Hepatoprotective Effects of Lactobacillus on Carbon Tetrachloride-Induced Acute Liver Injury in Mice

 and
and
Abstract
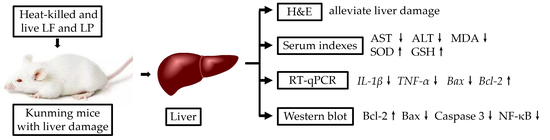
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
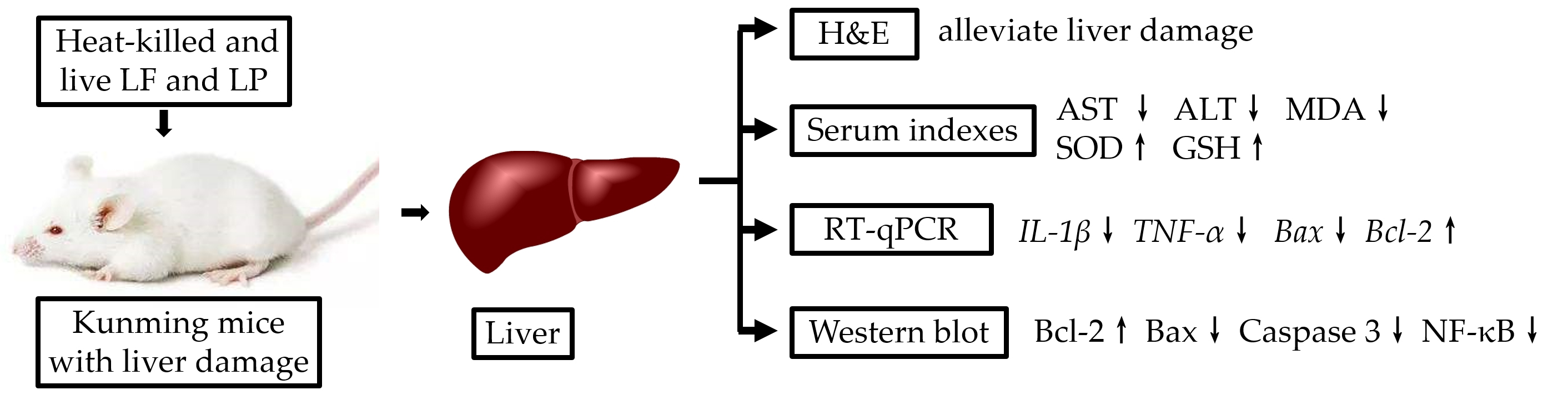
2.1. Morphological Characteristics
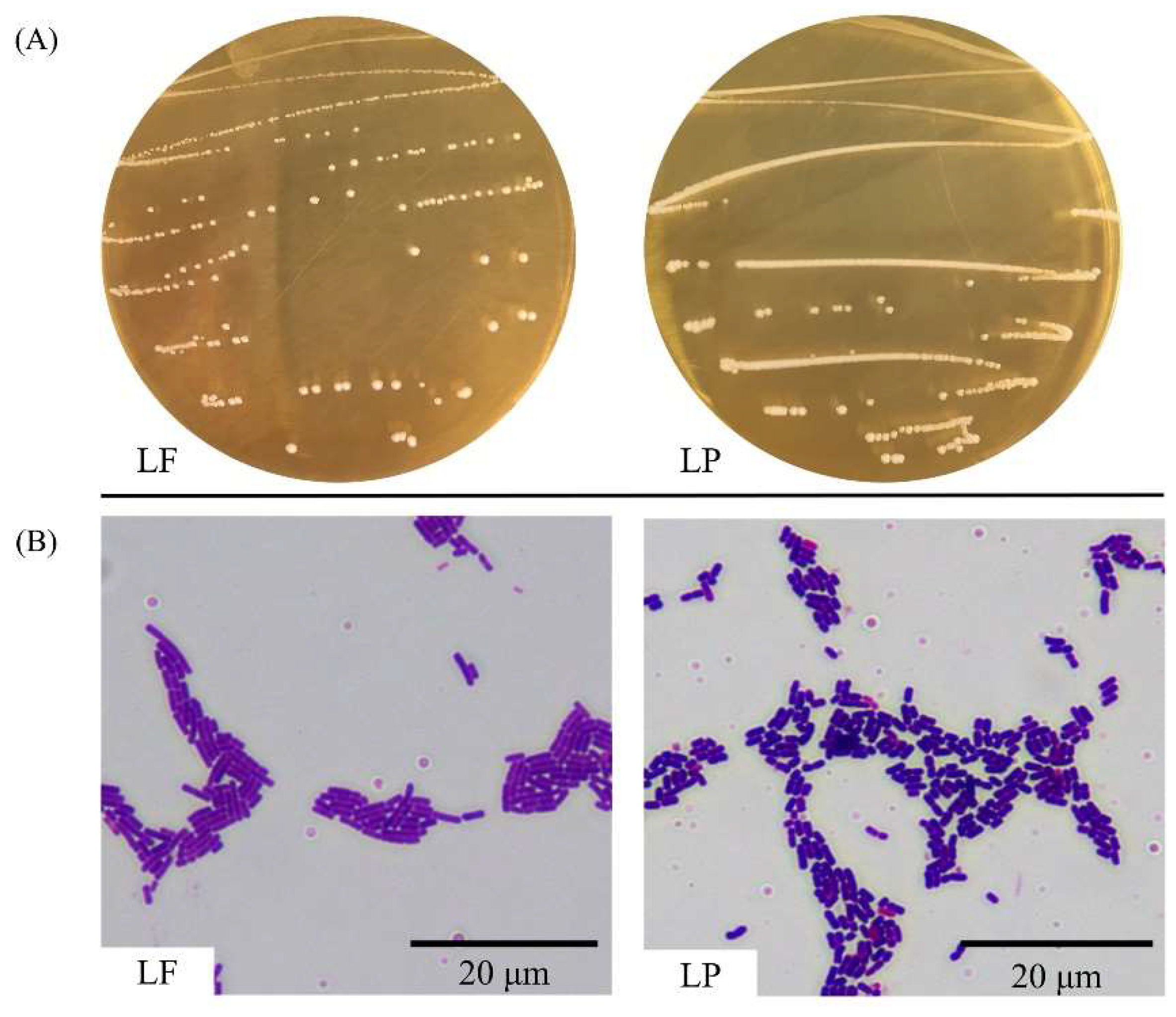
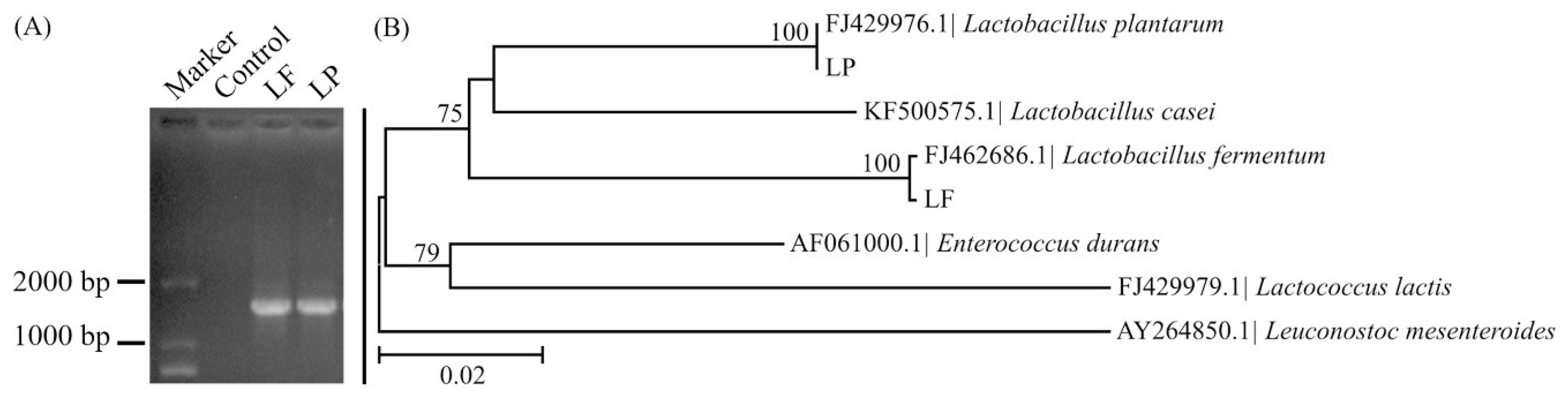
2.2. Species Analysis
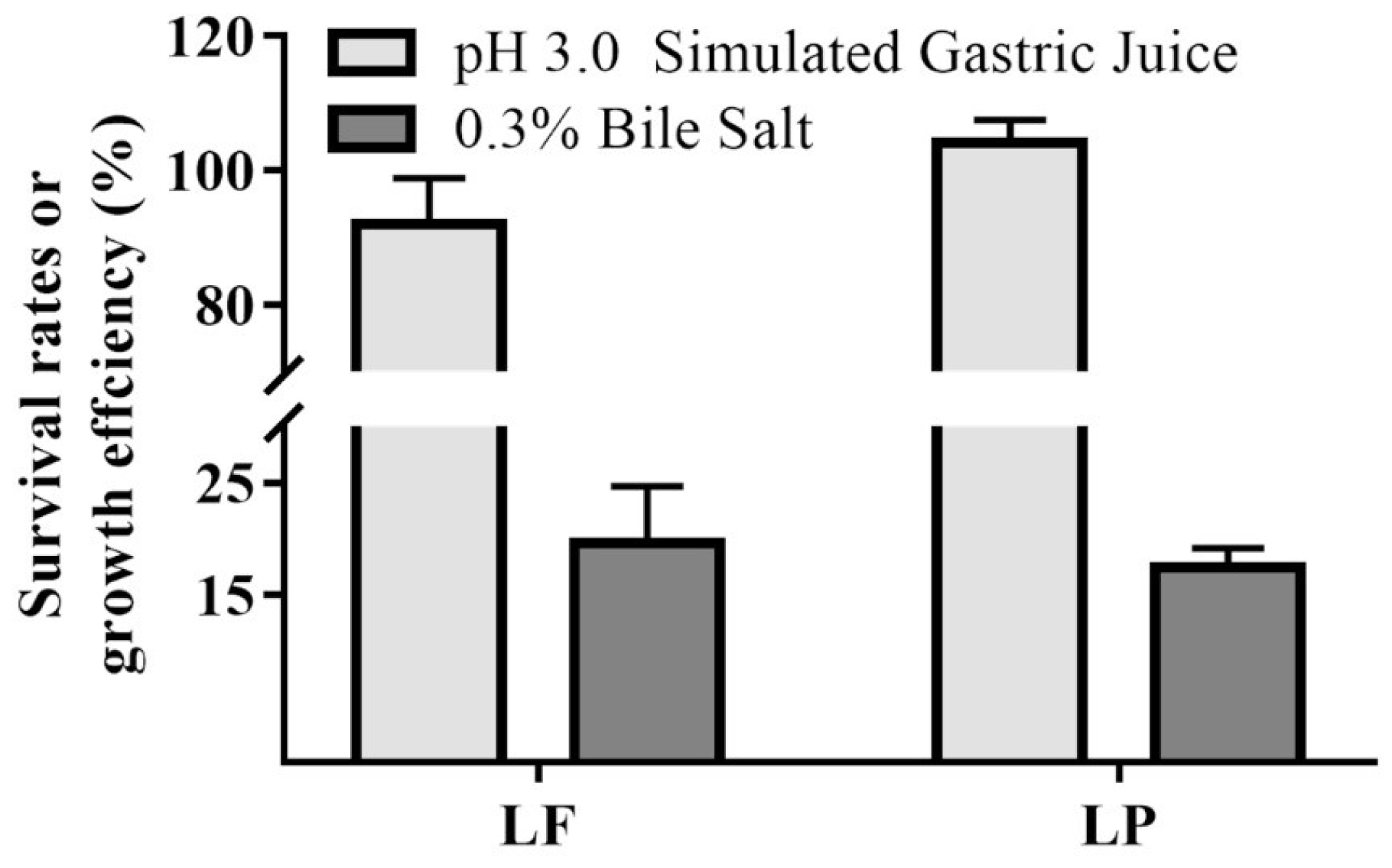
2.3. Tolerance to Stimulated Gastric Juice and Bile Salt
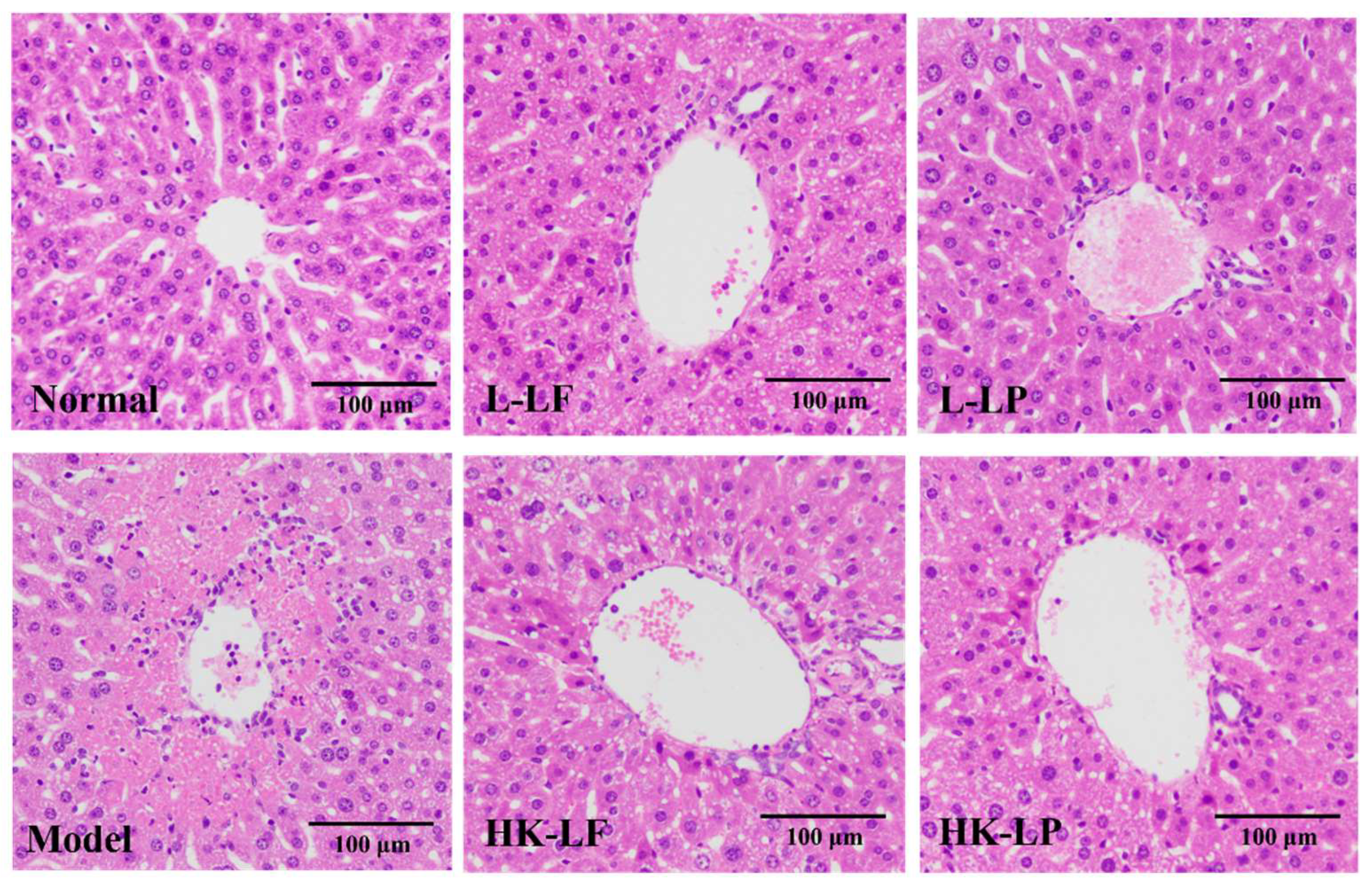
2.4. Pathological Observation
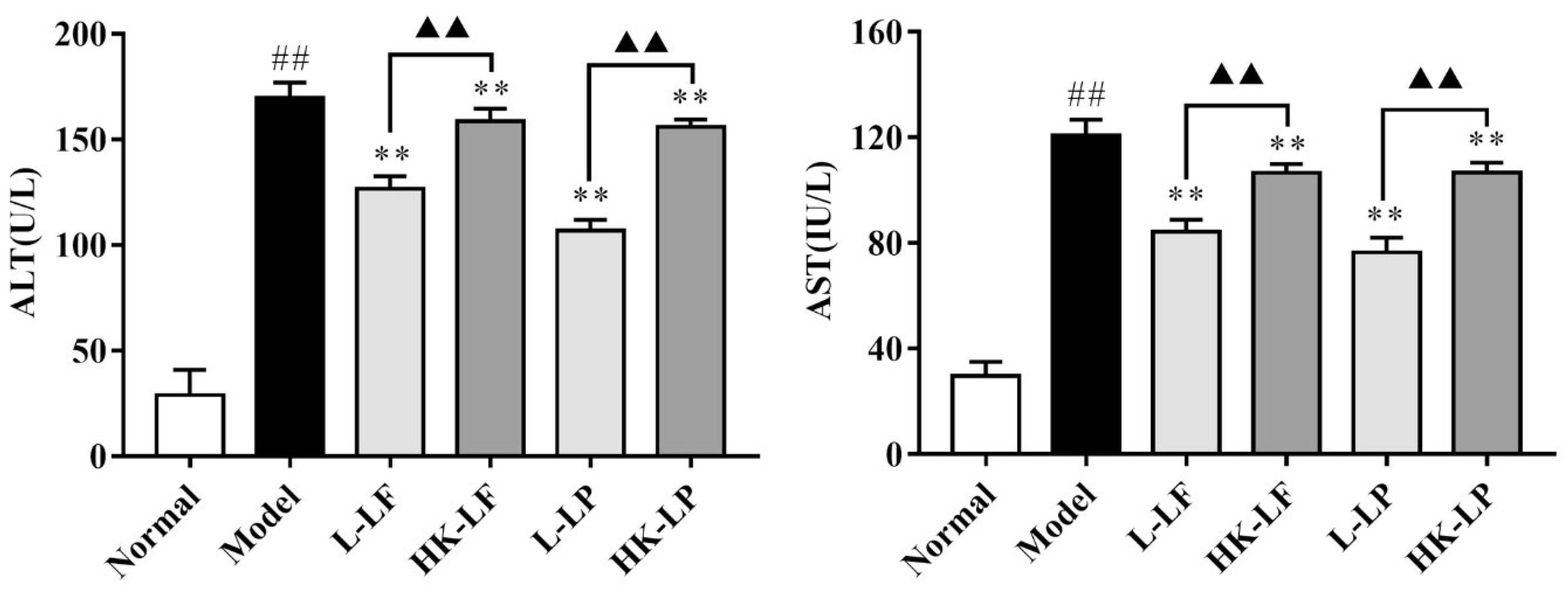
2.5. Measurement of ALT and AST
2.6. Measurement of SOD, GSH, and MDA
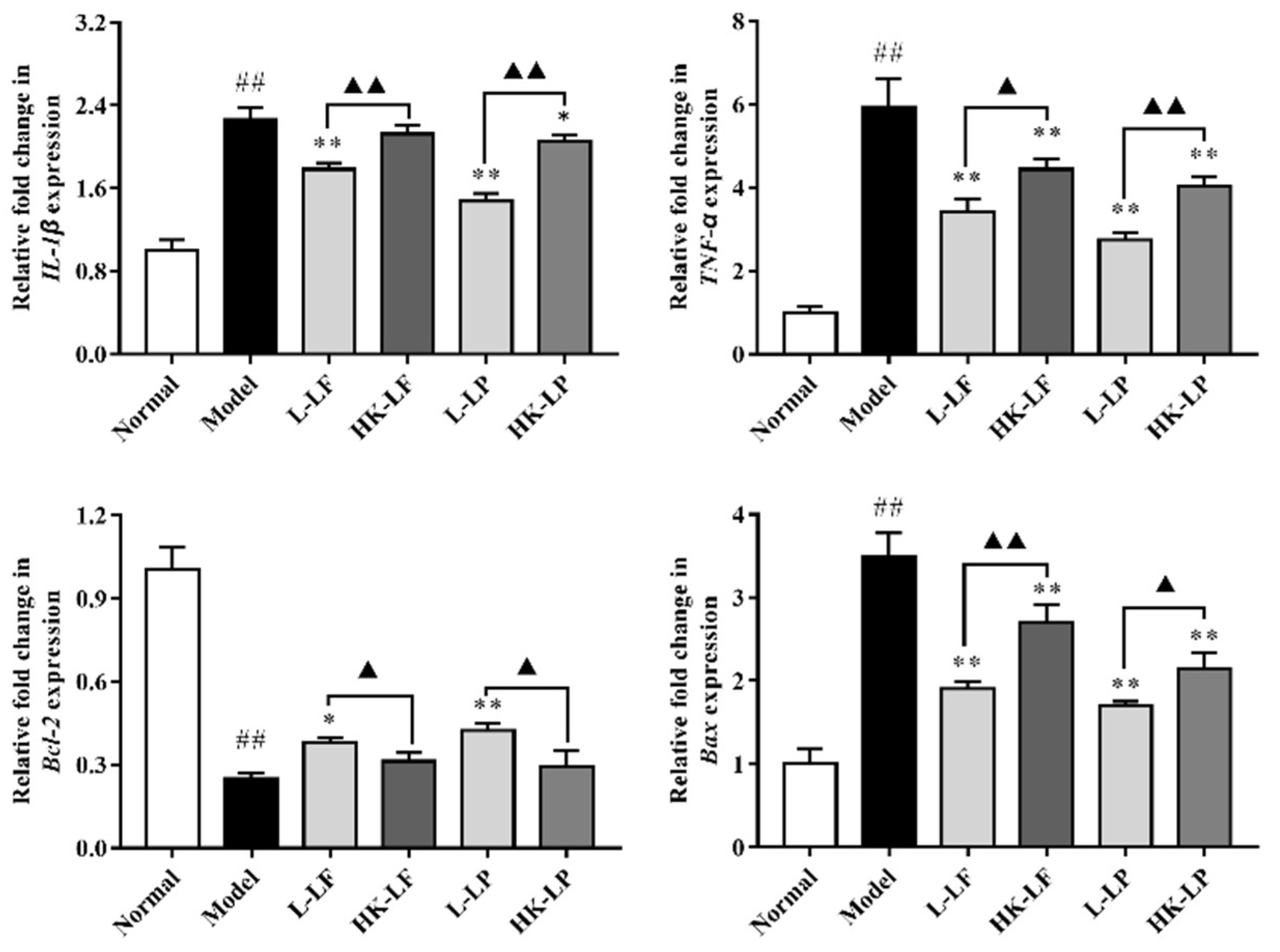
2.7. RT-qPCR
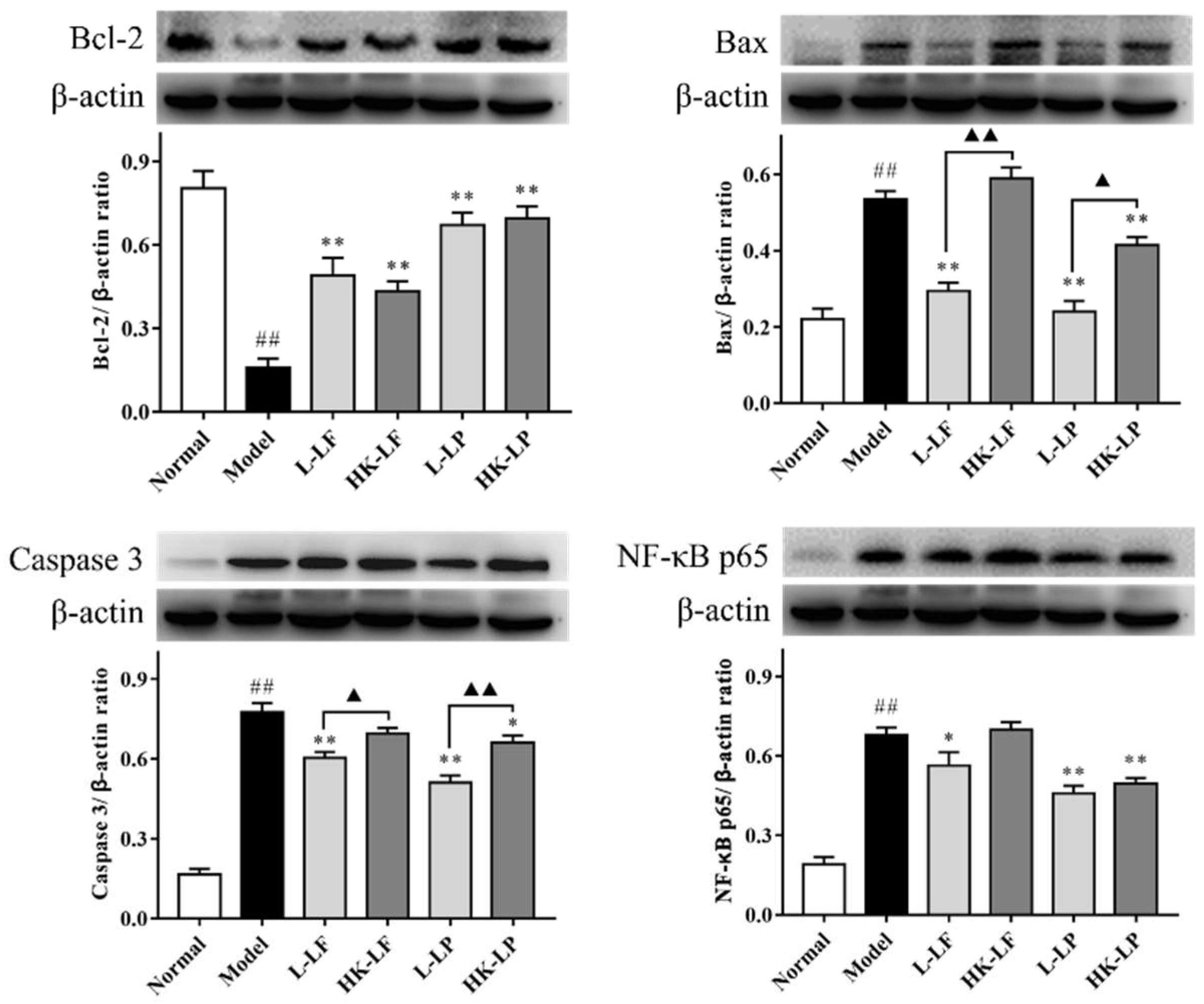
2.8. Western Blot
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Evaluation Gastrointestinal Tolerability in Vitro
4.3. Animal Models and Treatment
4.4. Histological Observations
4.5. Measurement of Biochemical Parameters in Serum
4.6. RT-qPCR (Real-Time Quantitative Polymerase Chain Reaction) Analyses
4.7. Western Blot
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, H.; Qiu, Y.; Shu, Z.; Zhang, X.; Li, R.; Liu, S.; Chen, L.; Liu, H.; Chen, N. Protective effect of Trillium tschonoskii saponin on CCl4-induced acute liver injury of rats through apoptosis inhibition. Can. J. Physiol. Pharmacol. 2016, 94, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Liu, X.; Tang, G.; Liu, H.; Zhang, Y.; Zhang, B.; Zhao, X.; Wang, W. Ethanol extract of Portulaca Oleracea, L. Reduced the carbon tetrachloride induced liver injury in mice involving enhancement of NF-κB activity. Am. J. Transl. Res. 2014, 6, 746–755. [Google Scholar] [PubMed]
- Dong, Y.; Liu, Y.; Kou, X.; Jing, Y.; Sun, K.; Sheng, D.; Yu, G.; Yu, D.; Zhao, Q.; Zhao, X.; et al. The protective or damaging effect of Tumor necrosis factor-α in acute liver injury is concentration-dependent. Cell Biosci. 2016, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, M.; Makki, K.; Storelli, G.; Machuca-Gayet, I.; Srutkova, D.; Hermanova, P.; Martino, M.E.; Balmand, S.; Hudcovic, T.; Heddi, A.; et al. Lactobacillus plantarum strain maintains growth of infant mice during chronic undernutrition. Science 2016, 351, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Ozaki, M.; Tamura, A.; Yamada, N.; Ishida, T.; Hosoda, M.; Hosono, A. Antidiabetic effect of Lactobacillus GG in streptozotocin-induced diabetic rats. Biosci. Biotechnol. Biochem. 2003, 67, 1421–1424. [Google Scholar] [CrossRef] [PubMed]
- Tien, M.T.; Girardin, S.E.; Regnault, B.; Le Bourhis, L.; Dillies, M.A.; Coppee, J.Y.; Bourdet-Sicard, R.; Sansonetti, P.J.; Pedron, T. Anti-inflammatory effect of Lactobacillus casei on Shigella-infected human intestinal epithelial cells. J. Immunol. 2006, 176, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Ostad, S.N.; Salarian, A.A.; Ghahramani, M.H.; Fazeli, M.R.; Samadi, N.; Jamalifar, H. Live and heat-inactivated lactobacilli from feces inhibit Salmonella typhi and Escherichia coli adherence to Caco-2 cells. Folia Microbiol. 2009, 54, 157–160. [Google Scholar] [CrossRef] [PubMed]
- De, D.C.; Rodrigues, G.; Bona, S.; Meurer, L.; González-gallego, J.; Tuñón, M.J.; Marroni, N.P. Role of quercetin in preventing thioacetamide-induced liver injury in rats. Toxicol. Pathol. 2011, 39, 949–957. [Google Scholar]
- Chen, D.; McKallip, R.J.; Zeytun, A.; Do, Y.; Lombard, C.; Robertson, J.L.; Mak, T.W.; Nagarkatti, P.S.; Nagarkatti, M. CD44-deficient mice exhibit enhanced hepatitis after concanavalin A injection: Evidence for involvement of CD44 in activation-induced cell death. J. Immunol. 2001, 166, 5889–5897. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Kim, J.; Bang, H.; Moon, J.; Kim, G.O.; Shin, T. Hepatoprotective effects of allyl isothiocyanate against carbon tetrachloride-induced hepatotoxicity in rat. Chem. Biol. Interact. 2016, 254, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Ren, N.; Gao, H.; Lei, X.; Zheng, J.; Cao, W. Antioxidant and hepatoprotective effects of Schisandra chinensis pollen extract on CCl4-induced acute liver damage in mice. Food Chem. Toxicol. 2013, 55, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Pan, D.D.; Zhou, J.; Jiang, Y.Z. Protective effect of selenium-enriched Lactobacillus on CCl4-induced liver injury in mice and its possible mechanisms. World J. Gastroenterol. 2005, 11, 5795–5800. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fu, Y.; Zhang, H.; Wang, J.; Zhu, J.; Wang, Y.; Guo, Y.; Wang, G.; Xu, T.; Chu, M.; et al. The hepatoprotective effect of the probiotic Clostridium butyricum against carbon tetrachloride-induced acute liver damage in mice. Food Funct. 2017, 8, 4042–4052. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ding, J.; Zhang, L.; Liu, C. Hepatoprotective properties of sesamin against CCl4 induced oxidative stress-mediated apoptosis in mice via JNK pathway. Food Chem. Toxicol. 2014, 64, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Fardet, A.; Rock, E. In vitro and in vivo antioxidant potential of milks, yoghurts, fermented milks and cheeses: A narrative review of evidence. Nutr. Res. Rev. 2017, 2, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Persichetti, E.; De Michele, A.; Codini, M.; Traina, G. Antioxidative capacity of Lactobacillus fermentum LF31 evaluated in vitro by oxygen radical absorbance capacity assay. Nutrition 2014, 30, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Ge, P.; Jiang, D.; Du, N.; Chen, J.; Yuan, L.; Yu, H.; Xu, X.; Wu, M.; Zhang, W.; et al. A novel and simple cell-based electrochemical biosensor for evaluating the antioxidant capacity of Lactobacillus plantarum strains isolated from Chinese dry-cured ham. Biosens. Bioelectron. 2018, 99, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Marazza, J.A.; Nazareno, M.A.; de Giori, G.S.; Garro, M.S. Enhancement of the antioxidant capacity of soymilk by fermentation with Lactobacillus rhamnosus. J. Funct. Foods. 2012, 4, 594–601. [Google Scholar] [CrossRef]
- Hunaefi, D.; Akumo, D.N.; Riedel, H.; Smetanska, I. The Effect of Lactobacillus plantarum ATCC 8014 and Lactobacillus acidophilus NCFM Fermentation on Antioxidant Properties of Selected in Vitro Sprout Culture of Orthosiphon aristatus (Java Tea) as a Model Study. Antioxidants 2012, 1, 4–32. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, F.; Lan, C.E.; Huang, T.; Chen, K.; Chai, C.; Chen, W.; Fang, A.; Chen, Y.; Wu, C. Heat-killed and live Lactobacillus reuteri GMNL-263 exhibit similar effects on improving metabolic functions in high-fat diet-induced obese rats. Food Funct. 2016, 7, 2374–2388. [Google Scholar] [CrossRef] [PubMed]
- Yuki, N.; Watanabe, K.; Mike, A.; Tagami, Y.; Tanaka, R.; Ohwaki, M.; Morotomi, M. Survival of a probiotic, Lactobacillus casei strain Shirota, in the gastrointestinal tract: Selective isolation from faeces and identification using monoclonal antibodies. Int. J. Food Microbiol. 1999, 48, 51–57. [Google Scholar] [CrossRef]
- Gomathi, S.; Sasikumar, P.; Anbazhagan, K.; Sasikumar, S.; Kavitha, M.; Selvi, M.S.; Selvam, G.S. Screening of indigenous oxalate degrading lactic acid bacteria from human faeces and South Indian fermented foods: Assessment of probiotic potential. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Suo, H.; Zhao, X.; Qian, Y.; Li, G.; Liu, Z.; Xie, J.; Li, J. Therapeutic effect of activated carbon-induced constipation mice with Lactobacillus fermentum Suo on treatment. Int. J. Mol. Sci. 2014, 15, 21875–21895. [Google Scholar] [CrossRef] [PubMed]
- Brautbar, N.; Williams, J.N. Industrial solvents and liver toxicity: Risk assessment, risk factors and mechanisms. Int. J. Hyg. Environ. Health 2002, 205, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Al-Jameil Noura, N. Liver Function Tests as Probable Markers of Preeclampsia—A Prospective Study Conducted in Riyadh. JCAM 2015, 6, 461–464. [Google Scholar] [CrossRef]
- Jain, A.; Soni, M.; Deb, L.; Jain, A.; Rout, S.P.; Gupta, V.B.; Krishna, K.L. Antioxidant and hepatoprotective activity of ethanolic and aqueous extracts of Momordica dioica Roxb. leaves. J. Ethnopharmacol. 2008, 115, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Miesel, R.; Sanocka, D.; Kurpisz, M.; Kr Ger, H. Antiinflammatory effects of NADPH oxidase inhibitors. Inflammation 1995, 19, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wang, F.; Wu, C. Hepatoprotective Effects of Apple Polyphenols on CCl4-Induced Acute Liver Damage in Mice. J. Agric. Food Chem. 2010, 58, 6525–6531. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Horvitz, H.R. A first insight into the molecular mechanisms of apoptosis. Cell 2004, 116, S53–S56. [Google Scholar] [CrossRef]
- Garrido, C.; Galluzzi, L.; Brunet, M.; Puig, P.E.; Didelot, C.; Kroemer, G. Mechanisms of cytochrome c release from mitochondria. Cell Death Differ. 2006, 13, 1423–1433. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Schwabe, R.F. NF-κB in the liver-linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroentrol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Qamar, A.A. Probiotics in nonalcoholic fatty liver disease, nonalcoholic steatohepatitis, and cirrhosis. J. Clin. Gastroenterol. 2015, 49, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Bensalah, F.; Delorme, C.; Renault, P. Characterisation of thermotolerant cocci from indigenous flora of ‘leben’ in algerian arid area and DNA identification of atypical Lactococcus lactis strains. Curr. Microbiol. 2009, 59, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of potential probiotic properties of Lactobacillus fermentum isolated from traditional dairy products. Food Control. 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Walker, D.K.; Gilliland, S.E. Relationship among bile tolerance, bile salt deconjugation, and assimilation of cholesterol by Lactobacillus acidophilus. J. Dairy Sci. 1993, 76, 956–961. [Google Scholar] [CrossRef]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Zhang, J.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Hepatoprotective Effects of Lactobacillus on Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Int. J. Mol. Sci. 2018, 19, 2212. https://doi.org/10.3390/ijms19082212
Chen X, Zhang J, Yi R, Mu J, Zhao X, Yang Z. Hepatoprotective Effects of Lactobacillus on Carbon Tetrachloride-Induced Acute Liver Injury in Mice. International Journal of Molecular Sciences. 2018; 19(8):2212. https://doi.org/10.3390/ijms19082212
Chicago/Turabian StyleChen, Xiaoyong, Jing Zhang, Ruokun Yi, Jianfei Mu, Xin Zhao, and Zhennai Yang. 2018. "Hepatoprotective Effects of Lactobacillus on Carbon Tetrachloride-Induced Acute Liver Injury in Mice" International Journal of Molecular Sciences 19, no. 8: 2212. https://doi.org/10.3390/ijms19082212
APA StyleChen, X., Zhang, J., Yi, R., Mu, J., Zhao, X., & Yang, Z. (2018). Hepatoprotective Effects of Lactobacillus on Carbon Tetrachloride-Induced Acute Liver Injury in Mice. International Journal of Molecular Sciences, 19(8), 2212. https://doi.org/10.3390/ijms19082212