Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
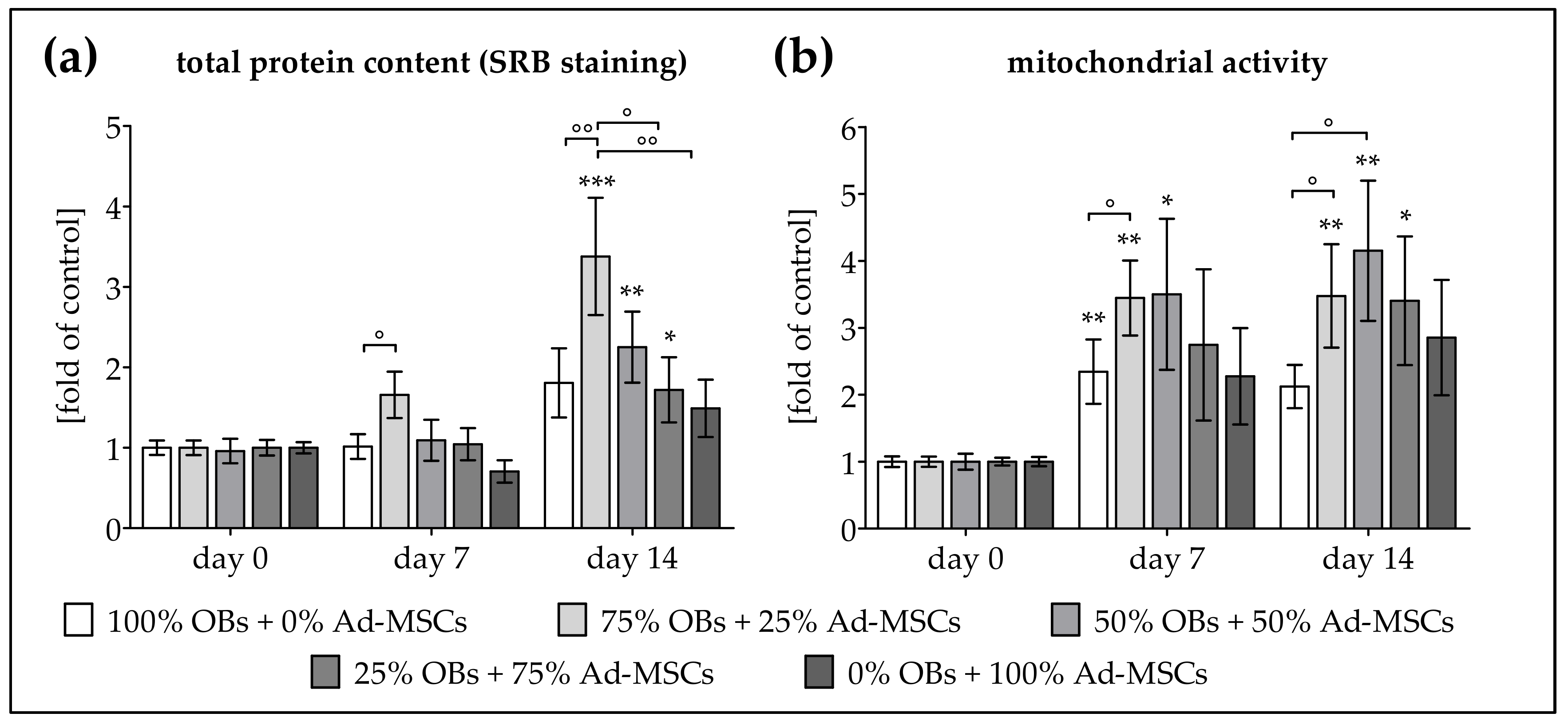
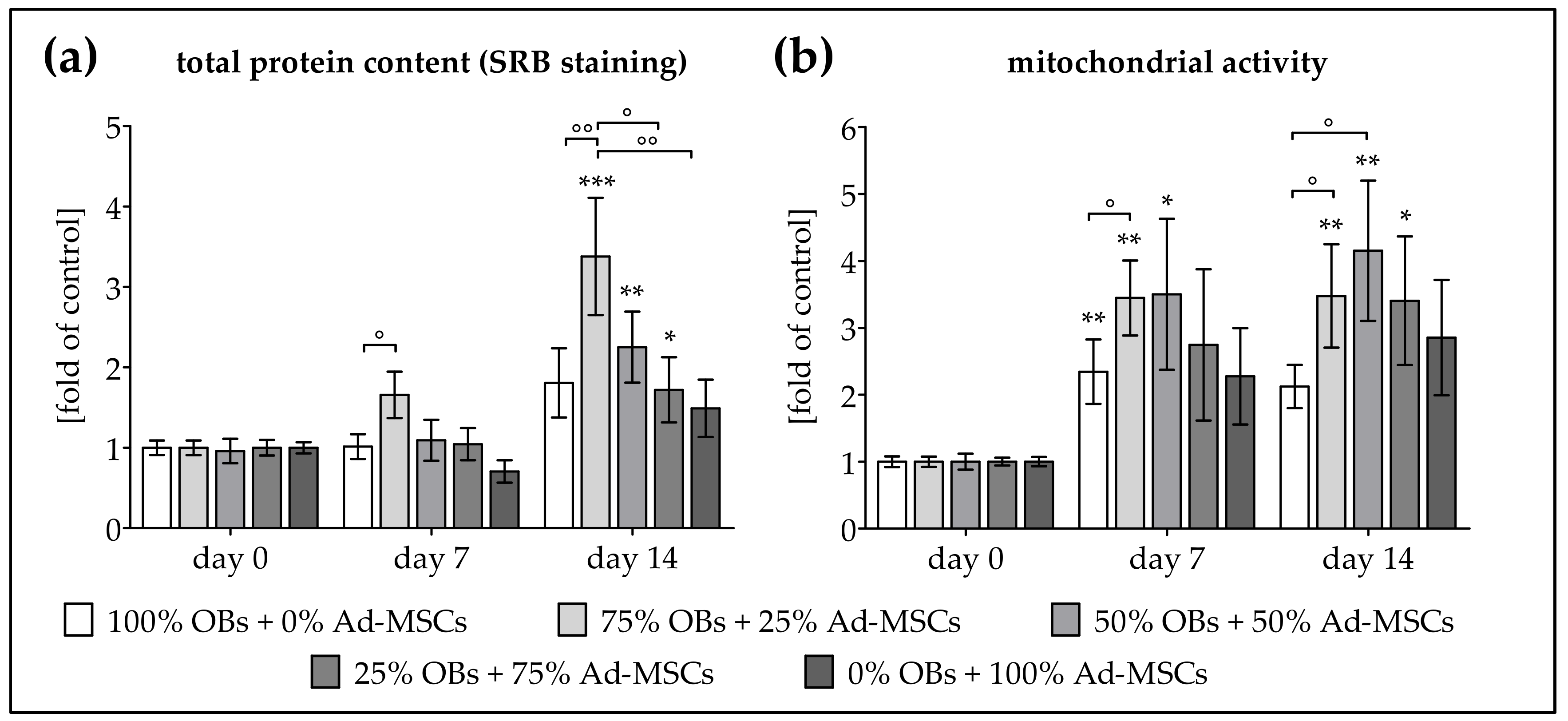
2.1. Co-Culture Improves Proliferation of OBs and Ad-MSCs
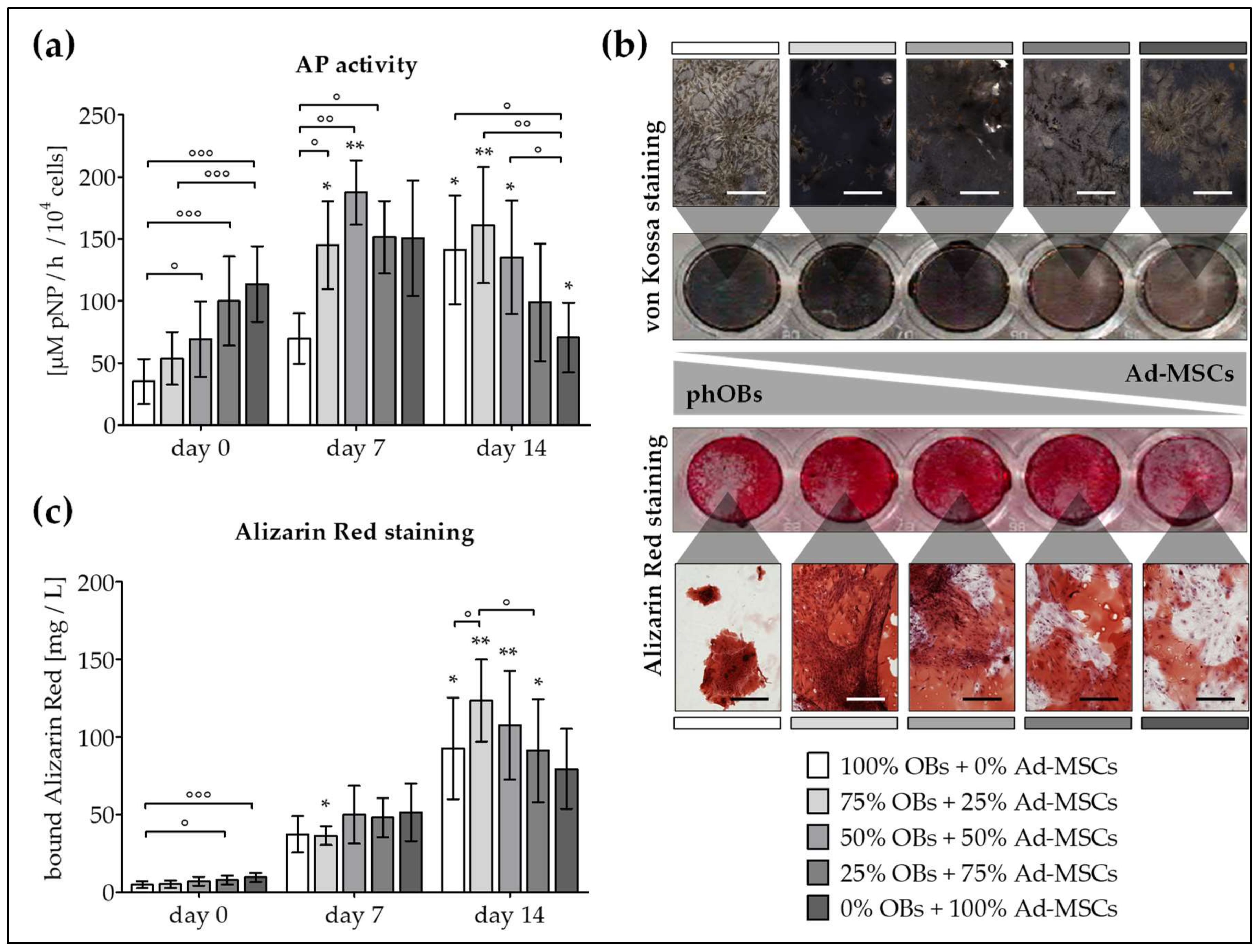
2.2. Co-Culture Improves AP Activity and Matrix Mineralization of OBs and Ad-MSCs
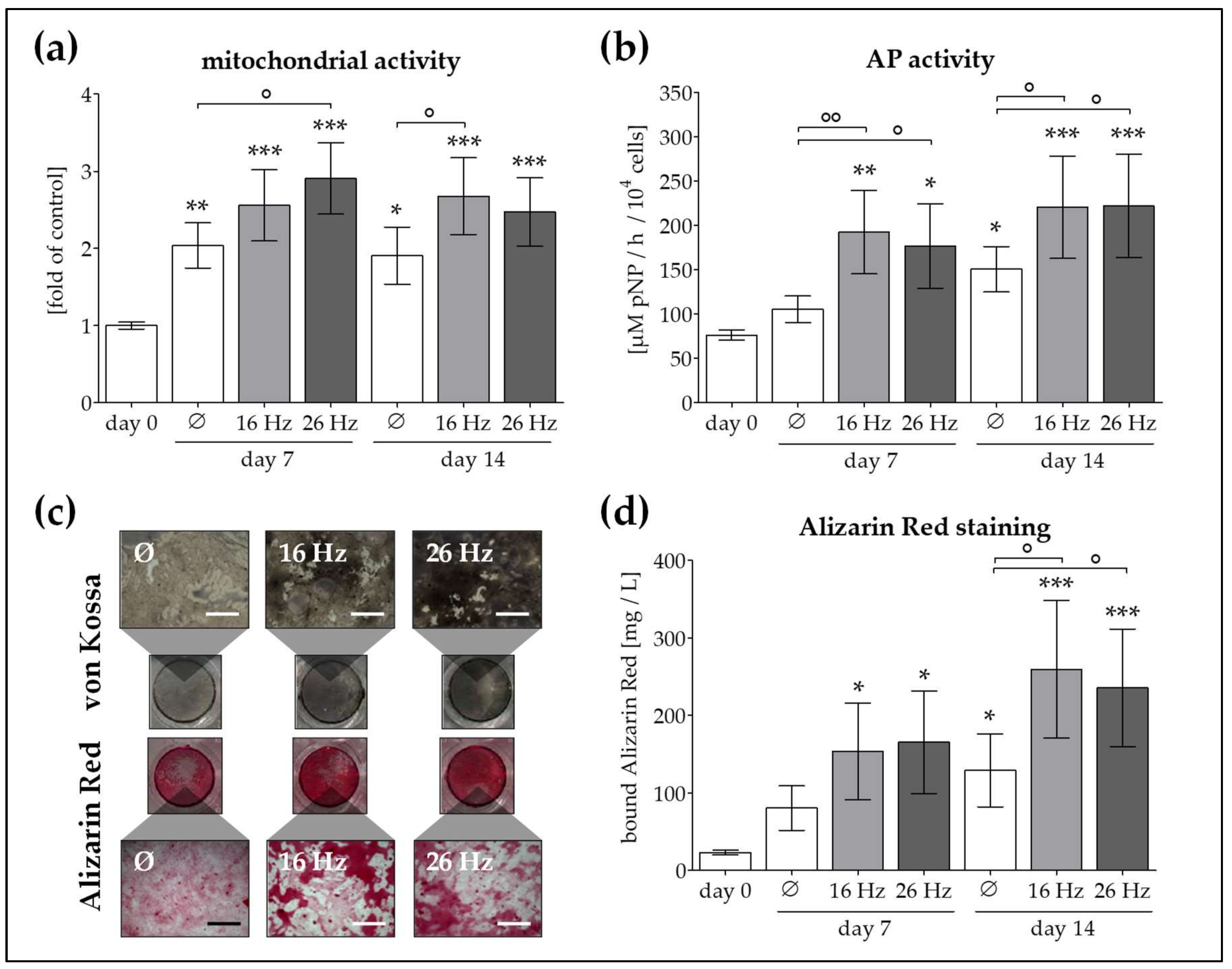
2.3. ELF-PEMF Exposure Improves Viability and Function of OB and Ad-MSC Co-Cultures
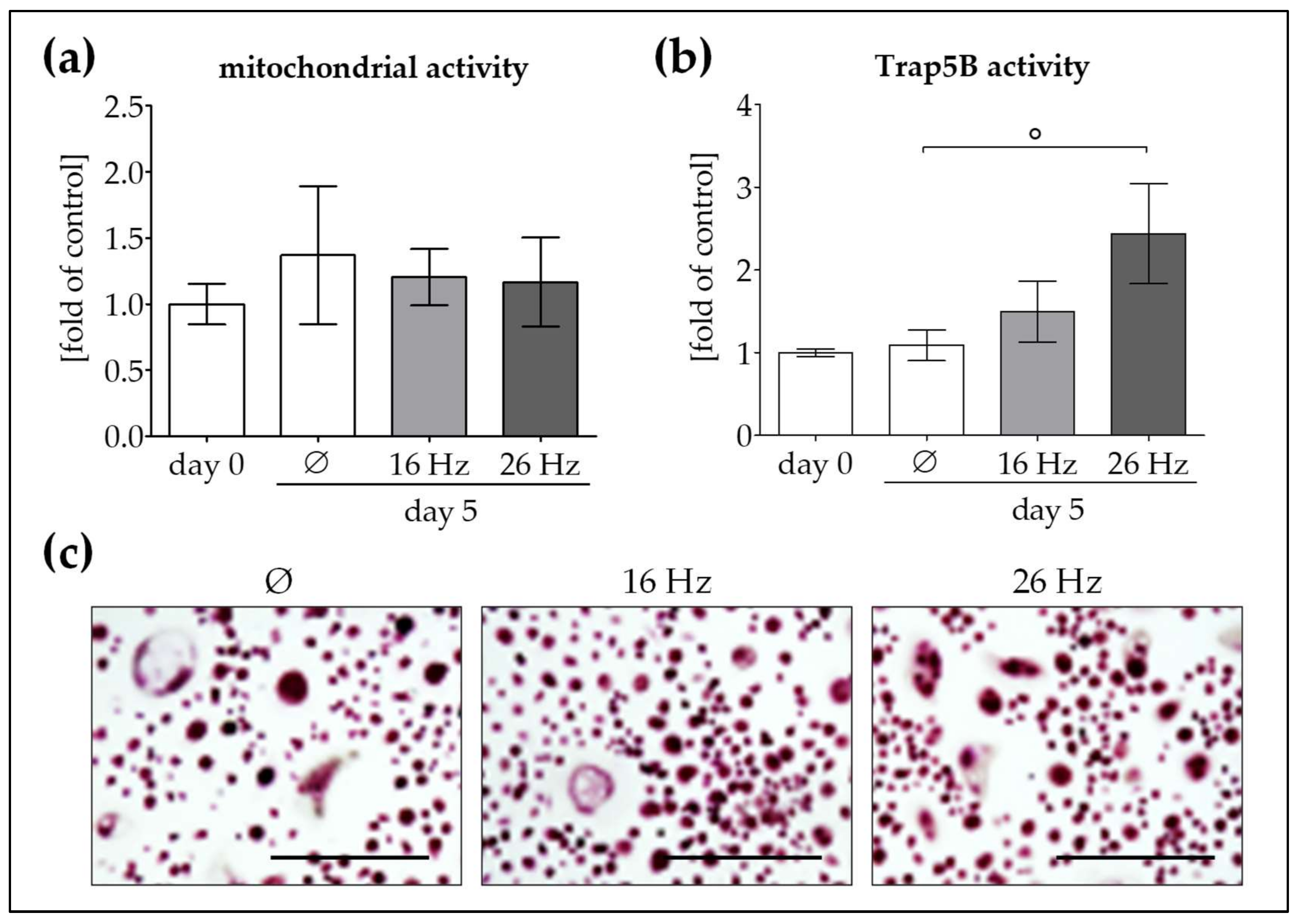
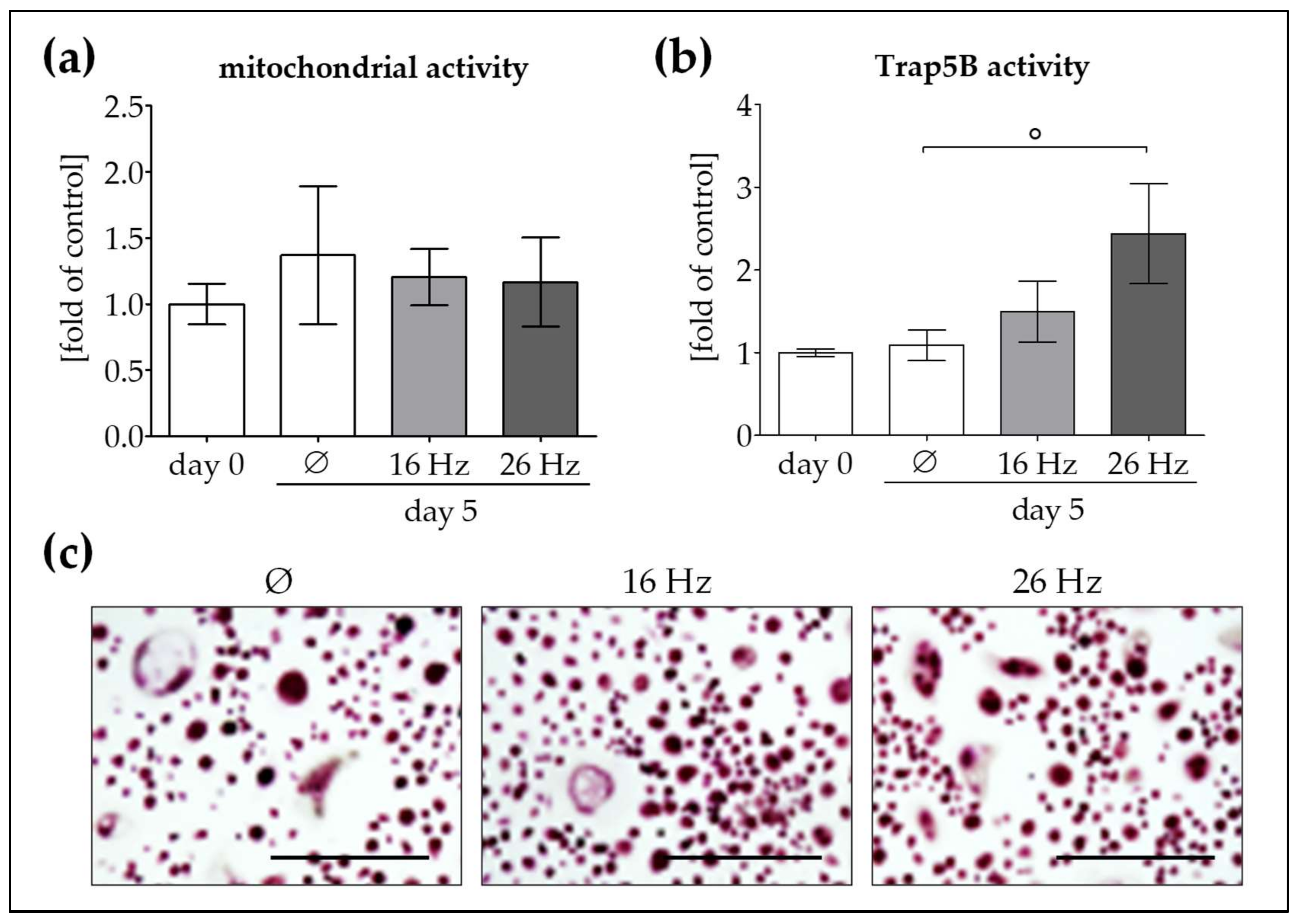
2.4. Increased OC Function by Repetitive Exposure to 26 Hz ELF-PEMF but Not 16 Hz ELF-PEMF
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Isolation and Expansion of OBs
4.3. Isolation and Expansion of Ad-MSCs
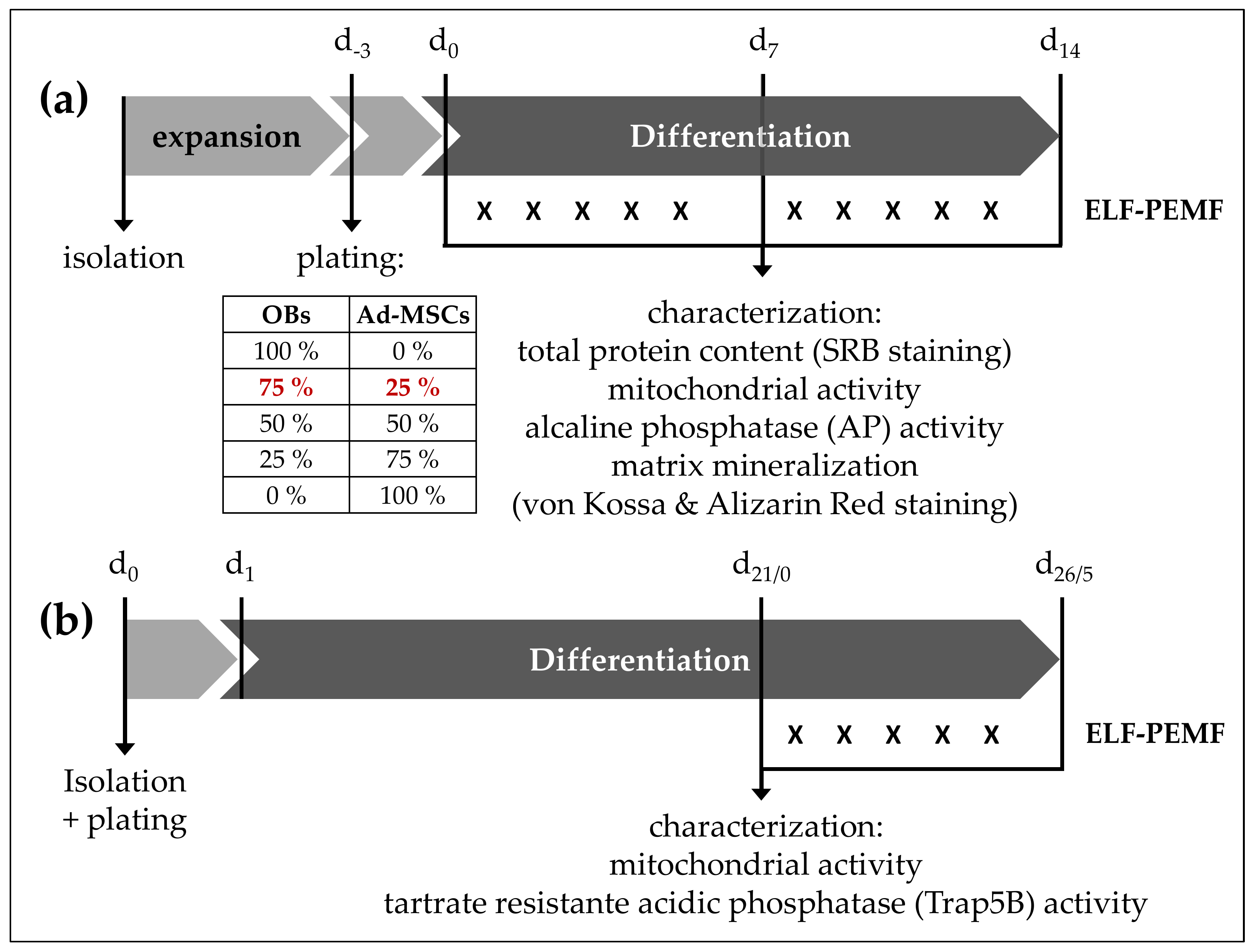
4.4. Osteogenic Differentiation of OBs and Ad-MSCs Mono- and Co-Cultures
4.5. Generation of OCs
4.6. Electromagnetic Field Application with the Somagen® Device
4.7. Sulforhodamine B (SRB) Staining to Assess Total Protein Content
4.8. Resazurin Conversion Assay to Assess Mitochondrial Activity
4.9. AP Activity Measurement
4.10. Assessing Matrix Mineralization by von Kossa and Alizarin Red Staining
4.11. Trap5B Activity Measurement
4.12. Trap5B Staining
4.13. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Ad-MSCs | primary human adipose-derived mesenchymal stem cells |
| B-MSCs | bone marrow-derived mesenchymal stem cells |
| AP | alkaline phosphatase |
| ELF-PEMF | extremely low frequency pulsed electromagnetic fields |
| FCS | fetal calf serum |
| hrM-CSF | human recombinant macrophage colony-stimulating factor |
| hrRANKL | human recombinant receptor activator of nuclear factor κ-B ligand |
| OBs | primary human osteoblasts |
| OCs | primary human osteoclasts |
| Trap5B | tartrate resistant acidic phosphatase |
References
- Smith, J.O.; Aarvold, A.; Tayton, E.R.; Dunlop, D.G.; Oreffo, R.O. Skeletal tissue regeneration: Current approaches, challenges, and novel reconstructive strategies for an aging population. Tissue Eng. Part B Rev. 2011, 17, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Harada, N.; Sato, K.; Abe, S.; Yamanaka, K.; Matushita, T. Stem cell therapy: Is there a future for reconstruction of large bone defects? Injury 2016, 47, S47–S51. [Google Scholar] [CrossRef]
- Granero-Molto, F.; Weis, J.A.; Miga, M.I.; Landis, B.; Myers, T.J.; O’Rear, L.; Longobardi, L.; Jansen, E.D.; Mortlock, D.P.; Spagnoli, A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells 2009, 27, 1887–1898. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, X.; Han, Z.P.; Qu, F.F.; Shao, L.; Shi, Y.F. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Spees, J.L.; Lee, R.H.; Gregory, C.A. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res. Ther. 2016, 7, 125. [Google Scholar] [CrossRef] [PubMed]
- Granero-Molto, F.; Weis, J.A.; Longobardi, L.; Spagnoli, A. Role of mesenchymal stem cells in regenerative medicine: Application to bone and cartilage repair. Expert Opin. Biol. Ther. 2008, 8, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Rando, T.A. Manifestations and mechanisms of stem cell aging. J. Cell Biol. 2011, 193, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, C.; Xie, Z.; Song, P.; Zhao, R.C.; Guo, L.; Liu, Z.; Wu, Y. Epigenetic dysregulation in mesenchymal stem cell aging and spontaneous differentiation. PLoS ONE 2011, 6, e20526. [Google Scholar] [CrossRef] [PubMed]
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Ehnert, S.; Culmes, M.; Bachmann, A.; Seeliger, C.; Schyschka, L.; Wang, Z.; Rahmanian-Schwarz, A.; Stockle, U.; de Sousa, P.A.; et al. 5-azacytidine improves the osteogenic differentiation potential of aged human adipose-derived mesenchymal stem cells by DNA demethylation. PLoS ONE 2014, 9, e90846. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, P.; Fechner, K.; Milz, S.; Richter, W.; Suedkamp, N.P.; Mehlhorn, A.T.; Pearce, S.; Kasten, P. Comparison of mesenchymal stem cells from bone marrow and adipose tissue for bone regeneration in a critical size defect of the sheep tibia and the influence of platelet-rich plasma. Biomaterials 2010, 31, 3572–3579. [Google Scholar] [CrossRef] [PubMed]
- Dawson, J.I.; Oreffo, R.O. Bridging the regeneration gap: Stem cells, biomaterials and clinical translation in bone tissue engineering. Arch. Biochem. Biophys. 2008, 473, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Gershovich, J.G.; Dahlin, R.L.; Kasper, F.K.; Mikos, A.G. Enhanced osteogenesis in cocultures with human mesenchymal stem cells and endothelial cells on polymeric microfiber scaffolds. Tissue Eng. Part A 2013, 19, 2565–2576. [Google Scholar] [CrossRef] [PubMed]
- Bulnheim, U.; Muller, P.; Neumann, H.G.; Peters, K.; Unger, R.E.; Kirkpatrick, C.J.; Rychly, J. Endothelial cells stimulate osteogenic differentiation of mesenchymal stem cells on calcium phosphate scaffolds. J. Tissue Eng. Regen. Med. 2014, 8, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Birmingham, E.; Niebur, G.L.; McHugh, P.E.; Shaw, G.; Barry, F.P.; McNamara, L.M. Osteogenic differentiation of mesenchymal stem cells is regulated by osteocyte and osteoblast cells in a simplified bone niche. Eur. Cell Mater. 2012, 23, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Bassett, C.A.; Pawluk, R.J.; Pilla, A.A. Augmentation of bone repair by inductively coupled electromagnetic fields. Science 1974, 184, 575–577. [Google Scholar] [PubMed]
- Hannouche, D.; Petite, H.; Sedel, L. Current trends in the enhancement of fracture healing. J. Bone Jt. 2001, 83, 157–164. [Google Scholar] [CrossRef]
- Ciombor, D.M.; Aaron, R.K. The role of electrical stimulation in bone repair. Foot Ankle Clin. 2005, 10, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, J.Q.; Ge, B.F.; Ma, X.N.; Ma, H.P.; Xian, C.J.; Chen, K.M. Different electromagnetic field waveforms have different effects on proliferation, differentiation and mineralization of osteoblasts in vitro. Bioelectromagnetics 2014, 35, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, H.; Hirao, K.; Tominaga, T.; Bando, N.; Takahashi, K.; Matsuo, T. Electromagnetic Wave Irradiation Promotes Osteoblastic Cell Proliferation and Up-Regulates Growth Factors via Activation of the ERK1/2 and p38 MAPK Pathways. Cell Physiol. Biochem. 2015, 35, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Falldorf, K.; Fentz, A.-K.; Ziegler, P.; Schröter, S.; Freude, T.; Ochs, B.G.; Stacke, C.; Ronniger, M.; Sachtleben, J.; et al. Primary human osteoblasts with reduced alkaline phosphatase and matrix mineralization baseline capacity are responsive to extremely low frequency pulsed electromagnetic field exposure—Clinical implication possible. Bone Rep. 2015, 3, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Fentz, A.K.; Schreiner, A.; Birk, J.; Wilbrand, B.; Ziegler, P.; Reumann, M.K.; Wang, H.; Falldorf, K.; Nussler, A.K. Extremely low frequency pulsed electromagnetic fields cause antioxidative defense mechanisms in human osteoblasts via induction of •O2− and H2O2. Sci. Rep. 2017, 7, 14544. [Google Scholar] [CrossRef] [PubMed]
- Poh, P.S.P.; Unger, M.; Seelinger, C.; Balmayor, E.R.; van Griensven, M. Low-frequency pulsed electromagnetic fields as treatment for bone healing. Eur. Cell Mater. 2016, 31, 122. [Google Scholar]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon Med. School 2010, 77, 4–12. [Google Scholar] [CrossRef]
- Shetty, S.; Kapoor, N.; Bondu, J.D.; Thomas, N.; Paul, T.V. Bone turnover markers: Emerging tool in the management of osteoporosis. Indian J. Endocrinol. Metab. 2016, 20, 846–852. [Google Scholar] [PubMed]
- Halleen, J.M.; Alatalo, S.L.; Janckila, A.J.; Woitge, H.W.; Seibel, M.J.; Vaananen, H.K. Serum tartrate-resistant acid phosphatase 5b is a specific and sensitive marker of bone resorption. Clin. Chem. 2001, 47, 597–600. [Google Scholar] [PubMed]
- Heino, T.J.; Hentunen, T.A.; Vaananen, H.K. Conditioned medium from osteocytes stimulates the proliferation of bone marrow mesenchymal stem cells and their differentiation into osteoblasts. Exp. Cell Res. 2004, 294, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.S.; Abuna, R.P.; Castro Raucci, L.M.; Teixeira, L.N.; de Oliveira, P.T.; Beloti, M.M.; Rosa, A.L. Mesenchymal Stem Cells Repress Osteoblast Differentiation Under Osteogenic-Inducing Conditions. J. Cell Biochem. 2015, 116, 2896–2902. [Google Scholar] [CrossRef] [PubMed]
- Glueck, M.; Gardner, O.; Czekanska, E.; Alini, M.; Stoddart, M.J.; Salzmann, G.M.; Schmal, H. Induction of Osteogenic Differentiation in Human Mesenchymal Stem Cells by Crosstalk with Osteoblasts. Biores. Open Access 2015, 4, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ming, L.G.; Ge, B.F.; Wang, J.Q.; Zhu, R.Q.; Wei, Z.; Ma, H.P.; Xian, C.J.; Chen, K.M. Effects of 50 Hz sinusoidal electromagnetic fields of different intensities on proliferation, differentiation and mineralization potentials of rat osteoblasts. Bone 2011, 49, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.; Pan, L.; Lee, I. Magnetic fields at extremely low-frequency (50 Hz, 0.8 mT) can induce the uptake of intracellular calcium levels in osteoblasts. Biochem. Biophys. Res. Commun. 2010, 396, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: A revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.M.; Kang, K.S.; Yi, H.G.; Kim, S.Y.; Cho, D.W. Electromagnetically controllable osteoclast activity. Bone 2014, 62, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.; McLeod, K.J.; Titus, L.; Nanes, M.S.; Catherwood, B.D.; Rubin, C.T. Formation of osteoclast-like cells is suppressed by low frequency, low intensity electric fields. J. Orthop. Res. 1996, 14, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.; Unger, M.; van Griensven, M.; Balmayor, E.R. Adipose-derived mesenchymal stem cells from liposuction and resected fat are feasible sources for regenerative medicine. Eur. J. Med. Res. 2017, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Prause, M.; Seeliger, C.; Unger, M.; van Griensven, M.; Haug, A.T. Pantoprazole increases cell viability and function of primary human osteoblasts in vitro. Injury 2014, 45, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Seeliger, C.; Vester, H.; Schmitt, A.; Saidy-Rad, S.; Lin, J.; Neumaier, M.; Gillen, S.; Kleeff, J.; Friess, H.; et al. Autologous serum improves yield and metabolic capacity of monocyte-derived hepatocyte-like cells: Possible implication for cell transplantation. Cell Transpl. 2011, 20, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Kelch, S.; Balmayor, E.R.; Seeliger, C.; Vester, H.; Kirschke, J.S.; van Griensven, M. miRNAs in bone tissue correlate to bone mineral density and circulating miRNAs are gender independent in osteoporotic patients. Sci. Rep. 2017, 7, 15861. [Google Scholar] [CrossRef] [PubMed]
- Prause, M.; Seeliger, C.; Unger, M.; Rosado Balmayor, E.; van Griensven, M.; Haug, A.T. Pantoprazole decreases cell viability and function of human osteoclasts in vitro. Mediat. Inflamm. 2015, 2015, 413097. [Google Scholar] [CrossRef] [PubMed]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Ehnert, S.; Schyschka, L.; Buschner, P.; Kuhnl, A.; Dobele, S.; Siebenlist, S.; Lucke, M.; Stockle, U.; Nussler, A.K. Monocytes do not transdifferentiate into proper osteoblasts. Sci. World J. 2012, 2012, 384936. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ehnert, S.; Van Griensven, M.; Unger, M.; Scheffler, H.; Falldorf, K.; Fentz, A.-K.; Seeliger, C.; Schröter, S.; Nussler, A.K.; Balmayor, E.R. Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 994. https://doi.org/10.3390/ijms19040994
Ehnert S, Van Griensven M, Unger M, Scheffler H, Falldorf K, Fentz A-K, Seeliger C, Schröter S, Nussler AK, Balmayor ER. Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2018; 19(4):994. https://doi.org/10.3390/ijms19040994
Chicago/Turabian StyleEhnert, Sabrina, Martijn Van Griensven, Marina Unger, Hanna Scheffler, Karsten Falldorf, Anne-Kristin Fentz, Claudine Seeliger, Steffen Schröter, Andreas K. Nussler, and Elizabeth R. Balmayor. 2018. "Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells" International Journal of Molecular Sciences 19, no. 4: 994. https://doi.org/10.3390/ijms19040994
APA StyleEhnert, S., Van Griensven, M., Unger, M., Scheffler, H., Falldorf, K., Fentz, A.-K., Seeliger, C., Schröter, S., Nussler, A. K., & Balmayor, E. R. (2018). Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences, 19(4), 994. https://doi.org/10.3390/ijms19040994