cGMP Signaling in the Cardiovascular System—The Role of Compartmentation and Its Live Cell Imaging

Abstract
1. cGMP Signaling in the Cardiovascular System
1.1. cGMP Synthesis
1.2. cGMP Effector Activation
1.3. cGMP Catabolism by Phosphodiesterases
2. Live Cell Imaging of cGMP
3. Compartmentation of cGMP Signaling in the Cardiovascular System
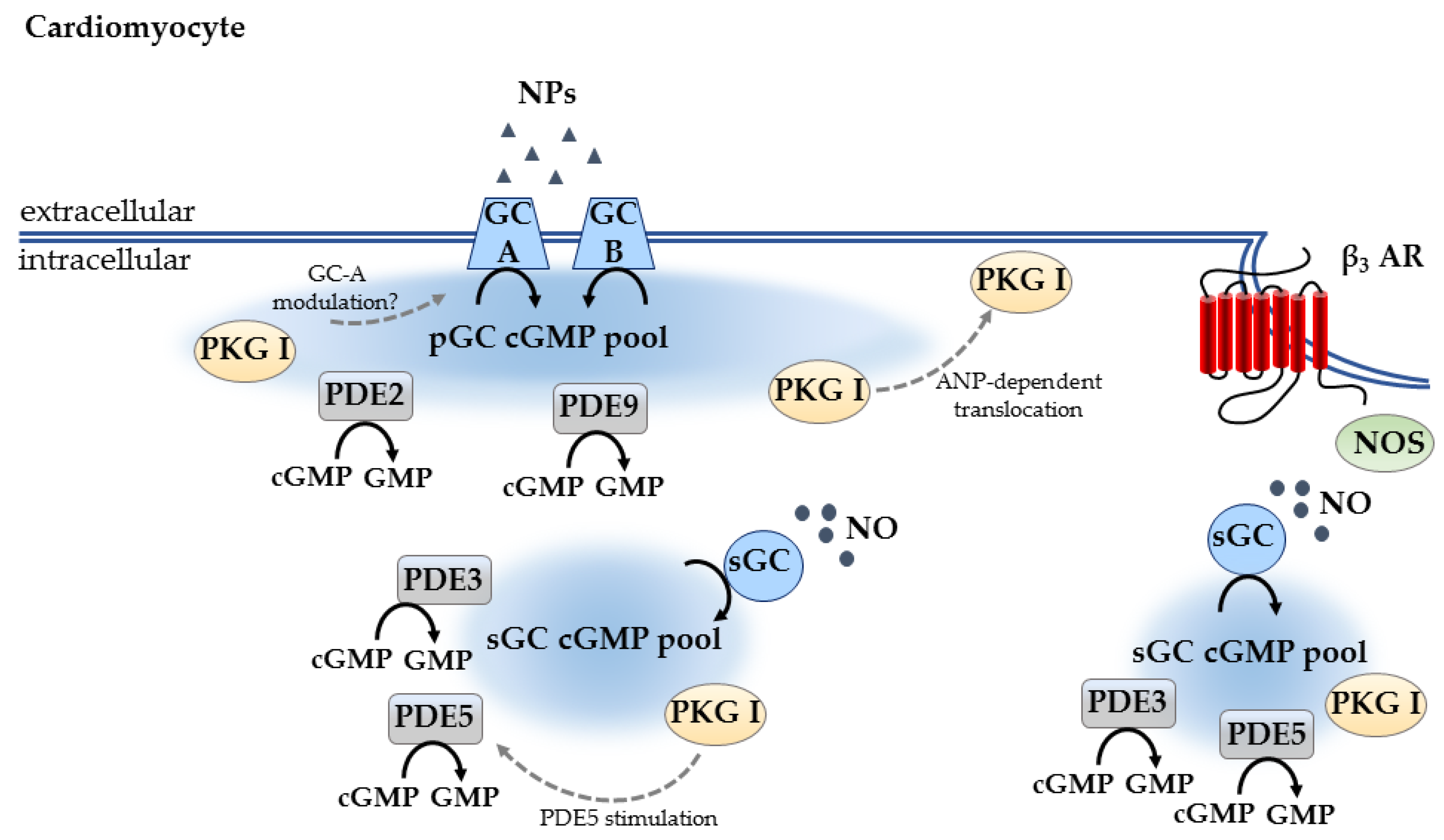
3.1. Compartmentation in Cardiomyocytes
3.2. Compartmentation in Other Cell Types
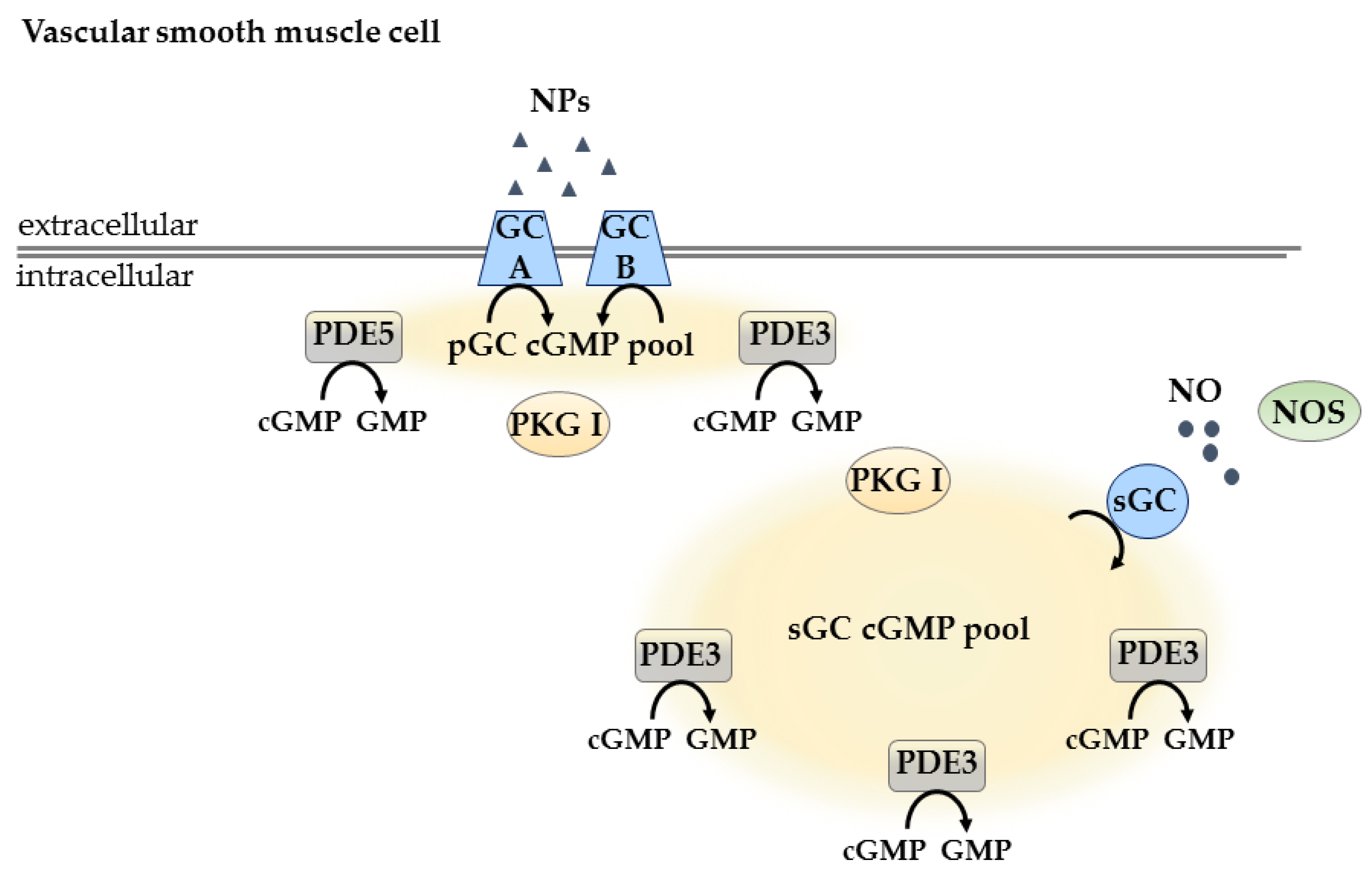
3.2.1. Compartmentation in Vascular Smooth Muscle Cells
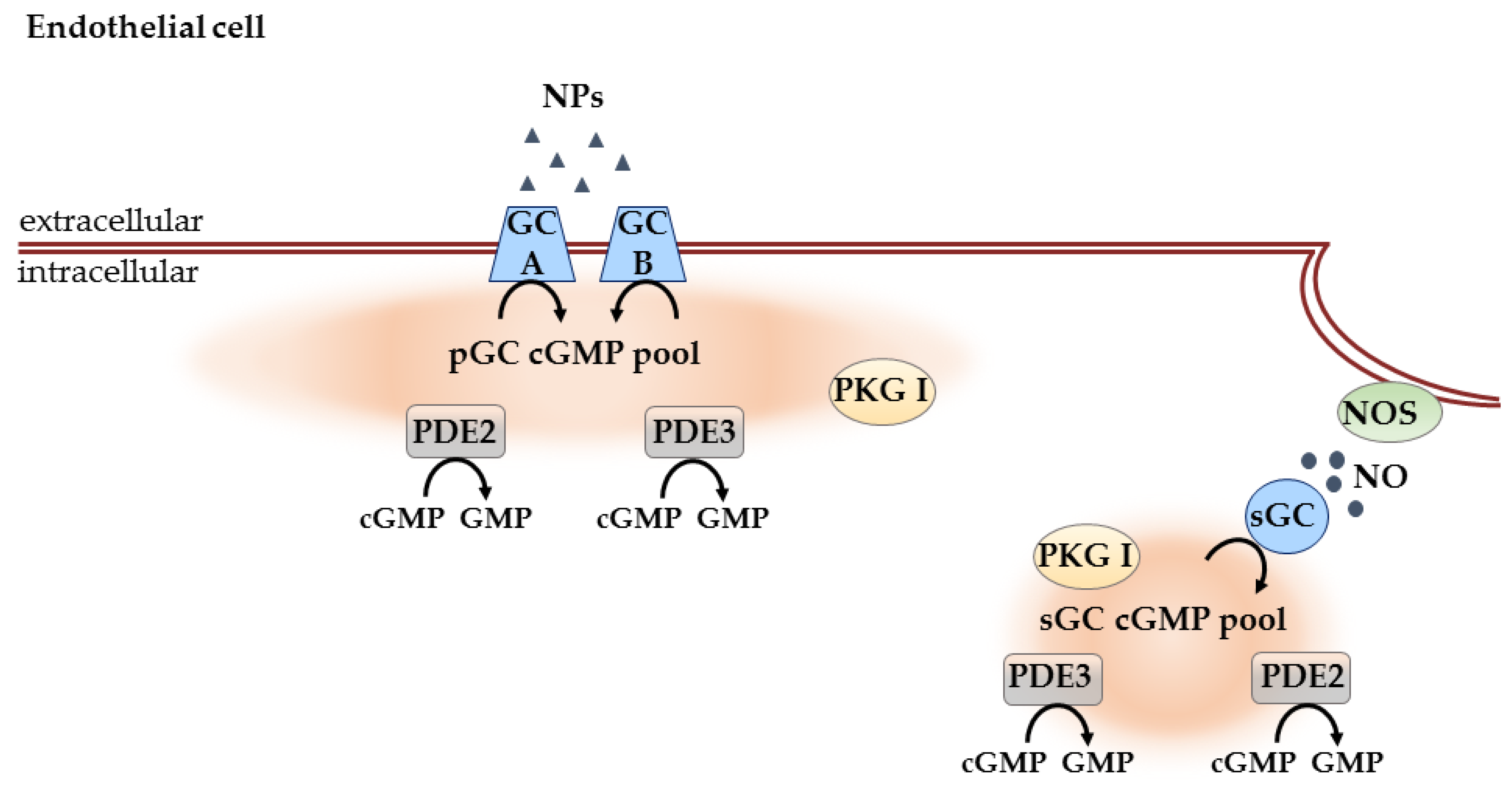
3.2.2. Compartmentation in Endothelial Cells
3.2.3. Compartmentation in Cardiac Fibroblasts
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACh | Acetylcholine |
| AChR | Acetylcholine receptor |
| AMP | Adenosine monophosphate |
| ANP | Atrial natriuretic peptide |
| BNP | Brain natriuretic peptide |
| CAG | Cytomegalovirus early enhancer/chicken β-actin/β-globin |
| cAMP | Adenosine monophosphate |
| CF | Cardiac fibroblast |
| cGMP | Cyclic guanosine 3′,5′-monophosphate |
| CM | Cardiomyocyte |
| CNG | Cyclic nucleotide gated channels |
| CNP | C-type natriuretic peptide |
| DEA/NO | 2-(N,N-dethylamino)-diazenolate-2-oxide dethylammonium salt |
| EC | Endothelial cell |
| ECFP | Enhanced cyan fluorescent protein |
| ECM | Extracellular matrix |
| EDRF | Endothelium derived relaxation factor |
| ELISA | Enzyme-linked immunoassay |
| eNOS | Endothelial NO synthase |
| EYFP | Enhanced yellow fluorescent protein |
| FRET | Förster resonance energy transfer |
| GC | Guanylyl cyclase |
| GMP | Guanosine monophosphate |
| GTP | Guanosine triphosphate |
| HUVEC | Human umbilical vein endothelial cell |
| IBMX | 3-isobutyl-1-methylxanthine |
| mAChR | Muscarinic acetylcholine receptor |
| iNOS | Inducible NO synthase |
| nAChR | Nicotinic acetylcholine receptor |
| NO | Nitric oxide |
| NOS | NO synthase |
| NP | Natriuretic peptide |
| PDE | Phosphodiesterase |
| pGC | Particulate guanylyl cyclase |
| PKG | cGMP-dependent protein kinase |
| PKG I | cGMP-dependent protein kinase type I |
| PKG II | cGMP-dependent protein kinase type II |
| RGD | Arg-Gly-Asp |
| sGC | Soluble guanylyl cyclase |
| VSMC | Vascular smooth muscle cell |
References
- Tsai, E.J.; Kass, D.A. Cyclic GMP signaling in cardiovascular pathophysiology and therapeutics. Pharmacol. Ther. 2009, 122, 216–238. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.; Zeller, M.; Guan, K.; Wunder, F.; Wagner, M.; El-Armouche, A. PDE2 at the crossway between cAMP and cGMP signalling in the heart. Cell. Signal. 2017, 38, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Feil, R.; Lohmann, S.M.; de Jonge, H.; Walter, U.; Hofmann, F. Cyclic GMP-Dependent Protein Kinases and the Cardiovascular System. Circ. Res. 2003, 93, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, E. Cyclic GMP-dependent signaling in cardiac myocytes. Circ. J. Off. J. Jpn. Circ. Soc. 2012, 76, 1819–1825. [Google Scholar] [CrossRef]
- Ashman, D.F.; Lipton, R.; Melicow, M.M.; Price, T.D. Isolation of adenosine 3′,5′-monophosphate and guanosine 3′,5′-monophosphate from rat urine. Biochem. Biophys. Res. Commun. 1963, 11, 330–334. [Google Scholar] [CrossRef]
- Beavo, J.A.; Brunton, L.L. Cyclic nucleotide research—Still expanding after half a century. Nat. Rev. Mol. Cell Biol. 2002, 3, 710. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, R.; Krieg, T.; Rybalkin, S.D.; Beavo, J.; Hofmann, F. Turning on cGMP-dependent pathways to treat cardiac dysfunctions: Boom, bust, and beyond. Trends Pharmacol. Sci. 2014, 35, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M. Endothelial actions of atrial and B-type natriuretic peptides. Br. J. Pharmacol. 2012, 166, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Baxter, G.F. Natriuretic peptides and myocardial ischaemia. Basic Res. Cardiol. 2004, 99, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Chinkers, M.; Garbers, D.L.; Chang, M.S.; Lowe, D.G.; Chin, H.M.; Goeddel, D.V.; Schulz, S. A membrane form of guanylate cyclase is an atrial natriuretic peptide receptor. Nature 1989, 338, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, E.; Champion, H.C.; Li, M.; Belardi, D.; Ren, S.; Rodriguez, E.R.; Bedja, D.; Gabrielson, K.L.; Wang, Y.; Kass, D.A. Chronic inhibition of cyclic GMP phosphodiesterase 5A prevents and reverses cardiac hypertrophy. Nat. Med. 2005, 11, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Cerra, M.C.; Pellegrino, D. Cardiovascular cGMP-generating systems in physiological and pathological conditions. Curr. Med. Chem. 2007, 14, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Russwurm, M.; Wittau, N.; Koesling, D. Guanylyl cyclase/PSD-95 interaction: Targeting of the nitric oxide-sensitive α2β1 guanylyl cyclase to synaptic membranes. J. Biol. Chem. 2001, 276, 44647–44652. [Google Scholar] [CrossRef] [PubMed]
- Montfort, W.R.; Wales, J.A.; Weichsel, A. Structure and Activation of Soluble Guanylyl Cyclase, the Nitric Oxide Sensor. Antioxid. Redox Signal. 2017, 26, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Moody, B.F.; Calvert, J.W. Emergent role of gasotransmitters in ischemia-reperfusion injury. Med. Gas. Res. 2011, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Rastaldo, R.; Pagliaro, P.; Cappello, S.; Penna, C.; Mancardi, D.; Westerhof, N.; Losano, G. Nitric oxide and cardiac function. Life Sci. 2007, 81, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, R.M.; Hare, J.M. Nitric oxide signaling in the cardiovascular system: Implications for heart failure. Curr. Opin. Cardiol. 2006, 21, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M. Structure, regulation, and function of mammalian membrane guanylyl cyclase receptors, with a focus on guanylyl cyclase-A. Circ. Res. 2003, 93, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M. Molecular Physiology of Membrane Guanylyl Cyclase Receptors. Physiol. Rev. 2016, 96, 751–804. [Google Scholar] [CrossRef] [PubMed]
- Garbers, D.L.; Koesling, D.; Schultz, G. Guanylyl cyclase receptors. Mol. Biol. Cell 1994, 5, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Woodard, G.E.; Rosado, J.A. Natriuretic peptides in vascular physiology and pathology. Int. Rev. Cell Mol. Biol. 2008, 268, 59–93. [Google Scholar] [PubMed]
- Nakao, K.; Ogawa, Y.; Suga, S.; Imura, H. Molecular biology and biochemistry of the natriuretic peptide system. II: Natriuretic peptide receptors. J. Hypertens. 1992, 10, 1111–1114. [Google Scholar] [CrossRef] [PubMed]
- Koller, K.J.; Lowe, D.G.; Bennett, G.L.; Minamino, N.; Kangawa, K.; Matsuo, H.; Goeddel, D.V. Selective activation of the B natriuretic peptide receptor by C-type natriuretic peptide (CNP). Science 1991, 252, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Zaccolo, M.; Movsesian, M.A. cAMP and cGMP Signaling Cross-Talk: Role of Phosphodiesterases and Implications for Cardiac Pathophysiology. Circ. Res. 2007, 100, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Evgenov, O.V.; Pacher, P.; Schmidt, P.M.; Hasko, G.; Schmidt, H.H.; Stasch, J.P. NO-independent stimulators and activators of soluble guanylate cyclase: Discovery and therapeutic potential. Nat. Rev. Drug. Discov. 2006, 5, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Linder, A.E.; McCluskey, L.P.; Cole, K.R., 3rd; Lanning, K.M.; Webb, R.C. Dynamic association of nitric oxide downstream signaling molecules with endothelial caveolin-1 in rat aorta. J. Pharmacol. Exp. Ther. 2005, 314, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Venema, R.C.; Venema, V.J.; Ju, H.; Harris, M.B.; Snead, C.; Jilling, T.; Dimitropoulou, C.; Maragoudakis, M.E.; Catravas, J.D. Novel complexes of guanylate cyclase with heat shock protein 90 and nitric oxide synthase. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H669–H678. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Karlin, A.; Cox, R.N.; Dipaola, M.; Holtzman, E.; Kao, P.N.; Lobel, P.; Wang, L.; Yodh, N. Functional domains of the nicotinic acetylcholine receptor. Ann. N. Y. Acad. Sci. 1986, 463, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.D.; Belevych, A.E. Muscarinic regulation of cardiac ion channels. Br. J. Pharmacol. 2003, 139, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.P.; Ghebremariam, Y.T. Endothelial nicotinic acetylcholine receptors and angiogenesis. Trends Cardiovasc. Med. 2008, 18, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Eglen, R.M. Overview of muscarinic receptor subtypes. Handb. Exp. Pharmacol. 2012, 3–28. [Google Scholar] [CrossRef]
- Attina, T.M.; Oliver, J.J.; Malatino, L.S.; Webb, D.J. Contribution of the M3 muscarinic receptors to the vasodilator response to acetylcholine in the human forearm vascular bed. Br. J. Clin. Pharmacol. 2008, 66, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Eglen, R.M.; Reddy, H.; Watson, N.; Challiss, R.A. Muscarinic acetylcholine receptor subtypes in smooth muscle. Trends Pharmacol. Sci. 1994, 15, 114–119. [Google Scholar] [CrossRef]
- Goutsouliak, V.; Wang, Y.; Cynader, M.S.; Rabkin, S.W. Visualization of muscarinic cholinergic receptors on chick cardiomyocytes and their involvement in phosphatidylcholine hydrolysis. Biochem. Cell Biol. 1997, 75, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.; Lips, K.S.; Bruggmann, D.; Slavikova, J.; Kuncova, J.; Kummer, W. Developmental changes in the expression of nicotinic acetylcholine receptor α-subunits in the rat heart. Cell Tissue Res. 2005, 319, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Buga, G.M.; Wood, K.S.; Byrns, R.E.; Chaudhuri, G. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci. USA 1987, 84, 9265–9269. [Google Scholar] [CrossRef] [PubMed]
- Furchgott, R.F.; Vanhoutte, P.M. Endothelium-derived relaxing and contracting factors. FASEB J. 1989, 3, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, R.M.; Murad, F. Agonist-induced endothelium-dependent relaxation in rat thoracic aorta may be mediated through cGMP. Circ. Res. 1983, 52, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Griffith, T.M.; Edwards, D.H.; Lewis, M.J.; Newby, A.C.; Henderson, A.H. The nature of endothelium-derived vascular relaxant factor. Nature 1984, 308, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Forstermann, U.; Mulsch, A.; Bohme, E.; Busse, R. Stimulation of soluble guanylate cyclase by an acetylcholine-induced endothelium-derived factor from rabbit and canine arteries. Circ. Res. 1986, 58, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Brodde, O.E.; Michel, M.C. Adrenergic and muscarinic receptors in the human heart. Pharmacol. Rev. 1999, 51, 651–690. [Google Scholar] [PubMed]
- Jungen, C.; Scherschel, K.; Eickholt, C.; Kuklik, P.; Klatt, N.; Bork, N.; Salzbrunn, T.; Alken, F.; Angendohr, S.; Klene, C.; et al. Disruption of cardiac cholinergic neurons enhances susceptibility to ventricular arrhythmias. Nat. Commun. 2017, 8, 14155. [Google Scholar] [CrossRef] [PubMed]
- Mavropoulos, S.A.; Khan, N.S.; Levy, A.C.J.; Faliks, B.T.; Sison, C.P.; Pavlov, V.A.; Zhang, Y.; Ojamaa, K. Nicotinic acetylcholine receptor-mediated protection of the rat heart exposed to ischemia reperfusion. Mol. Med. 2017, 23, 120. [Google Scholar] [CrossRef] [PubMed]
- Gileadi, O. Structures of soluble guanylate cyclase: Implications for regulatory mechanisms and drug development. Biochem. Soc. Trans. 2014, 42, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, R.; Rybalkin, S.D.; Loga, F.; Leiss, V.; Beavo, J.A.; Hofmann, F. Cardiac hypertrophy is not amplified by deletion of cGMP-dependent protein kinase I in cardiomyocytes. Proc. Natl. Acad. Sci. USA 2010, 107, 5646–5651. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, F.; Bernhard, D.; Lukowski, R.; Weinmeister, P. cGMP regulated protein kinases (cGK). Handb. Exp. Pharmacol. 2009, 137–162. [Google Scholar] [CrossRef]
- Lorenz, R.; Bertinetti, D.; Herberg, F.W. cAMP-Dependent Protein Kinase and cGMP-Dependent Protein Kinase as Cyclic Nucleotide Effectors. Handb. Exp. Pharmacol. 2017, 238, 105–122. [Google Scholar] [PubMed]
- Kim, G.E.; Kass, D.A. Cardiac Phosphodiesterases and Their Modulation for Treating Heart Disease. Handb. Exp. Pharmacol. 2017, 243, 249–269. [Google Scholar] [PubMed]
- Biel, M.; Michalakis, S. Cyclic nucleotide-gated channels. Handb. Exp. Pharmacol. 2009, 111–136. [Google Scholar] [CrossRef]
- Kaupp, U.B.; Seifert, R. Cyclic nucleotide-gated ion channels. Physiol. Rev. 2002, 82, 769–824. [Google Scholar] [CrossRef] [PubMed]
- Bradley, J.; Reisert, J.; Frings, S. Regulation of cyclic nucleotide-gated channels. Curr. Opin. Neurobiol. 2005, 15, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 58, 488–520. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, K.; Kass, D.A. Nanodomain Regulation of Cardiac Cyclic Nucleotide Signaling by Phosphodiesterases. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 455–479. [Google Scholar] [CrossRef] [PubMed]
- Brescia, M.; Zaccolo, M. Modulation of Compartmentalised Cyclic Nucleotide Signalling via Local Inhibition of Phosphodiesterase Activity. Int. J. Mol. Sci. 2016, 17, 1672. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Annu. Rev. Biochem. 2007, 76, 481–511. [Google Scholar] [CrossRef] [PubMed]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef] [PubMed]
- Degerman, E.; Belfrage, P.; Manganiello, V.C. Structure, localization, and regulation of cGMP-inhibited phosphodiesterase (PDE3). J. Biol. Chem. 1997, 272, 6823–6826. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.I.; Kass, D.A. Phosphodiesterases and cyclic GMP regulation in heart muscle. Physiology 2012, 27, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.L.; Yan, C. Targeting cyclic nucleotide phosphodiesterase in the heart: Therapeutic implications. J. Cardiovasc. Transl. Res. 2010, 3, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kass, D.A. Phosphodiesterases and cardiac cGMP: Evolving roles and controversies. Trends Pharmacol. Sci. 2011, 32, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Götz, K.; Sprenger, J.; Perera, R.K.; Steinbrecher, J.H.; Lehnart, S.E.; Kuhn, M.; Gorelik, J.; Balligand, J.-L.; Nikolaev, V.O. Transgenic Mice for Real Time Visualization of cGMP in Intact Adult Cardiomyocytes. Circ. Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, J.U.; Nikolaev, V.O. Biophysical techniques for detection of cAMP and cGMP in living cells. Int. J. Mol. Sci. 2013, 14, 8025–8046. [Google Scholar] [CrossRef] [PubMed]
- Barsony, J.; Marx, S.J. Immunocytology on microwave-fixed cells reveals rapid and agonist-specific changes in subcellular accumulation patterns for cAMP or cGMP. Proc. Natl. Acad. Sci. USA 1990, 87, 1188–1192. [Google Scholar] [CrossRef] [PubMed]
- Brooker, G.; Harper, J.F.; Terasaki, W.L.; Moylan, R.D. Radioimmunoassay of cyclic AMP and cyclic GMP. Adv. Cycl. Nucleotide Res. 1979, 10, 1–33. [Google Scholar]
- Williams, C. cAMP detection methods in HTS: Selecting the best from the rest. Nat. Rev. Drug. Discov. 2004, 3, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Adams, S.R.; Sawyer, C.L.; Lev-Ram, V.; Tsien, R.Y.; Dostmann, W.R. Spatiotemporal dynamics of guanosine 3′,5′-cyclic monophosphate revealed by a genetically encoded, fluorescent indicator. Proc. Natl. Acad. Sci. USA 2001, 98, 2437–2442. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, B.; Kramer, R.H. Real-time patch-cram detection of intracellular cGMP reveals long-term suppression of responses to NO and muscarinic agonists. Neuron 1998, 21, 895–906. [Google Scholar] [CrossRef]
- Goulding, E.H.; Tibbs, G.R.; Siegelbaum, S.A. Molecular mechanism of cyclic-nucleotide-gated channel activation. Nature 1994, 372, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.R.; Verde, I.; Cooper, D.M.; Fischmeister, R. Cyclic guanosine monophosphate compartmentation in rat cardiac myocytes. Circulation 2006, 113, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Visweswariah, S.S. Illuminating Cyclic Nucleotides: Sensors for cAMP and cGMP and Their Application in Live Cell Imaging. J. Indian Inst. Sci. 2017, 97, 109–128. [Google Scholar] [CrossRef]
- Kraft, A.E.; Nikolaev, V.O. FRET Microscopy for Real-Time Visualization of Second Messengers in Living Cells. In Light Microscopy: Methods and Protocols; Markaki, Y., Harz, H., Eds.; Springer: New York, NY, USA, 2017; pp. 85–90. [Google Scholar] [CrossRef]
- Förster, T. Zwischenmolekulare Energiewanderung und Fluoreszenz. Annalen der Physik 1948, 437, 55–75. [Google Scholar] [CrossRef]
- Miyawaki, A. Visualization of the Spatial and Temporal Dynamics of Intracellular Signaling. Dev. Cell 2003, 4, 295–305. [Google Scholar] [CrossRef]
- Shrestha, D.; Jenei, A.; Nagy, P.; Vereb, G.; Szollosi, J. Understanding FRET as a research tool for cellular studies. Int. J. Mol. Sci. 2015, 16, 6718–6756. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, V.O.; Gambaryan, S.; Lohse, M.J. Fluorescent sensors for rapid monitoring of intracellular cGMP. Nat. Methods 2006, 3, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Hida, N.; Ozawa, T.; Umezawa, Y. Fluorescent indicators for cyclic GMP based on cyclic GMP-dependent protein kinase Iα and green fluorescent proteins. Anal. Chem. 2000, 72, 5918–5924. [Google Scholar] [CrossRef] [PubMed]
- Stangherlin, A.; Gesellchen, F.; Zoccarato, A.; Terrin, A.; Fields, L.A.; Berrera, M.; Surdo, N.C.; Craig, M.A.; Smith, G.; Hamilton, G.; et al. cGMP signals modulate cAMP levels in a compartment-specific manner to regulate catecholamine-dependent signaling in cardiac myocytes. Circ. Res. 2011, 108, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Herget, S.; Lohse, M.J.; Nikolaev, V.O. Real-time monitoring of phosphodiesterase inhibition in intact cells. Cell. Signal. 2008, 20, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Couto, A.; Oda, S.; Nikolaev, V.O.; Soltesz, Z.; de Bono, M. In vivo genetic dissection of O2-evoked cGMP dynamics in a Caenorhabditis elegans gas sensor. Proc. Natl. Acad. Sci. USA 2013, 110, E3301–E3310. [Google Scholar] [CrossRef] [PubMed]
- Russwurm, M.; Mullershausen, F.; Friebe, A.; Jager, R.; Russwurm, C.; Koesling, D. Design of fluorescence resonance energy transfer (FRET)-based cGMP indicators: A systematic approach. Biochem. J. 2007, 407, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Thunemann, M.; Wen, L.; Hillenbrand, M.; Vachaviolos, A.; Feil, S.; Ott, T.; Han, X.; Fukumura, D.; Jain, R.K.; Russwurm, M.; et al. Transgenic mice for cGMP imaging. Circ. Res. 2013, 113, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Belge, C.; Hammond, J.; Dubois-Deruy, E.; Manoury, B.; Hamelet, J.; Beauloye, C.; Markl, A.; Pouleur, A.C.; Bertrand, L.; Esfahani, H.; et al. Enhanced expression of β3-adrenoceptors in cardiac myocytes attenuates neurohormone-induced hypertrophic remodeling through nitric oxide synthase. Circulation 2014, 129, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Niino, Y.; Hotta, K.; Oka, K. Simultaneous Live Cell Imaging Using Dual FRET Sensors with a Single Excitation Light. PLoS ONE 2009, 4, e6036. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, Y.; Hampden-Smith, K.; Chachlaki, K.; Wood, K.C.; Vernon, J.; Allerston, C.K.; Batchelor, A.M.; Garthwaite, J. Improved genetically-encoded, FlincG-type fluorescent biosensors for neural cGMP imaging. Front. Mol. Neurosci. 2013, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Nausch, L.W.; Ledoux, J.; Bonev, A.D.; Nelson, M.T.; Dostmann, W.R. Differential patterning of cGMP in vascular smooth muscle cells revealed by single GFP-linked biosensors. Proc. Natl. Acad. Sci. USA 2008, 105, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, P.; Cardinal, R.; De Lean, A.; Lambert, C. Direct chronotropic effects of atrial and C-type natriuretic peptides in anaesthetized dogs. Br. J. Pharmacol. 1996, 118, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Hirose, M.; Furukawa, Y.; Miyashita, Y.; Kurogouchi, F.; Nakajima, K.; Tsuboi, M.; Chiba, S. CNP causes receptor-mediated positive dromotropic effects in anesthetized dog hearts. Am. J. Physiol. 1998, 275, H717–H720. [Google Scholar] [CrossRef] [PubMed]
- Nir, A.; Zhang, D.F.; Fixler, R.; Burnett, J.C., Jr.; Eilam, Y.; Hasin, Y. C-type natriuretic peptide has a negative inotropic effect on cardiac myocytes. Eur. J. Pharmacol. 2001, 412, 195–201. [Google Scholar] [CrossRef]
- Fixler, R.; Hasin, Y.; Eilam, Y.; Zhang, D.F.; Nir, A. Opposing effects of endothelin-1 on C-type natriuretic peptide actions in rat cardiomyocytes. Eur. J. Pharmacol. 2001, 423, 95–98. [Google Scholar] [CrossRef]
- Brusq, J.M.; Mayoux, E.; Guigui, L.; Kirilovsky, J. Effects of C-type natriuretic peptide on rat cardiac contractility. Br. J. Pharmacol. 1999, 128, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Pierkes, M.; Gambaryan, S.; Boknik, P.; Lohmann, S.M.; Schmitz, W.; Potthast, R.; Holtwick, R.; Kuhn, M. Increased effects of C-type natriuretic peptide on cardiac ventricular contractility and relaxation in guanylyl cyclase A-deficient mice. Cardiovasc. Res. 2002, 53, 852–861. [Google Scholar] [CrossRef]
- Wollert, K.C.; Yurukova, S.; Kilic, A.; Begrow, F.; Fiedler, B.; Gambaryan, S.; Walter, U.; Lohmann, S.M.; Kuhn, M. Increased effects of C-type natriuretic peptide on contractility and calcium regulation in murine hearts overexpressing cyclic GMP-dependent protein kinase I. Br. J. Pharmacol. 2003, 140, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Neyses, L.; Vetter, H. Action of atrial natriuretic peptide and angiotensin II on the myocardium: Studies in isolated rat ventricular cardiomyocytes. Biochem. Biophys. Res. Commun. 1989, 163, 1435–1443. [Google Scholar] [CrossRef]
- McCall, D.; Fried, T.A. Effect of atriopeptin II on Ca influx, contractile behavior and cyclic nucleotide content of cultured neonatal rat myocardial cells. J. Mol. Cell. Cardiol. 1990, 22, 201–212. [Google Scholar] [CrossRef]
- Flesch, M.; Kilter, H.; Cremers, B.; Lenz, O.; Sudkamp, M.; Kuhn-Regnier, F.; Bohm, M. Acute effects of nitric oxide and cyclic GMP on human myocardial contractility. J. Pharmacol. Exp. Ther. 1997, 281, 1340–1349. [Google Scholar] [PubMed]
- Kojda, G.; Kottenberg, K.; Nix, P.; Schluter, K.D.; Piper, H.M.; Noack, E. Low increase in cGMP induced by organic nitrates and nitrovasodilators improves contractile response of rat ventricular myocytes. Circ. Res. 1996, 78, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, A.S.; Ma, X.L.; Buerke, M.; Murohara, T.; Armstead, V.E.; Lefer, A.M.; Nicolas, J.M.; Thomas, A.P.; Lefer, D.J.; Vinten-Johansen, J. Physiological concentrations of nitric oxide do not elicit an acute negative inotropic effect in unstimulated cardiac muscle. Circ. Res. 1994, 75, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Nawrath, H.; Baumner, D.; Rupp, J.; Oelert, H. The ineffectiveness of the NO-cyclic GMP signaling pathway in the atrial myocardium. Br. J. Pharmacol. 1995, 116, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Wegener, J.W.; Godecke, A.; Schrader, J.; Nawrath, H. Effects of nitric oxide donors on cardiac contractility in wild-type and myoglobin-deficient mice. Br. J. Pharmacol. 2002, 136, 415–420. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dittrich, M.; Jurevicius, J.; Georget, M.; Rochais, F.; Fleischmann, B.; Hescheler, J.; Fischmeister, R. Local response of L-type Ca(2+) current to nitric oxide in frog ventricular myocytes. J. Physiol 2001, 534, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Fischmeister, R.; Castro, L.R.; Abi-Gerges, A.; Rochais, F.; Jurevicius, J.; Leroy, J.; Vandecasteele, G. Compartmentation of cyclic nucleotide signaling in the heart: The role of cyclic nucleotide phosphodiesterases. Circ. Res. 2006, 99, 816–828. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.R.; Schittl, J.; Fischmeister, R. Feedback control through cGMP-dependent protein kinase contributes to differential regulation and compartmentation of cGMP in rat cardiac myocytes. Circ. Res. 2010, 107, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Airhart, N.; Yang, Y.F.; Roberts, C.T., Jr.; Silberbach, M. Atrial natriuretic peptide induces natriuretic peptide receptor-cGMP-dependent protein kinase interaction. J. Biol. Chem. 2003, 278, 38693–38698. [Google Scholar] [CrossRef] [PubMed]
- Stratton, R.C.; Squires, P.E.; Green, A.K. ANP stimulates hepatocyte Ca2+ efflux via plasma membrane recruitment of PKGIα. Biochem. Biophys. Res. Commun. 2008, 368, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, E.; Belardi, D.; Tocchetti, C.G.; Vahebi, S.; Cormaci, G.; Ketner, E.A.; Moens, A.L.; Champion, H.C.; Kass, D.A. Compartmentalization of cardiac β-adrenergic inotropy modulation by phosphodiesterase type 5. Circulation 2007, 115, 2159–2167. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, T.; Zhang, M.; Hsu, S.; Takimoto, E.; Kass, D.A. Sustained soluble guanylate cyclase stimulation offsets nitric-oxide synthase inhibition to restore acute cardiac modulation by sildenafil. J. Pharmacol. Exp. Ther. 2008, 326, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.I.; Vahebi, S.; Tocchetti, C.G.; Barouch, L.A.; Solaro, R.J.; Takimoto, E.; Kass, D.A. PDE5A suppression of acute β-adrenergic activation requires modulation of myocyte β-3 signaling coupled to PKG-mediated troponin I phosphorylation. Basic Res. Cardiol. 2010, 105, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.I.; Zhu, G.; Sasaki, T.; Cho, G.-S.; Hamdani, N.; Holewinski, R.; Jo, S.-H.; Danner, T.; Zhang, M.; Rainer, P.P.; et al. Phosphodiesterase 9A controls nitric-oxide-independent cGMP and hypertrophic heart disease. Nature 2015, 519, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.K.; Sprenger, J.U.; Steinbrecher, J.H.; Hubscher, D.; Lehnart, S.E.; Abesser, M.; Schuh, K.; El-Armouche, A.; Nikolaev, V.O. Microdomain switch of cGMP-regulated phosphodiesterases leads to ANP-induced augmentation of β-adrenoceptor-stimulated contractility in early cardiac hypertrophy. Circ. Res. 2015, 116, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, J.U.; Perera, R.K.; Steinbrecher, J.H.; Lehnart, S.E.; Maier, L.S.; Hasenfuss, G.; Nikolaev, V.O. In vivo model with targeted cAMP biosensor reveals changes in receptor-microdomain communication in cardiac disease. Nat. Commun. 2015, 6, 6965. [Google Scholar] [CrossRef] [PubMed]
- Morgado, M.; Cairrão, E.; Santos-Silva, A.J.; Verde, I. Cyclic nucleotide-dependent relaxation pathways in vascular smooth muscle. Cell. Mol. Life Sci. 2012, 69, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Sausbier, M.; Schubert, R.; Voigt, V.; Hirneiss, C.; Pfeifer, A.; Korth, M.; Kleppisch, T.; Ruth, P.; Hofmann, F. Mechanisms of NO/cGMP-Dependent Vasorelaxation. Circ. Res. 2000, 87, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Murad, F. Regulation of Cytosolic Guanylyl Cyclase by Nitric Oxide: The NO-Cyclic GMP Signal Transduction System. In Advances in Pharmacology; Murad, F., Ed.; Academic Press: Cambridge, MA, USA, 1994; Volume 26, pp. 19–33. [Google Scholar]
- Piggott, L.A.; Hassell, K.A.; Berkova, Z.; Morris, A.P.; Silberbach, M.; Rich, T.C. Natriuretic peptides and nitric oxide stimulate cGMP synthesis in different cellular compartments. J. Gen. Physiol. 2006, 128, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Feiteiro, J.; Verde, I.; Cairrao, E. Cyclic guanosine monophosphate compartmentation in human vascular smooth muscle cells. Cell. Signal. 2016, 28, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Surapisitchat, J.; Jeon, K.I.; Yan, C.; Beavo, J.A. Differential regulation of endothelial cell permeability by cGMP via phosphodiesterases 2 and 3. Circ. Res. 2007, 101, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Spitzl, A.; Mathes, D.; Nikolaev, V.O.; Werner, F.; Weirather, J.; Spiranec, K.; Rock, K.; Fischer, J.W.; Kammerer, U.; et al. Endothelial Actions of ANP Enhance Myocardial Inflammatory Infiltration in the Early Phase After Acute Infarction. Circ. Res. 2016, 119, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, V.O.; Gambaryan, S.; Engelhardt, S.; Walter, U.; Lohse, M.J. Real-time monitoring of the PDE2 activity of live cells: Hormone-stimulated cAMP hydrolysis is faster than hormone-stimulated cAMP synthesis. J. Biol. Chem. 2005, 280, 1716–1719. [Google Scholar] [CrossRef] [PubMed]
- MacKenna, D.; Summerour, S.R.; Villarreal, F.J. Role of mechanical factors in modulating cardiac fibroblast function and extracellular matrix synthesis. Cardiovasc. Res. 2000, 46, 257–263. [Google Scholar] [CrossRef]
- Huntley, B.K.; Sandberg, S.M.; Noser, J.A.; Cataliotti, A.; Redfield, M.M.; Matsuda, Y.; Burnett, J.C., Jr. BNP-induced activation of cGMP in human cardiac fibroblasts: Interactions with fibronectin and natriuretic peptide receptors. J. Cell. Physiol. 2006, 209, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Souders, C.A.; Bowers, S.L.K.; Baudino, T.A. Cardiac Fibroblast. Circ. Res. 2009, 105, 1164. [Google Scholar] [CrossRef] [PubMed]
- Smolenski, A.; Schultess, J.; Danielewski, O.; Garcia Arguinzonis, M.I.; Thalheimer, P.; Kneitz, S.; Walter, U.; Lohmann, S.M. Quantitative analysis of the cardiac fibroblast transcriptome—Implications for NO/cGMP signaling. Genomics 2004, 83, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, Å.B.; Brunton, L.L. Attenuation of cAMP accumulation in adult rat cardiac fibroblasts by IL-1β and NO: Role of cGMP-stimulated PDE2. Am. J. Physiol. Cell Physiol. 2002, 283, C463–C471. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huntley, B.K.; Ichiki, T.; Sangaralingham, S.J.; Chen, H.H.; Burnett, J.C., Jr. B-type natriuretic peptide and extracellular matrix protein interactions in human cardiac fibroblasts. J. Cell. Physiol. 2010, 225, 251–255. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Method | Advantages/Disadvantages | References |
|---|---|---|
| Traditional biochemical methods | ||
| Immunohistochemistry | Quite sensitive and specific | [64,65,66,67] |
| Radioimmunoassays | Cell destructive assays | |
| Enzyme-linked immunoassays | Only measure total cGMP levels | |
| Immunoblots for PKG substrate phosphorylation | Often require PDE inhibition to obtain adequate sensitivity | |
| Real-time cGMP detection | ||
| Electrophysiological recordings of CNG ion channels | High temporal resolution Technically challenging and time consuming Temporal resolution limited to subsarcolemmal microdomains | [69,70,71] |
| Förster resonance energy transfer (FRET) based cGMP sensors CGY-Del1 Cynget-1/2 cGES-DE2/5 cGi-500/3000/6000 red cGES-DE5 | High temporal and spatial resolution Sensitivity for cGMP measurements in some cell types challenging cGMP/cAMP selectivity important | [77,78] [68,79] [77,80] [81,82,83] [63,84,85] |
| Non-FRET based cGMP sensors | Good cGMP sensitivity | [86,87] |
| FlincG1-3 | Relatively low cGMP/cAMP selectivity |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bork, N.I.; Nikolaev, V.O. cGMP Signaling in the Cardiovascular System—The Role of Compartmentation and Its Live Cell Imaging. Int. J. Mol. Sci. 2018, 19, 801. https://doi.org/10.3390/ijms19030801
Bork NI, Nikolaev VO. cGMP Signaling in the Cardiovascular System—The Role of Compartmentation and Its Live Cell Imaging. International Journal of Molecular Sciences. 2018; 19(3):801. https://doi.org/10.3390/ijms19030801
Chicago/Turabian StyleBork, Nadja I., and Viacheslav O. Nikolaev. 2018. "cGMP Signaling in the Cardiovascular System—The Role of Compartmentation and Its Live Cell Imaging" International Journal of Molecular Sciences 19, no. 3: 801. https://doi.org/10.3390/ijms19030801
APA StyleBork, N. I., & Nikolaev, V. O. (2018). cGMP Signaling in the Cardiovascular System—The Role of Compartmentation and Its Live Cell Imaging. International Journal of Molecular Sciences, 19(3), 801. https://doi.org/10.3390/ijms19030801