Integrins Were Involved in Soybean Agglutinin Induced Cell Apoptosis in IPEC-J2




Abstract

1. Introduction
2. Results
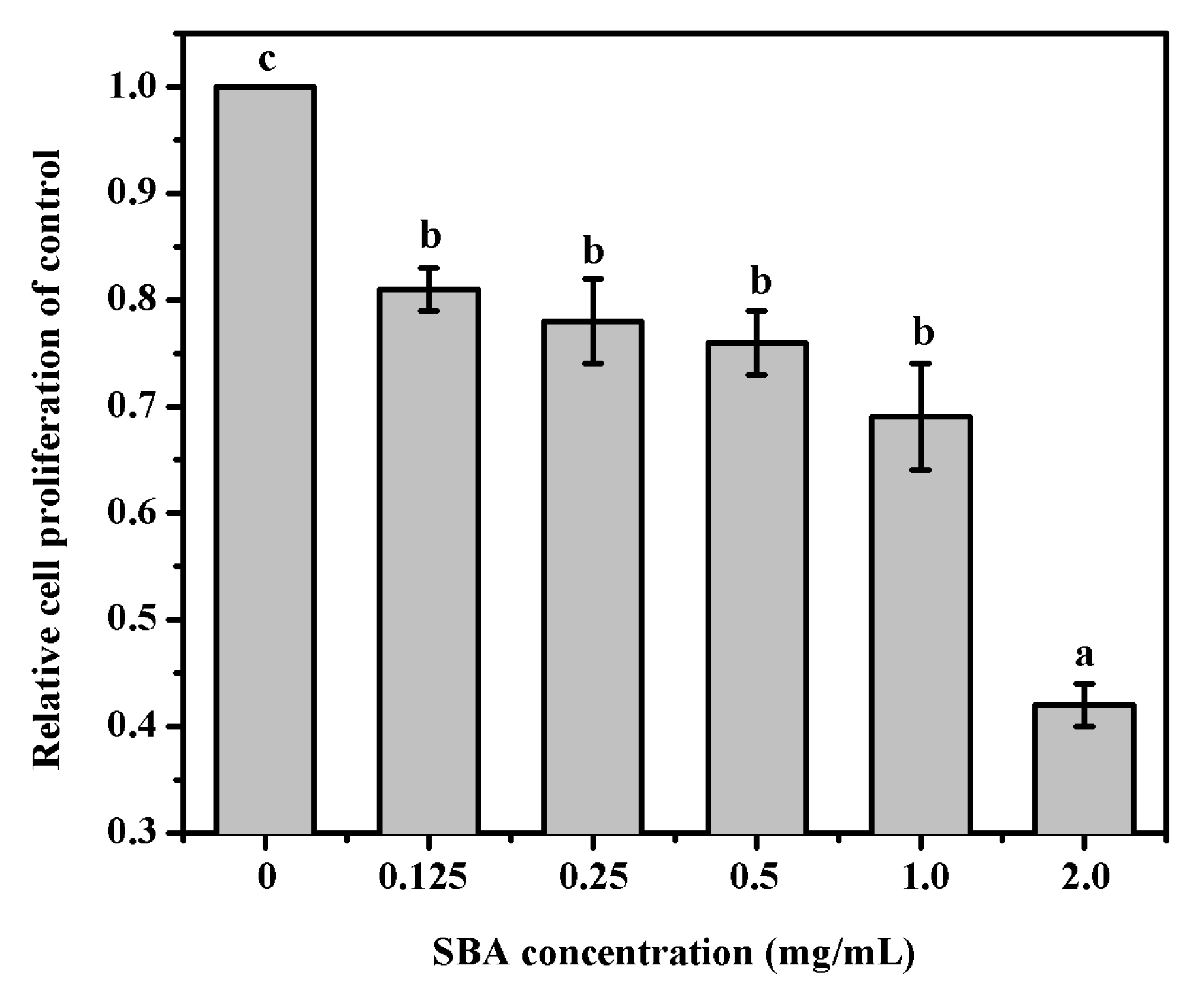
2.1. SBA Decreased IPEC-J2 Cell Proliferation Detected by 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
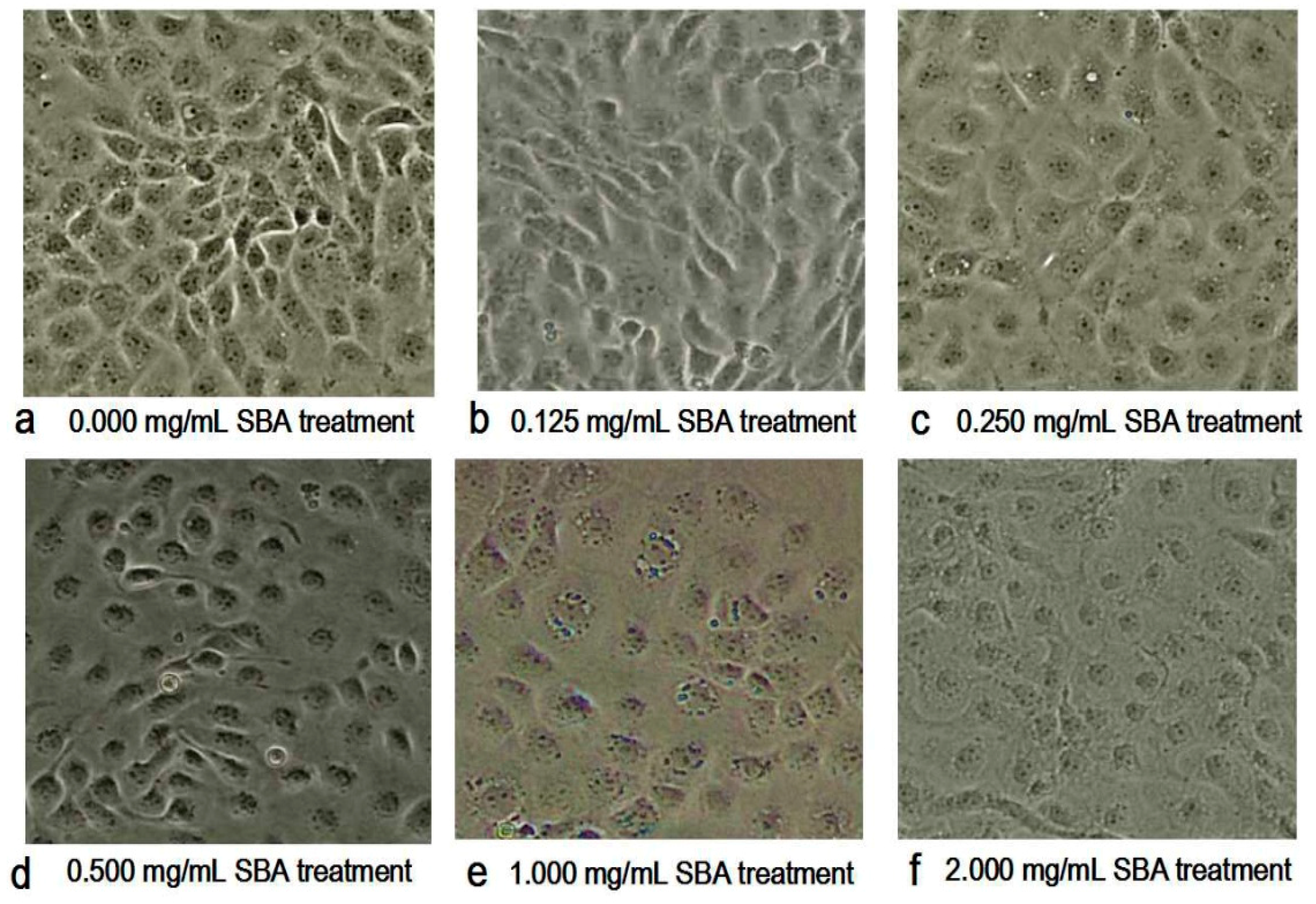
2.2. Morphometric Analysis by Contrast Microscopy
2.3. SBA Induced IPEC-J2 Cell Apoptosis
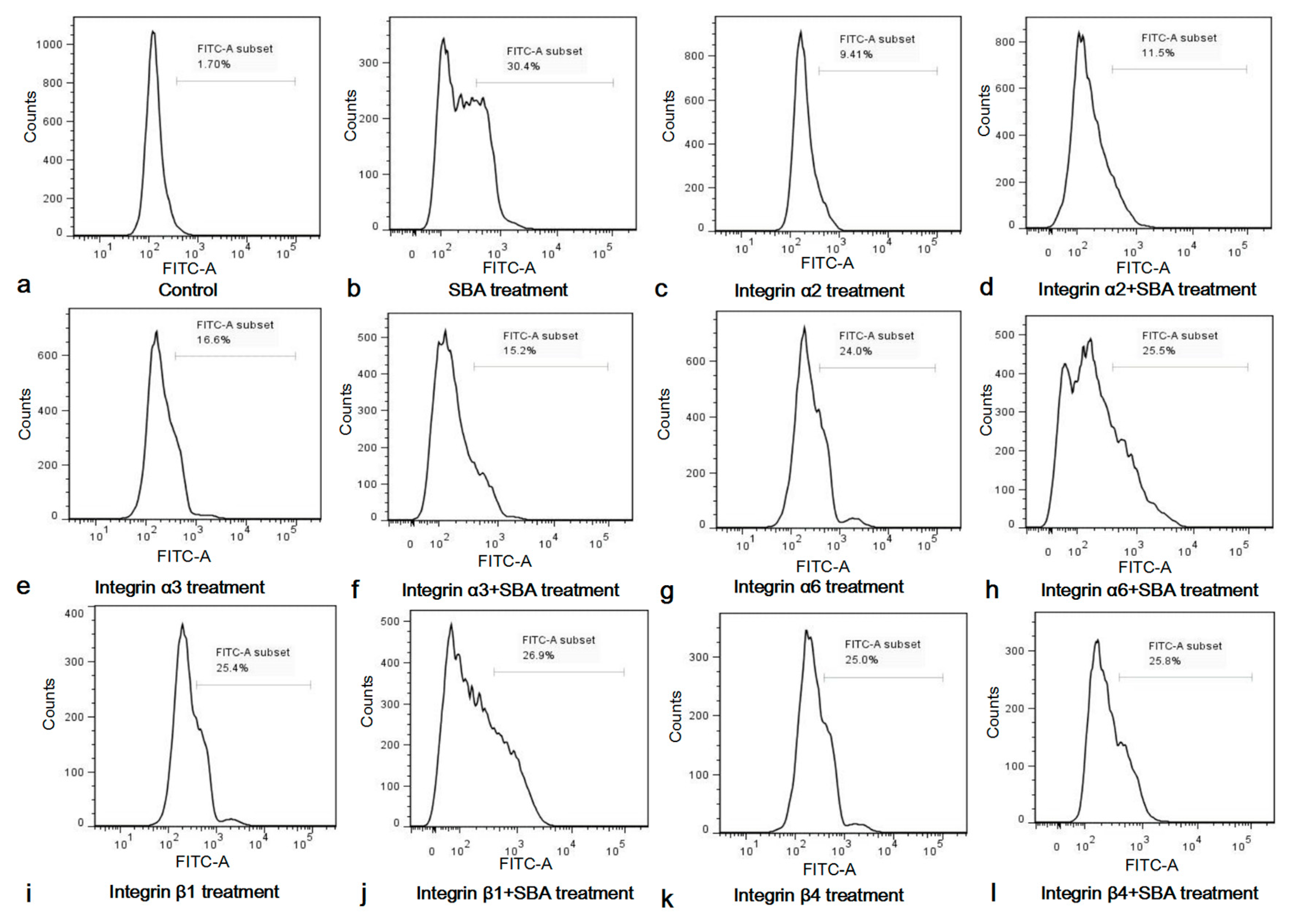
2.4. The Integrins Were Involved in SBA-Induced IPEC-J2 Cell Apoptosis
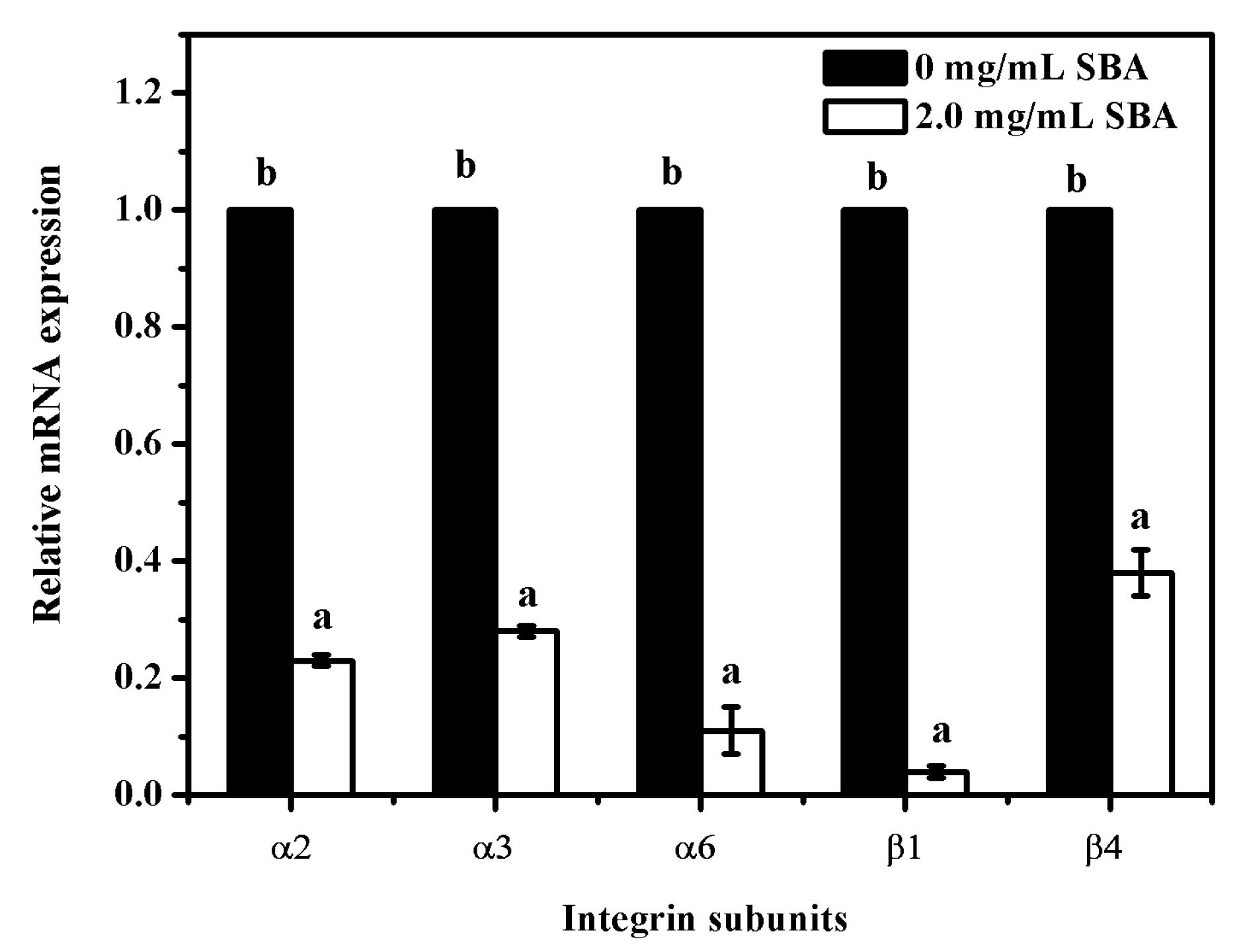
2.5. SBA Lowered the mRNA Expression of Integrins Detected by qRT-PCR
2.6. Identification of the Connection Type between SBA and Integrins in IPEC-J2
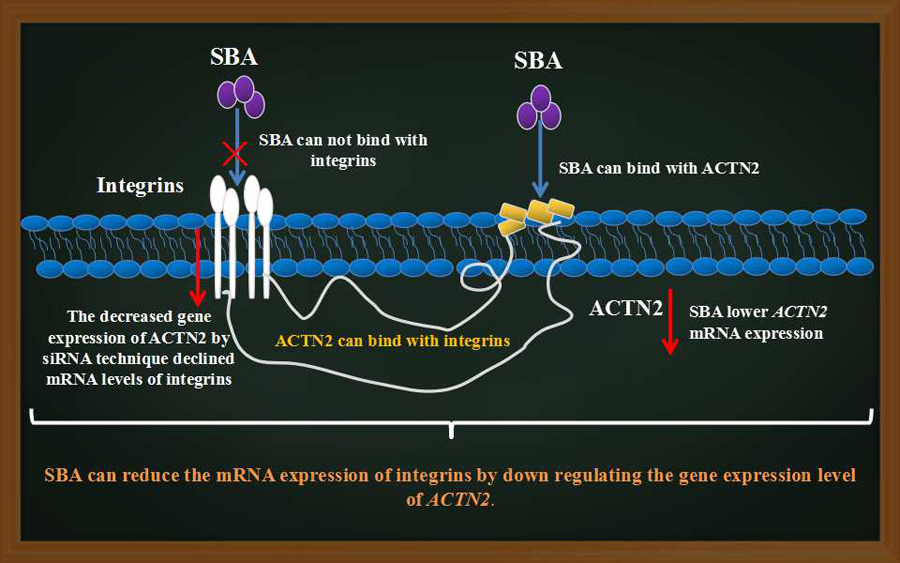
2.7. The Relationship between SBA, Integrins and ACTN2
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Morphological Observation
4.3. Cell Proliferation Assays
4.4. Cell Apoptosis Assessment by Flow Cytometry
4.5. qRT-PCR
4.6. Integrin Functional Inhibition Test
4.7. Effects of SBA on Integrin Gene Expression by qRT-PCR
4.8. Preparation of Cell Membrane Protein Samples
4.9. SDS-PAGE
4.10. Co-Immunoprecipitation
4.11. Liquid Chromatography Coupled to Tandem Mass Spectrometry (LC-MS-MS (Q-E)) Detection
4.12. In Vitro Gene Silencing of ACTN2
4.13. Determination of the Interaction between SBA, ACTN2 and Integrins from the Gene Levels by qRT-PCR
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACTN2 | α-Actinin-2 |
| FCM | Flow cytometry |
| IPEC-J2 | Porcine intestinal columnar epithelial cells |
| LC-MS-MS | Liquid chromatography coupled to tandem mass spectrometry |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NCA | Normal cell apoptosis |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| SBA | Soybean agglutinin |
| SCA | SBA-induced cell apoptosis |
References
- Halder, S.; Surolia, A.; Mukhopadhyay, C. Dynamics simulation of soybean agglutinin (SBA) dimer reveals the impact of glycosylation on its enhanced structural stability. Carbohydr. Res. 2016, 428, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Jindal, S.; Soni, G.; Singh, R. Biochemical and histopathological studies in albino rats fed on soybean lectin. Nutr. Rep. Int. 1984, 29, 95–106. [Google Scholar]
- Pan, L.; Qin, G.; Zhao, Y.; Wang, J.; Liu, F.; Che, D. Effects of soybean agglutinin on mechanical barrier function and tight junction protein expression in intestinal epithelial cells from piglets. Int. J. Mol. Sci. 2013, 14, 21689–21704. [Google Scholar] [CrossRef] [PubMed]
- De Boeck, H.; Lis, H.; van Tilbeurgh, H.; Sharon, N.; Loontiens, F.G. Binding of simple carbohydrates and some of their chromophoric derivatives to soybean agglutinin as followed by titrimetric procedures and stopped flow kinetics. J. Biol. Chem. 1984, 259, 7067–7074. [Google Scholar] [PubMed]
- Horowitz, M. Gastrointestinal glycoproteins. In The Glycoconjugates; Horowitz, M., Pigman, W., Eds.; Academic Press: New York, NY, USA, 2012; pp. 189–213. [Google Scholar]
- Babot, J.D.; Argañaraz-Martínez, E.; Lorenzo-Pisarello, M.J.; Apella, M.C.; Perez Chaia, A. Cytotoxic damage of soybean agglutinin on intestinal epithelial cells of broiler chicks: in vitro protection by Bifidobacterium infantis CRL1395. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qin, G.; Sun, Z.; Che, D.; Bao, N.; Zhang, X. Effects of soybean agglutinin on intestinal barrier permeability and tight junction protein expression in weaned piglets. Int. J. Mol. Sci. 2011, 12, 8502–8512. [Google Scholar] [CrossRef] [PubMed]
- Srichai, M.; Zent, R. Integrin structure and function. In Cell-Extracellular Matrix Interactions in Cancer; Zent, R., Pozzi, A., Eds.; Springer Science + Business Media, LLC: New York, NY, USA, 2010; pp. 19–41. [Google Scholar]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Lee, S.; Qiao, J.; Paul, P.; Chung, D.H. Integrin β1 is critical for gastrin-releasing peptide receptor-mediated neuroblastoma cell migration and invasion. Surgery 2013, 154, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.R.; Felgueiras, J.; Fardilha, M. Signaling pathways in anchoring junctions of epithelial cells: Cell-to-cell and cell-to-extracellular matrix interactions. J. Recept. Signal Transduct. Res. 2015, 35, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Layseca, P.; Streuli, C.H. Signalling pathways linking integrins with cell cycle progression. Matrix Biol. 2014, 34, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Eberwein, P.; Laird, D.; Schulz, S.; Reinhard, T.; Steinberg, T.; Tomakidi, P. Modulation of focal adhesion constituents and their down-stream events by EGF: On the cross-talk of integrins and growth factor receptors. Biochim. Biophys. Acta 2015, 1853, 2183–2198. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhao, Y.; Yuan, Z.; Qin, G. Research advances on structure and biological functions of integrins. SpringerPlus 2016, 5, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhao, Y.; Yuan, Z.; Farouk, M.H.; Zhang, S.; Bao, N.; Qin, G. The integrins involved in soybean agglutinin-induced cell cycle alterations in IPEC-J2. Mol. Cells 2017, 40, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Eldstrom, J.; Choi, W.S.; Steele, D.F.; Fedida, D. SAP97 increases Kv1.5 currents through an indirect N-terminal mechanism. FEBS Lett. 2003, 547, 205–211. [Google Scholar]
- Bakke-McKellep, A.; Penn, M.; Chikwati, E.; Hage, E.; Cai, C.; Krogdahl, A. In Vitro Effects of Various Anti-Nutritional Factors on Intestinal Glucose Absorption and Histology in Atlantic Salmon; Book of Abstracts from XIII ISFNF-International Symposium on Fish Nutr Feeding: Florianopolis, Brazil, 2008; pp. 1–5. [Google Scholar]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Luft, S.; Arrizabalaga, O.; Kulish, I.; Nasonova, E.; Durante, M.; Ritter, S.; Schroeder, I.S. Ionizing radiation alters human embryonic stem cell properties and differentiation capacity by diminishing the expression of activin receptors. Stem Cells Dev. 2017, 26, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, D.; Ishihara, N.; Bujo, H.; Shirai, K.; Tatsuno, I. Effects of serotonin on expression of the LDL receptor family member LR11 and 7-ketocholesterol-induced apoptosis in human vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2014, 446, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Martinou, J.C.; Youle, R.J. Mitochondria in apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev. Cell 2011, 21, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Kalashnikova, I.; Mazar, J.; Neal, C.J.; Rosado, A.L.; Das, S.; Westmoreland, T.J.; Seal, S. Nanoparticle delivery of curcumin induces cellular hypoxia and ROS-mediated apoptosis via modulation of Bcl-2/Bax in human neuroblastoma. Nanoscale 2017, 9, 10375–10387. [Google Scholar] [CrossRef] [PubMed]
- Pusztai, A.; Ewen, S.W.B.; Grant, G.; Peumans, W.J.; van Damme, E.J.M.; Rubio, L.; Bardocz, S. Relationship between survival and binding of plant lectins during small intestinal passage and their effectiveness as growth factors. Digestion 1990, 46, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Hadari, Y.R.; Arbel-Goren, R.; Levy, Y.; Amsterdam, A.; Alon, R.; Zakut, R.; Zick, Y. Galectin-8 binding to integrins inhibits cell adhesion and induces apoptosis. J. Cell Sci. 2000, 113, 2385–2397. [Google Scholar] [PubMed]
- Kabir, S.R.; Reza, M.A. Antibacterial activity of Kaempferia rotunda Rhizome lectin and its induction of apoptosis in Ehrlich Ascites carcinoma cells. Appl. Biochem. Biotechnol. 2014, 172, 2866–2876. [Google Scholar] [CrossRef] [PubMed]
- Gilcrease, M.Z. Integrin signaling in epithelial cells. Cancer Lett. 2007, 247. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Su, L.; Yin, D.; Sun, C.; Zhao, J.; Zhang, S.; Miao, J. Knockdown of integrin β4 in primary cultured mouse neurons blocks survival and induces apoptosis by elevating NADPH oxidase activity and reactive oxygen species level. Int. J. Biochem. Cell Biol. 2008, 40, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Morita-Fujimura, Y.; Tokitake, Y.; Matsui, Y. Heterogeneity of mouse primordial germ cells reflecting the distinct status of their differentiation, proliferation and apoptosis can be classified by the expression of cell surface proteins integrin α6 and c-Kit. Dev. Growth Differ. 2009, 51, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Jeanes, A.I.; Wang, P.; Moreno-Layseca, P.; Paul, N.; Cheung, J.; Tsang, R.; Akhtar, N.; Foster, F.M.; Brennan, K.; Streuli, C.H. Specific β-containing integrins exert differential control on proliferation and two-dimensional collective cell migration in mammary epithelial cells. J. Biol. Chem. 2012, 287, 24103–24112. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.M.; Yang, L.; Wang, Z.; Liu, Y.W.; Fan, J.Z.; Fan, J.; Liu, J.; Luo, Z.J. Overexpression of integrin a2 promotes osteogenic differentiation of hBMSCs from senile osteoporosis through the ERK pathway. Int. J. Clin. Exp. Pathol. 2013, 6, 841–852. [Google Scholar] [PubMed]
- Wang, Y.; Shenouda, S.; Baranwal, S.; Rathinam, R.; Jain, P.; Bao, L.; Hazari, S.; Dash, S.; Alahari, S.K. Integrin subunits alpha5 and alpha6 regulate cell cycle by modulating the chk1 and Rb/E2F pathways to affect breast cancer metastasis. Mol. Cancer 2011, 10, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Mercurio, A.M. Contributions of the α6 integrins to breast carcinoma survival and progression. Mol. Cells 2004, 17, 203–209. [Google Scholar] [PubMed]
- Dowling, J.; Yu, Q.C.; Fuchs, E. Beta4 integrin is required for hemidesmosome formation, cell adhesion and cell survival. J.Cell Biol. 1996, 134, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Kaltner, H.; Stierstorfer, B. Animal lectins as cell adhesion molecules. Cells Tissues Organs 1998, 161, 162–179. [Google Scholar] [CrossRef]
- Maxwell, J. Soybeans: Cultivation, Uses and Nutrition; Nova Science Publishers, Inc.: New York, NY, USA, 2011. [Google Scholar]
- Burridge, K.; McCullough, L. The association of α-actinin with the plasma membrane. J. Supramol. Struct. 1980, 13, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Otey, C.A.; Pavalko, F.M.; Burridge, K. An interaction between α-actinin and the beta 1 integrin subunit in vitro. J. Cell Biol. 1990, 111, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Yu, Y.; Chi, X.; Xu, W.; Lu, D.; Song, Y.; Zhang, Y.; Zhang, H. Kindlin-2 interacts with α-actinin-2 and β1 integrin to maintain the integrity of the Z-disc in cardiac muscles. FEBS Lett. 2015, 589, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Tadokoro, S.; Nakazawa, T.; Kamae, T.; Kiyomizu, K.; Kashiwagi, H.; Honda, S.; Kanakura, Y.; Tomiyama, Y. A potential role for α-actinin in inside-out αIIbβ3 signaling. Blood 2011, 117, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Roca-Cusachs, P.; del Rio, A.; Puklin-Faucher, E.; Gauthier, N.C.; Biais, N.; Sheetz, M.P. Integrin-dependent force transmission to the extracellular matrix by α-actinin triggers adhesion maturation. Proc. Nat. Acad. Sci. 2013, 110, E1361–E1370. [Google Scholar] [CrossRef] [PubMed]
- Van der Flier, A.; Sonnenberg, A. Function and interactions of integrins. Cell Tissue Res. 2001, 305, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Che, D.; Qin, G.; Kong, X.; Farouk, M.H. Effects of dietary non-fiber carbohydrates on composition and function of gut microbiome in monogastrics: A review. Protein Pept. Lett. 2017, 24, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M. Comparative Study on Digestive Dynamics and Antinutritional Effects of Soybean Agglutinin in Different Species of Animals; Jilin Agricultural University: Changchun, China, 2007. [Google Scholar]
- Li, Z.; Li, D.; Qiao, S.; Zhu, X.; Huang, C. Anti-nutritional effects of a moderate dose of soybean agglutinin in the rat. Arch. Anim. Nutr. 2003, 57, 267–277. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef]
- SPSS Inc. SPSS Advanced Statistics 16 Update; SPSS Inc.: Chicago, IL, USA, 2008. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Name | Accession No. | Score | mw/PI | Sequence Coverage |
|---|---|---|---|---|---|
| 1 | KRT1 | F1SGG3 | 1973 | 65.381/8.18 | 14% |
| 2 | KRT14 | F1S0L1 | 1309 | 51.811/4.94 | 40% |
| 3 | KRT10 | I3LDS3 | 1265 | 58.243/4.93 | 17% |
| 4 | KRT5 | F1SGG6 | 870 | 63.668/7.74 | 20% |
| 5 | KRT75 | F1SGI7 | 697 | 65.354/9.27 | 12% |
| 6 | KRT84 | F1SGI2 | 619 | 54.121/6.12 | 5% |
| 7 | LOC100737483 | F1SGG9 | 564 | 60.375/8.53 | 19% |
| 8 | MYH13 | 71SS66 | 550 | 223.705/5.54 | 1% |
| 9 | KRT2A | A5A759 | 539 | 66.510/7.47 | 8% |
| 10 | KRT3 | I3LDM6 | 525 | 65.497/8.03 | 11% |
| 11 | PYGM | F1RQQ8 | 507 | 97.665/6.65 | 7% |
| 12 | LOC100302368 | F1SRS2 | 464 | 26.563/6.85 | 26% |
| 13 | KRT8 | F1SGG2 | 440 | 54.394/5.70 | 7% |
| 14 | CKM | I3LBJ8 | 411 | 43.114/6.91 | 10% |
| 15 | ALB | F1RUN2 | 409 | 71.621/5.98 | 8% |
| 16 | Jup | Q8WNW3 | 399 | 82.539/5.75 | 24% |
| 17 | ACTA1 | P68137 | 368 | 42.366/5.23 | 17% |
| 18 | KRT15 | F1S0K2 | 317 | 49.501/4.87 | 8% |
| 19 | l-lactate dehydrogenase | D2SW96 | 275 | 36.582/8.18 | 6% |
| 20 | Myosin-4 | Q9TV62 | 245 | 224.0.1/5.60 | 2% |
| 21 | LOC100519531 | F1RU49 | 174 | 103.873/5.31 | 5% |
| 22 | SLC25A4 | F1RZQ6 | 173 | 33.233/9.82 | 14% |
| 23 | GFAP | F1RR02 | 166 | 49.463/5.32 | 4% |
| 24 | HUMMLC2B | A1XQT5 | 143 | 19.066/4.82 | 10% |
| 25 | CS | Q0QEK1 | 142 | 24.886/6.59 | 5% |
| 26 | ACTC1 | B6VNT8 | 141 | 42.334/5.23 | 18% |
| 27 | MYH4 | Q9TV62 | 139 | 224.010/5.60 | 1% |
| 28 | DSP | F1RW75 | 129 | 310.122/6.49 | 4% |
| 29 | LOC407246 | I3LL80 | 108 | 36.884/5.57 | 11% |
| 30 | LOC100515788 | F1RII5 | 107 | 15.682/7.93 | 6% |
| 31 | ATP2A2 | P11607 | 93 | 116.371/5.23 | 3% |
| 32 | PCBP1 | I3LEC2 | 93 | 37.991/6.66 | 5% |
| 33 | LDHB | F1SR05 | 89 | 37.108/5.45 | 5% |
| 34 | GAPDH | G3CKJ2 | 87 | 29.621/9.29 | 8% |
| 35 | MB | P02189 | 85 | 17.074/6.75 | 10% |
| 36 | Uncharacterized protein | F1SIX1 | 84 | 17.814/5.82 | 15% |
| 37 | PKM | F1SHL9 | 82 | 59.256/7.95 | 2% |
| 38 | HUMMLC2B | Q5EEL9 | 81 | 10.704/4.80 | 16% |
| 39 | AK1 | P00571 | 75 | 21.739/8.38 | 6% |
| 40 | BLVRB | I3LQH7 | 75 | 22.400/6.35 | 10% |
| 41 | TMEM38A | F1S9V9 | 74 | 29.180/8.79 | 5% |
| 42 | tnnt3 | Q75NG6 | 70 | 29.811/7.74 | 6% |
| 43 | MYH7 | F1S9D6 | 67 | 223.795/5.59 | 0% |
| 44 | HUMMLC2B | Q5EEL9 | 59 | 10.704/4.80 | 16% |
| 45 | YWHAG | F2Z4Z1 | 55 | 28.456/4.80 | 13% |
| 46 | TPI1 | D0G7F6 | 51 | 26.879/6.54 | 5% |
| 47 | SOD2 | F1SB60 | 49 | 24.923/8.59 | 9% |
| 48 | Uncharacterized protein | F1RWF5 | 49 | 28.518/5.50 | 7% |
| 49 | MLC2V | A1XQV9 | 45 | 16.862/4.57 | |
| 50 | ENO2 | I3LCN1 | 44 | 47.438/4.93 | 5% |
| 51 | LOC100621642 | I3LJ52 | 39 | 28.128/6.69 | 3% |
| 52 | ANXA2 | P19620 | 38 | 38.795/6.49 | 13% |
| 53 | KRT18 | A8D349 | 37 | 18.038/4.63 | 10% |
| 54 | PCBP2 | F1SFQ4 | 37 | 37.606/6.60 | 5% |
| 55 | Beta-actin | A0SNU5 | 37 | 29.678/5.50 | 11% |
| 56 | PKP1 | I3LGN8 | 36 | 53.569/9.12 | 6% |
| 57 | ACTN2 | F1RHL9 | 34 | 104.237/5.31 | 2% |
| 58 | Glyceraldehyde-3-phosphate dehydrogenase | A0SNU7 | 34 | 28.035/6.77 | 5% |
| 59 | HADH | P00348 | 32 | 34.197/9/02 | 11% |
| 60 | PFMK | Q1HL06 | 30 | 82.443/8.57 | 2% |
| 61 | ANK2 | F1S146 | 28 | 428.715/4.93 | 0% |
| 62 | CA3 | Q5S1S4 | 28 | 29.679/7.72 | 9% |
| 63 | ATP2A1 | F1RFH9 | 27 | 110.403/5.17 | 3% |
| 64 | METTL3 | F1S8J8 | 26 | 65.501/6.12 | 1% |
| 65 | CABLES1 | F1SBB7 | 26 | 64.246/9.61 | 1% |
| 66 | CKMT2 | Q2HYU1 | 25 | 47.961/8.47 | 3% |
| 67 | VDAC1 | Q9MZ16 | 24 | 30.882/8.62 | 7% |
| Gene | Accession No. | Forward Primer | Reverse Primer |
|---|---|---|---|
| Bcl-2 | NM_214285.1 | TGTGCGTGGAGAGCGTAGA | TAGGTGGTCATTCAGGTAAGTGG |
| α2 (ITGA2) | NM_001244272 | CGCAGATTACGCTCCTCAAAA | GCTGAACAACAGTCCCACTCC |
| α3 (ITGA3) | XM_005668879 | AGCGTCCCCACCATCAA | CTCACAGCCACAAGCACCA |
| α6 (ITGA6) | XM_005657544 | TGTGACTGTGTTTCCCTCAAAGA | AGCATCAAAATCCCAGCAAGA |
| β1 (ITGB1) | XM_003131195 | TGAGTGCAACCCCAACTACAC | CAGACCCCACATTCACAGACA |
| β4 (ITGB4) | NM_213968.1 | CAGGGCTACAGCGTGGAGTA | TAGGAGTGGTTGGGCAGAAGA |
| ACTN2 | NM_001243666 | GGCATACGGCAAAGAGCA | GGCGTCGTGATAGTCCAGTT |
| GAPDH | NM_001206359 | TGCACCACCAACTGCTTGGC | GGCATGGACCGTGGTCATGAG |
| Treatment Number | SBA Level | Integrin Inhibitor Type |
|---|---|---|
| 1 (control) | - | - |
| 2 | SBA | - |
| 3 | - | α2 inhibitor |
| 4 | SBA | α2 inhibitor |
| 5 | - | α3 inhibitor |
| 6 | SBA | α3 inhibitor |
| 7 | - | α6 inhibitor |
| 8 | SBA | α6 inhibitor |
| 9 | - | β1 inhibitor |
| 10 | SBA | β1 inhibitor |
| 11 | - | β4 inhibitor |
| 12 | SBA | β4 inhibitor |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| ACTN2-siRNA1 | GCACUCAGAUCGAGAACAUTT | AUGUUCUCGAUCUGAGUGCTT |
| ACTN2-siRNA2 | GCAUGAUCUGGACCAUUAUTT | AUAAUGGUCCAGAUCAUGCTT |
| ACTN2-siRNA3 | GGUUCUUGCUGUGAAUCAATT | UUGAUUCACAGCAAGAACCTT |
| NC-siRNA | GACTTCATAAGGCGCATGC | GCATGCGCCTTATGAAGTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, L.; Zhao, Y.; Farouk, M.H.; Bao, N.; Wang, T.; Qin, G. Integrins Were Involved in Soybean Agglutinin Induced Cell Apoptosis in IPEC-J2. Int. J. Mol. Sci. 2018, 19, 587. https://doi.org/10.3390/ijms19020587
Pan L, Zhao Y, Farouk MH, Bao N, Wang T, Qin G. Integrins Were Involved in Soybean Agglutinin Induced Cell Apoptosis in IPEC-J2. International Journal of Molecular Sciences. 2018; 19(2):587. https://doi.org/10.3390/ijms19020587
Chicago/Turabian StylePan, Li, Yuan Zhao, Mohammed Hamdy Farouk, Nan Bao, Tao Wang, and Guixin Qin. 2018. "Integrins Were Involved in Soybean Agglutinin Induced Cell Apoptosis in IPEC-J2" International Journal of Molecular Sciences 19, no. 2: 587. https://doi.org/10.3390/ijms19020587
APA StylePan, L., Zhao, Y., Farouk, M. H., Bao, N., Wang, T., & Qin, G. (2018). Integrins Were Involved in Soybean Agglutinin Induced Cell Apoptosis in IPEC-J2. International Journal of Molecular Sciences, 19(2), 587. https://doi.org/10.3390/ijms19020587