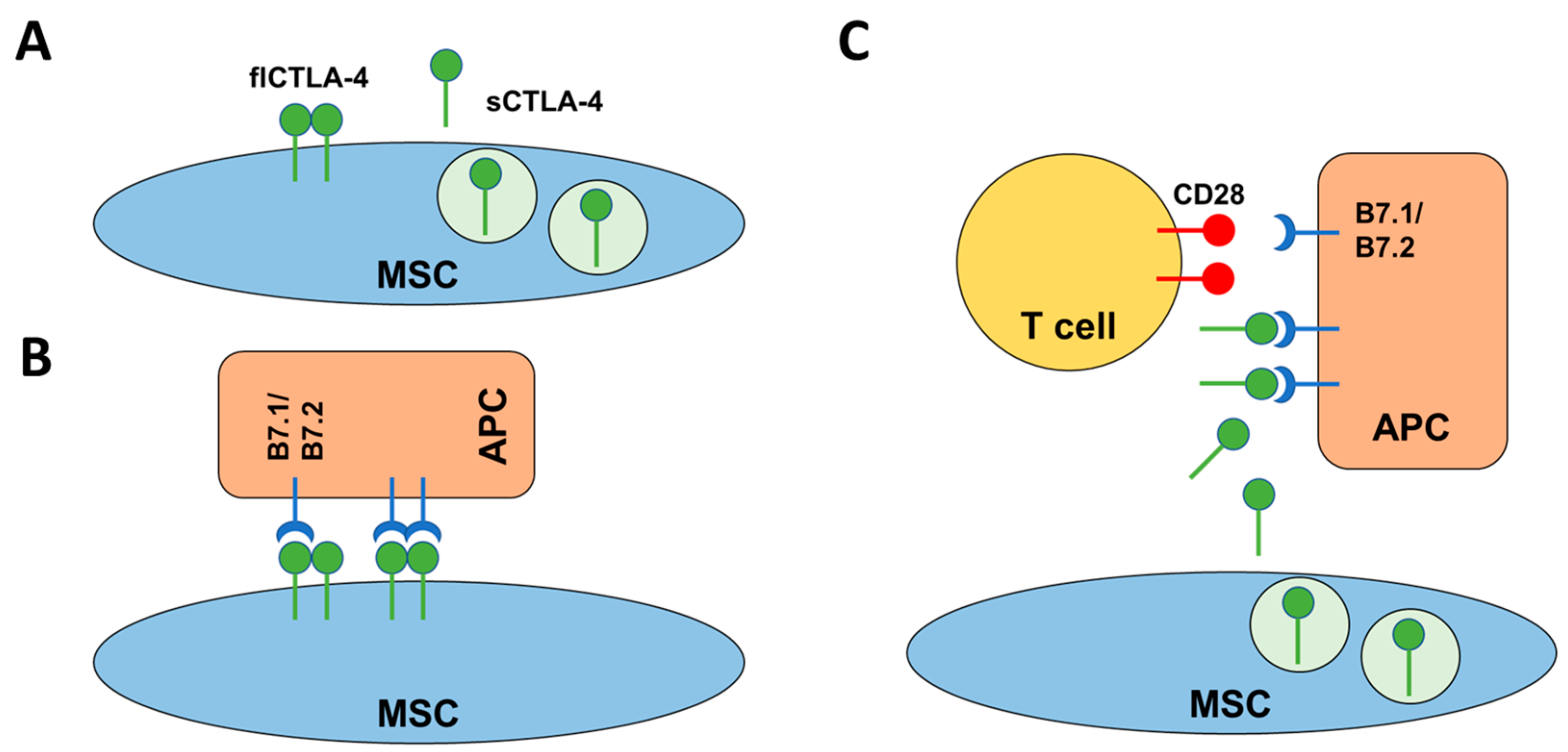
2.4. flCTLA-4 and sCTLA-4 Mediate the Inhibitory Effect of MSCs
3. Discussion
4. Material and Methods
4.1. Antibodies
4.2. Isolation and Expansion and Characterization of MSCs
4.3. Induction of Hypoxia
4.4. Induction of Osteogenic and Adipogenic Differentiation of MSCs
4.5. Oil Red O Staining of Adipogenic Differentiated MSCs
4.6. Alizarin S Red Staining of Osteogenic Differentiated MSCs
4.7. RNA Isolation and Quantitative PCR (qPCR) of CTLA-4 Genes of MSCs
4.8. Immunoblot of CTLA-4 and β-Actin
4.9. Quantification of Secreted CTLA-4
4.10. Quantification of Secreted TNFα
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexander, T.; Arnold, R.; Hiepe, F.; Radbruch, A. Resetting the immune system with immunoablation and autologous haematopoietic stem cell transplantation in autoimmune diseases. Clin. Exp. Rheumatol. 2016, 34, 53–57. [Google Scholar] [PubMed]
- Freitag, J.; Berod, L.; Kamradt, T.; Sparwasser, T. Immunometabolism and autoimmunity. Immunol. Cell Biol. 2016, 94, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Schett, G. Autoimmunity as a trigger for structural bone damage in rheumatoid arthritis. Mod. Rheumatol. 2017, 27, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Mauri, C.; Blair, P.A. Regulatory B cells in autoimmunity: Developments and controversies. Nat. Rev. Rheumatol. 2010, 6, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Miyara, M.; Costantino, C.M.; Hafler, D.A. FOXP3+ regulatory T cells in the human immune system. Nat. Rev. Immunol. 2010, 10, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Wehrens, E.J.; Prakken, B.J.; van Wijk, F. T cells out of control—Impaired immune regulation in the inflamed joint. Nat. Rev. Rheumatol. 2013, 9, 34–42. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, K.F.; Clark, K.; Naqvi, S.; McGuire, V.A.; Noehren, G.; Kristariyanto, Y.; van den Bosch, M.; Mudaliar, M.; McCarthy, P.C.; Pattison, M.J.; et al. PGE(2) induces macrophage IL-10 production and a regulatory-like phenotype via a protein kinase A-SIK-CRTC3 pathway. J. Immunol. 2013, 190, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Allavena, P.; Sica, A.; Solinas, G.; Porta, C.; Mantovani, A. The inflammatory micro-environment in tumor progression: The role of tumor-associated macrophages. Crit. Rev. Oncol. Hematol. 2008, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lippens, C.; Duraes, F.V.; Dubrot, J.; Brighouse, D.; Lacroix, M.; Irla, M.; Aubry-Lachainaye, J.P.; Reith, W.; Mandl, J.N.; Hugues, S. IDO-orchestrated crosstalk between pDCs and Tregs inhibits autoimmunity. J. Autoimmun. 2016, 75, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, D.; Trivedi, R.N.; Kathuria, A.; Khandia, R.; Goswami, T.K.; Munjal, A.K. In vitro and In vivo Immunomodulating Properties of Mesenchymal Stem Cells. Recent Pat. Inflamm. Allergy Drug Discov. 2018. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Lima, F.; da Rocha Romero, A.B.; Hastreiter, A.; Nogueira-Pedro, A.; Makiyama, E.; Colli, C.; Fock, R.A. An insight into the role of magnesium in the immunomodulatory properties of mesenchymal stem cells. J. Nutr. Biochem. 2018, 55, 200–208. [Google Scholar] [CrossRef] [PubMed]
- De Ugarte, D.A.; Morizono, K.; Elbarbary, A.; Alfonso, Z.; Zuk, P.A.; Zhu, M.; Dragoo, J.L.; Ashjian, P.; Thomas, B.; Benhaim, P.; et al. Comparison of multi-lineage cells from human adipose tissue and bone marrow. Cells Tissues Organs 2003, 174, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kadle, R.L.; Abdou, S.A.; Villarreal-Ponce, A.P.; Soares, M.A.; Sultan, D.L.; David, J.A.; Massie, J.; Rifkin, W.J.; Rabbani, P.; Ceradini, D.J. Microenvironmental cues enhance mesenchymal stem cell-mediated immunomodulation and regulatory T-cell expansion. PLoS ONE 2018, 13, e0193178. [Google Scholar] [CrossRef] [PubMed]
- O'Driscoll, S.W.; Saris, D.B.; Ito, Y.; Fitzimmons, J.S. The chondrogenic potential of periosteum decreases with age. J. Orthop. Res. 2001, 19, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Sandrasaigaran, P.; Algraittee, S.J.R.; Ahmad, A.R.; Vidyadaran, S.; Ramasamy, R. Characterisation and immunosuppressive activity of human cartilage-derived mesenchymal stem cells. Cytotechnology 2018, 70, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Wagegg, M.; Gaber, T.; Lohanatha, F.L.; Hahne, M.; Strehl, C.; Fangradt, M.; Tran, C.L.; Schonbeck, K.; Hoff, P.; Ode, A.; et al. Hypoxia promotes osteogenesis but suppresses adipogenesis of human mesenchymal stromal cells in a hypoxia-inducible factor-1 dependent manner. PLoS ONE 2012, 7, e46483. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, A.; Sturgeon, C.; Siatskas, M.; Ferrer, K.; McIntosh, K.; Patil, S.; Hardy, W.; Devine, S.; Ucker, D.; Deans, R.; et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp. Hematol. 2002, 30, 42–48. [Google Scholar] [CrossRef]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stagg, J. Immune regulation by mesenchymal stem cells: Two sides to the coin. Tissue Antigens 2007, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lohan, P.; Treacy, O.; Griffin, M.D.; Ritter, T.; Ryan, A.E. Anti-Donor Immune Responses Elicited by Allogeneic Mesenchymal Stem Cells and Their Extracellular Vesicles: Are We Still Learning? Front. Immunol. 2017, 8, 1626. [Google Scholar] [CrossRef] [PubMed]
- Faiella, W.; Atoui, R. Immunotolerant Properties of Mesenchymal Stem Cells: Updated Review. Stem Cells Int. 2016, 2016, 1859567. [Google Scholar] [CrossRef] [PubMed]
- Burr, S.P.; Dazzi, F.; Garden, O.A. Mesenchymal stromal cells and regulatory T cells: The Yin and Yang of peripheral tolerance? Immunol. Cell Biol. 2013, 91, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Gaber, T.; Schmidt-Bleek, K.; Senturk, U.; Tran, C.L.; Blankenstein, K.; Lutkecosmann, S.; Bredahl, J.; Schuler, H.J.; Simon, P.; et al. Immunologically restricted patients exhibit a pronounced inflammation and inadequate response to hypoxia in fracture hematomas. Immunol. Res. 2011, 51, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Gaber, T.; Strehl, C.; Schmidt-Bleek, K.; Lang, A.; Huscher, D.; Burmester, G.R.; Schmidmaier, G.; Perka, C.; Duda, G.N.; et al. Immunological characterization of the early human fracture hematoma. Immunol. Res. 2016, 64, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Maschmeyer, P.; Gaber, T.; Schutze, T.; Raue, T.; Schmidt-Bleek, K.; Dziurla, R.; Schellmann, S.; Lohanatha, F.L.; Rohner, E.; et al. Human immune cells’ behavior and survival under bioenergetically restricted conditions in an in vitro fracture hematoma model. Cell Mol. Immunol. 2013, 10, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Rakow, A.; Gaber, T.; Hahne, M.; Senturk, U.; Strehl, C.; Fangradt, M.; Schmidt-Bleek, K.; Huscher, D.; Winkler, T.; et al. Preoperative irradiation for the prevention of heterotopic ossification induces local inflammation in humans. Bone 2013, 55, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Kolar, P.; Gaber, T.; Perka, C.; Duda, G.N.; Buttgereit, F. Human early fracture hematoma is characterized by inflammation and hypoxia. Clin. Orthop. Relat. Res. 2011, 469, 3118–3126. [Google Scholar] [CrossRef] [PubMed]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The Early Fracture Hematoma and Its Potential Role in Fracture Healing. Tissue Eng. Part. B Rev. 2010, 16, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Hoff, P.; Gaber, T.; Strehl, C.; Jakstadt, M.; Hoff, H.; Schmidt-Bleek, K.; Lang, A.; Rohner, E.; Huscher, D.; Matziolis, G.; et al. A Pronounced Inflammatory Activity Characterizes the Early Fracture Healing Phase in Immunologically Restricted Patients. Int. J. Mol. Sci. 2017, 18, 583. [Google Scholar] [CrossRef] [PubMed]
- Barry, F.; Murphy, M. Mesenchymal stem cells in joint disease and repair. Nat. Rev. Rheumatol. 2013, 9, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.; Zhang, Z.; Chen, L.; Ji, H.L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal stem cells and immunomodulation: Current status and future prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, J.F.; Denizot, F.; Luciani, M.F.; Roux-Dosseto, M.; Suzan, M.; Mattei, M.G.; Golstein, P. A new member of the immunoglobulin superfamily–CTLA-4. Nature 1987, 328, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Brunner-Weinzierl, M.C.; Hoff, H.; Burmester, G.R. Multiple functions for CD28 and cytotoxic T lymphocyte antigen-4 during different phases of T cell responses: Implications for arthritis and autoimmune diseases. Arthritis Res. Ther. 2004, 6, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpert, M.M.; Konduri, V.; Liang, D.; Chen, Y.; Wing, J.B.; Paust, S.; Levitt, J.M.; Decker, W.K. Dendritic Cell-Secreted Cytotoxic T-Lymphocyte-Associated Protein-4 Regulates the T-cell Response by Downmodulating Bystander Surface B7. Stem Cells Dev. 2016, 25, 774–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quandt, D.; Hoff, H.; Rudolph, M.; Fillatreau, S.; Brunner-Weinzierl, M.C. A new role of CTLA-4 on B cells in thymus-dependent immune responses in vivo. J. Immunol. 2007, 179, 7316–7324. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, U.; Orabona, C.; Fallarino, F.; Vacca, C.; Calcinaro, F.; Falorni, A.; Candeloro, P.; Belladonna, M.L.; Bianchi, R.; Fioretti, M.C.; et al. CTLA-4-Ig regulates tryptophan catabolism in vivo. Nat. Immunol. 2002, 3, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Hoff, H.; Knieke, K.; Cabail, Z.; Hirseland, H.; Vratsanos, G.; Burmester, G.R.; Jorch, G.; Nadler, S.G.; Broker, B.; Hebel, K.; et al. Surface CD152 (CTLA-4) expression and signaling dictates longevity of CD28null T cells. J. Immunol. 2009, 182, 5342–5351. [Google Scholar] [CrossRef] [PubMed]
- Hoff, H.; Kolar, P.; Ambach, A.; Radbruch, A.; Brunner-Weinzierl, M.C. CTLA-4 (CD152) inhibits T cell function by activating the ubiquitin ligase Itch. Mol. Immunol. 2010, 47, 1875–1881. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Howson, J.M.; Esposito, L.; Heward, J.; Snook, H.; Chamberlain, G.; Rainbow, D.B.; Hunter, K.M.; Smith, A.N.; Di Genova, G.; et al. Association of the T-cell regulatory gene CTLA4 with susceptibility to autoimmune disease. Nature 2003, 423, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Chen, Y.; Wei, L.; Li, S.; Jin, X.; Cong, C.; Yuan, Y.; Long, D.; Li, Y.; Cheng, J.; et al. Inhibitory effect of mesenchymal stem cells on lymphocyte proliferation. Cell Biochem. Funct. 2008, 26, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, I.; Ringden, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit lymphocyte proliferation by mitogens and alloantigens by different mechanisms. Exp. Cell Res. 2005, 305, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Chinnadurai, R.; Copland, I.B.; Patel, S.R.; Galipeau, J. IDO-independent suppression of T cell effector function by IFN-gamma-licensed human mesenchymal stromal cells. J. Immunol. 2014, 192, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Mancheno-Corvo, P.; Menta, R.; del Rio, B.; Franquesa, M.; Ramirez, C.; Hoogduijn, M.J.; DelaRosa, O.; Dalemans, W.; Lombardo, E. T Lymphocyte Prestimulation Impairs in a Time-Dependent Manner the Capacity of Adipose Mesenchymal Stem Cells to Inhibit Proliferation: Role of Interferon gamma, Poly I:C, and Tryptophan Metabolism in Restoring Adipose Mesenchymal Stem Cell Inhibitory Effect. Stem Cells Dev. 2015, 24, 2158–2170. [Google Scholar] [PubMed]
- Jones, S.; Horwood, N.; Cope, A.; Dazzi, F. The antiproliferative effect of mesenchymal stem cells is a fundamental property shared by all stromal cells. J. Immunol. 2007, 179, 2824–2831. [Google Scholar] [CrossRef] [PubMed]
- Zomorodian, E.; Baghaban Eslaminejad, M. Mesenchymal stem cells as a potent cell source for bone regeneration. Stem Cells Int. 2012, 2012, 980353. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Wu, K.C.; Liu, H.W.; Chu, T.Y.; Ding, D.C. Human umbilical cord-derived mesenchymal stem cells reduce monosodium iodoacetate-induced apoptosis in cartilage. Ci Ji Yi Xue Za Zhi 2018, 30, 71–80. [Google Scholar] [PubMed]
- Gugjoo, M.B.; Amarpal, G.T.; Aithal, H.P.; Kinjavdekar, P. Cartilage tissue engineering: Role of mesenchymal stem cells along with growth factors & scaffolds. Indian J. Med. Res. 2016, 144, 339–347. [Google Scholar] [PubMed]
- Hermeto, L.C.; DeRossi, R.; Oliveira, R.J.; Pesarini, J.R.; Antoniolli-Silva, A.C.; Jardim, P.H.; Santana, A.E.; Deffune, E.; Rinaldi, J.C.; Justulin, L.A. Effects of intra-articular injection of mesenchymal stem cells associated with platelet-rich plasma in a rabbit model of osteoarthritis. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Granero-Molto, F.; Weis, J.A.; Miga, M.I.; Landis, B.; Myers, T.J.; O’Rear, L.; Longobardi, L.; Jansen, E.D.; Mortlock, D.P.; Spagnoli, A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells 2009, 27, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
- Lamo-Espinosa, J.M.; Mora, G.; Blanco, J.F.; Granero-Molto, F.; Nunez-Cordoba, J.M.; Sanchez-Echenique, C.; Bondia, J.M.; Aquerreta, J.D.; Andreu, E.J.; Ornilla, E.; et al. Intra-articular injection of two different doses of autologous bone marrow mesenchymal stem cells versus hyaluronic acid in the treatment of knee osteoarthritis: Multicenter randomized controlled clinical trial (phase I/II). J. Transl. Med. 2016, 14, 246. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | Accession Number | Forward Primer | Reverse Primer | Remarks |
|---|---|---|---|---|---|
| flCTLA4 | Homo sapiens cytotoxic T-lymphocyte associated protein 4 (CTLA4), transcript variant 1, mRNA | NM_005214 | ACC CAG ATT TAT GTA ATT GAT CCA GAA | CCG AAC TAA CTG CAA GGA | Primer combination detects transcript variant 1, which represents the longer transcript and encodes the longer membrane-bound isoform CTLA4-TM (also known as full-length (fl)CTLA4) |
| sCTLA4 | Homo sapiens cytotoxic T-lymphocyte associated protein 4 (CTLA4), transcript variant 2, mRNA | NM_001037631 | ATG TAA TTG CTA AAG AAA AGA AGC CCT C | GCCTCAGCTCTTGGAAATTGAAAT | Primer combination detects the encoded isoform CTLA-4delTM (also known as sCTLA4) which is soluble and lacks the transmembrane domain, compared to isoform |
| totCTLA4 | Homo sapiens cytotoxic T-lymphocyte associated protein 4 (CTLA4), transcript variant 1 and 2, mRNA | NM_005214 and NM_001037631 | TGT TGA CAT GTG CTT TGG GG | GCT GCC TTC TGT CCA TG | Primer combination detects all transcript variants of CTLA4 including flCTLA-4 and sCTLA-4 but also the ligand independent CTLA-4 and the 1/4 CTLA-4 that lacks both the ligand-binding domain and the transmembrane domain |
| ACTB | Homo sapiens actin beta (ACTB), mRNA | NM_001101 | GAC AGG ATG CAG AAG GAG ATC ACT | TGA TCC ACA TCT GCT GGA AGG T | Primer combination detects full length ACTB |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaber, T.; Schönbeck, K.; Hoff, H.; Tran, C.L.; Strehl, C.; Lang, A.; Ohrndorf, S.; Pfeiffenberger, M.; Röhner, E.; Matziolis, G.; et al. CTLA-4 Mediates Inhibitory Function of Mesenchymal Stem/Stromal Cells. Int. J. Mol. Sci. 2018, 19, 2312. https://doi.org/10.3390/ijms19082312
Gaber T, Schönbeck K, Hoff H, Tran CL, Strehl C, Lang A, Ohrndorf S, Pfeiffenberger M, Röhner E, Matziolis G, et al. CTLA-4 Mediates Inhibitory Function of Mesenchymal Stem/Stromal Cells. International Journal of Molecular Sciences. 2018; 19(8):2312. https://doi.org/10.3390/ijms19082312
Chicago/Turabian StyleGaber, Timo, Kerstin Schönbeck, Holger Hoff, Cam Loan Tran, Cindy Strehl, Annemarie Lang, Sarah Ohrndorf, Moritz Pfeiffenberger, Eric Röhner, Georg Matziolis, and et al. 2018. "CTLA-4 Mediates Inhibitory Function of Mesenchymal Stem/Stromal Cells" International Journal of Molecular Sciences 19, no. 8: 2312. https://doi.org/10.3390/ijms19082312