Insulin-Like Growth Factor (IGF) System in Liver Diseases


Abstract
1. Introduction
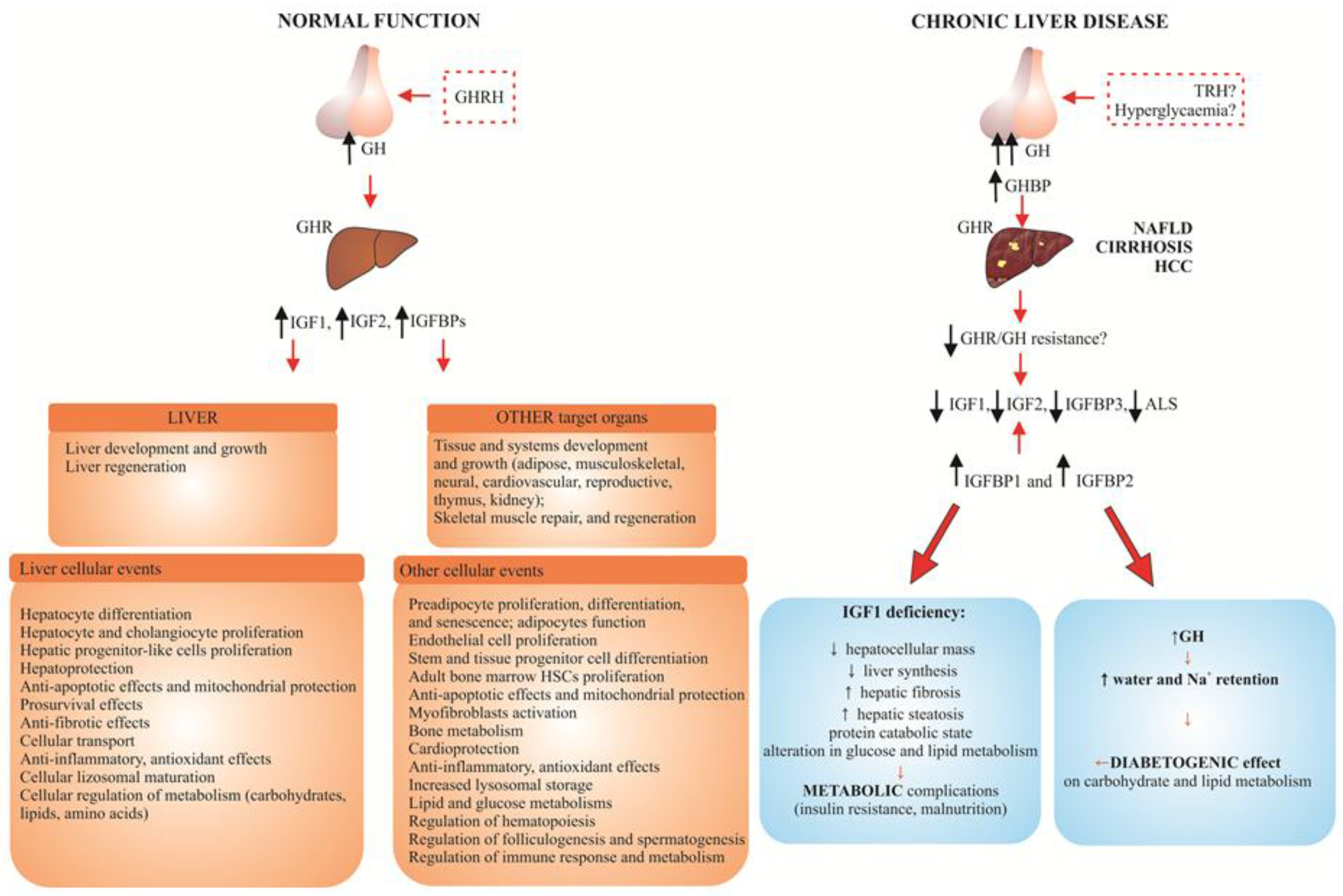
2. Role of IGF System in the Pathogenesis of Chronic Liver Disease (CLD)
3. Evidence from Animal Models
3.1. Role in Nonalcoholic Fatty Liver Disease (NAFLD)
3.2. Role in Liver Cirrhosis
3.3. Role in Hepatocellular Carcinoma (HCC)
4. Evidence from In Vitro Studies
4.1. Role in Nonalcoholic Fatty Liver Disease (NAFLD)
4.2. Role in Liver Cirrhosis
4.3. Role in Hepatocellular Carcinoma (HCC)
5. Evidence from Epidemiologic and Clinicopathological Studies
5.1. Role in Nonalcoholic Fatty Liver Disease (NAFLD)
5.2. Role in Liver Cirrhosis
5.3. Role in Hepatocellular Carcinoma (HCC)
6. Genomic Alterations of IGF System in CLD
7. IGF System in CLD—Summary and Outstanding Questions
- The studies on the IGF1R signaling pathway in CLD using animal models and in vitro studies, allowed for the clarification of general aspects of hepatocarcinogenesis (cell cycle progression, uncontrolled proliferation, cell survival, migration, inhibition of apoptosis, protein synthesis and cell growth), as well as show that systemic IGF1 administration can reduce fibrosis and ameliorate general liver function. This allowed them to become promising factors in development of new methods of managing the most common chronic liver diseases.
- The results of several careful epidemiologic studies concerning CLD show that low IGF1 serum level is a typical indicator of decreased hepatic reserve in NAFLD, liver cirrhosis and HCC (Table 1, Figure 1). Different values of lowered serum concentration of both IGFs were proposed as a negative prognostic factor in patients with liver cirrhosis (IGF1: 13, 30 ng/mL; IGF2: below 200 ng/mL). In the majority of HCC patients, the inverse association between serum IGFBP3 level and the risk of cancer was observed.
- Clinicopathological data involving tissue expression of IGF system components in patients with CLD confirmed that decreased IGF1 secretion and increased expression of IGF1R, accompanied by a reduced expression of IGFBPs (mainly of IGFBP3), as compared to the control, are negative prognostic markers in CLD, independent prognostic factors of survival (IGFBP1), or may function as a tumor suppressor (IGFBP7) in HCC.
- The main outstanding question still remains—is the decreased synthesis of the IGF system components due to chronic damage of liver parenchyma, or is damaged parenchyma an effect of an altered function of GH/IGF/IGFR system?
- Promising results of the studies on IGF system are coming from genetic and epigenetic investigations, as well as from research on the role of various isoforms of IGF system components in hepatocarcinogenesis, and cooperation between functioning IGF1R and many viral and cellular oncogenes.
- Concluding the article, we believe that addressing some of those questions will be essential to fully understand the mechanisms of IGF system signaling in chronic liver diseases and to develop new IGF-based biomarkers and treatment strategies for liver cancer.
Author Contributions
Conflicts of Interest
Abbreviations
| AA | adenine-adenine |
| ADAMTS9 | a disintegrin and metalloproteinase with thrombospondin motifs 9 |
| AFP | alpha-fetoprotein |
| Akt (AKT) (protein kinase B) | serine/threonine protein kinase |
| ALD | alcoholic liver disease |
| ALS | acid-labile protein subunit |
| aPTT | Activated Partial Thromboplastin Time |
| BMI | body mass index |
| CLD | chronic liver diseases |
| CT | cytosine-thymine |
| ECM | extracellular matrix |
| EGF/R | epidermal growth factor/receptor |
| ERK | extracellular signal-regulated kinase |
| GG | guanine-guanine |
| GH | growth hormone |
| HBV/HCV | hepatitis B/C virus |
| HCC | hepatocellular carcinoma |
| HGF | hepatocyte growth factor |
| HIV | human immunodeficiency virus |
| HSCs | human stellate cells |
| IGF | insulin-like growth factor |
| IGFR | IGF receptor |
| IGFBPs | IGF binding proteins |
| IGFBPrP1-10 | IGF IGFBP-related proteins 1-10 |
| IL | interleukin |
| INR | International Normalized Ratio of prothrombin time of blood coagulation |
| IR | insulin receptor |
| IRS-1/2 | insulin receptor substrate 1/2 |
| kDa | kilodalton |
| LH | luteinizing hormone |
| MAPK | mitogen-activated protein kinase (originally called ERK) |
| MELD | Model of End-Stage Liver Disease |
| NAFLD | nonalcoholic fatty liver disease |
| NASH | nonalcoholic steatohepatitis |
| PI3K | phosphatidylinositol 3-kinase |
| STAT | signal transducer and activator of transcription |
| TGF-α, -β | transforming growth factor α, -β |
| TNF-α | tumor necrosis factor α |
| TNM | classification of Malignant Tumors (T—tumor; N—lymph nodes; M—metastasis) |
| TT | thymine-thymine |
References
- Postic, C.; Dentin, R.; Girard, J. Role of the liver in the control of carbohydrate and lipid homeostasis. Diabetes Metab. 2004, 30, 398–408. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, W.; Pu, Z.; Li, X.; Lei, X.; Yao, J.; Yang, X. Developmental changes of Insulin-like growth factors in the liver and muscle of chick embryos. Poult. Sci. 2016, 95, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Voci, A.; Arvigo, M.; Massajoli, M.; Garrone, S.; Bottazzi, C.; Demori, I.; Gallo, G. IGF-I production by adult rat hepatocytes is stimulated by transforming growth factor-alpha and transforming growth factor-beta1. Eur. J. Endocrinol. 1999, 140, 577–582. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rosen, C.J. Serum insulin-like growth factors and insulin-like growth factor-binding proteins: Clinical implications. Clin. Chem. 1999, 45, 1384–1390. [Google Scholar] [PubMed]
- Shin, M.; Kang, H.S.; Park, J.H.; Bae, J.H.; Song, D.K.; Im, S.S. Recent Insights into Insulin-Like Growth Factor Binding Protein 2 Transcriptional Regulation. Endocrinol. Metab. 2017, 32, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Daughaday, W.H.; Rotwein, P. Insulin-like growth factors I and II. Peptide, messenger ribonucleic acid and gene structures, serum, and tissue concentrations. Endocr. Rev. 1989, 10, 68–91. [Google Scholar] [CrossRef] [PubMed]
- Weroha, S.J.; Haluska, P. The insulin-like growth factor system in cancer. Endocrinol. Metab. Clin. N. Am. 2012, 41, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Humbel, R.E. Insulin-like growth factors I and II. Eur. J. Biochem. 1990, 190, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, C. IGF2 and cancer. Endocr. Relat. Cancer 2013, 20, R321–R339. [Google Scholar] [CrossRef] [PubMed]
- Rotwein, P.; Naylor, S.L.; Chirgwin, J.M. Human insulin-related DNA sequences map to chromosomes 2 and 11. Somat. Cell Mol. Genet. 1986, 12, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Von Horn, H.; Ekström, C.; Ellis, E.; Olivecrona, H.; Einarsson, C.; Tally, M.; Ekström, T.J. GH is a regulator of IGF2 promoter-specific transcription in human liver. J. Endocrinol. 2002, 172, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Rotwein, P. Diversification of the insulin-like growth factor 1 gene in mammals. PLoS ONE 2017, 12, e0189642. [Google Scholar] [CrossRef] [PubMed]
- Rotwein, P.; Chia, D.J. Gene regulation by growth hormone. Pediatr. Nephrol. 2010, 25, 651–658. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Durzyńska, J. IGF axis and other factors in HPV-related and HPV-unrelated carcinogenesis (review). Oncol. Rep. 2014, 32, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Kwasniewski, W.; Adamek, A.; Gozdzicka-Jozefiak, A. Insulin-like growth factor (IGF) axis in cancerogenesis. Mutat. Res. Rev. Mutat. Res. 2017, 772, 78–104. [Google Scholar] [CrossRef] [PubMed]
- Rechler, M.M.; Clemmonds, D.R. Regulatory actions of insulin-like growth factor-binding proteins. Trends Endocrinol. Metab. 1998, 9, 176–183. [Google Scholar] [CrossRef]
- Clemmons, D.R. Role of insulin-like growth factor in maintaining normal glucose homeostasis. Horm. Res. 2004, 62 (Suppl. S1), 77–82. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, D.R. The relative roles of growth hormone and IGF-1 in controlling insulin sensitivity. J. Clin. Investig. 2004, 113, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Hwa, V.; Oh, Y.; Rosenfeld, R.G. The insulin-like growth factor-binding protein (IGFBP) superfamily. Endocr. Rev. 1999, 20, 761–787. [Google Scholar] [CrossRef] [PubMed]
- Wheatcroft, S.B.; Kearney, M.T. IGF-dependent and IGF-independent actions of IGF-binding protein-1 and -2: Implications for metabolic homeostasis. Trends Endocrinol. Metab. 2009, 20, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Phillips, L.S.; Pao, C.I.; Villafuerte, B.C. Molecular regulation of insulin-like growth factor-I and its principal binding protein, IGFBP-3. Prog. Nucleic Acid. Res. Mol. Biol. 1998, 60, 195–265. [Google Scholar] [PubMed]
- Murphy, L.J. Insulin-like growth factor-binding proteins: Functional diversity or redundancy? J. Mol. Endocrinol. 1998, 21, 97–107. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Baserga, R. The insulin-like growth factor I receptor: A key to tumor growth? Cancer Res. 1995, 55, 249–255. [Google Scholar] [PubMed]
- LeRoith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The somatomedin hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Adamek, A. The insulin-like growth factor (IGF) signaling axis and hepatitis C virus-associated carcinogenesis (review). Int. J. Oncol. 2012, 41, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Nalbantoglu, I.L.; Brunt, E.M. Role of liver biopsy in nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 9026–9037. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.; Shazi, L. Pattern and profile of chronic liver disease in acute on chronic liver failure. Hepatol. Int. 2015, 9, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Wree, A.; Broderick, L.; Canbay, A.; Hoffman, H.M.; Feldstein, A.E. From NAFLD to NASH to cirrhosis-new insights into disease mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment. Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Issa, D.; Patel, V.; Sanyal, A. Future therapy for non-alcoholic fatty liver disease. Liver Int. 2018, 38, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Finelli, C. What about non-alcoholic fatty liver disease as a new criterion to define metabolic syndrome? World J. Gastroenterol. 2013, 19, 3375–3384. [Google Scholar] [CrossRef] [PubMed]
- Hribal, M.L.; Procopio, T.; Petta, S.; Sciacqua, A.; Grimaudo, S.; Pipitone, R.M.; Perticone, F.; Sesti, G. Insulin-like growth factor-I, inflammatory proteins, and fibrosis in subjects with nonalcoholic fatty liver disease. J. Clin. Endocrinol. Metab. 2013, 98, E304–E308. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y. The Role of Growth Hormone and Insulin-Like Growth Factor-I in the Liver. Int. J. Mol. Sci. 2017, 18, 1447. [Google Scholar] [CrossRef] [PubMed]
- Bonefeld, K.; Møller, S. Insulin-like growth factor-I and the liver. Liver Int. 2011, 31, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Puche, J.E.; Castilla-Cortázar, I. Human conditions of insulin-like growth factor-I (IGF-I) deficiency. J. Transl. Med. 2012, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Mazzanti, R.; Arena, U.; Tassi, R. Hepatocellular carcinoma: Where are we? World J. Exp. Med. 2016, 6, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Nakao, K.; Hamasaki, K.; Furukawa, R.; Tsuruta, S.; Ueda, Y.; Taura, N.; Shibata, H.; Fujimoto, M.; Toriyama, K.; et al. Role of growth hormone, insulin-like growth factor 1 and insulin-like growth factor-binding protein 3 in development of non-alcoholic fatty liver disease. Hepatol. Int. 2007, 1, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Carbone, D.L.; Zuloaga, D.G.; Hiroi, R.; Foradori, C.D.; Legare, M.E.; Handa, R.J. Prenatal dexamethasone exposure potentiates diet-induced hepatosteatosis and decreases plasma IGF-I in a sex-specific fashion. Endocrinology 2012, 153, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Hogg, K.; Wood, C.; McNeilly, A.S.; Duncan, W.C. The in utero programming effect of increased maternal androgens and a direct fetal intervention on liver and metabolic function in adult sheep. PLoS ONE 2011, 6, e24877. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Takahashi, M.; Fukuoka, H.; Iguchi, G.; Kitazawa, R.; Takahashi, Y. GH-independent IGF-I action is essential to prevent the development of nonalcoholic steatohepatitis in a GH-deficient rat model. Biochem. Biophys. Res. Commun. 2012, 423, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Iguchi, G.; Fukuoka, H.; Takahashi, M.; Suda, K.; Bando, H.; Matsumoto, R.; Yoshida, K.; Odake, Y.; Ogawa, W.; et al. IGF-I induces senescence of hepatic stellate cells and limits fibrosis in a p53-dependent manner. Sci. Rep. 2016, 6, 34605. [Google Scholar] [CrossRef] [PubMed]
- Makovicky, P.; Tumova, E.; Volek, Z.; Makovicky, P.; Vodickova, L.; Slyskova, J.; Svoboda, M.; Rejhova, A.; Vodicka, P.; Samasca, G.; et al. Histopathological aspects of liver under variable food restriction: Has the intense one-week food restriction a protective effect on non-alcoholic-fatty-liver-disease (NAFLD) development? Pathol. Res. Pract. 2014, 210, 855–862. [Google Scholar] [CrossRef] [PubMed]
- De Minicis, S.; Agostinelli, L.; Rychlicki, C.; Sorice, G.P.; Saccomanno, S.; Candelaresi, C.; Giaccari, A.; Trozzi, L.; Pierantonelli, I.; Mingarelli, E.; et al. HCC development is associated to peripheral insulin resistance in a mouse model of NASH. PLoS ONE 2014, 9, e97136. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.M.; Laggai, S.; Van Wonterg, E.; Gemperlein, K.; Müller, R.; Haybaeck, J.; Vandenbroucke, R.E.; Ogris, M.; Libert, C.; Kiemer, A.K. Transient Hepatic Overexpression of Insulin-Like Growth Factor 2 Induces Free Cholesterol and Lipid Droplet Formation. Front. Physiol. 2016, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.; Sobrevals, L.; Zaratiegui, M.; Martinez, L.; Palencia, B.; Rodríguez, C.M.; Prieto, J.; Fortes, P. Liver transduction with a simian virus 40 vector encoding insulin-like growth factor I reduces hepatic damage and the development of liver cirrhosis. Gene Ther. 2007, 14, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Picardi, A.; de Oliveira, A.C.; Muguerza, B.; Tosar, A.; Quiroga, J.; Castilla-Cortazar, I.; Santidrian, S.; Prieto, J. Low doses of insulin-like growth factor-I improve nitrogen retention and food efficiency in rats with early cirrhosis. J. Hepatol. 1997, 26, 191–202. [Google Scholar] [CrossRef]
- Castilla-Cortazar, I.; Prieto, J.; Urdaneta, E.; Pascual, M.; Nunez, M.; Zudaire, E.; Garcia, M.; Quiroga, J.; Santidrian, S. Impaired intestinal sugar transport in cirrhotic rats: Correction by low doses of insulin-like growth factor I. Gastroenterology 1997, 113, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Castilla-Cortazar, I.; Picardi, A.; Tosar, A.; Ainzua, J.; Urdaneta, E.; Garcia, M.; Pascual, M.; Quiroga, J.; Prieto, J. Effect of insulin-like growth factor I on in vivo intestinal absorption of d-galactose in cirrhotic rats. Am. J. Physiol. 1999, 276, G37–G42. [Google Scholar] [CrossRef] [PubMed]
- Cemborain, A.; Castilla-Cortazar, I.; Garcia, M.; Quiroga, J.; Muguerza, B.; Picardi, A.; Santidrian, S.; Prieto, J. Osteopenia in rats with liver cirrhosis: Beneficial effects of IGF-I treatment. J. Hepatol. 1998, 28, 122–131. [Google Scholar] [CrossRef]
- Castilla-Cortazar, I.; Garcia, M.; Quiroga, J.; Diez, N.; Diez-Caballero, F.; Calvo, A.; Diaz, M.; Prieto, J. Insulin-like growth factor-I reverts testicular atrophy in rats with advanced cirrhosis. Hepatology 2000, 31, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Castilla-Cortazar, I.; Diez, N.; Garcia-Fernandez, M.; Puche, J.E.; Diez-Caballero, F.; Quiroga, J.; Diaz-Sanchez, M.; Castilla, A.; Casares, A.D.; Varela-Nieto, I.; et al. Hematotesticular barrier is altered from early stages of liver cirrhosis: Effect of insulin-like growth factor 1. World J. Gastroenterol. 2004, 10, 2529–2534. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, M.; Delgado, G.; Puche, J.E.; González-Barón, S.; Castilla Cortázar, I. Low doses of insulin-like growth factor I improve insulin resistance, lipid metabolism, and oxidative damage in aging rats. Endocrinology 2008, 149, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Castilla-Cortazar, I.; Garcia, M.; Muguerza, B.; Quiroga, J.; Perez, R.; Santidrian, S.; Prieto, J. Hepatoprotective effects of insulin-like growth factor I in rats with carbon tetrachloride-induced cirrhosis. Gastroenterology 1997, 113, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Zuniga, V.; Rodriquez-Ortigosa, C.M.; Bartoli, R.; Martinez-Chantar, M.L.; Martinez-Peralta, L.; Pardo, A.; Ojanguren, I.; Quiroga, J.; Planas, R.; Prieto, J. Insulin-like growth factor I improves intestinal barrier function in cirrhotic rats. Gut 2000, 55, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Muguerza, B.; Castilla-Cortazar, I.; Garcia, M.; Quiroga, J.; Santidrian, S.; Prieto, J. Antifibrogenic effect in vivo of low doses of insulin-like growth factor-I in cirrhotic rats. Biochim. Biophys. Acta 2001, 1536, 185–195. [Google Scholar] [CrossRef]
- Sobrevals, L.; Rodriguez, C.; Romero-Trevejo, J.L.; Gondi, G.; Monreal, I.; Pañeda, A.; Juanarena, N.; Arcelus, S.; Razquin, N.; Guembe, L.; et al. Insulin-like growth factor I gene transfer to cirrhotic liver induces fibrolysis and reduces fibrogenesis leading to cirrhosis reversion in rats. Hepatology 2010, 51, 912–921. [Google Scholar] [PubMed]
- Zhang, Y.; Zhang, Q.Q.; Guo, X.H.; Zhang, H.Y.; Liu, L.X. IGFBPrP1 induces liver fibrosis by inducing hepatic stellate cell activation and hepatocyte apoptosis via Smad2/3 signaling. World J. Gastroenterol. 2014, 20, 6523–6533. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Kondo, M.; Takeda, S.; Osada, H.; Takahashi, T.; Nakao, A.; Takahashi, T. Altered transcriptional regulation of the insulin-like growth factor 2 gene in human hepatocellular carcinoma. Mol. Carcinog. 1997, 18, 193–198. [Google Scholar] [CrossRef]
- Wang, Z.; Ruan, Y.B.; Guan, Y.; Liu, S.H. Expression of IGF-II in early experimental hepatocellular carcinomas and its significance in early diagnosis. World J. Gastroenterol. 2003, 9, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, B.; Ghosh, S.; Das, T.; Doloi, M. Characterization of insulin-like-growth factor II (IGF II) mRNA positive hepatic altered foci and IGF II expression in hepatocellular carcinoma during diethylnitrosamine-induced hepatocarcinogenesis in rats. J. Carcinog. 2005, 4, 12–25. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Qiu, L.W.; Yao, D.F.; Zong, L.; Lu, Y.Y.; Huang, H.; Wu, W.; Wu, X.H. Abnormal expression of insulin-like growth factor-II and its dynamic quantitative analysis at different stages of hepatocellular carcinoma development. Hepatobiliary Pancreat. Dis. Int. 2008, 7, 406–411. [Google Scholar] [PubMed]
- Scharf, J.G.; Ramadori, G.; Dombrowski, F. Analysis of the IGF axis in preneoplastic hepatic foci and hepatocellular neoplasms developing after low-number pancreatic islet transplantation into the livers of streptozotocin diabetic rats. Lab. Investig. 2000, 80, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Aleem, E.; Nehrbass, D.; Klimek, F.; Mayer, D.; Bannasch, P. Upregulation of the insulin receptor and type I insulin-like growth factor receptor are early events in hepatocarcinogenesis. Toxicol. Pathol. 2011, 39, 524–543. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.D.; Yao, M.; Wang, L.; Zhang, H.J.; Yan, M.J.; Gu, X.; Shi, Y.; Chen, J.; Dong, Z.Z.; Yao, D.F. Overexpression of insulin-like growth factor-I receptor as a pertinent biomarker for hepatocytes malignant transformation. World J. Gastroenterol. 2013, 19, 6084–6092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Z.X.; Fang, Q.; Gao, F.Y.; Zhao, Q.L.; Yang, Y.; Su, H.M.; Liu, Y.K. Expression and significance of insulin-like growth factor-1 and insulin-like growth factor binding protein-3 in hepatocyte steatosis model. Zhonghua Gan Zang Bing Za Zhi 2012, 20, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Svegliati-Baroni, G.; Ridolfi, F.; Di Sario, A.; Casini, A.; Marucci, L.; Gaggiotti, G.; Orlandoni, P.; Macarri, G.; Perego, L.; Benedetti, A.; et al. Insulin and insulin-like growth factor-1 stimulate proliferation and type I collagen accumulation by human hepatic stellate cells: Differential effects on signal transduction pathways. Hepatology 1999, 29, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.N.; Geng, Y.J.; Li, F.; Yang, T.; Su, D.F.; Duan, J.L.; Li, Y. Insulin-like growth factor-1 receptor activation prevents hydrogen peroxide-induced oxidative stress, mitochondrial dysfunction and apoptosis. Apoptosis 2011, 16, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Tapia, N.C.; Rosso, N.; Tiribelli, C. Effect of intracellular lipid accumulation in a new model of non-alcoholic fatty liver disease. BMC Gastroenterol. 2012, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.E.; Friedman, S.L. Mechanisms of hepatic fibrogenesis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Kawada, N. Inflammation and fibrogenesis in steatohepatitis. J. Gastroenterol. 2012, 47, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Scharf, J.G.; Knittel, T.; Dombrowski, F.; Müller, L.; Saile, B.; Braulke, T.; Hartmann, H.; Ramadori, G. Characterization of the IGF axis components in isolated rat hepatic stellate cells. Hepatology 1998, 27, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Brenzel, A.; Gressner, A.M. Characterization of insulin-like growth factor (IGF)-I-receptor binding sites during in vitro transformation of rat hepatic stellate cells to myofibroblasts. Eur. J. Clin. Chem. Clin. Biochem. 1996, 34, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Skrtic, S.; Wallenius, V.; Ekberg, S.; Brenzel, A.; Gressner, A.M.; Jansson, J.O. Insulin-like growth factors stimulate expression of hepatocyte growth factor but not transforming growth factor beta1 in cultured hepatic stellate cells. Endocrinology 1997, 138, 4683–4689. [Google Scholar] [CrossRef] [PubMed]
- Desbois-Mouthon, C.; Wendum, D.; Cadoret, A.; Rey, C.; Leneuve, P.; Blaise, A.; Housset, C.; Tronche, F.; Le Bouc, Y.; Holzenberger, M. Hepatocyte proliferation during liver regeneration is impaired in mice with liver-specific IGF-1R knockout. FASEB J. 2006, 20, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Fausto, N.; Laird, A.D.; Webber, E.M. Liver regeneration. 2. Role of growth factors and cytokines in hepatic regeneration. FASEB J. 1995, 9, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Cressman, D.E.; Greenbaum, L.E.; DeAngelis, R.A.; Ciliberto, G.; Furth, E.E.; Poli, V.; Taub, R. Liver failure and defective hepatocyte regeneration in interleukin-6-deficient mice. Science 1996, 22, 1379–1383. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, B.Z.; Guan, L.Y.; Jiang, T. Hepatic insulin-like growth factor receptor is upregulated by activation of the GSK3B-FOXO3 pathway after partial hepatectomy. J. Biol. Regul. Homeost. Agents 2017, 31, 549–555. [Google Scholar]
- Godwin, J.W.; Pinto, A.R.; Rosenthal, N.A. Chasing the recipe for a pro-regenerative immune system. Semin. Cell Dev. Biol. 2017, 61, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Vannella, K.M.; Wynn, T.A. Mechanisms of Organ Injury and Repair by Macrophages. Annu. Rev. Physiol. 2017, 79, 593–617. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.F.; Yauk, Y.K.; Chou, C.K.; Ting, L.P.; Chang, C.; Hu, C.P.; Han, S.H.; Su, T.S. Evidence of autocrine regulation in human hepatoma cell lines. Biochem. Biophys. Res. Commun. 1988, 153, 39–45. [Google Scholar] [CrossRef]
- Scharf, J.G.; Schmidt-Sandte, W.; Pahernik, S.A.; Ramadori, G.; Braulke, T.; Hartmann, H. Characterization of the insulin-like growth factor axis in a human hepatoma cell line (PLC). Carcinogenesis 1998, 19, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Ling, X. IGF-1 promotes the growth and metastasis of hepatocellular carcinoma via the inhibition of proteasome-mediated cathepsin B degradation. World J. Gastroenterol. 2015, 21, 10137–10149. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.H.; Lee, M.J.; Bae, S.K.; Lee, O.H.; Lee, Y.M.; Park, B.C.; Kim, K.W. Insulin-like growth factor II (IGF-II) secreted from HepG2 human hepatocellular carcinoma cells shows angiogenic activity. Cancer Lett. 1998, 128, 41–46. [Google Scholar] [CrossRef]
- Lahm, H.; Gittner, K.; Krebs, O.; Sprague, L.; Deml, E.; Oesterle, D.; Hoeflich, A.; Wanke, R.; Wolf, E. Diethylnitrosamine induces long-lasting re-expression of insulin-like growth factor II during early stages of liver carcinogenesis in mice. Growth Horm. IGF Res. 2002, 12, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Breuhahn, K.; Vreden, S.; Haddad, R.; Beckebaum, S.; Stippel, D.; Flemming, P.; Nussbaum, T.; Caselmann, W.H.; Haab, B.B.; Schirmacher, P. Molecular profiling of human hepatocellular carcinoma defines mutually exclusive interferon regulation and insulin-like growth factor II overexpression. Cancer Res. 2004, 64, 6058–6064. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.B.; Hsieh, S.H.; Hsu, H.L.; Lai, M.Y.; Kan, L.S.; Au, L.C. Antisense oligodeoxynucleotides of IGF-II selectively inhibit growth of human hepatoma cells overproducing IGF-II. J. Biochem. 1997, 122, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Tovar, V.; Alsinet, C.; Villanueva, A.; Hoshida, Y.; Chiang, D.Y.; Solé, M.; Thung, S.; Moyano, S.; Toffanin, S.; Mínguez, B.; et al. IGF activation in a molecular subclass of hepatocellular carcinoma and pre-clinical efficacy of IGF-1R blockage. J. Hepatol. 2010, 52, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Desbois-Mouthon, C.; Baron, A.; Blivet-Van Eggelpoël, M.J.; Fartoux, L.; Venot, C.; Bladt, F.; Housset, C.; Rosmorduc, O. Insulin-like growth factor-1 receptor inhibition induces a resistance mechanism via the epidermal growth factor receptor/HER3/AKT signaling pathway: Rational basis for cotargeting insulin-like growth factor-1 receptor and epidermal growth factor receptor in hepatocellular carcinoma. Clin. Cancer Res. 2009, 15, 5445–5456. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Tatebe, H.; Nakagawa, T.; Hara, Y.; Weinstein, I.B.; Moriwaki, H. EGCG inhibits activation of the insulin-like growth factor (IGF)/IGF-1 receptor axis in human hepatocellular carcinoma cells. Cancer Lett. 2008, 262, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Höpfner, M.; Huether, A.; Sutter, A.P.; Baradari, V.; Schuppan, D.; Scherübl, H. Blockade of IGF-1 receptor tyrosine kinase has antineoplastic effects in hepatocellular carcinoma cells. Biochem. Pharmacol. 2006, 71, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Ruan, B.; Wu, J.; Wang, J.; Shang, R.; Sun, W.; Li, X.; Dou, K.; Wang, K.; Li, Y. Insulin-like growth factor binding protein-1 inhibits cancer cell invasion and is associated with poor prognosis in hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 5645–5654. [Google Scholar] [PubMed]
- Kondoh, N.; Wakatsuki, T.; Ryo, A.; Hada, A.; Aihara, T.; Horiuchi, S.; Goseki, N.; Matsubara, O.; Takenaka, K.; Shichita, M.; et al. Identification and characterization of genes associated with human hepatocellular carcinogenesis. Cancer Res. 1999, 59, 4990–4996. [Google Scholar] [PubMed]
- LeRoith, D. Insulin-like growth factor I receptor signaling–overlapping or redundant pathways. Endocrinology 2000, 141, 1287–1288. [Google Scholar] [CrossRef] [PubMed]
- Coutant, A.; Rescan, C.; Gilot, D.; Loyer, P.; Guguen-Guillouzo, C.; Baffet, G. PI3K-FRAP/mTOR pathway is critical for hepatocyte proliferation whereas MEK/ERK supports both proliferation and survival. Hepatology 2000, 36, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Alexia, C.; Fallot, G.; Lasfer, M.; Schweizer-Groyer, G.; Groyer, A. An evaluation of the role of insulin-like growth factors (IGF) and of type-I IGF receptor signaling in hepatocarcinogenesis and in the resistance of hepatocellular cells against drug-induced apoptosis. Biochem. Pharmacol. 2004, 86, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Dal, K.; Bulur, O.; Ata, N.; Yeniova, A.O.; Baser, S.; Karakaya, S.; Unsal, O.; Dagdeviren, M.; Karadag, I.; Beyan, E.; et al. The role of insulin—Like growth factor—1 on steatohepatitis. Acta Gastroenterol. Belg. 2017, 80, 21–24. [Google Scholar] [PubMed]
- Arturi, F.; Succurro, E.; Procopio, C.; Pedace, E.; Mannino, G.C.; Lugarà, M.; Procopio, T.; Andreozzi, F.; Sciacqua, A.; Hribal, M.L.; et al. Nonalcoholic fatty liver disease is associated with low circulating levels of insulin-like growth factor-I. J. Clin. Endocrinol. Metab. 2011, 96, E1640–E1644. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Yonei, Y.; Tanaka, S.; Mori, K.; Kanemasa, K.; Imai, S.; Taketani, H.; Hara, T.; Seko, Y.; Ishiba, H.; et al. Lower levels of insulin-like growth factor-1 standard deviation score are associated with histological severity of non-alcoholic fatty liver disease. Hepatol. Res. 2015, 45, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, R.; Koga, M.; Kasayama, S.; Fukuoka, H.; Iguchi, G.; Odake, Y.; Yoshida, K.; Bando, H.; Suda, K.; Nishizawa, H.; et al. Factors correlated with serum insulin-like growth factor-I levels in health check-up subjects. Growth Horm. IGF Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Cheng, X.; Hu, Y.; Song, R.; Li, G. Insulin-like growth factor 1 and metabolic parameters are associated with nonalcoholic fatty liver disease in obese children and adolescents. Acta Paediatr. 2017, 106, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Yu, Z.; Song, X.; Wang, Y.; Li, M.; Xue, J. Reduced growth hormone secretion is associated with nonalcoholic fatty liver disease in obese children. Horm. Metab. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Colak, Y.; Senates, E.; Ozturk, O.; Yilmaz, Y.; Zemheri, E.; Yilmaz Enc, F.; Ulasoglu, C.; Aksaray, S.; Bozbeyoglu, S.G.; Kiziltas, S.; et al. Serum concentrations of human insulin-like growth factor-1 and levels of insulin-like growth factor-binding protein-5 in patients with nonalcoholic fatty liver disease: Association with liver histology. Eur. J. Gastroenterol. Hepatol. 2012, 24, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Dichtel, L.E.; Corey, K.E.; Misdraji, J.; Bredella, M.A.; Schorr, M.; Osganian, S.A.; Young, B.J.; Sung, J.C.; Miller, K.K. The association between IGF-1 levels and the histologic severity of nonalcoholic fatty liver disease. Clin. Transl. Gastroenterol. 2017, 8, e217. [Google Scholar] [CrossRef] [PubMed]
- Hagström, H.; Stal, P.; Hultcrantz, R.; Brismar, K.; Ansurudeen, I. IGFBP-1 and IGF-I as markers for advanced fibrosis in NAFLD—A pilot study. Scand. J. Gastroenterol. 2017, 52, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Cianfarani, S.; Inzaghi, E.; Alisi, A.; Germani, D.; Puglianiello, A.; Nobili, V. Insulin-like growth factor-I and -II levels are associated with the progression of nonalcoholic fatty liver disease in obese children. J. Pediatr. 2014, 165, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Chishima, S.; Kogiso, T.; Matsushita, N.; Hashimoto, E.; Tokushige, K. The Relationship between the growth hormone/insulin-like growth factor system and the histological features of nonalcoholic fatty liver disease. Intern. Med. 2017, 56, 473–480. [Google Scholar] [CrossRef] [PubMed]
- García-Galiano, D.; Sánchez-Garrido, M.A.; Espejo, I.; Montero, J.L.; Costán, G.; Marchal, T.; Membrives, A.; Gallardo-Valverde, J.M.; Muñoz-Castañeda, J.R.; Arévalo, E.; et al. IL-6 and IGF-1 are independent prognostic factors of liver steatosis and non-alcoholic steatohepatitis in morbidly obese patients. Obes. Surg. 2007, 17, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Mallea-Gil, M.S.; Ballarino, M.C.; Spiraquis, A.; Iriarte, M.; Kura, M.; Gimenez, S.; Oneto, A.; Guitelman, M.; Machado, R.; Miguel, C.M. IGF-1 levels in different stages of liver steatosis and its association with metabolic syndrome. Acta Gastroenterol. Latinoam. 2012, 42, 20–26. [Google Scholar] [PubMed]
- Fusco, A.; Miele, L.; D’Uonnolo, A.; Forgione, A.; Riccardi, L.; Cefalo, C.; Barini, A.; Bianchi, A.; Giampietro, A.; Cimino, V.; et al. Nonalcoholic fatty liver disease is associated with increased GHBP and reduced GH/IGF-I levels. Clin. Endocrinol. 2012, 77, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Petäjä, E.M.; Zhou, Y.; Havana, M.; Hakkarainen, A.; Lundbom, N.; Ihalainen, J.; Yki-Järvinen, H. Phosphorylated IGFBP-1 as a non-invasive predictor of liver fat in NAFLD. Sci. Rep. 2016, 6, 24740. [Google Scholar] [CrossRef] [PubMed]
- Völzke, H.; Nauck, M.; Rettig, R.; Dörr, M.; Higham, C.; Brabant, G.; Wallaschofski, H. Association between hepatic steatosis and serum IGF1 and IGFBP-3 levels in a population-based sample. Eur. J. Endocrinol. 2009, 161, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Runchey, S.S.; Boyko, E.J.; Ioannou, G.N.; Utzschneider, K.M. Relationship between serum circulating insulin-like growth factor-1 and liver fat in the United States. J. Gastroenterol. Hepatol. 2014, 29, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zheng, X.M.; Jiang, F.; Qiu, W.Q. MicroRNA-190b regulates lipid metabolism and insulin sensitivity by targeting IGF-1 and ADAMTS9 in non-alcoholic fatty liver disease. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Vyzantiadis, T.; Theodoridou, S.; Giouleme, O.; Harsoulis, P.; Evgenidis, N.; Vyzantiadis, A. Serum concentrations of insulin-like growth factor-I (IGF-I) in patients with liver cirrhosis. Hepatogastroenterology 2003, 50, 814–816. [Google Scholar] [PubMed]
- Wu, Y.L.; Ye, J.; Zhang, S.; Zhong, J.; Xi, R.P. Clinical significance of serum IGF-I, IGF-II and IGFBP-3 in liver cirrhosis. World J. Gastroenterol. 2004, 10, 2740–2743. [Google Scholar] [CrossRef] [PubMed]
- Colakoğlu, O.; Taşkiran, B.; Colakoğlu, G.; Kizildağ, S.; Ari Ozcan, F.; Unsal, B. Serum insulin like growth factor-1 (IGF-1) and insulin like growth factor binding protein-3 (IGFBP-3) levels in liver cirrhosis. Turk. J. Gastroenterol. 2007, 18, 245–249. [Google Scholar] [PubMed]
- Raslan, H.M.; Ezzat, W.M.; Ahmed, M.M.; Rasheed, E.A. Insulin growth factor-1 and insulin growth factor binding protein-3 in Egyptian patients with chronic hepatitis C. Arch. Med. Sci. 2007, 3, 46–51. [Google Scholar]
- Raslan, H.M.; Elhosary, Y.; Ezzat, W.M.; Rasheed, E.A.; Rasheed, M.A. The potential role of insulin-like growth factor 1, insulin-like growth factor binding protein 3 and bone mineral density in patients with chronic hepatitis C virus in Cairo, Egypt. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Jeyaratnaganthan, N.; Grønbaek, H.; Holland-Fischer, P.; Espelund, U.; Chen, J.W.; Flyvbjerg, A.; Vilstrup, H.; Frystyk, J. Ascites from patients with alcoholic liver cirrhosis contains higher IGF-I bioactivity than serum. Clin. Endocrinol. 2010, 72, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Adamek, A.; Kasprzak, A.; Seraszek, A.; Mikoś, H.; Bura, A.; Mozer-Lisewska, I. Alterations of insulin-like growth factor I (IGF-I) and estradiol serum levels in chronic hepatitis C. Contemp. Oncol. 2012, 16, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Aleem, E.; Elshayeb, A.; Elhabachi, N.; Mansour, A.R.; Gowily, A.; Hela, A. Serum IGFBP-3 is a more effective predictor than IGF-1 and IGF-2 for the development of hepatocellular carcinoma in patients with chronic HCV infection. Oncol. Lett. 2012, 3, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Lupoli, R.; Di Minno, A.; Spadarella, G.; Ambrosino, P.; Panico, A.; Tarantino, L.; Lupoli, G.; Lupoli, G.; Di Minno, M.N. The risk of osteoporosis in patients with liver cirrhosis: A meta-analysis of literature studies. Clin. Endocrinol. 2016, 84, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, S.M.; Karamifar, H.; Hamzavi, S.S.; Haghighat, M.; Malek-Hosseini, S.A. Serum insulinlike growth factor-1 and its binding protein-3 levels in children with cirrhosis waiting for a liver transplant. Exp. Clin. Transplant. 2012, 10, 252–257. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brabant, G.; Wallaschofski, H. Normal levels of serum IGF-I: Determinants and validity of current reference ranges. Pituitary 2007, 10, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Ronsoni, M.F.; Lazzarotto, C.; Fayad, L.; Silva, M.C.; Nogueira, C.L.; Bazzo, M.L.; Narciso-Schiavon, J.L.; Dantas-Corrêa, E.B.; Schiavon Lde, L. IGF-I and IGFBP-3 serum levels in patients hospitalized for complications of liver cirrhosis. Ann. Hepatol. 2013, 12, 456–463. [Google Scholar] [PubMed]
- Khoshnood, A.; Nasiri Toosi, M.; Faravash, M.J.; Esteghamati, A.; Froutan, H.; Ghofrani, H.; Kalani, M.; Miroliaee, A.; Abdollahi, A.; Yasir, A. A survey of correlation between insulin-like growth factor-I (igf-I) levels and severity of liver cirrhosis. Hepat. Mon. 2013, 13, e6181. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.; Astemborski, J.; Mehta, S.H.; Kirk, G.D.; Thomas, D.L.; Balagopal, A. HIV/HCV Co-infection, Liver Disease Progression, and Age-Related IGF-1 Decline. Pathog. Immun. 2017, 2, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Castro, G.R.; Coelho, J.C.; Parolin, M.B.; Matias, J.E.; de Freitas, A.C. Insulin-like growth factor I correlates with MELD and returns to normal level after liver transplantation. Ann. Transplant. 2013, 18, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Colombo, B.D.; Ronsoni, M.F.; Soares E Silva, P.E.; Fayad, L.; Wildner, L.M.; Bazzo, M.L.; Dantas-Correa, E.B.; Narciso-Schiavon, J.L.; Schiavon, L.L. Prognostic significance of insulin-like growth factor-I serum levels in acute decompensation of cirrhosis. Biomarkers 2017, 22, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, G.; Sorvillo, F.; Morisco, F.; Carbone, A.; Rotondi, M.; Stornaiuolo, G.; Precone, D.F.; Cioffi, M.; Gaeta, G.B.; Caporaso, N.; et al. Serum insulin-like growth factor I evaluation as a useful tool for predicting the risk of developing hepatocellular carcinoma in patients with hepatitis C virus-related cirrhosis: A prospective study. Cancer 2002, 95, 2539–2545. [Google Scholar] [CrossRef] [PubMed]
- Salso, A.; Tisone, G.; Tariciotti, L.; Lenci, I.; Manzia, T.M.; Baiocchi, L. Relationship between GH/IGF-1 axis, graft recovery, and early survival in patients undergoing liver transplantation. Biomed. Res. Int. 2014, 2014, 240873. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.M.; Auernhammer, C.J.; Lee, P.D.; Engelhardt, D.; Zachoval, R. Insulin-like growth factors and insulin-like growth factor binding proteins in adult patients with severe liver disease before and after orthotopic liver transplantation. Horm. Res. 2002, 57, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Rehem, R.N.; El-Shikh, W.M. Serum IGF-1, IGF-2 and IGFBP-3 as parameters in the assessment of liver dysfunction in patients with hepatic cirrhosis and in the diagnosis of hepatocellular carcinoma. Hepatogastroenterology 2011, 58, 949–954. [Google Scholar] [PubMed]
- Correa, C.G.; Colombo Bda, S.; Ronsoni, M.F.; e Silva, P.E.S.; Fayad, L.; Silva, T.E.; Wildner, L.M.; Bazzo, M.L.; Dantas-Correa, E.B.; Narciso-Schiavon, J.L.; et al. Circulating insulin-like growth factor-binding protein 3 as prognostic biomarker in liver cirrhosis. World J. Hepatol. 2016, 8, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Donaghy, A.J.; Delhanty, P.J.; Ho, K.K.; Williams, R.; Baxter, R.C. Regulation of the growth hormone receptor/binding protein, insulin-like growth factor ternary complex system in human cirrhosis. J. Hepatol. 2002, 36, 751–758. [Google Scholar] [CrossRef]
- Sedlaczek, N.; Hasilik, A.; Neuhaus, P.; Schuppan, D.; Herbst, H. Focal overexpression of insulin-like growth factor 2 by hepatocytes and cholangiocytes in viral liver cirrhosis. Br. J. Cancer 2003, 88, 733–739. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Seo, J.H.; Park, B.C. Expression of insulin-like growth factor II in chronic hepatitis B, liver cirrhosis, and hepatocellular carcinoma. Gan Kagaku Ryoho 1995, 22, 292–307. [Google Scholar]
- Cariani, E.; Lasserre, C.; Seurin, D.; Hamelin, B.; Kemeny, F.; Franco, D.; Czech, M.P.; Ullrich, A.; Brechot, C. Differential expression of insulin-like growth factor II mRNA in human primary liver cancers, benign liver tumors, and liver cirrhosis. Cancer Res. 1988, 48, 6844–6849. [Google Scholar] [PubMed]
- Guo, X.H.; Liu, L.X.; Zhang, H.Y.; Zhang, Q.Q.; Li, Y.; Tian, X.X.; Qiu, Z.H. Insulin-like growth factor binding protein-related protein 1 contributes to hepatic fibrogenesis. J. Dig. Dis. 2014, 15, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Espelund, U.; Grønbæk, H.; Villadsen, G.E.; Simonsen, K.; Vestergaard, P.F.; Jørgensen, J.O.; Flyvbjerg, A.; Vilstrup, H.; Frystyk, J. The Circulating IGF System in Hepatocellular Carcinoma: The Impact of Liver Status and Treatment. Growth Horm. IGF Res. 2015, 25, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.C.; Deng, M.; Jiang, H.Y.; Guo, L.H.; Zhou, W.J.; Ruan, B. Serum insulin-like growth factor-1 and its binding protein 3 as prognostic factors for the incidence, progression, and outcome of hepatocellular carcinoma: A systematic review and meta-analysis. Oncotarget 2017, 8, 81098–81108. [Google Scholar] [CrossRef] [PubMed]
- Nedic, O.; Malenkovic, V.; Dukanovic, B.; Baricevic, I. Association of elevated IGFBP-1 with increased IGF-II concentration in patients with carcinoma of the liver. Int. J. Biol. Markers 2008, 23, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Stuver, S.O.; Kuper, H.; Tzonou, A.; Lagiou, P.; Spanos, E.; Hsieh, C.C.; Mantzoros, C.; Trichopoulos, D. Insulin-like growth factor 1 in hepatocellular carcinoma and metastatic liver cancer in men. Int. J. Cancer 2000, 87, 118–121. [Google Scholar] [CrossRef]
- Su, W.W.; Lee, K.T.; Yeh, Y.T.; Soon, M.S.; Wang, C.L.; Yu, M.L.; Wang, S.N. Association of circulating insulin-like growth factor 1 with hepatocellular carcinoma: One cross-sectional correlation study. J. Clin. Lab. Anal. 2010, 24, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Kaseb, A.O.; Morris, J.S.; Hassan, M.M.; Siddiqui, A.M.; Lin, E.; Xiao, L.; Abdalla, E.K.; Vauthey, J.N.; Aloia, T.A.; Krishnan, S.; et al. Clinical and prognostic implications of plasma insulin-like growth factor-1 and vascular endothelial growth factor in patients with hepatocellular carcinoma. J. Clin. Oncol. 2011, 29, 3892–3899. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.Y.; Huang, C.C.; Lin, S.D.; Hsu, C.H.; Cheng, A.L. Serum insulin-like growth factor-1 levels predict outcomes of patients with advanced hepatocellular carcinoma receiving antiangiogenic therapy. Clin. Cancer Res. 2012, 18, 3992–3997. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, R.; Shehata, S.; Hassan, M.M.; Xiao, L.; Lee, J.S.; Cheung, S.; Essa, H.H.; Hassabo, H.M.; Shalaby, A.S.; Mosad, E.; et al. Validation of an IGF-CTP scoring system for assessing hepatic reserve in Egyptian patients with hepatocellular carcinoma. Oncotarget 2015, 6, 21193–21207. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elmashad, N.; Ibrahim, W.S.; Mayah, W.W.; Farouk, M.; Ali, L.A.; Taha, A.; Elmashad, W. Predictive value of serum insulin-like growth factor-1 in hepatocellular carcinoma. Asian Pac. J. Cancer Prev. 2015, 16, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, J.H.; Jung, Y.J.; Gim, J.; Kim, W.; Kim, B.G.; Lee, K.L.; Cho, Y.; Yoo, J.J.; Lee, M.; et al. Validation of a Modified Child-Turcotte-Pugh Classification System Utilizing Insulin-Like Growth Factor-1 for Patients with Hepatocellular Carcinoma in an HBV Endemic Area. PLoS ONE 2017, 12, e0170394. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Lee, J.H.; Yoo, J.J.; Choi, W.M.; Lee, M.J.; Cho, Y.; Lee, D.H.; Lee, Y.B.; Kwon, J.H.; Yu, S.J.; et al. Serum insulin-like growth factor-I level is an independent predictor of recurrence and survival in early hepatocellular carcinoma: A prospective cohort study. Clin. Cancer Res. 2013, 19, 4218–4227. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Kim, H.C.; Lee, J.H.; Yoo, J.-J.; Choi, W.M.; Cho, Y.Y.; Lee, M.J.; Cho, Y.; Lee, D.H.; Lee, Y.B.; et al. Serum insulin-like growth factor-1 predicts disease progression and survival in patients with hepatocellular carcinoma who undergo transarterial chemoembolization. PLoS ONE 2014, 9, e90862. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, Y.; Jiang, X. Prognostic significance of serum insulin-like growth factor-1 in patients with hepatocellular carcinoma following transarterial chemoembolization. Exp. Ther. Med. 2016, 11, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Mao, W.; Dong, M.; Yang, D.; Li, W.; Chen, Y. Serum Insulin-Like Growth Factor-1 (IGF-1): A Novel Prognostic Factor for Early Recurrence of Hepatocellular Carcinoma (HCC). Clin. Lab. 2017, 63, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.Z.; Yao, D.F.; Yao, D.B.; Wu, X.H.; Wu, W.; Qiu, L.W.; Jiang, D.R.; Zhu, J.H.; Meng, X.Y. Expression and alteration of insulin-like growth factor II-messenger RNA in hepatoma tissues and peripheral blood of patients with hepatocellular carcinoma. World J. Gastroenterol. 2005, 11, 4655–4660. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.F.; Jeng, J.E.; Chuang, L.Y.; You, H.L.; Wang, L.Y.; Hsieh, M.Y.; Chen, S.C.; Chuang, W.L.; Lin, Z.Y.; Yu, M.L.; et al. Serum insulin-like growth factor-II as a serologic marker of small hepatocellular carcinoma. Scand. J. Gastroenterol. 2005, 40, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Morace, C.; Cucunato, M.; Bellerone, R.; De Caro, G.; Crinò, S.; Fortiguerra, A.; Spadaro, F.; Zirilli, A.; Alibrandi, A.; Consolo, P.; et al. Insulin-like growth factor-II is a useful marker to detect hepatocellular carcinoma? Eur. J. Intern. Med. 2012, 23, e157–e161. [Google Scholar] [CrossRef] [PubMed]
- Mattera, D.; Capuano, G.; Colao, A.; Pivonello, R.; Manguso, F.; Puzziello, A.; D’Agostino, L. Increased IGF-I: IGFBP-3 ratio in patients with hepatocellular carcinoma. Clin. Endocrinol. 2003, 59, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Ranke, M.B.; Maier, K.P.; Schweizer, R.; Stadler, B.; Schleicher, S.; Elmlinger, M.W.; Flehmig, B. Pilot study of elevated levels of insulin-like growth factor-binding protein-2 as indicators of hepatocellular carcinoma. Horm. Res. 2003, 60, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Nojima, M.; Mori, M.; Matsunaga, Y.; Akutsu, N.; Sasaki, S.; Endo, T.; Kurozawa, Y.; Wakai, K.; Tamakoshi, A.; et al. Insulin-like growth factor-related components and the risk of liver cancer in a nested case-control study. Tumour Biol. 2016, 37, 15125–15132. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Fan, Y.C.; Gao, S.; Sun, F.K.; Yang, Y.; Wang, K. Methylation of serum insulin-like growth factor-binding protein 7 promoter in hepatitis B virus-associated hepatocellular carcinoma. Genes Chromosomes Cancer 2014, 53, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H.; Chow, P.K.; Ooi, L.L.; Soo, K.C. A possible role for insulin-like growth factor-binding protein-3 autocrine/paracrine loops in controlling hepatocellular carcinoma cell proliferation. Cell Growth Differ. 2002, 13, 115–122. [Google Scholar] [PubMed]
- Adamek, A.; Kasprzak, A.; Mikoś, H.; Przybyszewska, W.; Seraszek-Jaros, A.; Czajka, A.; Sterzyńska, K.; Mozer-Lisewska, I. The insulin-like growth factor-1 and expression of its binding protein-3 in chronic hepatitis C and hepatocellular carcinoma. Oncol. Rep. 2013, 30, 1337–1345. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ikeda, Y.; Kajiyama, K.; Yamashita, Y.; Ikegami, T.; Uchiyama, H.; Soejima, Y.; Kawanaka, H.; Ikeda, T.; Morita, M.; Oki, E.; et al. Differential expression of insulin-like growth factor 1 in human primary liver cancer. Fukuoka Igaku Zasshi 2013, 104, 334–338. [Google Scholar] [PubMed]
- Chun, Y.S.; Huang, M.; Rink, L.; Von Mehren, M. Expression levels of insulin-like growth factors and receptors in hepatocellular carcinoma: A retrospective study. World J. Surg. Oncol. 2014, 12, 231. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.M.; Tan, W.M.; Deng, W.X.; Zhuang, S.M.; Luo, J.W. Expression of albumin, IGF-1, IGFBP-3 in tumor tissues and adjacent non-tumor tissues of hepatocellular carcinoma patients with cirrhosis. World J. Gastroenterol. 2005, 11, 4272–4276. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Cui, L.; Minuk, G.Y. The expression of insulin-like growth factor binding proteins in human hepatocellular carcinoma. Mol. Cell. Biochem. 2000, 207, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yang, X.; Li, L.; Liu, P.; Wu, H.; Liu, Z.; Li, Q.; Liao, G.; Wang, X. Low expression levels of insulin-like growth factor binding protein-3 are correlated with poor prognosis for patients with hepatocellular carcinoma. Oncol. Lett. 2017, 13, 3395–3402. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yoo, B.K.; Santhekadur, P.K.; Gredler, R.; Bhutia, S.K.; Das, S.K.; Fuller, C.; Su, Z.Z.; Fisher, P.B.; Sarkar, D. Insulin-like growth factor-binding protein-7 functions as a potential tumor suppressor in hepatocellular carcinoma. Clin. Cancer Res. 2011, 17, 6693–6701. [Google Scholar] [CrossRef] [PubMed]
- Tomimaru, Y.; Eguchi, H.; Wada, H.; Kobayashi, S.; Marubashi, S.; Tanemura, M.; Umeshita, K.; Kim, T.; Wakasa, K.; Doki, Y.; et al. IGFBP7 downregulation is associated with tumor progression and clinical outcome in hepatocellular carcinoma. Int. J. Cancer 2012, 130, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Yao, M.; Wang, L.; Yan, X.; Gu, X.; Shi, Y.; Yao, N.; Qiu, L.; Wu, W.; Yao, D. Abnormal expression of insulin-like growth factor-I receptor in hepatoma tissue and its inhibition to promote apoptosis of tumor cells. Tumour Biol. 2013, 34, 3397–3405. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.H.; Lee, B.S.; Lee, E.S.; Kim, S.H.; Lee, H.Y.; Kang, D.Y. Prognostic significance of p53, mTOR, c-Met, IGF-1R, and HSP70 overexpression after the resection of hepatocellular carcinoma. Gut Liver 2014, 8, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Adamek, A.; Przybyszewska, W.; Pyda, P.; Szmeja, J.; Seraszek-Jaros, A.; Lanzafame, A.; Surdacka, A.; Mozer-Lisewska, I.; Koczorowska, M. Insulin-like growth factor-1 mRNA isoforms and insulin-like growth factor-1 receptor mRNA expression in chronic hepatitis C. World J. Gastroenterol. 2015, 21, 3867–3875. [Google Scholar] [CrossRef] [PubMed]
- Couvert, P.; Carrié, A.; Pariès, J.; Vaysse, J.; Miroglio, A.; Kerjean, A.; Nahon, P.; Chelly, J.; Trinchet, J.C.; Beaugrand, M.; et al. Liver insulin-like growth factor 2 methylation in hepatitis C virus cirrhosis and further occurrence of hepatocellular carcinoma. World J. Gastroenterol. 2008, 14, 5419–5427. [Google Scholar] [CrossRef]
- Couvert, P.; Carrié, A.; Tezenas du Montcel, S.; Vaysse, J.; Sutton, A.; Barget, N.; Trinchet, J.C.; Beaugrand, M.; Ganne, N.; Giral, P.; et al. Insulin-like growth factor 2 gene methylation in peripheral blood mononuclear cells of patients with hepatitis C related cirrhosis or hepatocellular carcinoma. Clin. Res. Hepatol. Gastroenterol. 2012, 36, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Hsieh, Y.H.; Tsai, C.M.; Chu, Y.H.; Ueng, K.C.; Liu, Y.F.; Yeh, Y.H.; Su, S.C.; Chen, Y.C.; Chen, M.K.; et al. Relationship of insulin-like growth factors system gene polymorphisms with the susceptibility and pathological development of hepatocellular carcinoma. Ann. Surg. Oncol. 2010, 17, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- Rashad, N.M.; El-Shal, A.S.; Abd Elbary, E.H.; Abo Warda, M.H.; Hegazy, O. Impact of insulin-like growth factor 2, insulin-like growth factor receptor 2, insulin receptor substrate 2 genes polymorphisms on susceptibility and clinicopathological features of hepatocellular carcinoma. Cytokine 2014, 68, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Chung, Y.H.; Kim, J.A.; Lee, D.; Jin, Y.J.; Shim, J.H.; Jang, M.K.; Cho, E.Y.; Shin, E.S.; Lee, J.E.; et al. Single nucleotide polymorphisms associated with metastatic tumour antigen 1 overexpression in patients with hepatocellular carcinoma. Liver Int. 2012, 32, 457–466. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene/Protein | NAFLD | Liver Cirrhosis | HCC | |
|---|---|---|---|---|
| IGF1 | S | ↓[97,100,101] ♣[37,98,102,103,104,106] | ↓[114,115,116,117,118,119,120,121,122] ♦[115,116,124,125,126,128] | ↓[133,140,142] ⇓[121,133,140,141] #[140,141,145,146,147,148,149,150,151,152] |
| T | ↓[65] ♣[32] | nd | ↓[161,162,163] | |
| IGF2 | S | ♣[105] | ↓[115] ♦[115,133] | ⇑, ↓[121,140,154,155,156] |
| T | nd | ↑[136] | ↑[138,154,164] | |
| IGF1R | S | nd | nd | nd |
| T | nd | nd | ↓[161,164] ↑[154,171] | |
| IGFBP1 | S | ↑[104] | ↑[119] | ↑[142] |
| T | nd | nd | ↓⇓[166] #[91] | |
| IGFBP2 | S | nd | ↑[119] | ↑[158] |
| T | nd | nd | nd | |
| IGFBP3 | S | ↑[106,109] | ↓[118,121,123,125,128] | ⇓[121,133,141,157] |
| T | ↓[65] | nd | ↓[161,165,166] ⇓[166] | |
| IGFBP4 | S | nd | nd | nd |
| T | nd | nd | ↓⇓[166] | |
| IGFBP5 | S | ♣[102] | nd | nd |
| T | nd | nd | nd | |
| IGFBP7 | S | nd | nd | ↑[160] |
| T | nd | ↓[139] | ↓ #[168,169] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. https://doi.org/10.3390/ijms19051308
Adamek A, Kasprzak A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. International Journal of Molecular Sciences. 2018; 19(5):1308. https://doi.org/10.3390/ijms19051308
Chicago/Turabian StyleAdamek, Agnieszka, and Aldona Kasprzak. 2018. "Insulin-Like Growth Factor (IGF) System in Liver Diseases" International Journal of Molecular Sciences 19, no. 5: 1308. https://doi.org/10.3390/ijms19051308
APA StyleAdamek, A., & Kasprzak, A. (2018). Insulin-Like Growth Factor (IGF) System in Liver Diseases. International Journal of Molecular Sciences, 19(5), 1308. https://doi.org/10.3390/ijms19051308