Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi


Abstract
1. Introduction
2. Results
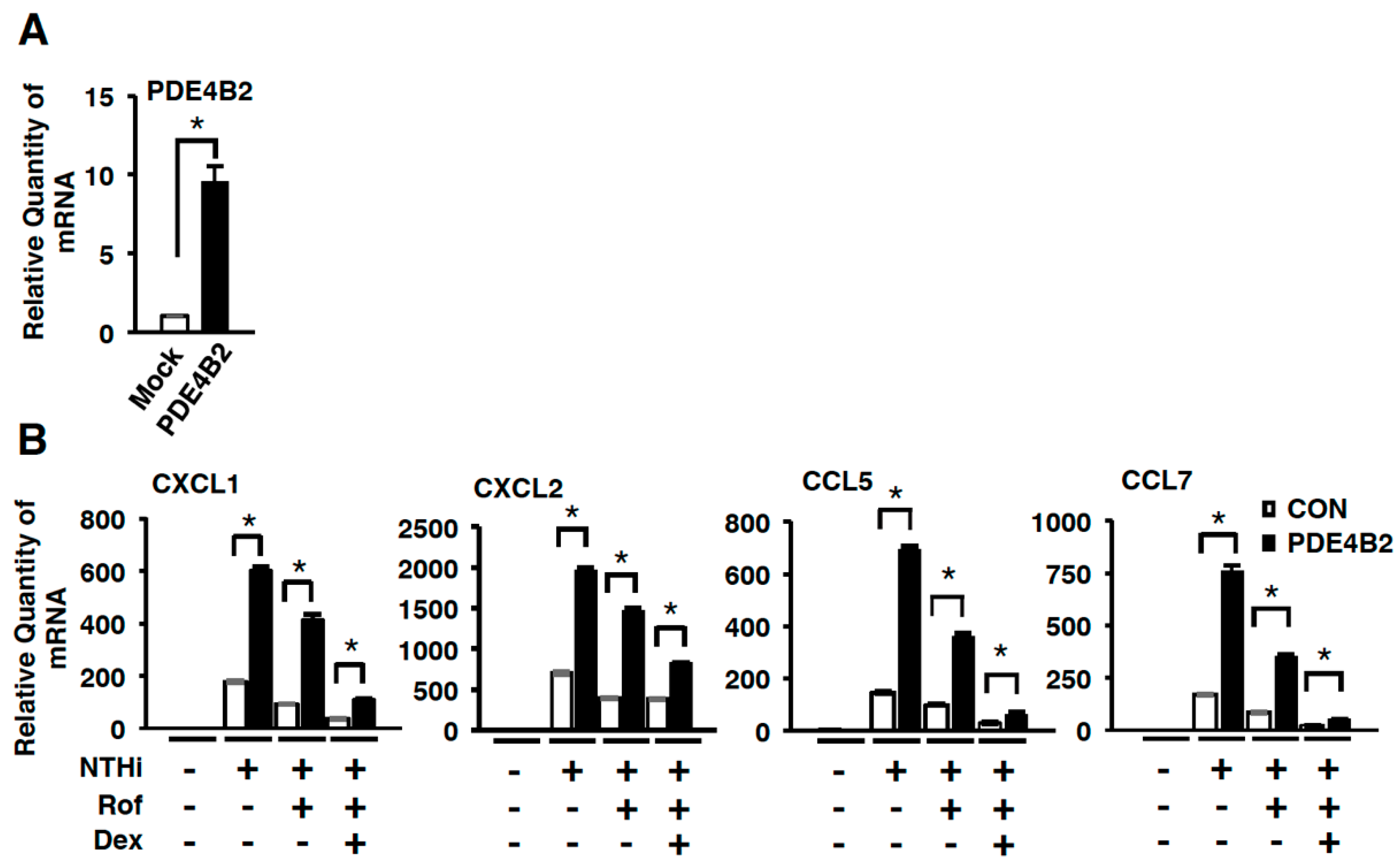
2.1. Dexamethasone Suppresses Synergistic Induction of PDE4B Expression by Roflumilast and NTHi In Vitro and In Vivo
2.2. Dexamethasone Suppresses NTHi-Induced Inflammation In Vitro and Mouse Lung In Vivo
2.3. Dexamethasone Improves the Inhibitory Effect of Roflumilast on NTHi-Induced Inflammation by Suppressing Up-Regulated PDE4B
2.4. Dexamethasone Suppresses PDE4B Expression in a GR-Dependent Manner
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Bacterial Strains and Culture Condition
4.3. Cell Culture
4.4. Real-Time Quantitative and Semi-Quantitative RT-PCR Analyses
4.5. Plasmids and Transfections
4.6. Immunoblotting
4.7. Mice and Animal Experiments
4.8. Histology and Immunofluorescence Assay
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PDE4B | Phosphodiesterase 4B |
| NTHi | Nontypeable Haemophilus influenzae |
| GR | Glucocorticoid receptor |
| COPD | Chronic obstructive pulmonary disease |
| CF | Cystic fibrosis |
| GCs | Glucocorticoids |
| PMN | Polymorphonuclear neutrophils |
| BAL | Bronchoalveolar lavage |
References
- Ollivier, V.; Parry, G.C.; Cobb, R.R.; de Prost, D.; Mackman, N. Elevated cyclic AMP inhibits NF-kappaB-mediated transcription in human monocytic cells and endothelial cells. J. Biol. Chem. 1996, 271, 20828–20835. [Google Scholar] [CrossRef] [PubMed]
- Hertz, A.L.; Bender, A.T.; Smith, K.C.; Gilchrist, M.; Amieux, P.S.; Aderem, A.; Beavo, J.A. Elevated cyclic AMP and PDE4 inhibition induce chemokine expression in human monocyte-derived macrophages. Proc. Natl. Acad. Sci. USA 2009, 106, 21978–21983. [Google Scholar] [CrossRef] [PubMed]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-kappaB action. Cell. Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef] [PubMed]
- Oldenburger, A.; Roscioni, S.S.; Jansen, E.; Menzen, M.H.; Halayko, A.J.; Timens, W.; Meurs, H.; Maarsingh, H.; Schmidt, M. Anti-inflammatory role of the cAMP effectors Epac and PKA: Implications in chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e31574. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Beavo, J. Biochemistry and physiology of cyclic nucleotide phosphodiesterases: Essential components in cyclic nucleotide signaling. Annu. Rev. Biochem. 2007, 76, 481–511. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Mika, D.; Richter, W. Cyclic AMP compartments and signaling specificity: Role of cyclic nucleotide phosphodiesterases. J. Gen. Physiol. 2014, 143, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 58, 488–520. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.K.; Connett, J.; Bailey, W.C.; Casaburi, R.; Cooper, J.A., Jr.; Criner, G.J.; Curtis, J.L.; Dransfield, M.T.; Han, M.K.; Lazarus, S.C.; et al. Azithromycin for prevention of exacerbations of COPD. N. Engl. J. Med. 2011, 365, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, E.; Zlock, L.; Lao, A.; Mika, D.; Namkung, W.; Xie, M.; Scheitrum, C.; Gruenert, D.C.; Verkman, A.S.; Finkbeiner, W.E.; et al. Anchored PDE4 regulates chloride conductance in wild-type and DeltaF508-CFTR human airway epithelia. FASEB J. 2014, 28, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Lipworth, B.J. Phosphodiesterase-4 inhibitors for asthma and chronic obstructive pulmonary disease. Lancet 2005, 365, 167–175. [Google Scholar] [CrossRef]
- Milara, J.; Armengot, M.; Banuls, P.; Tenor, H.; Beume, R.; Artigues, E.; Cortijo, J. Roflumilast N-oxide, a PDE4 inhibitor, improves cilia motility and ciliated human bronchial epithelial cells compromised by cigarette smoke in vitro. Br. J. Pharmacol. 2012, 166, 2243–2262. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Peiro, T.; Serrano, A.; Artigues, E.; Aparicio, J.; Tenor, H.; Sanz, C.; Cortijo, J. Simvastatin Increases the Ability of Roflumilast N-oxide to Inhibit Cigarette Smoke-Induced Epithelial to Mesenchymal Transition in Well-differentiated Human Bronchial Epithelial Cells in vitro. COPD 2015, 12, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Peiro, T.; Serrano, A.; Guijarro, R.; Zaragoza, C.; Tenor, H.; Cortijo, J. Roflumilast N-oxide inhibits bronchial epithelial to mesenchymal transition induced by cigarette smoke in smokers with COPD. Pulm. Pharmacol. Ther. 2014, 28, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Mehats, C.; Tanguy, G.; Dallot, E.; Robert, B.; Rebourcet, R.; Ferre, F.; Leroy, M.J. Selective up-regulation of phosphodiesterase-4 cyclic adenosine 3′,5′-monophosphate (cAMP)-specific phosphodiesterase variants by elevated cAMP content in human myometrial cells in culture. Endocrinology 1999, 140, 3228–3237. [Google Scholar] [CrossRef] [PubMed]
- Campos-Toimil, M.; Keravis, T.; Orallo, F.; Takeda, K.; Lugnier, C. Short-term or long-term treatments with a phosphodiesterase-4 (PDE4) inhibitor result in opposing agonist-induced Ca(2+) responses in endothelial cells. Br. J. Pharmacol. 2008, 154, 82–92. [Google Scholar] [CrossRef] [PubMed]
- D’Sa, C.; Tolbert, L.M.; Conti, M.; Duman, R.S. Regulation of cAMP-specific phosphodiesterases type 4B and 4D (PDE4) splice variants by cAMP signaling in primary cortical neurons. J. Neurochem. 2002, 81, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Hallstrand, T.S.; Hackett, T.L.; Altemeier, W.A.; Matute-Bello, G.; Hansbro, P.M.; Knight, D.A. Airway epithelial regulation of pulmonary immune homeostasis and inflammation. Clin. Immunol. 2014, 151, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Michalski, J.M.; Golden, G.; Ikari, J.; Rennard, S.I. PDE4: A novel target in the treatment of chronic obstructive pulmonary disease. Clin. Pharmacol. Ther. 2012, 91, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Price, D.; Chisholm, A.; Ryan, D.; Crockett, A.; Jones, R. The use of roflumilast in COPD: A primary care perspective. Prim. Care Respir. J. 2010, 19, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Tashkin, D.P. Roflumilast: The new orally active, selective phophodiesterase-4 inhibitor, for the treatment of COPD. Expert Opin. Pharmacother. 2014, 15, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Grootendorst, D.C.; Gauw, S.A.; Verhoosel, R.M.; Sterk, P.J.; Hospers, J.J.; Bredenbroker, D.; Bethke, T.D.; Hiemstra, P.S.; Rabe, K.F. Reduction in sputum neutrophil and eosinophil numbers by the PDE4 inhibitor roflumilast in patients with COPD. Thorax 2007, 62, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Tannheimer, S.L.; Sorensen, E.A.; Haran, A.C.; Mansfield, C.N.; Wright, C.D.; Salmon, M. Additive anti-inflammatory effects of beta 2 adrenoceptor agonists or glucocorticosteroid with roflumilast in human peripheral blood mononuclear cells. Pulm. Pharmacol. Ther. 2012, 25, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Milara, J.; Morell, A.; Ballester, B.; Sanz, C.; Freire, J.; Qian, X.; Alonso-Garcia, M.; Morcillo, E.; Cortijo, J. Roflumilast improves corticosteroid resistance COPD bronchial epithelial cells stimulated with toll like receptor 3 agonist. Respir. Res. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef] [PubMed]
- Rabe, K.F.; Bateman, E.D.; O’Donnell, D.; Witte, S.; Bredenbroker, D.; Bethke, T.D. Roflumilast--an oral anti-inflammatory treatment for chronic obstructive pulmonary disease: A randomised controlled trial. Lancet 2005, 366, 563–571. [Google Scholar] [CrossRef]
- Calverley, P.M.; Sanchez-Toril, F.; McIvor, A.; Teichmann, P.; Bredenbroeker, D.; Fabbri, L.M. Effect of 1-year treatment with roflumilast in severe chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2007, 176, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Vestbo, J.; Tan, L.; Atkinson, G.; Ward, J.; Uk, G.S.T. A controlled trial of 6-weeks’ treatment with a novel inhaled phosphodiesterase type-4 inhibitor in COPD. Eur. Respir. J. 2009, 33, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Susuki-Miyata, S.; Miyata, M.; Lee, B.C.; Xu, H.; Kai, H.; Yan, C.; Li, J.D. Cross-talk between PKA-Cbeta and p65 mediates synergistic induction of PDE4B by roflumilast and NTHi. Proc. Natl. Acad. Sci. USA 2015, 112, E1800–E1809. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.; Sargent, C.; Singh, D. Glucocorticoid sensitivity of lipopolysaccharide-stimulated chronic obstructive pulmonary disease alveolar macrophages. Clin. Exp. Immunol. 2009, 158, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.; Robinson, L.; Lea, S.; Banyard, A.; Blaikley, J.; Ray, D.; Bizzi, A.; Volpi, G.; Facchinetti, F.; Singh, D. Evaluation of glucocorticoid receptor function in COPD lung macrophages using beclomethasone-17-monopropionate. PLoS ONE 2013, 8, e64257. [Google Scholar] [CrossRef] [PubMed]
- Toward, T.J.; Broadley, K.J. Airway function, oedema, cell infiltration and nitric oxide generation in conscious ozone-exposed guinea-pigs: Effects of dexamethasone and rolipram. Br. J. Pharmacol. 2002, 136, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.L.; Milara, J.; Lluch, J.; De Diego, A.; Sanz, C.; Cortijo, J. Phosphodiesterase-4 inhibition improves corticosteroid insensitivity in pulmonary endothelial cells under oxidative stress. Allergy 2013, 68, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Luecke, H.F.; Yamamoto, K.R. The glucocorticoid receptor blocks P-TEFb recruitment by NFkappaB to effect promoter-specific transcriptional repression. Genes Dev. 2005, 19, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Busillo, J.M.; Cidlowski, J.A. The five Rs of glucocorticoid action during inflammation: Ready, reinforce, repress, resolve, and restore. Trends Endocrinol. Metab. 2013, 24, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Ahlstrom, M.; Pekkinen, M.; Huttunen, M.; Lamberg-Allardt, C. Dexamethasone down-regulates cAMP-phosphodiesterase in human osteosarcoma cells. Biochem. Pharmacol. 2005, 69, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; De Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell. Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Tillis, C.C.; Huang, H.W.; Bi, W.; Pan, S.; Bruce, S.R.; Alcorn, J.L. Glucocorticoid regulation of human pulmonary surfactant protein-B (SP-B) mRNA stability is independent of activated glucocorticoid receptor. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, L940–L950. [Google Scholar] [CrossRef] [PubMed]
- Otulakowski, G.; Duan, W.; Sarangapani, A.; Gandhi, S.; O’Brodovich, H. Glucocorticoid-mediated repression of REDD1 mRNA expression in rat fetal distal lung epithelial cells. Pediatr. Res. 2009, 65, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, L.; Liu, B.; Blaxall, B.C.; Taubman, M.B. A novel role for the glucocorticoid receptor in the regulation of monocyte chemoattractant protein-1 mRNA stability. J. Biol. Chem. 2007, 282, 10146–10152. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Conti, M. Induction of the cyclic nucleotide phosphodiesterase PDE4B is essential for LPS-activated TNF-alpha responses. Proc. Natl. Acad. Sci. USA 2002, 99, 7628–7633. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Brooks, J.; Hagan, G.; Cahn, A.; O’Connor, B.J. Superiority of “triple” therapy with salmeterol/fluticasone propionate and tiotropium bromide versus individual components in moderate to severe COPD. Thorax 2008, 63, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Calverley, P.M.; Rabe, K.F.; Goehring, U.M.; Kristiansen, S.; Fabbri, L.M.; Martinez, F.J. M2-124 and M2-125 study groups. Roflumilast in symptomatic chronic obstructive pulmonary disease: Two randomised clinical trials. Lancet 2009, 374, 685–694. [Google Scholar] [CrossRef]
- Donnelly, L.E.; Barnes, P.J. Chemokine receptors as therapeutic targets in chronic obstructive pulmonary disease. Trends Pharmacol. Sci. 2006, 27, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.; Zhou, Y.; Iribarren, P.; Wang, J. Chemokines and chemokine receptors: Their manifold roles in homeostasis and disease. Cell. Mol. Immunol. 2004, 1, 95–104. [Google Scholar] [PubMed]
- Quint, J.K.; Wedzicha, J.A. The neutrophil in chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2007, 119, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Howarth, P.H. A comparison of the anti-inflammatory properties of intranasal corticosteroids and antihistamines in allergic rhinitis. Allergy 2000, 55 (Suppl. 62), 6–11. [Google Scholar] [CrossRef] [PubMed]
- Hidi, R.; Timmermans, S.; Liu, E.; Schudt, C.; Dent, G.; Holgate, S.T.; Djukanovic, R. Phosphodiesterase and cyclic adenosine monophosphate-dependent inhibition of T-lymphocyte chemotaxis. Eur. Respir. J. 2000, 15, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.; Plumb, J.; Kaur, M.; Ray, D.; Singh, D. Additive anti-inflammatory effects of corticosteroids and phosphodiesterase-4 inhibitors in COPD CD8 cells. Respir. Res. 2016, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.J.; Calverley, P.M.; Goehring, U.M.; Brose, M.; Fabbri, L.M.; Rabe, K.F. Effect of roflumilast on exacerbations in patients with severe chronic obstructive pulmonary disease uncontrolled by combination therapy (REACT): A multicentre randomised controlled trial. Lancet 2015, 385, 857–866. [Google Scholar] [CrossRef]
- Kanehiro, A.; Ikemura, T.; Makela, M.J.; Lahn, M.; Joetham, A.; Dakhama, A.; Gelfand, E.W. Inhibition of phosphodiesterase 4 attenuates airway hyperresponsiveness and airway inflammation in a model of secondary allergen challenge. Am. J. Respir. Crit. Care Med. 2001, 163, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.G.; Deng, Y.M.; Wu, X.; Tang, H.F.; Deng, J.F.; Chen, J.Q.; Yang, S.Y.; Xie, Q.M. Inhibition of phosphodiesterase activity, airway inflammation and hyperresponsiveness by PDE4 inhibitor and glucocorticoid in a murine model of allergic asthma. Life Sci. 2006, 79, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Guess, A.; Agrawal, S.; Wei, C.C.; Ransom, R.F.; Benndorf, R.; Smoyer, W.E. Dose- and time-dependent glucocorticoid receptor signaling in podocytes. Am. J. Physiol. Renal. Physiol. 2010, 299, F845–F853. [Google Scholar] [CrossRef] [PubMed]
- Almawi, W.Y.; Melemedjian, O.K. Negative regulation of nuclear factor-kappaB activation and function by glucocorticoids. J. Mol. Endocrinol. 2002, 28, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Haddad, J.J.; Land, S.C.; Tarnow-Mordi, W.O.; Zembala, M.; Kowalczyk, D.; Lauterbach, R. Immunopharmacological potential of selective phosphodiesterase inhibition. I. Differential regulation of lipopolysaccharide-mediated proinflammatory cytokine (interleukin-6 and tumor necrosis factor-alpha) biosynthesis in alveolar epithelial cells. J. Pharmacol. Exp. Ther. 2002, 300, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.A.; Taverna, J.; Chaves, J.; Makkinje, A.; Lerner, A. Phosphodiesterase 4 inhibitors augment levels of glucocorticoid receptor in B cell chronic lymphocytic leukemia but not in normal circulating hematopoietic cells. Clin. Cancer Res. 2007, 13, 4920–4927. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Dong, H.; Kim, E.J.; Weintraub, L.; Epstein, P.M.; Lerner, A. Type 4 cAMP phosphodiesterase (PDE4) inhibitors augment glucocorticoid-mediated apoptosis in B cell chronic lymphocytic leukemia (B-CLL) in the absence of exogenous adenylyl cyclase stimulation. Biochem. Pharmacol. 2005, 69, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Barenkamp, S.J.; Leininger, E. Cloning, expression, and DNA sequence analysis of genes encoding nontypeable Haemophilus influenzae high-molecular-weight surface-exposed proteins related to filamentous hemagglutinin of Bordetella pertussis. Infect. Immun. 1992, 60, 1302–1313. [Google Scholar] [PubMed]
- Lee, B.C.; Miyata, M.; Lim, J.H.; Li, J.D. Deubiquitinase CYLD acts as a negative regulator for bacterium NTHi-induced inflammation by suppressing K63-linked ubiquitination of MyD88. Proc. Natl. Acad. Sci. USA 2016, 113, E165–E171. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Lee, J.Y.; Susuki-Miyata, S.; Wang, W.Y.; Xu, H.; Kai, H.; Kobayashi, K.S.; Flavell, R.A.; Li, J.D. Glucocorticoids suppress inflammation via the upregulation of negative regulator IRAK-M. Nat. Commun. 2015, 6, 6062. [Google Scholar] [CrossRef] [PubMed]
- Horvat, J.C.; Beagley, K.W.; Wade, M.A.; Preston, J.A.; Hansbro, N.G.; Hickey, D.K.; Kaiko, G.E.; Gibson, P.G.; Foster, P.S.; Hansbro, P.M. Neonatal chlamydial infection induces mixed T-cell responses that drive allergic airway disease. Am. J. Respir. Crit. Care Med. 2007, 176, 556–564. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| human PDE4B | CTATACCGATCGCATTCAGGTC | CTGTCCATTGCCGATACAATT |
| human PDE4B2 | AGCGGTGGTAGCGGTGACTC | GCAGCGTGCAGGCTGTTGTG |
| human CXCL1 | TGCAGGGAATTCACCCCAAG | AGCTTTCCGCCCATTCTTGA |
| human CXCL2 | GTGTGAAGGTGAAGTCCCCC | AGCTTTCTGCCCATTCTTGA |
| human CCL5 | CTACACCAGTGGCAAGTGC | CTTTCGGGTGACAAAGACGAC |
| human CCL7 | GGCTTGCTCAGCCAGTTG | GGTGGTCCTTCTGTAGCTCTC |
| human Cyclophillin A | CGGGTCCTGGCATCTTGT | GCAGATGAAAAACTGGGAACCA |
| mouse pde4b2 | GTAGAGGCCAGTTCCCATCA | CCAACACCTAGTGCAGAGC |
| mouse cxcl1 | CATGGCTGGGATTCACCTCA | CCTCGCGACCATTCTTGAGT |
| mouse cxcl2 | TCAATGCCTGAAGACCCTG | GCAAACTTTTTGACCGCCCT |
| mouse ccl5 | CCTCACCATATGGCTCGGAC | ACGACTGCAAGATTGGAGCA |
| mouse ccl7 | CAATGCATCCACATGCTGCT | GCAGACTTCCATGCCCTTCT |
| mouse gapdh | ACCCAGAAGACTGTGGATGG | GGATGCAGGGATGATGTTCT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.-C.; Susuki-Miyata, S.; Yan, C.; Li, J.-D. Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. Int. J. Mol. Sci. 2018, 19, 3511. https://doi.org/10.3390/ijms19113511
Lee B-C, Susuki-Miyata S, Yan C, Li J-D. Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. International Journal of Molecular Sciences. 2018; 19(11):3511. https://doi.org/10.3390/ijms19113511
Chicago/Turabian StyleLee, Byung-Cheol, Seiko Susuki-Miyata, Chen Yan, and Jian-Dong Li. 2018. "Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi" International Journal of Molecular Sciences 19, no. 11: 3511. https://doi.org/10.3390/ijms19113511
APA StyleLee, B.-C., Susuki-Miyata, S., Yan, C., & Li, J.-D. (2018). Dexamethasone Inhibits Synergistic Induction of PDE4B Expression by Roflumilast and Bacterium NTHi. International Journal of Molecular Sciences, 19(11), 3511. https://doi.org/10.3390/ijms19113511