Pan-Cancer Analysis Reveals Differential Susceptibility of Bidirectional Gene Promoters to DNA Methylation, Somatic Mutations, and Copy Number Alterations

Abstract
1. Introduction
2. Results
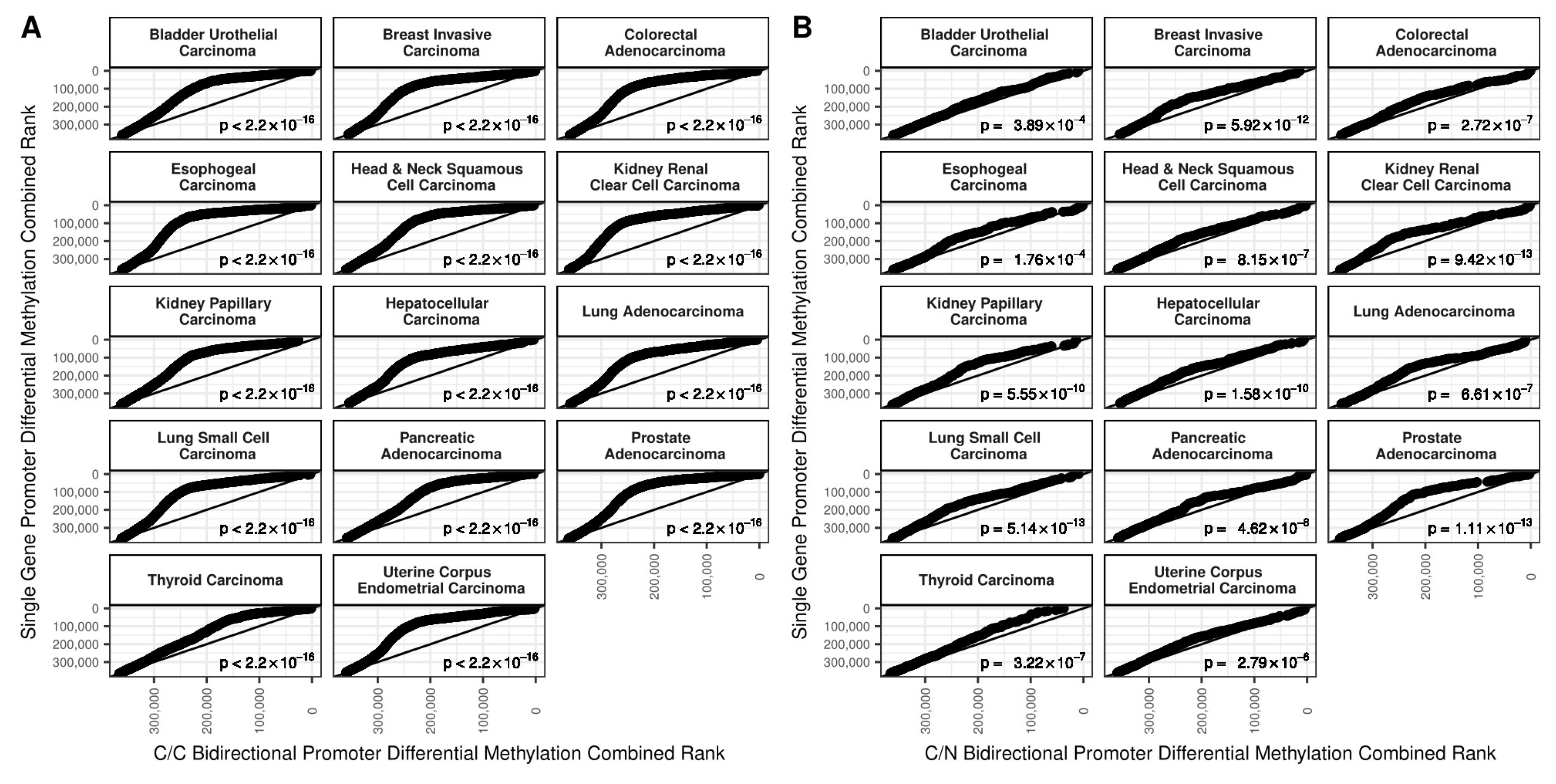
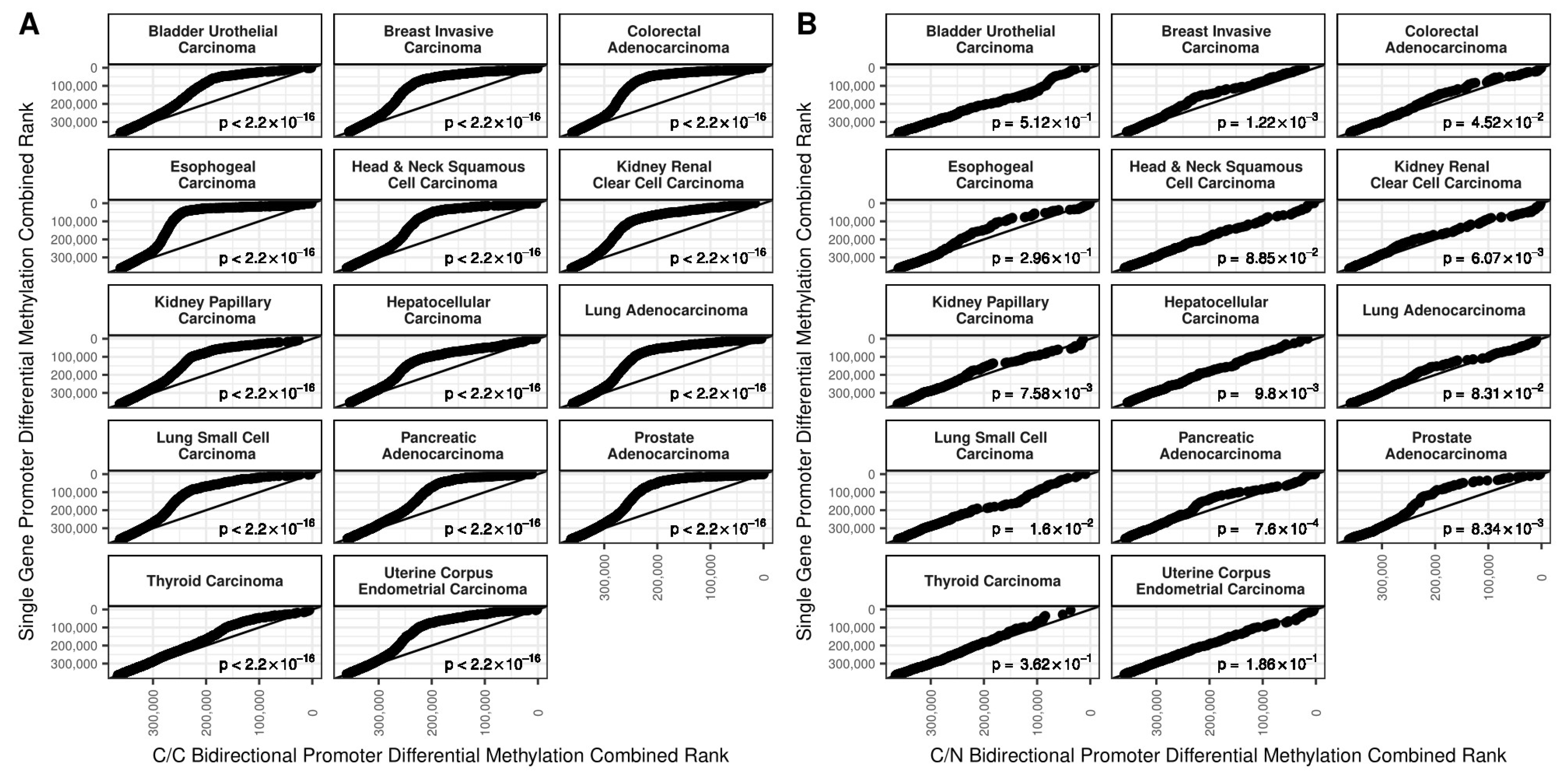
2.1. DNA Methylation
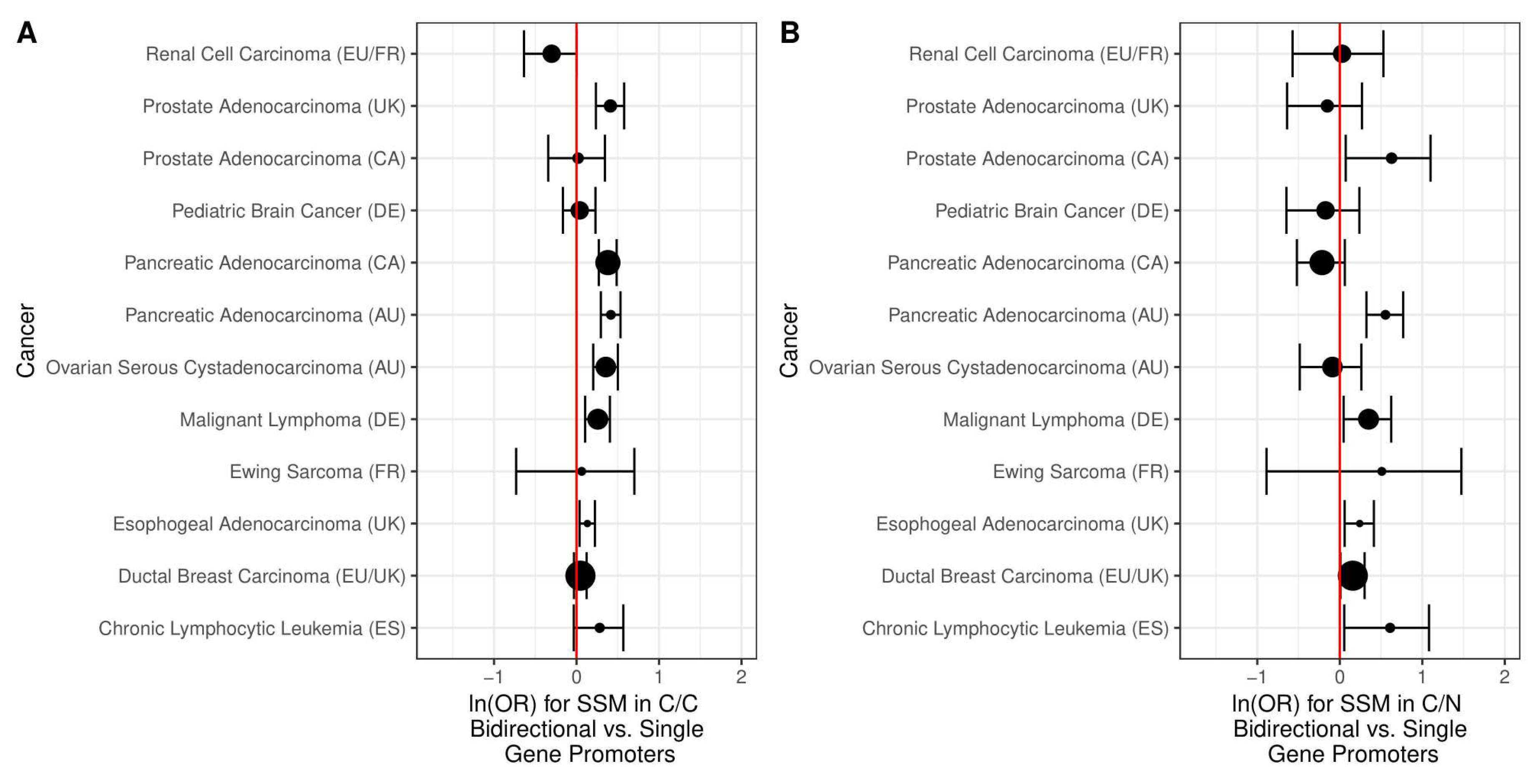
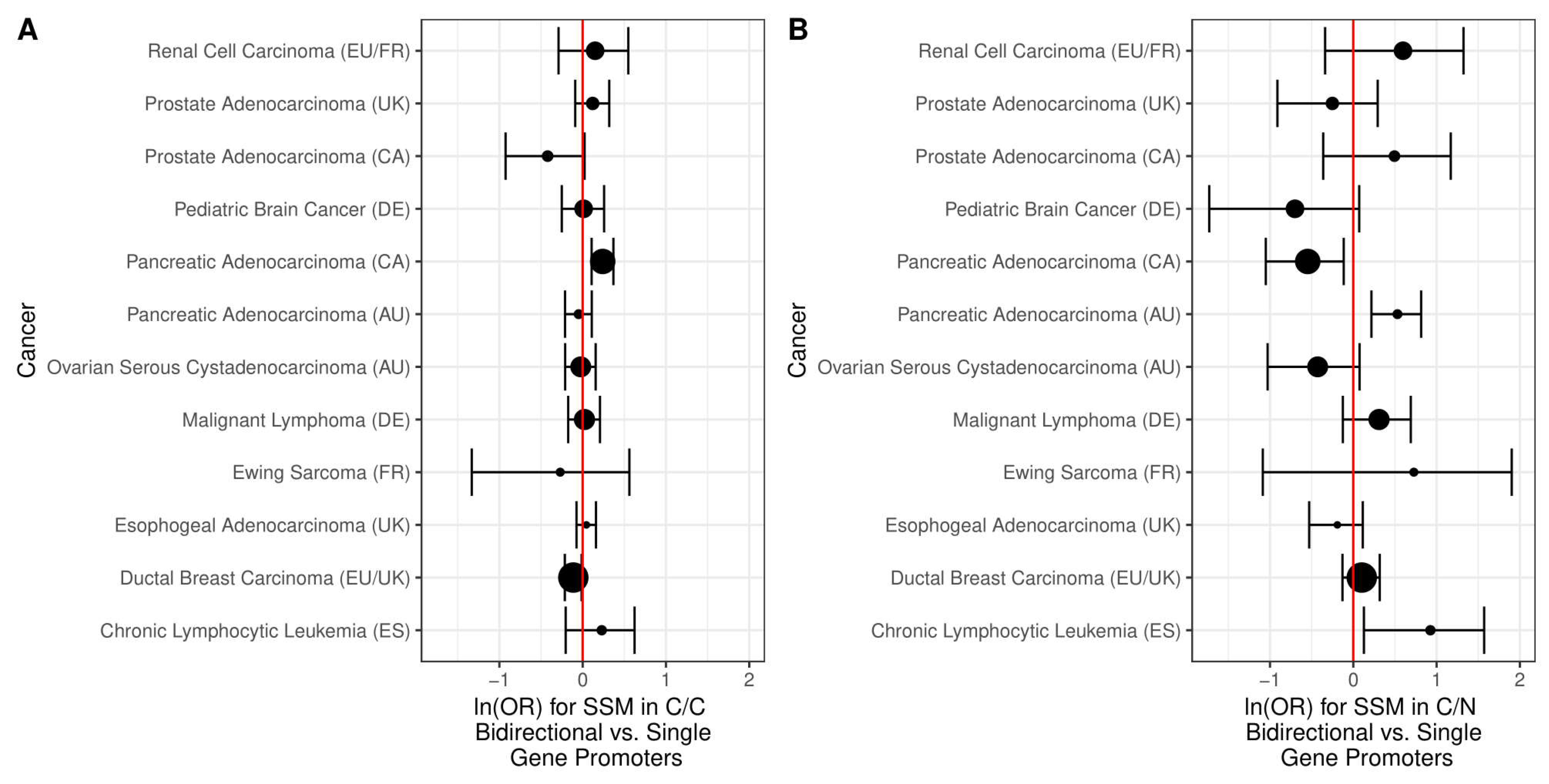
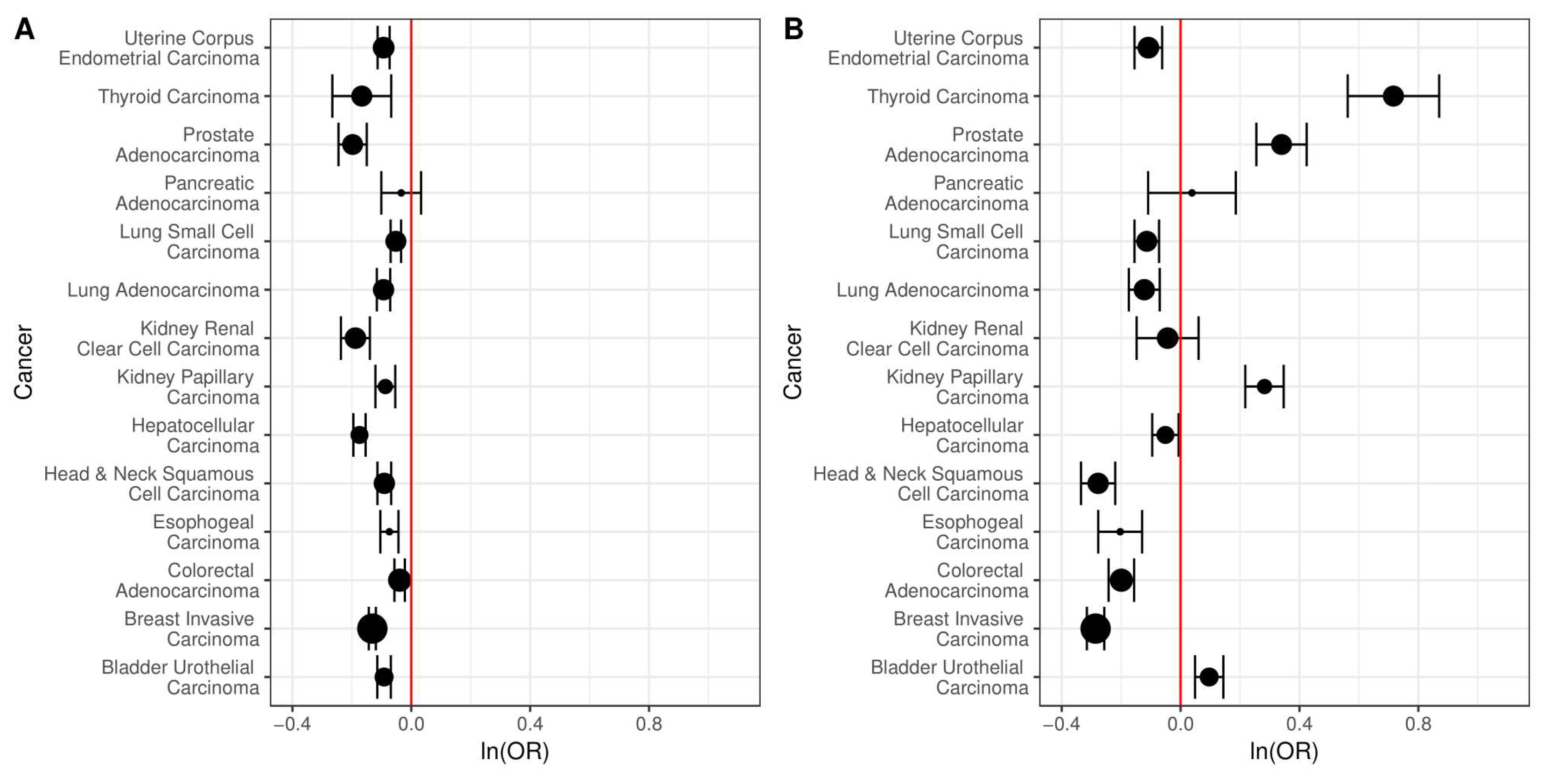
2.2. Simple Somatic Mutations
2.3. Somatic Copy Number Alterations
2.4. Cancer Genes
2.5. DNAse Hypersensitive Sites
2.6. Functional Enrichment
3. Discussion
4. Materials and Methods
4.1. Promoter Identification and Definitions
4.2. DNA Methylation Data
4.3. Simple Somatic Mutation Data
4.4. Somatic Copy Number Alteration Data
Author Contributions
Funding
Conflicts of Interest
References
- Xu, C.; Chen, J.; Shen, B. The Preservation of Bidirectional Promoter Architecture in Eukaryotes—Functional or Co-Regulation Constraint? In Proceedings of the IEE International Conference on Systems Biology (ISB), Zhuhai, China, 2–4 September 2011; pp. 211–218. [Google Scholar]
- Trinklein, N.D.; Aldred, S.F.; Hartman, S.J.; Schroeder, D.I.; Otillar, R.P.; Myers, R.M. An abundance of bidirectional promoters in the human genome. Genome Res. 2004, 14, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Pelechano, V.; Jarvelin, A.I.; Steinmetz, L.M. Functional consequences of bidirectional promoters. Trends Genet. 2011, 27, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Adachi, N.; Lieber, M.R. Bidirectional gene organization: A common architectural feature of the human genome. Cell 2002, 109, 807–809. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Li, Y.X.; Wei, J.; Li, Y.Y. Transcriptional regulation and spatial interactions of head-to-head genes. BMC Genom. 2014, 15, 519. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Q.; Koehly, L.M.; Elnitski, L. Comprehensive annotation of bidirectional promoters identifies co-regulation among breast and ovarian cancer genes. PLoS Comput. Biol. 2007, 3, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.M.; Jelinek, J.; Chang, H.; Shen, L.; Qin, T.; Chung, W.; Oki, Y.; Issa, J.P.J. Silencing of bidirectional promoters by DNA methylation in tumorigenesis. Cancer Res. 2006, 66, 5077–5084. [Google Scholar] [CrossRef] [PubMed]
- Wakano, C.; Byun, J.S.; Di, L.J.; Gardner, K. The dual lives of bidirectional promoters. BBA Gene Regul. Mech. 2012, 1819, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Illingworth, R.S.; Bird, A.P. CpG islands—‘A rough guide’. FEBS Lett. 2009, 583, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
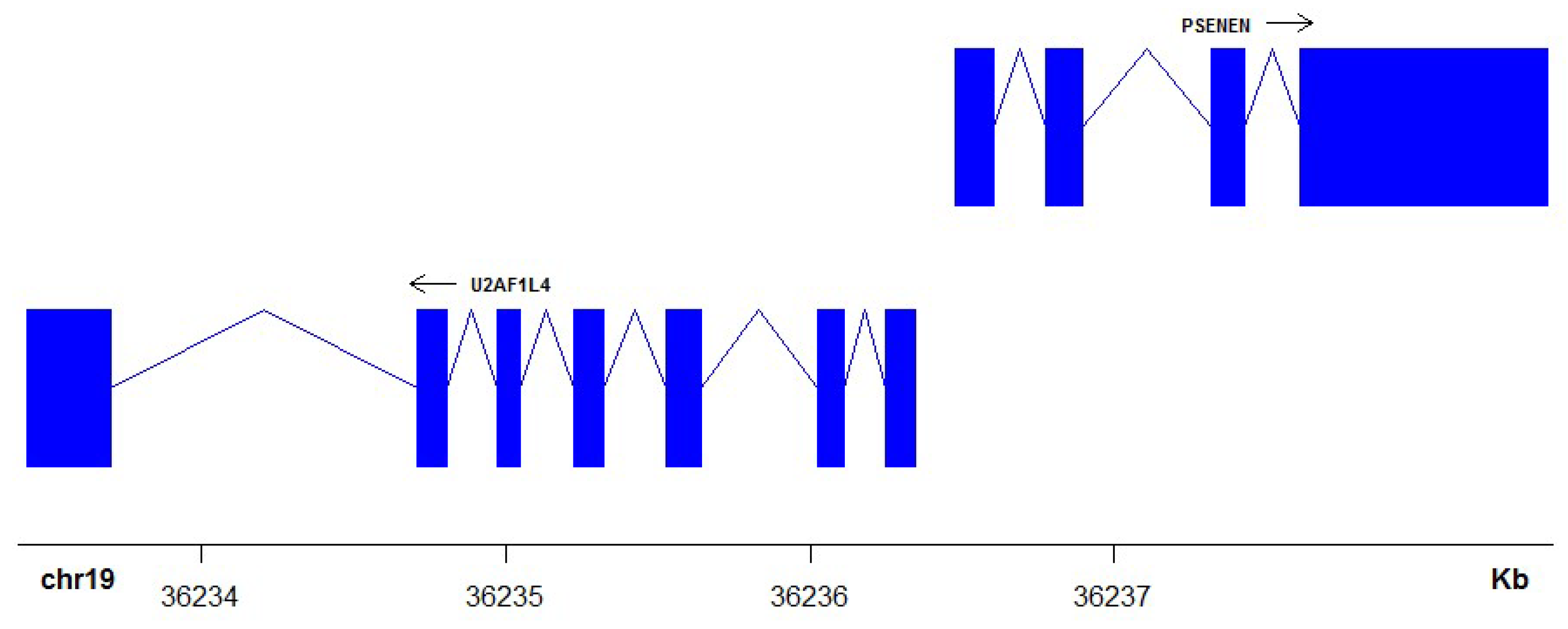
- Didych, D.A.; Shamsutdinov, M.F.; Smirnov, N.A.; Akopov, S.B.; Monastyrskaya, G.S.; Uspenskaya, N.Y.; Nikolaev, L.G.; Sverdlov, E.D. Human PSENEN and U2AF1L4 are concertedly regulated by a genuine bidirectional promoter. Gene 2013, 515, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wu, H.; Xu, H.; Xiong, H.; Chu, Q.; Yu, S.; Wu, G.S.; Wu, K. Notch signaling: An emerging therapeutic target for cancer treatment. Cancer Lett. 2015, 369, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Bellizzi, D.; Dato, S.; Cavalcante, P.; Covello, G.; Di Cianni, F.; Passarino, G.; Rose, G.; De Benedictis, G. Characterization of a bidirectional promoter shared between two human genes related to aging: SIRT3 and PSMD13. Genomics 2007, 89, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Iwahara, T.; Bonasio, R.; Narendra, V.; Reinberg, D. SIRT3 functions in the nucleus in the control of stress-related gene expression. Mol. Cell. Biol. 2012, 32, 5022–5034. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Q.; Elnitski, L. Orthology-driven mapping of bidirectional promoters in human and mouse genomes. BMC Bioinform. 2014, 15, S1. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Chen, J.; Shen, B. The preservation of bidirectional promoter architecture in eukaryotes: What is the driving force? BMC Syst. Biol. 2012, 6, S21. [Google Scholar] [CrossRef] [PubMed]
- Polak, P.; Lawrence, M.S.; Haugen, E.; Stoletzki, N.; Stojanov, P.; Thurman, R.E.; Garraway, L.A.; Mirkin, S.; Getz, G.; Stamatoyannopoulos, J.A.; et al. Reduced local mutation density in regulatory DNA of cancer genomes is linked to DNA repair. Nat. Biotechnol. 2014, 32, 71. [Google Scholar] [CrossRef] [PubMed]
- Crawford, G.E.; Holt, I.E.; Whittle, J.; Webb, B.D.; Tai, D.; Davis, S.; Margulies, E.H.; Chen, Y.D.; Bernat, J.A.; Ginsburg, D.; et al. Genome-wide mapping of DNase hypersensitive sites using massively parallel signature sequencing (MPSS). Genome Res. 2006, 16, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Dillon, L.W.; Burrow, A.A.; Wang, Y.H. DNA Instability at Chromosomal Fragile Sites in Cancer. Curr. Genomics 2010, 11, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Fungtammasan, A.; Walsh, E.; Chiaromonte, F.; Eckert, K.A.; Makova, K.D. A genome-wide analysis of common fragile sites: What features determine chromosomal instability in the human genome? Genome Res. 2012, 22, 993–1005. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.A.; Beare, D.; Gunasekaran, P.; Leung, K.; Bindal, N.; Boutselakis, H.; Ding, M.J.; Bamford, S.; Cole, C.; Ward, S.; et al. COSMIC: Exploring the world’s knowledge of somatic mutations in human cancer. Nucleic Acids Res. 2015, 43, D805–D811. [Google Scholar] [CrossRef] [PubMed]
- Kundaje, A.; Meuleman, W.; Ernst, J.; Bilenky, M.; Yen, A.; Heravi-Moussavi, A.; Kheradpour, P.; Zhang, Z.; Wang, J.; Ziller, M.J.; et al. Integrative analysis of 111 reference human epigenomes. Nature 2015, 518, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Polak, P.; Karlic, R.; Koren, A.; Thurman, R.; Sandstrom, R.; Lawrence, M.S.; Reynolds, A.; Rynes, E.; Vlahovicek, K.; Stamatoyannopoulos, J.A.; et al. Cell-of-origin chromatin organization shapes the mutational landscape of cancer. Nature 2015, 518, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Kirov, S.; Snoddy, J. WebGestalt: An integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res. 2005, 33, W741–W748. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, R.J.; Kahari, A.; Haider, S.; Zamora, J.; Proctor, G.; Spudich, G.; Almeida-King, J.; Staines, D.; Derwent, P.; Kerhornou, A.; et al. Ensembl BioMarts: A hub for data retrieval across taxonomic space. Database-Oxford 2011, 2011, bar030. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M.; Cancer Genome Atlas Research Network. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Grossman, R.L.; Heath, A.P.; Ferretti, V.; Varmus, H.E.; Lowy, D.R.; Kibbe, W.A.; Staudt, L.M. Toward a Shared Vision for Cancer Genomic Data. N. Engl. J. Med. 2016, 375, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.P.; Labbe, A.; Lemire, M.; Zanke, B.W.; Hudson, T.J.; Fertig, E.J.; Greenwood, C.M.T.; Hansen, K.D. Functional normalization of 450k methylation array data improves replication in large cancer studies. Genome Biol. 2014, 15, 503. [Google Scholar] [CrossRef] [PubMed]
- Assenov, Y.; Muller, F.; Lutsik, P.; Walter, J.; Lengauer, T.; Bock, C. Comprehensive analysis of DNA methylation data with RnBeads. Nat. Methods 2014, 11, 1138–1140. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zhang, X.; Huang, C.C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Baran, J.; Cros, A.; Guberman, J.M.; Haider, S.; Hsu, J.; Liang, Y.; Rivkin, E.; Wang, J.X.; Whitty, B.; et al. International Cancer Genome Consortium Data Portal-a one-stop shop for cancer genomics data. Database-Oxford 2011, 2011, bar026. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Gentleman, R.; Carey, V. rtracklayer: An R package for interfacing with genome browsers. Bioinformatics 2009, 25, 1841–1842. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | Methylation Matched Tumor and Normal Samples | Promoter Probes | Copy Number Samples |
|---|---|---|---|
| Bladder Urothelial Carcinoma | 21 | 37,532 | 412 |
| Breast Invasive Carcinoma | 96 | 37,124 | 1094 |
| Colorectal Adenocarcinoma | 37 | 37,088 | 614 |
| Esophogeal Carcinoma | 16 | 37,324 | 184 |
| Head & Neck Squamous Cell Carcinoma | 50 | 37,303 | 517 |
| Kidney Renal Clear Cell Carcinoma | 160 | 37,325 | 530 |
| Kidney Papillary Carcinoma | 45 | 37,227 | 290 |
| Hepatocellular Carcinoma | 49 | 36,845 | 375 |
| Lung Adenocarcinoma | 32 | 37,082 | 518 |
| Lung Small Cell Carcinoma | 42 | 37,581 | 503 |
| Pancreatic Adenocarcinoma | 10 | 37,365 | 184 |
| Prostate Adenocarcinoma | 50 | 37,416 | 497 |
| Thyroid Carcinoma | 56 | 37,779 | 505 |
| Uterine Corpus Endometrial Carcinoma | 46 | 37,296 | 540 |
| Cancer | ICGC Project Code | Samples | Countries |
|---|---|---|---|
| Chronic Lymphocytic Leukemia (ES) | CLLE-ES | 201 | Spain |
| Ductal Breast Carcinoma (EU/UK) | BRCA-EU | 560 | European Union, United Kingdom |
| Esophageal Adenocarcinoma (UK) | ESAD-UK | 203 | United Kingdom |
| Ewing Sarcoma (FR) | BOCA-FR | 98 | France |
| Malignant Lymphoma (DE) | MALY-DE | 100 | Germany |
| Ovarian Serous Cystadenocarcinoma (AU) | OV-AU | 93 | Australia |
| Pancreatic Adenocarcinoma (AU) | PACA-AU | 252 | Australia |
| Pancreatic Adenocarcinoma (CA) | PACA-CA | 259 | Canada |
| Pediatric Brain Cancer (DE) | PBCA-DE | 380 | Germany |
| Prostate Adenocarcinoma (CA) | PRAD-CA | 124 | Canada |
| Prostate Adenocarcinoma (UK) | PRAD-UK | 108 | United Kingdom |
| Renal Cell Carcinoma (EU/FR) | RECA-EU | 95 | European Union, France |
| Tissue Type | Promoter Type | Odds Ratio | 95% CI | p-Value |
|---|---|---|---|---|
| Breast | C/C Bidirectional | 23.73 | [18.72, 30.43] | <2.20 × 10−16 |
| C/N Bidirectional | 6.21 | [4.27, 9.15] | <2.20 × 10−16 | |
| Pancreas | C/C Bidirectional | 28.36 | [21.53, 38.02] | <2.20 × 10−16 |
| C/N Bidirectional | 6.53 | [4.41, 9.86] | <2.20 × 10−16 | |
| Placenta | C/C Bidirectional | 23.05 | [18.30, 29.30] | <2.20 × 10−16 |
| C/N Bidirectional | 7.02 | [4.80, 10.41] | <2.20 × 10−16 | |
| Ovary | C/C Bidirectional | 33.60 | [23.38, 43.66] | <2.20 × 10−16 |
| C/N Bidirectional | 10.42 | [6.60, 17.14] | <2.20 × 10−16 |
| Pathway | GO ID | Total | Observed | Expected | Ratio | adjP |
|---|---|---|---|---|---|---|
| DNA metabolic process | GO:0006259 | 899 | 138 | 63.98 | 2.16 | 0.00 × 100 |
| RNA processing | GO:0006396 | 851 | 130 | 60.56 | 2.15 | 0.00 × 100 |
| DNA repair | GO:0006281 | 472 | 87 | 33.59 | 2.59 | 3.11 × 10−13 |
| chromosome organization | GO:0051276 | 562 | 95 | 39.99 | 2.38 | 2.33 × 10−12 |
| ncRNA metabolic process | GO:0034660 | 535 | 91 | 38.07 | 2.39 | 5.41 × 10−12 |
| ncRNA processing | GO:0034470 | 379 | 68 | 26.97 | 2.52 | 1.29 × 10−9 |
| cellular response to DNA damage stimulus | GO:0006974 | 731 | 104 | 52.02 | 2 | 5.60 × 10−9 |
| organelle fission | GO:0048285 | 578 | 84 | 41.13 | 2.04 | 2.10 × 10−7 |
| mitochondrion organization | GO:0007005 | 599 | 86 | 42.63 | 2.02 | 2.10 × 10−7 |
| cell cycle | GO:0007049 | 1591 | 178 | 113.22 | 1.57 | 2.10 × 10−7 |
| double-strand break repair | GO:0006302 | 181 | 39 | 12.88 | 3.03 | 2.39 × 10−7 |
| nuclear division | GO:0000280 | 537 | 79 | 38.21 | 2.07 | 2.87 × 10−7 |
| cell cycle process | GO:0022402 | 1217 | 143 | 86.61 | 1.65 | 4.92 × 10−7 |
| DNA recombination | GO:0006310 | 244 | 46 | 17.36 | 2.65 | 5.42 × 10−7 |
| telomere maintenance | GO:0000723 | 119 | 29 | 8.47 | 3.42 | 1.69 × 10−6 |
| telomere organization | GO:0032200 | 122 | 29 | 8.68 | 3.34 | 2.94 × 10−6 |
| DNA conformation change | GO:0071103 | 235 | 43 | 16.72 | 2.57 | 4.03 × 10−6 |
| nucleic acid phosphodiester bond hydrolysis | GO:0090305 | 264 | 46 | 18.79 | 2.45 | 5.68 × 10−6 |
| DNA biosynthetic process | GO:0071897 | 187 | 36 | 13.31 | 2.71 | 1.53 × 10−5 |
| rRNA metabolic process | GO:0016072 | 250 | 43 | 17.79 | 2.42 | 2.25 × 10−5 |
| ribonucleoprotein complex biogenesis | GO:0022613 | 420 | 60 | 29.89 | 2.01 | 6.35 × 10−5 |
| mitotic cell cycle process | GO:1903047 | 842 | 100 | 59.92 | 1.67 | 7.95 × 10−5 |
| ribosome biogenesis | GO:0042254 | 302 | 47 | 21.49 | 2.19 | 1.07 × 10−4 |
| mRNA processing | GO:0006397 | 442 | 61 | 31.45 | 1.94 | 1.49 × 10−4 |
| rRNA processing | GO:0006364 | 243 | 40 | 17.29 | 2.31 | 1.72 × 10−4 |
| mitotic cell cycle | GO:0000278 | 926 | 106 | 65.9 | 1.61 | 1.76 × 10−4 |
| mitotic nuclear division | GO:0007067 | 411 | 57 | 29.25 | 1.95 | 2.70 × 10−4 |
| mRNA metabolic process | GO:0016071 | 631 | 78 | 44.9 | 1.74 | 3.11× 10−4 |
| chromatin organization | GO:0006325 | 676 | 82 | 48.11 | 1.7 | 3.41 × 10−4 |
| tRNA processing | GO:0008033 | 115 | 24 | 8.18 | 2.93 | 4.10 × 10−4 |
| DNA synthesis involved in DNA repair | GO:0000731 | 71 | 18 | 5.05 | 3.56 | 4.31 × 10−4 |
| mitochondrial translation | GO:0032543 | 117 | 24 | 8.33 | 2.88 | 5.28 × 10−4 |
| DNA-templated transcription, termination | GO:0006353 | 94 | 21 | 6.69 | 3.14 | 5.28 × 10−4 |
| chromosome segregation | GO:0007059 | 305 | 45 | 21.7 | 2.07 | 5.64 × 10−4 |
| cellular macromolecular complex assembly | GO:0034622 | 876 | 99 | 62.34 | 1.59 | 5.64 × 10−4 |
| protein folding | GO:0006457 | 204 | 34 | 14.52 | 2.34 | 6.34 × 10−4 |
| regulation of chromosome organization | GO:0033044 | 128 | 25 | 9.11 | 2.74 | 7.25 × 10−4 |
| regulation of organelle organization | GO:0033043 | 963 | 106 | 68.53 | 1.55 | 7.58 × 10−4 |
| DNA packaging | GO:0006323 | 155 | 28 | 11.03 | 2.54 | 9.00 × 10−4 |
| mitochondrial translational elongation | GO:0070125 | 83 | 19 | 5.91 | 3.22 | 9.01 × 10−4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, J.A.; Christensen, B.C.; Marsit, C.J. Pan-Cancer Analysis Reveals Differential Susceptibility of Bidirectional Gene Promoters to DNA Methylation, Somatic Mutations, and Copy Number Alterations. Int. J. Mol. Sci. 2018, 19, 2296. https://doi.org/10.3390/ijms19082296
Thompson JA, Christensen BC, Marsit CJ. Pan-Cancer Analysis Reveals Differential Susceptibility of Bidirectional Gene Promoters to DNA Methylation, Somatic Mutations, and Copy Number Alterations. International Journal of Molecular Sciences. 2018; 19(8):2296. https://doi.org/10.3390/ijms19082296
Chicago/Turabian StyleThompson, Jeffrey A., Brock C. Christensen, and Carmen J. Marsit. 2018. "Pan-Cancer Analysis Reveals Differential Susceptibility of Bidirectional Gene Promoters to DNA Methylation, Somatic Mutations, and Copy Number Alterations" International Journal of Molecular Sciences 19, no. 8: 2296. https://doi.org/10.3390/ijms19082296
APA StyleThompson, J. A., Christensen, B. C., & Marsit, C. J. (2018). Pan-Cancer Analysis Reveals Differential Susceptibility of Bidirectional Gene Promoters to DNA Methylation, Somatic Mutations, and Copy Number Alterations. International Journal of Molecular Sciences, 19(8), 2296. https://doi.org/10.3390/ijms19082296