Effects of Acanthopanax senticosus on Brain Injury Induced by Simulated Spatial Radiation in Mouse Model Based on Pharmacokinetics and Comparative Proteomics

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Pharmacokinetics of Active Compounds of Acanthopanax senticosus (AS) Extract in Mouse Model
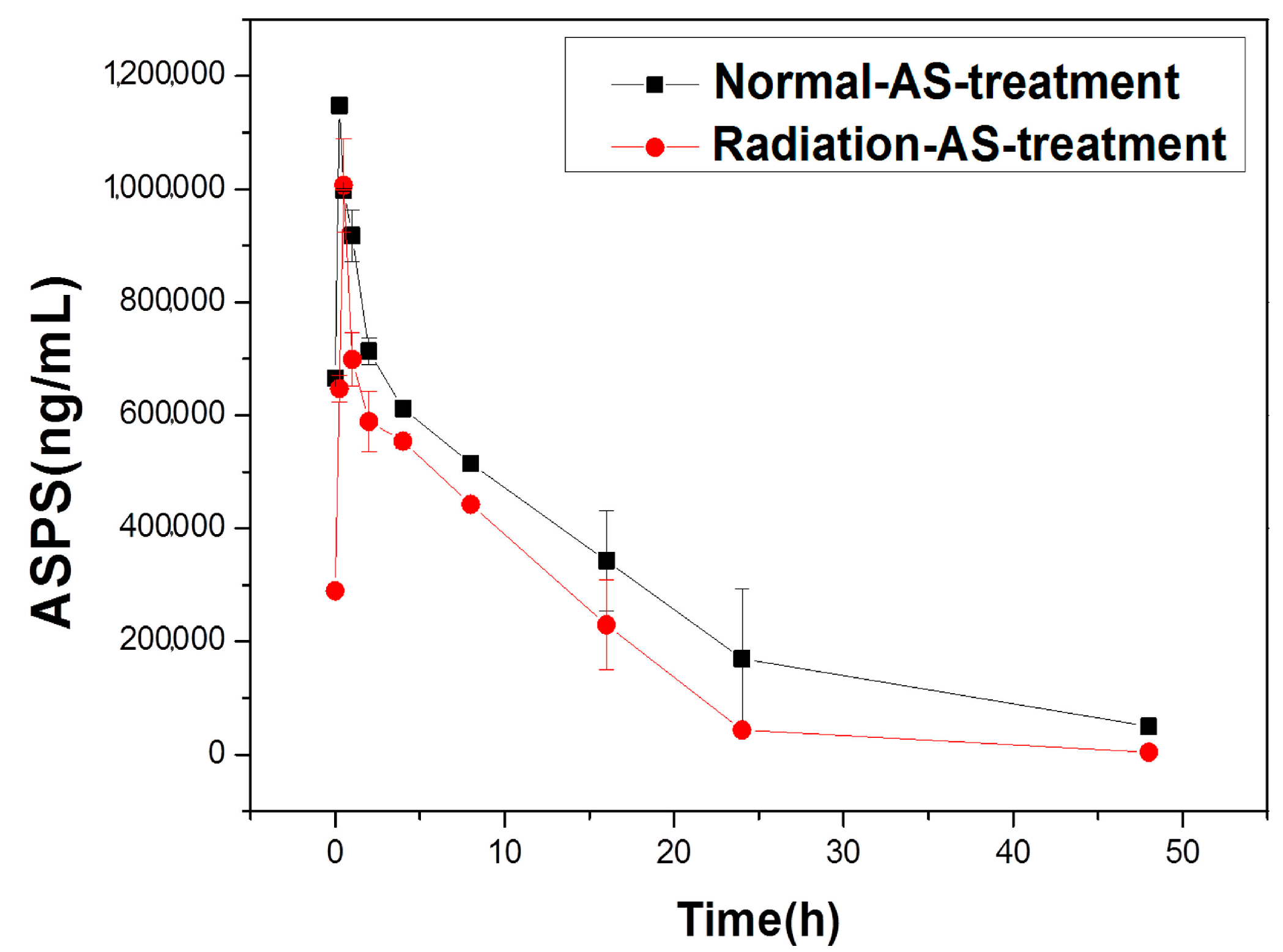
2.1.1. Pharmacokinetics of Acanthopanax senticosus polysaccharides (ASPS) of AS Extract in Irradiated Mice
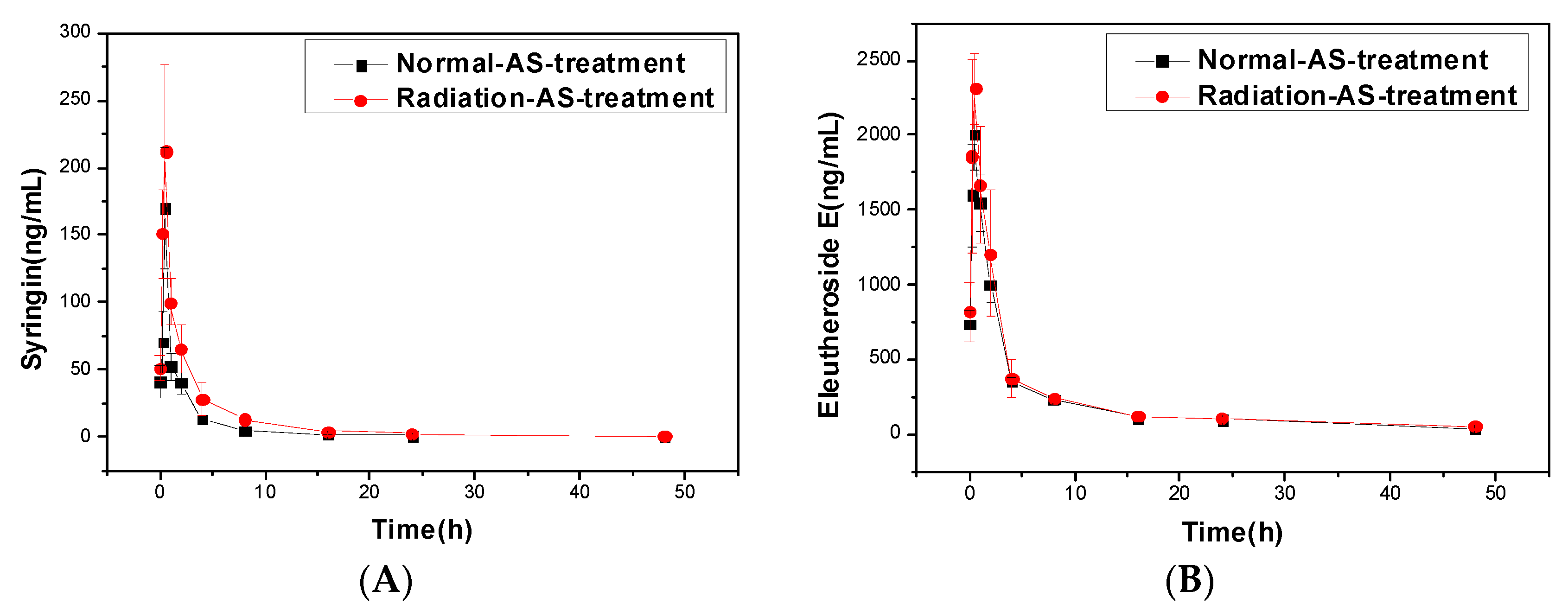
2.1.2. Pharmacokinetics of Syringin and Eleutheroside E of AS Extract in Irradiated Mice
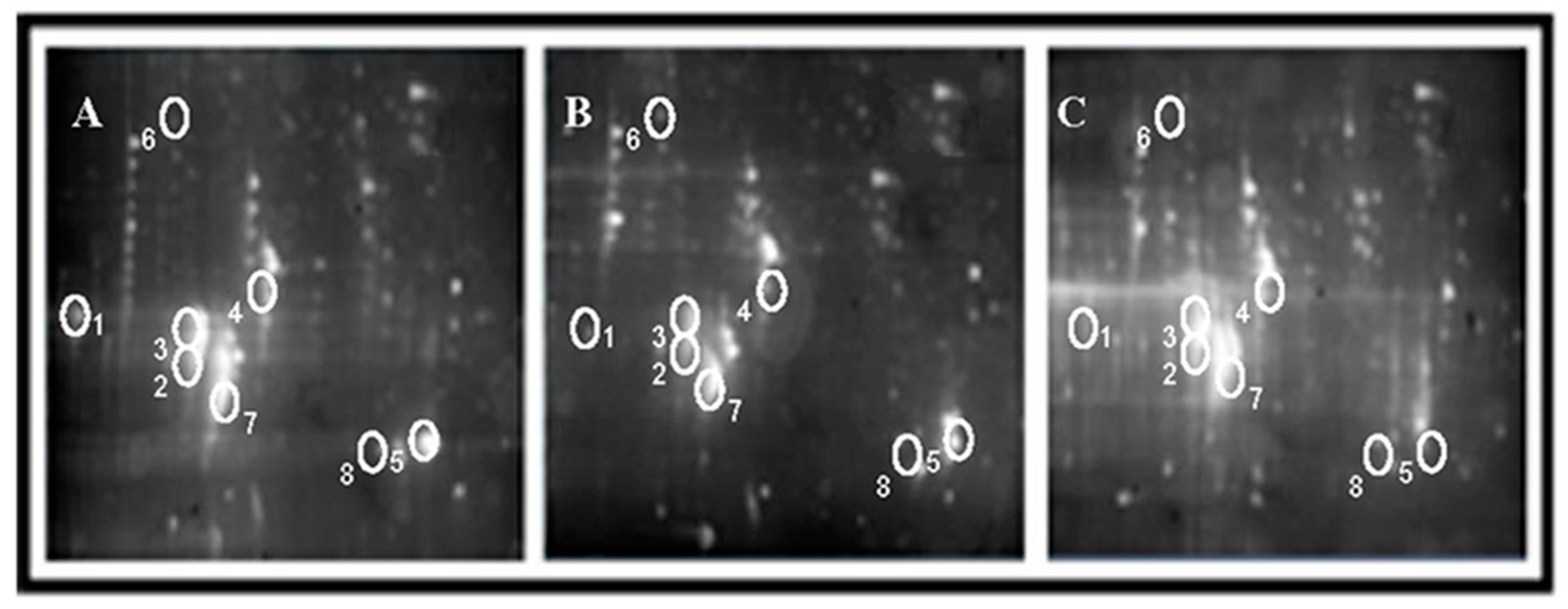
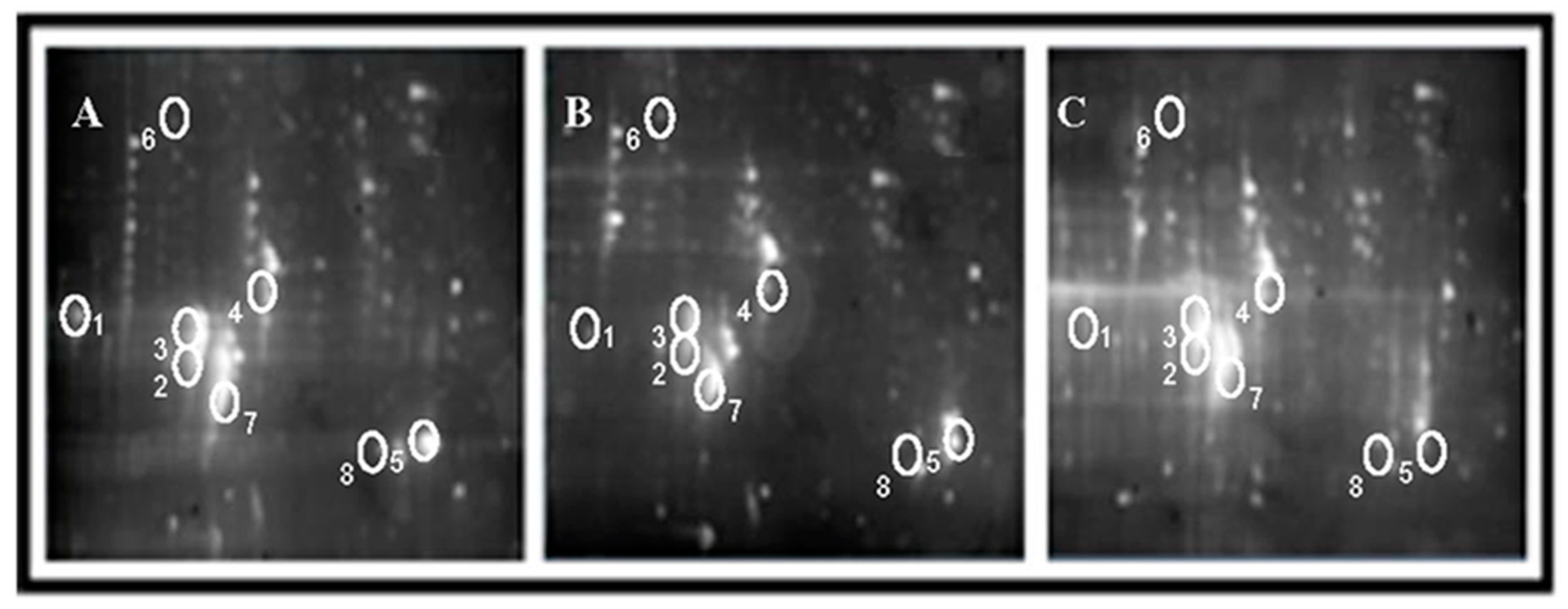
2.2. Effects of AS Extract on Proteins in PFC of Mice Brain after Radiation
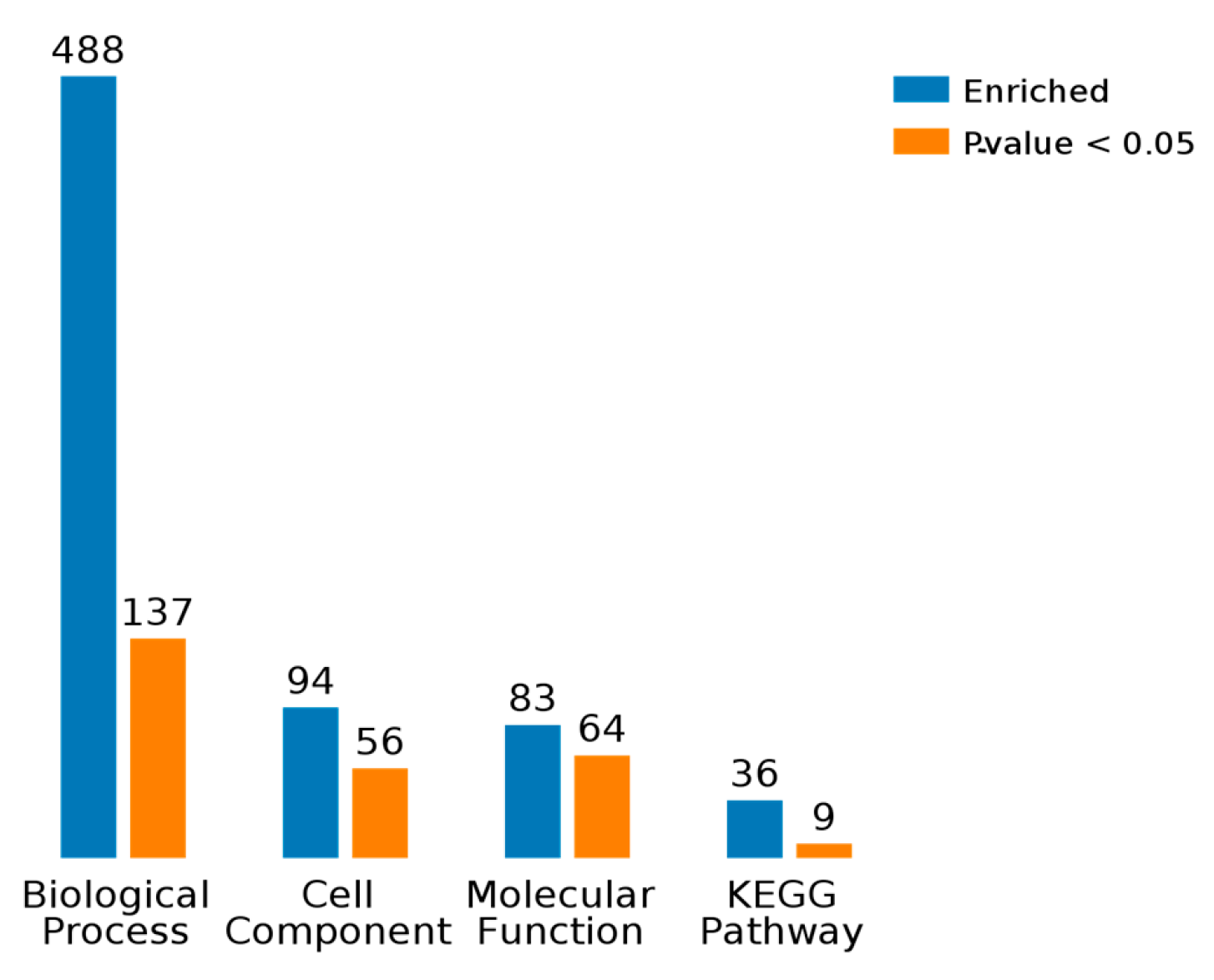
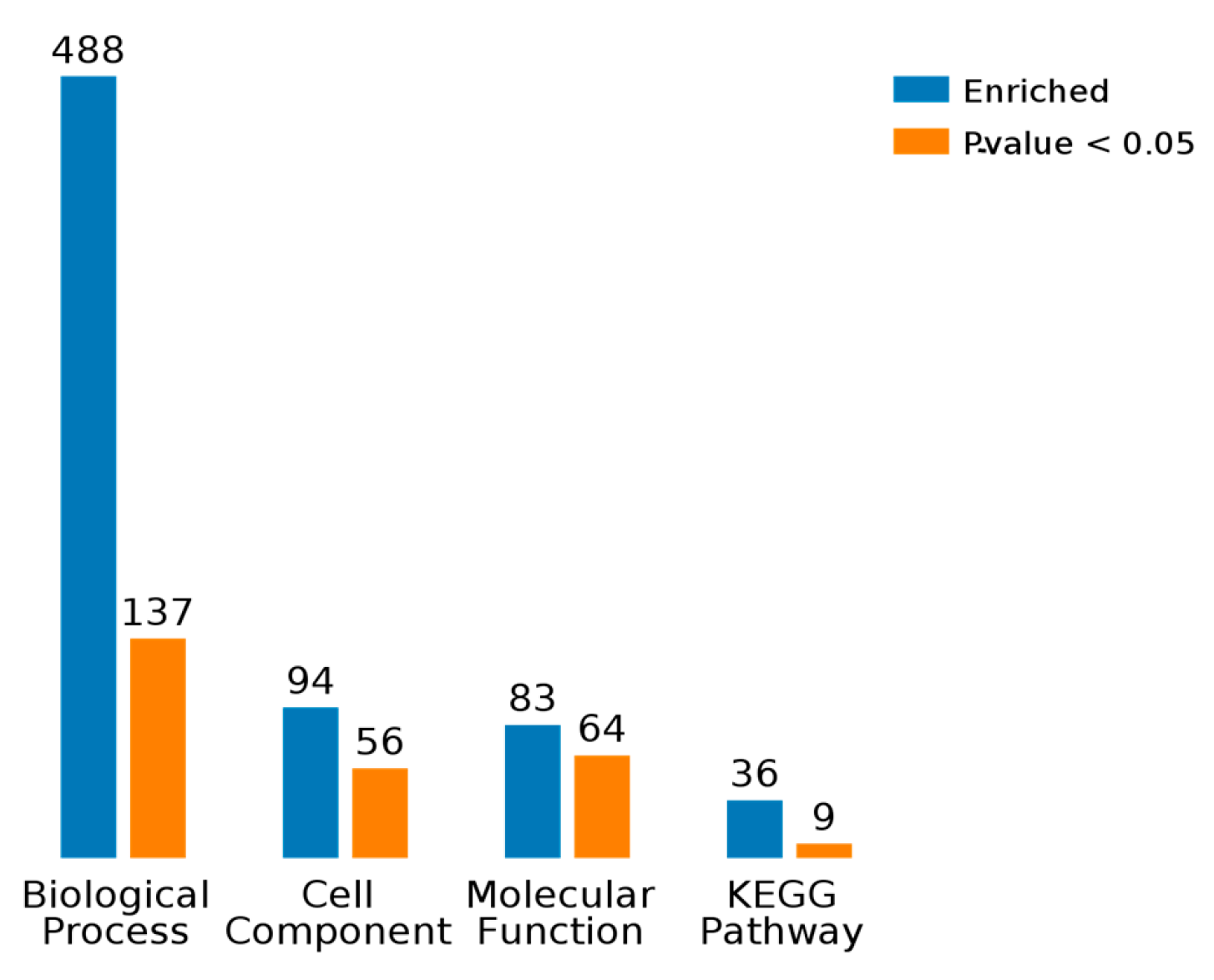
2.3. Results of Bioinformatics Analysis
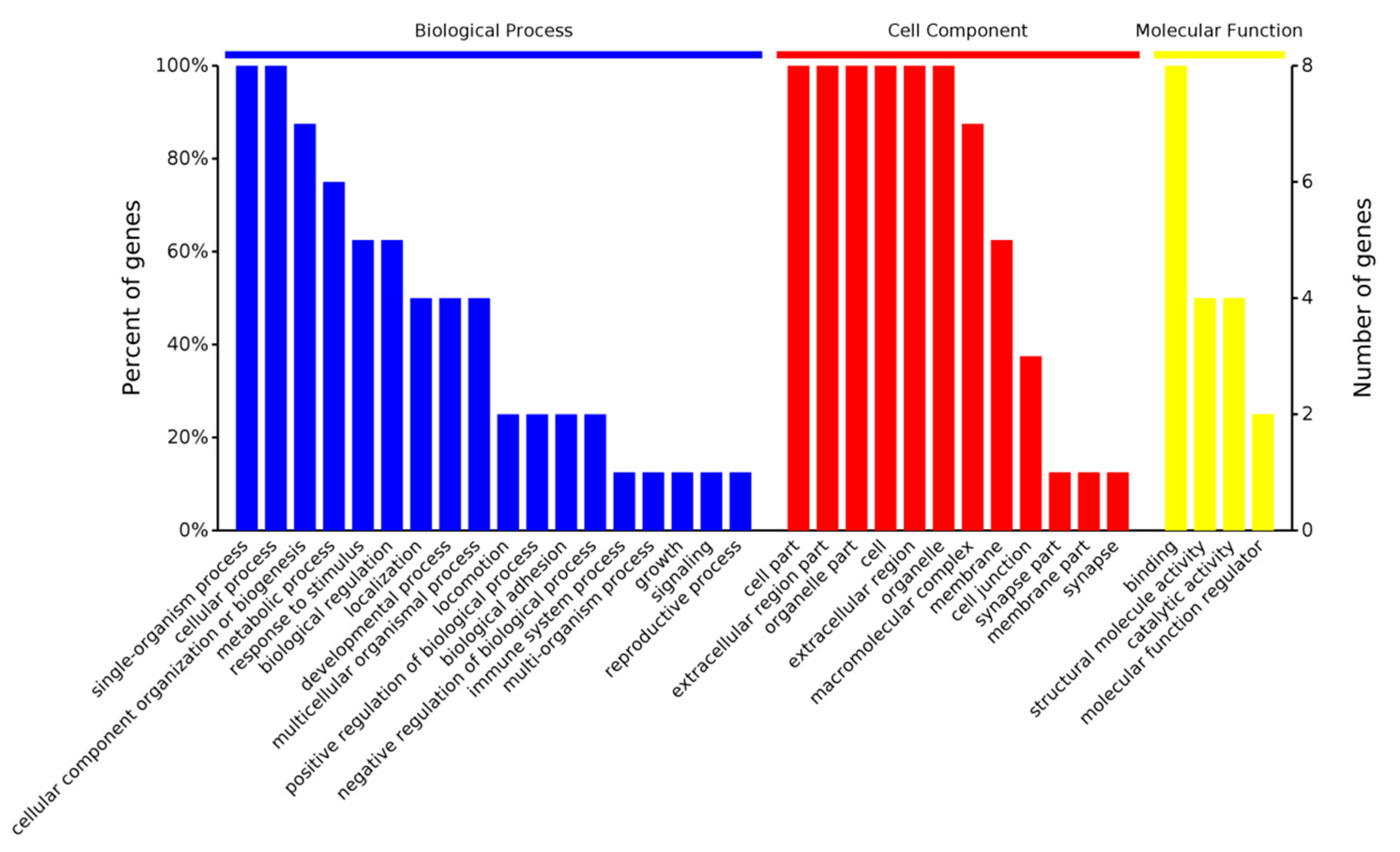
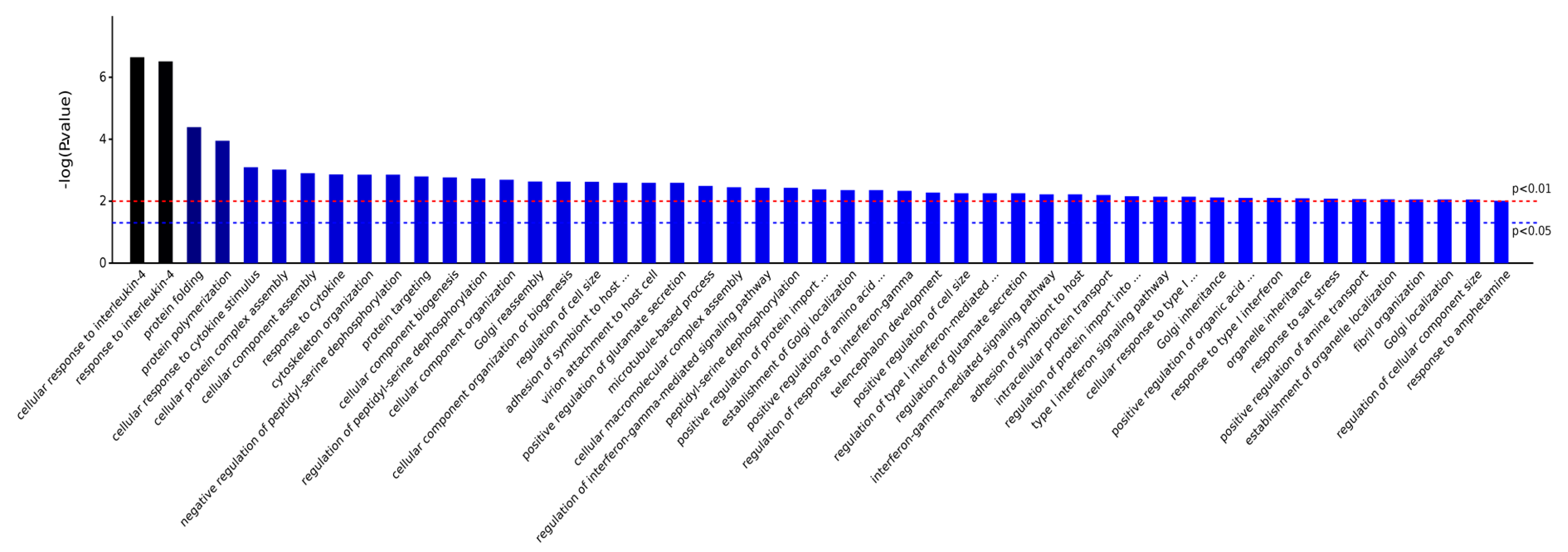
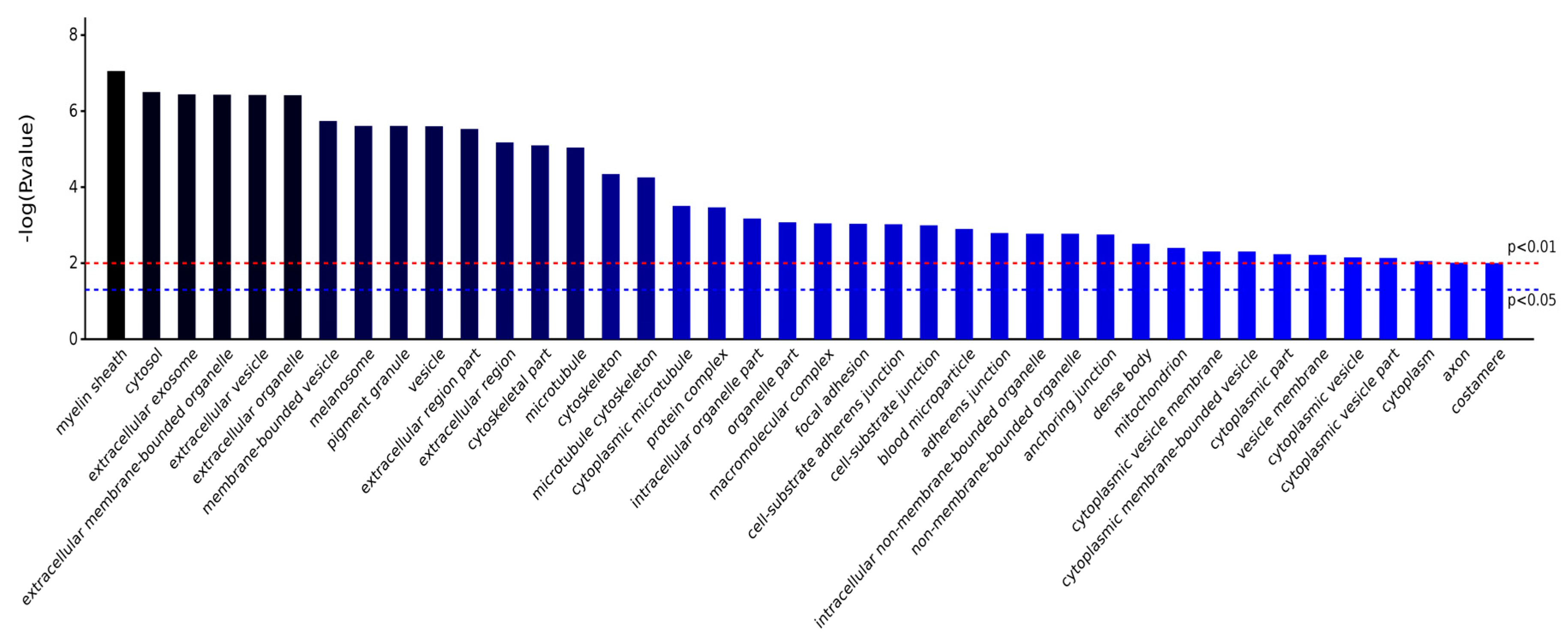
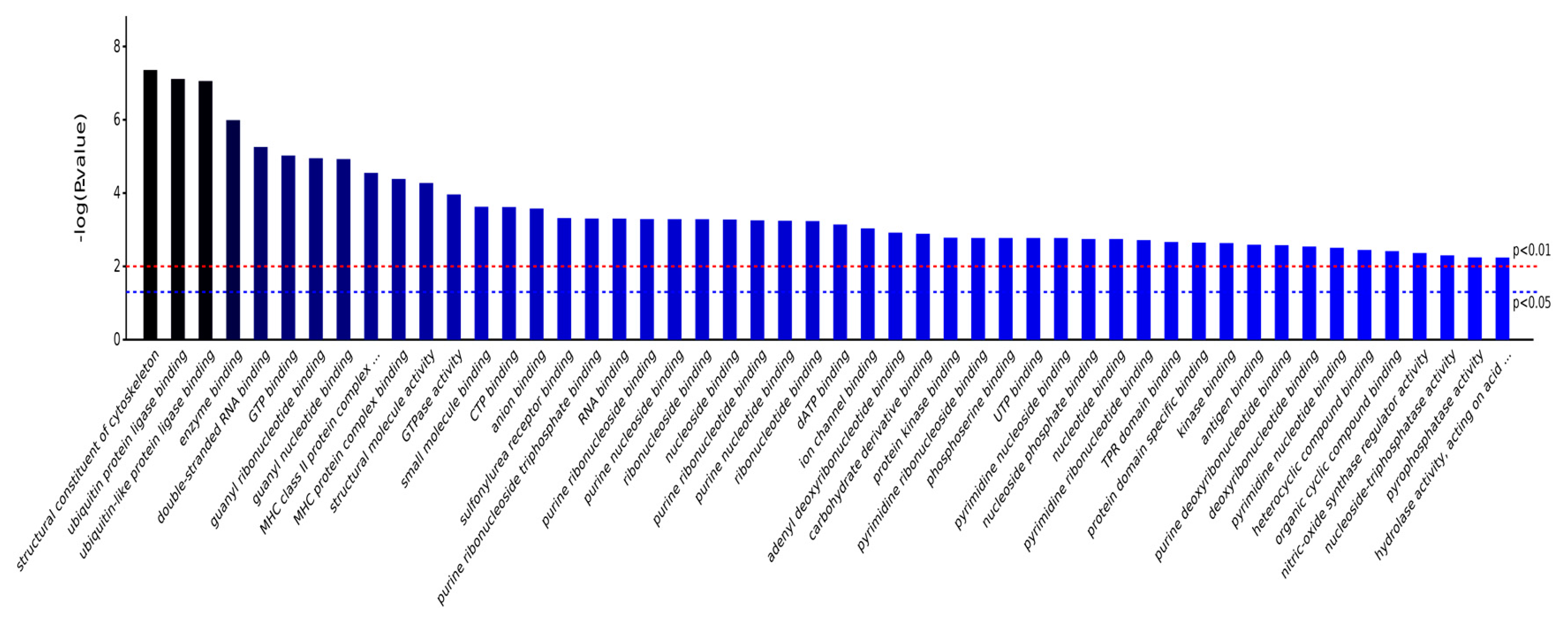
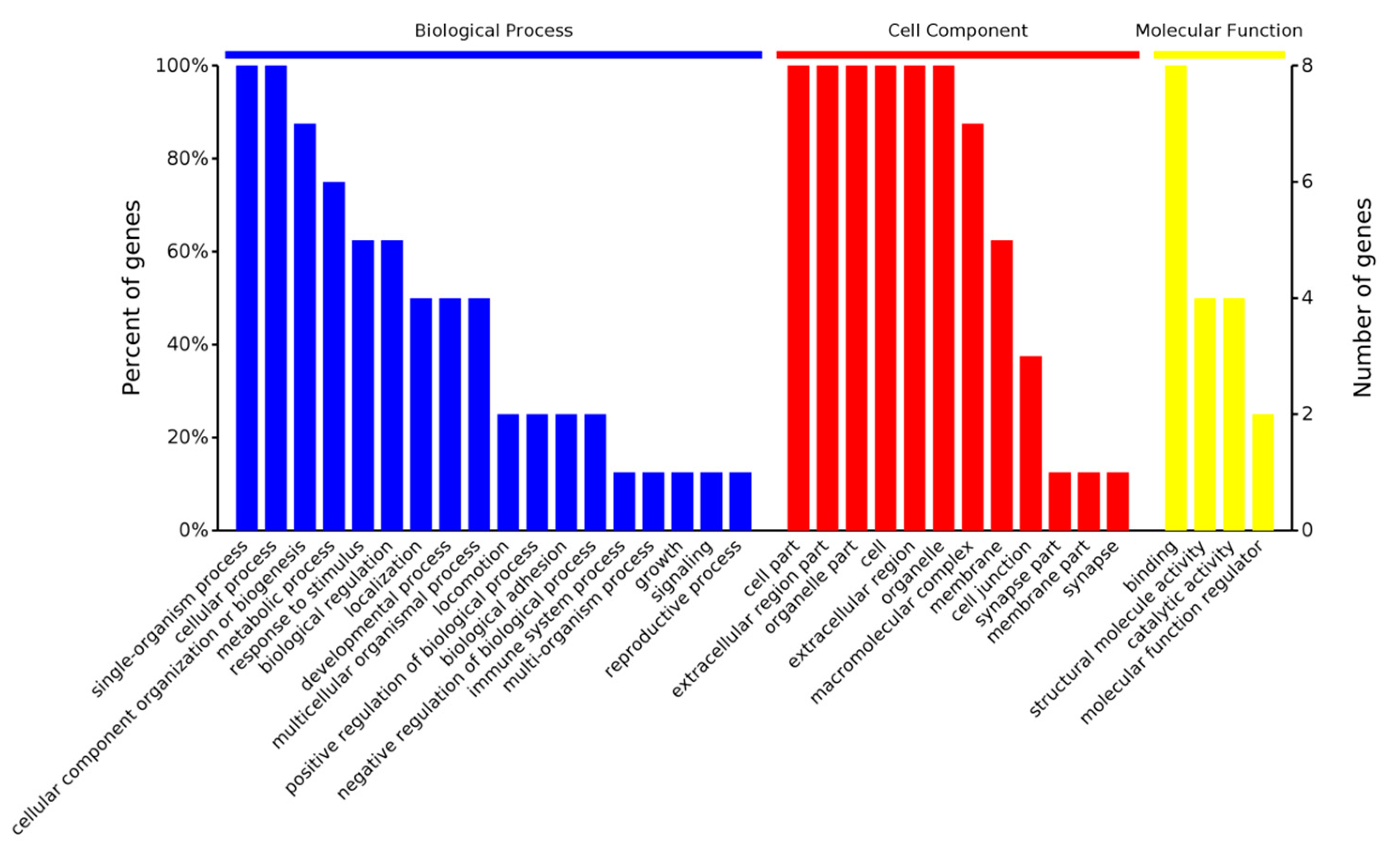
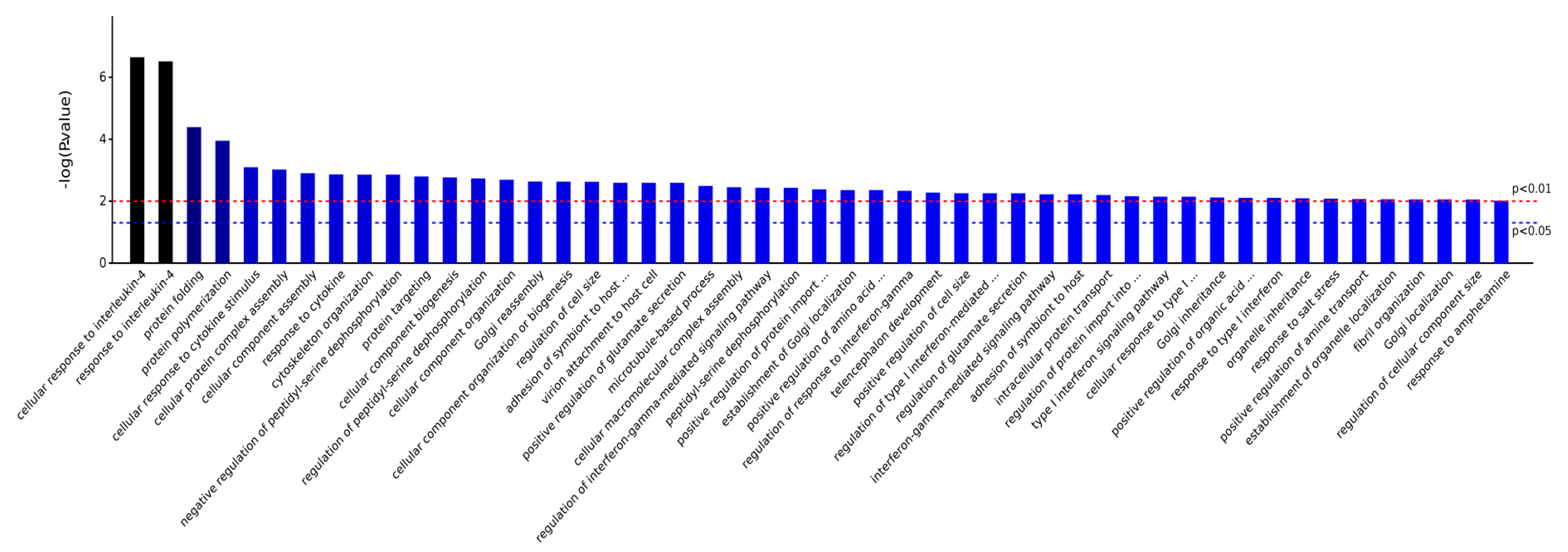
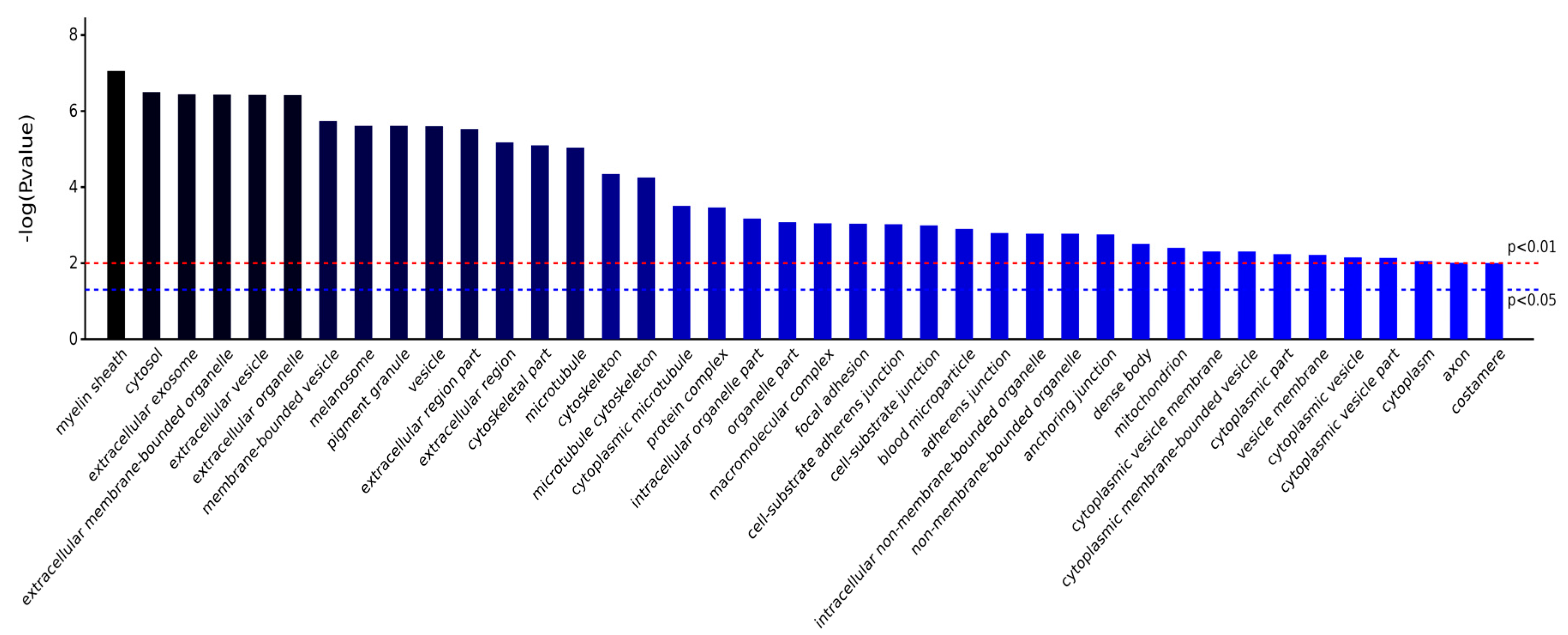
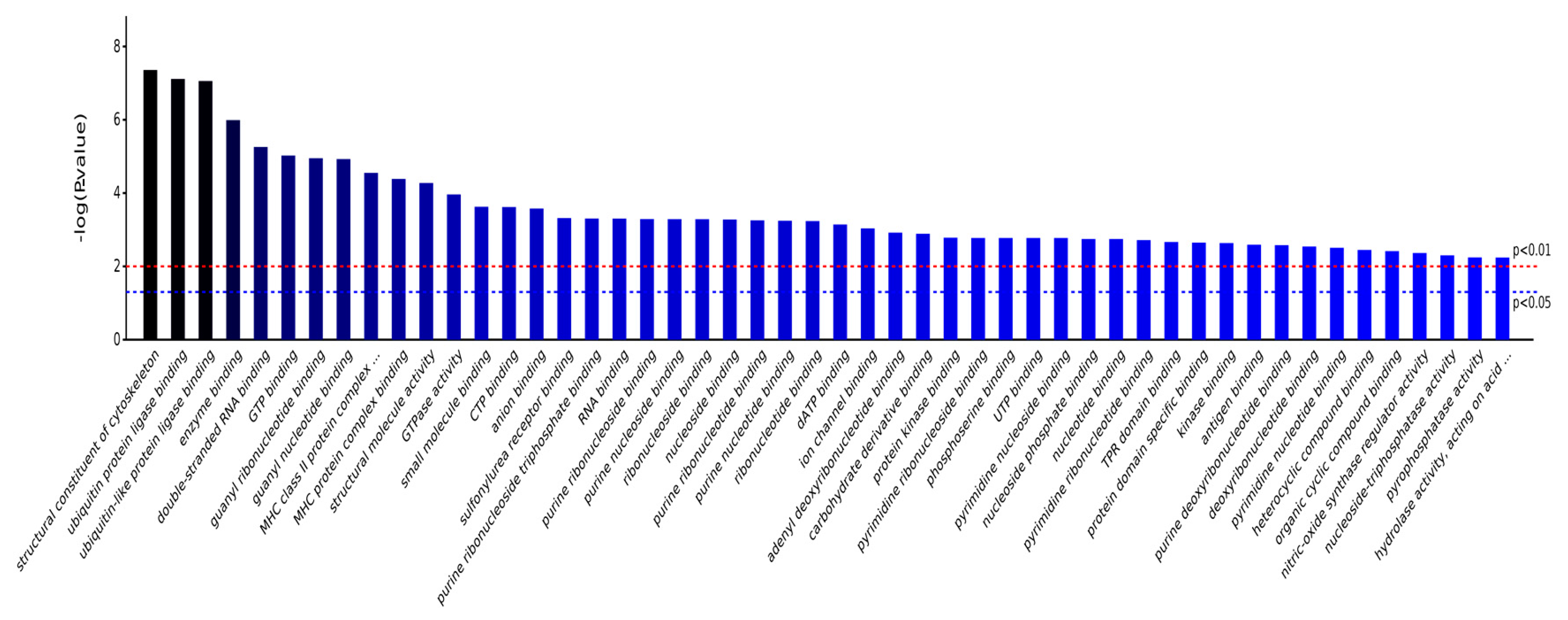
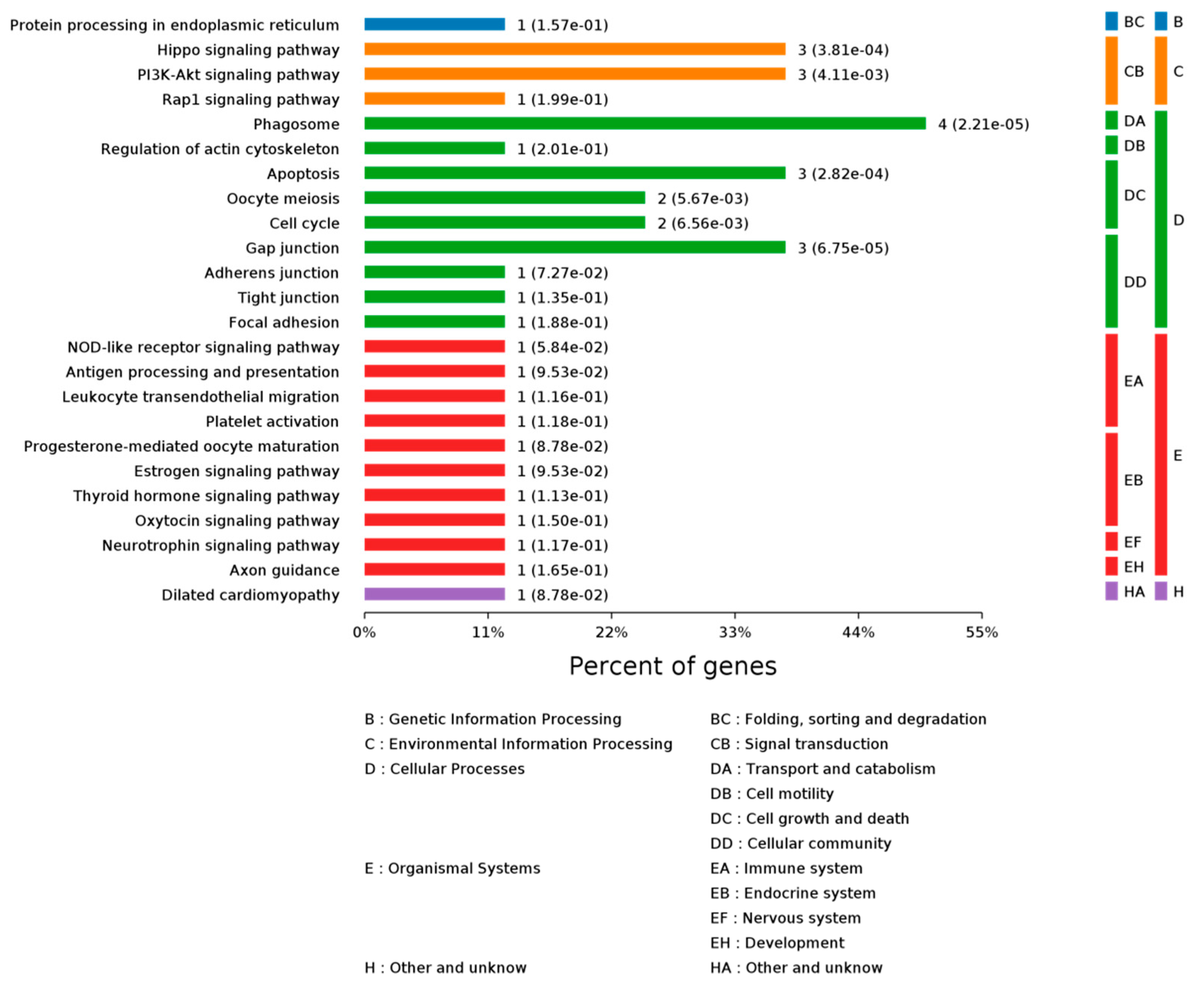
2.3.1. Gene Ontology Analysis
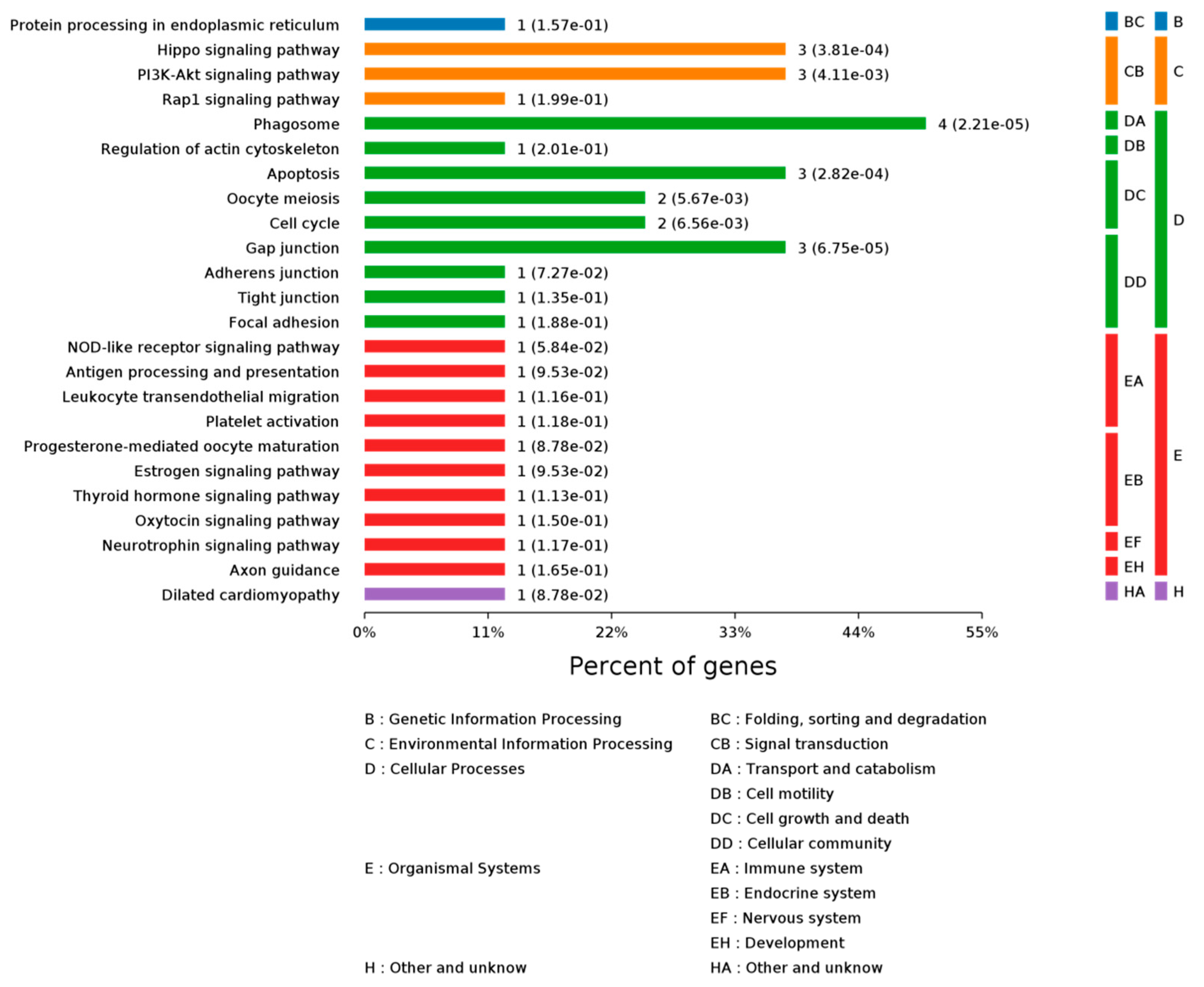
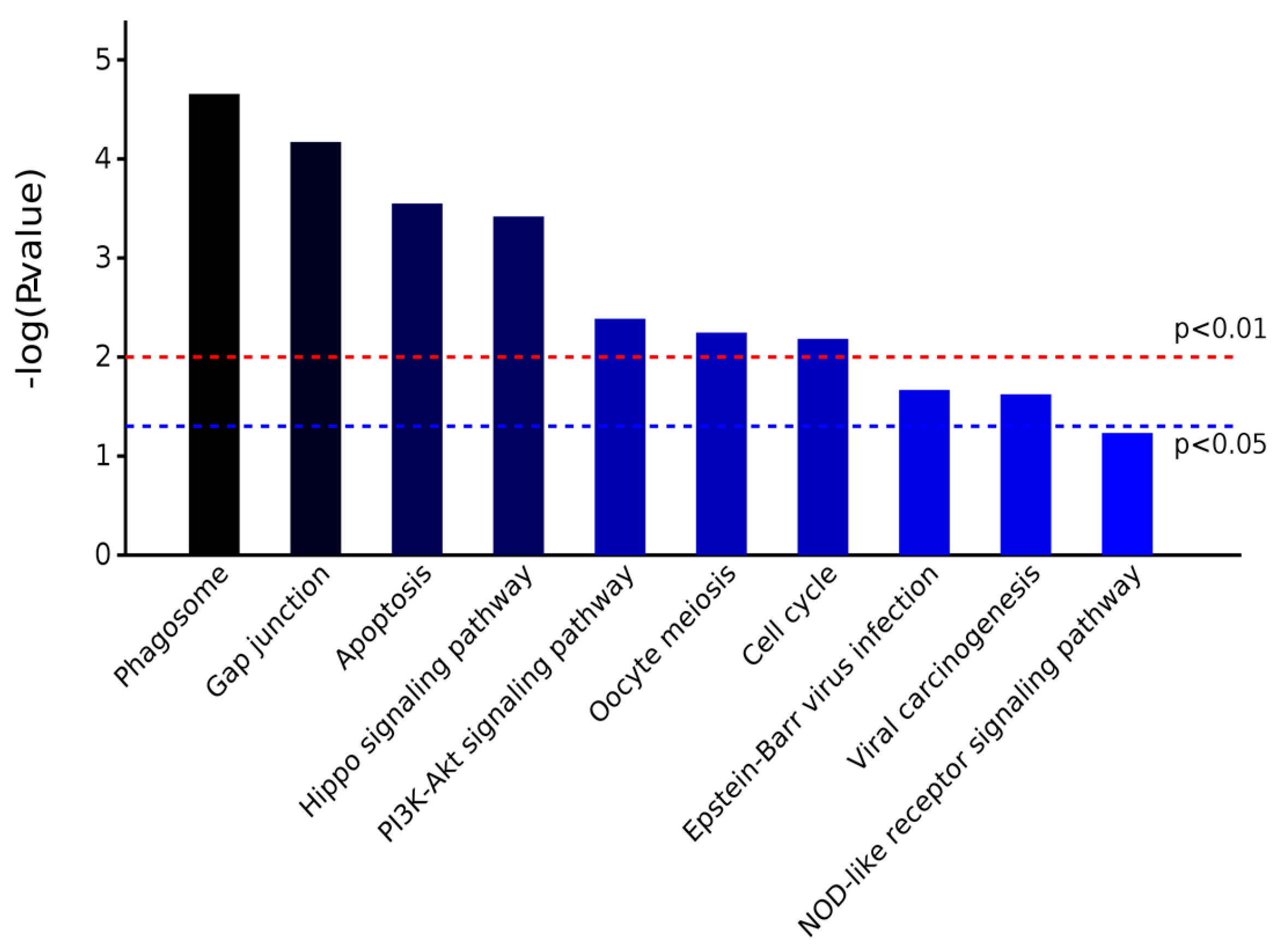
2.3.2. KEGG Pathway Analysis
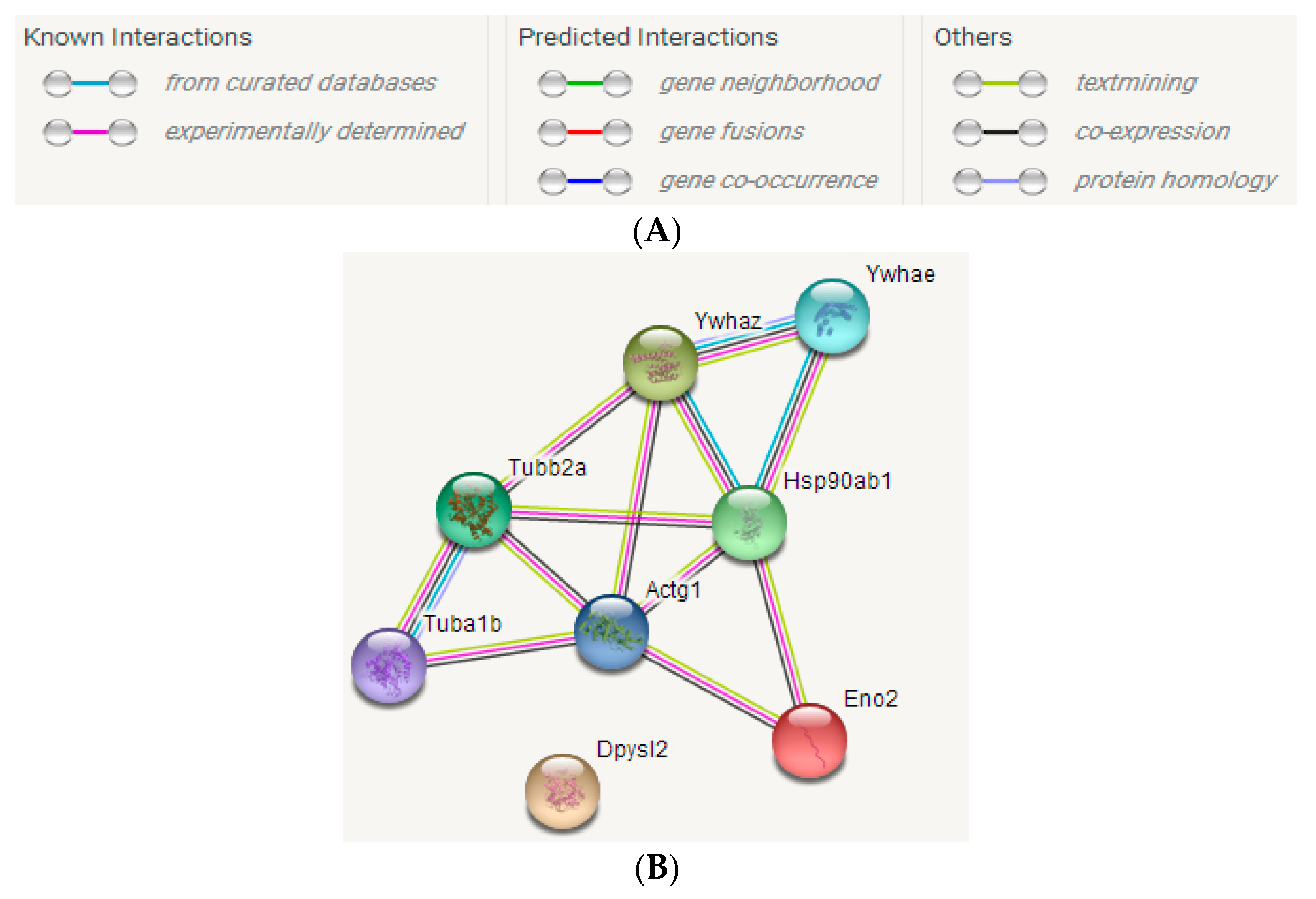
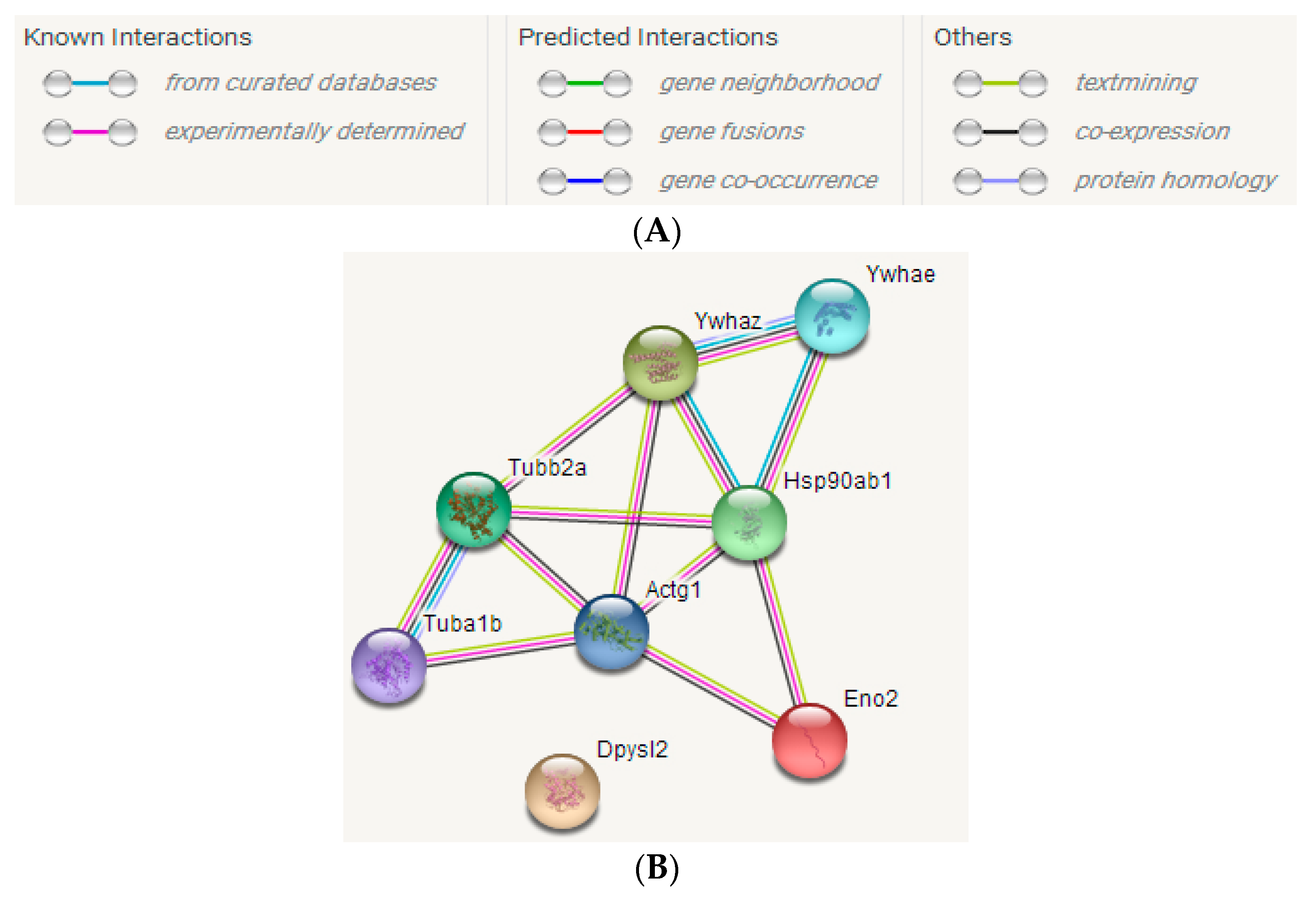
2.3.3. STRING Interactive Network Analysis
2.4. Analysis of Differentially Abundant Proteins
2.4.1. Tubulin Protein Family
2.4.2. Dihydropyrimidinase-Related Protein 2 (CRMP2)
2.4.3. Actin Proteins
2.4.4. 14-3-3 Protein Family
2.4.5. Heat Shock Protein (HSP) Family
2.4.6. Enolase 2
3. Materials and Methods
3.1. Plant Material
3.2. Animal Material
3.3. Multi-Component Pharmacokinetics of AS Extract
3.3.1. Pharmacokinetics of ASPS
3.3.2. Pharmacokinetics of Syringin and Eleutheroside E
3.4. Two-Dimensional Gel Electrophoresis (2-DE) Analysis
3.4.1. 2-DE of Prefrontal Cortex (PFC)
3.4.2. Image Analysis of 2-DE Gels
3.4.3. Identification of Proteins from 2DE Gels
3.4.4. Bioinformatics Analysis
3.4.5. Statistical Analysis
4. Conclusions and Expectations
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Song, Y.; Salbu, B.; Teien, H.C.; Evensen, Ø.; Lind, O.C.; Rosseland, B.O.; Tollefsen, K.E. Hepatic transcriptional responses in atlantic salmon (salmo salar) exposed to γ radiation and depleted uranium singly and in combination. Sci. Total Environ. 2016, 562, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Newman, V.L.; Berg, A.N.; Macvittie, T.J. Animal models for acute radiation syndrome drug discovery. Expert Opin. Drug Discov. 2015, 10, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Milligan, J.R.; Aguilera, J.A.; Paglinawan, R.A.; Ward, J.F.; Limoli, C.L. DNA strand break yields after post-high let irradiation incubation with endonuclease-III and evidence for hydroxyl radical clustering. Int. J. Radiat. Biol. 2001, 77, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Caffrey, M. Free radical mediated X-ray damage of model membranes. Biophys. J. 1996, 70, 2212–2222. [Google Scholar] [CrossRef]
- Berovic, N.; Pratontep, S.; Bryant, A.; Montouris, A.; Green, R.G. The kinetics of radiation damage to the protein luciferase and recovery of enzyme activity after irradiation. Radiat. Res. 2014, 157, 122–127. [Google Scholar] [CrossRef]
- Brook, I.; Elliott, T.B.; Ledney, G.D.; Shoemaker, M.O.; Knudson, G.B. Management of postirradiation infection: Lessons learned from animal models. Mil. Med. 2004, 169, 194–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Soo, M.Y.; Srinivasan, N.; Tan, B.K.; Chan, S.H. Activation of macrophages by polysaccharide-protein complex from Lycium barbarum L. Phytother. Res. 2009, 23, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Zhong, Y.; Yong, Y.I.; Wu, X.-R.; Fan, Q.C.; Zhao, S.; Bai, G.E. Effects of Acanthopanax senticosus saponinon blood lipid, blood glucose, immune function, liver and renal function in simulated weightlessness rats. China J. Tradit. Chin. Med. Pharm. 2017, 32, 4761–4764. [Google Scholar]
- Zhou, L.J.; Long, T.T.; Zhou, X.; Bao, Y.X. Immunomodulatory effects of Acanthopanax senticosus polysaccharides on lewis tumor-bearing mice through TLR4 signaling pathway. Chin. J. Immunol. 2017, 33, 849. [Google Scholar]
- Kang, M.C.; Kim, S.Y.; Kim, E.A.; Lee, J.H.; Kim, Y.S.; Yu, S.K.; Chae, J.B.; Choe, I.H.; Cho, J.H.; Jeon, Y.J. Antioxidant activity of polysaccharide purified from Acanthopanax koreanum nakai stems in vitro and in vivo zebrafish model. Carbohydr. Polym. 2015, 127, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ying, D.; Huang, D.; Jing, W.; Liu, Z.; Li, F. LC–MS/MS determination and pharmacokinetic study of four lignan components in rat plasma after oral administration of acanthopanax sessiliflorus extract. J. Ethnopharmacol. 2012, 141, 957–963. [Google Scholar]
- Liu, S.; Tang, B.; Lu, F.; Fan, Z.; Zhou, S. Pharmacokinetics of isofraxidin in extracellular fluids of striatum in rats using microdialysis-uplc method. World Sci. Technol. 2012, 14, 1206–1210. [Google Scholar]
- He, X.; Xing, D.; Ding, Y.; Li, Y.; Xu, L.; Du, L. Effects of cerebral ischemia-reperfusion on pharmacokinetic fate of paeoniflorin after intravenous administration of paeoniae radix extract in rats. J. Ethnopharmacol. 2004, 94, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Guipaud, O. Serum and plasma proteomics and its possible use as detector and predictor of radiation diseases. Adv. Exp. Med. Biol. 2013, 990, 61–86. [Google Scholar] [PubMed]
- Orre, L.M.; Pernemalm, M.; Lengqvist, J.; Lewensohn, R.; Lehtiö, J. Up-regulation, modification, and translocation of s100a6 induced by exposure to ionizing radiation revealed by proteomics profiling. Mol. Cell. Proteom. 2007, 6, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Craft, J.M.; Townsend, R.; Deasy, J.O.; Bradley, J.D.; Naqa, I.E. A bioinformatics approach for biomarker identification in radiation-induced lung inflammation from limited proteomics data. J. Proteome Res. 2011, 10, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Park, E.C.; Yoon, J.B.; Seong, J.S.; Choi, K.S.; Kong, E.S.; Kim, Y.J.; Park, Y.M.; Park, E.M. Effect of ionizing radiation on rat tissue: Proteomic and biochemical analysis. Prep. Biochem. Biotechnol. 2006, 36, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.X.; Zhao, H.B.; Li, C.R.; Sun, Y.N.; Qian, X.H.; Wang, S.Q. Proteomic analysis of ionizing radiation-induced proteins at the subcellular level. J. Proteome Res. 2009, 8, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Sun, Y.; Shi, J. Radioprotective effects of active compounds from Acanthopanax senticosus of Lesser Khingan mountain in China. RSC Adv. 2015, 6, 65–72. [Google Scholar]
- Nishibe, S.; Kinoshita, H.; Kitagawa, S.; Fujimoto, T.; Takeda, H.; Morita, I.; Okano, G.; Fujisawa, T.; Goto, T. PC48 phenolic compounds from Acanthopanax senticosus and their pharmacological effects on swimming exercise in rats. Int. Symp. Chem. Natl. Prod. 1988, 5, 504. [Google Scholar]
- Huang, Z.; Cen, H.; Wu, Q.; Li, D.; Jiang, H.; Zheng, Y.; Pharmacy, S.O. Antidepressant effects of Acanthopanax senticosus and Crataegi fructus aqueous extract with different proportion in mice. J. Pharm. Res. 2017, 36, 497–500. [Google Scholar]
- Yan, B.; Xing, D.; Ding, Y.; Tao, J.; Du, L.J. HPLC method for the determination and pharmacokinetic studies on puerarin in cerebral ischemia reperfusion rat plasma after intravenous administration of puerariae radix isoflavone. J. Pharm. Biomed. Anal. 2005, 37, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Son, D.; Ryu, J.; Lee, Y.S.; Sang, H.J.; Kang, J.; Sang, Y.L.; Kim, H.S.; Shin, K.H. Anti-oxidant activities of Acanthopanax senticosus stems and their lignan components. Arch. Pharm. Res. 2004, 27, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Z.; Huang, B.K.; Liang, J.; Zheng, C.J.; Han, T.; Zhang, Q.Y.; Qin, L.P. Antifatigue activity of the liposoluble fraction from Acanthopanax senticosus. Phytother. Res. PTR 2011, 25, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.M.; Yang, B.K.; Lu, F.; Bai, Y.; Mou, H.; Ma, D. Metabolism of eleutheroside E and B in rat intestinal flora in vitro. Chin. Pharm. 2011, 14, 1075–1077. [Google Scholar]
- Zhou, J.; Zeng, P.; Cheng, Z.H.; Liu, J.; Wang, F.Q.; Qian, R.J. Application of hollow fiber liquid phase microextraction coupled with high-performance liquid chromatography for the study of the osthole pharmacokinetics in cerebral ischemia hypoperfusion rat plasma. J. Chromatogr. B 2011, 879, 2304–2310. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Coffee, P.; Smith, G.; Liem, R.K.; Brady, S.T.; Black, M.M. Neurofilaments are transported rapidly but intermittently in axons: Implications for slow axonal transport. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 6849–6861. [Google Scholar]
- Lo, L.P.; Liu, S.H.; Chang, Y.C. Assembling microtubules disintegrate the postsynaptic density in vitro. Cell Motil. Cytoskelet. 2007, 64, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Poddar, R.; Paul, S.; Chaudhury, S.; Sarkar, P.K. Regulation of actin and tubulin gene expression by thyroid hormone during rat brain development. Mol. Brain Res. 1996, 35, 111–118. [Google Scholar] [CrossRef]
- Nishimura, T.; Fukata, Y.; Kato, K.; Yamaguchi, T.; Matsuura, Y.; Kamiguchi, H.; Kaibuchi, K. CRMP-2 regulates polarized numb-mediated endocytosis for axon growth. Nat. Cell Biol. 2003, 5, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Yoshimura, T.; Tsuboi, D.; Kawabata, S.; Kanekokawano, T.; Shirataki, H.; Takenawa, T.; Kaibuchi, K. CRMP-2 is involved in kinesin-1-dependent transport of the SRA-1/wave1 complex and axon formation. Mol. Cell. Biol. 2005, 25, 9920–9935. [Google Scholar] [CrossRef] [PubMed]
- Vuaillat, C.; Varrindoyer, M.; Bernard, A.; Sagardoy, I.; Cavagna, S.; Chounlamountri, I.; Lafon, M.; Giraudon, P. High CRMP-2 expression in peripheral T lymphocytes is associated with recruitment to the brain during virus-induced neuroinflammation. J. Neuroimmunol. 2008, 193, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.A.; Lee, J.E.; Lee, J.Y.; Ko, M.J.; Lee, S.T.; Kim, H.J. Alteration of collapsin response mediator protein-2 expression in focal ischemic rat brain. Neuroreport 2005, 16, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Shirakashi, Y.; Kawamoto, Y.; Tomimoto, H.; Takahashi, R.; Ihara, M. Α-synuclein is colocalized with 14-3-3 and synphilin-1 in A53T transgenic mice. Acta Neuropathol. 2006, 112, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Tanabe, Y.; Uchikawa, H.; Kobayashi, K.; Kubota, H.; Takanashi, J.; Kohno, Y. 14-3-3 protein detection in the cerebrospinal fluid of patients with influenza-associated encephalopathy. J. Child Neurol. 2006, 21, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.C.; Sun, Y.; Xu, R.; Sang, Y.; Zhao, J.; Liu, G.; Cai, L.; Li, C.; Zhao, S. The interaction between adam 22 and 14-3-3ζ: Regulation of cell adhesion and spreading. Biochem. Biophys. Res. Commun. 2003, 301, 991–999. [Google Scholar] [CrossRef]
- Sunayama, J.; Tsuruta, F.; Masuyama, N.; Gotoh, Y. JNK antagonizes AKT-mediated survival signals by phosphorylating 14-3-3. J. Cell Biol. 2005, 170, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Csermely, P.; Schnaider, T.; Soti, C.; Prohászka, Z.; Nardai, G. The 90-kDa molecular chaperone family: Structure, function, and clinical applications. A comprehensive review. Pharmacol. Ther. 1998, 79, 129–168. [Google Scholar] [CrossRef]
- Grad, I.; Cederroth, C.R.; Walicki, J.; Grey, C.; Barluenga, S.; Winssinger, N.; De, M.B.; Nef, S.; Picard, D. The molecular chaperone HSP90Α is required for meiotic progression of spermatocytes beyond pachytene in the mouse. PLoS ONE 2010, 5, e15770. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1955, 28, 350–356. [Google Scholar] [CrossRef]
- Yin, Y.; Gong, F.Y.; Wu, X.X.; Sun, Y.; Li, Y.H.; Chen, T.; Xu, Q. Anti-inflammatory and immunosuppressive effect of flavones isolated from artemisia vestita. J. Ethnopharmacol. 2008, 120, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.M.; Li, X.Z.; Huo, Y.; Lu, F. Protective effect of extract of Acanthopanax senticosus harms on dopaminergic neurons in Parkinson’s disease mice. Phytomedicine 2012, 19, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Kitteringham, N.R.; Abdullah, A.; Walsh, J.; Randle, L.; Jenkins, R.E.; Sison, R.; Goldring, C.E.; Powell, H.; Sanderson, C.; Williams, S. Proteomic analysis of NRF2 deficient transgenic mice reveals cellular defence and lipid metabolism as primary NRF2-dependent pathways in the liver. J. Proteom. 2010, 73, 1612–1631. [Google Scholar] [CrossRef] [PubMed]
- Görg, A.; Obermaier, C.; Boguth, G.; Harder, A.; Scheibe, B.; Wildgruber, R.; Weiss, W. The current state of two-dimensional electrophoresis with immobilized pH gradients. Electrophoresis 2006, 9, 175–188. [Google Scholar]
- Fountoulakis, M.; Berndt, P.; Boelsterli, U.A.; Crameri, F.; Winter, M.; Albertini, S.; Suter, L. Two-dimensional database of mouse liver proteins: Changes in hepatic protein levels following treatment with acetaminophen or its nontoxic regioisomer 3-acetamidophenol. Electrophoresis 2000, 21, 2148–2161. [Google Scholar] [CrossRef]
- Castegna, A.; Aksenov, M.; Thongboonkerd, V.; Klein, J.B.; Pierce, W.M.; Booze, R.; Markesbery, W.R.; Butterfield, D.A. Proteomic identification of oxidatively modified proteins in Alzheimer’s disease brain. Part II: Dihydropyrimidinase-related protein 2, α-enolase and heat shock cognate71. J. Neurochem. 2002, 33, 562–571. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Content of the Active Compounds (%) | ||||
|---|---|---|---|---|---|
| Tissue of Lesser Khingan Mountain Acanthopanax senticosus | |||||
| Root | Stem | Leaf | Fruit | Average | |
| Polysaccharides | 6.05 ± 0.25 | 8.09 ± 0.37 | 4.12 ± 0.19 | 3.08 ± 0.13 | 5.33 ± 0.24 |
| Flavones | 14.25 ± 0.66 | 9.26 ± 0.45 | 4.02 ± 0.17 | 9.58 ± 0.50 | 9.28 ± 0.45 |
| Syringin | 1.44 ± 0.06 | 1.86 ± 0.08 | 0.09 ± 0.01 | 0.02 ± 0.001 | 3.40 ± 0.04 |
| Eleutheroside E | 3.17 ± 0.12 | 4.14 ± 0.25 | 0.18 ± 0.01 | 0.04 ± 0.002 | 1.88 ± 0.10 |
| Parameters | ASPS | |
|---|---|---|
| Normal-AS-Treatment | Radiation-AS-Treatment | |
| 1.18 × 101 ± 6.85 × 10−1 | 6.14 × 100 ± 7.82 × 10−1 ** | |
| 2.50 × 10−1 ± 3.81 × 10−1 | 5.00 × 10−1 ± 1.44 × 10−1 | |
| 1.15 × 106 ± 8.13 × 104 | 1.01 × 106 ± 1.24 × 105 | |
| 1.43 × 107 ± 3.00 × 106 | 8.92 × 106 ± 8.81 × 105 * | |
| 1.59 × 101 ± 9.63 × 10−1 | 9.07 × 100 ± 1.95 × 10−1 ** | |
| 9.83 × 10−4 ± 1.76 × 10−4 | 8.19 × 10−4 ± 8.33 × 10−5 | |
| 5.76 × 10−5 ± 6.12 × 10−6 | 9.25 × 10−5 ± 8.50 × 10−6 * | |
| Parameters | Syringin | Eleutheroside E | ||
|---|---|---|---|---|
| Normal-AS-Treatment | Radiation-AS-Treatment | Normal-AS-Treatment | Radiation-AS-Treatment | |
| 6.47 × 100 ± 6.61 × 10−1 | 9.72 × 100 ± 3.81 × 10−1 *** | 1.72 × 101 ± 7.22 × 10−1 | 2.51 × 101 ± 5.56 × 100 ** | |
| 5.00 × 10−1 ± 1.83 × 10−1 | 5.00 × 10−1 ± 2.37 × 10−1 | 5.00 × 10−1 ± 1.92 × 10−1 | 5.00 × 10−1 ± 3.21 × 10−1 | |
| 1.70 × 102 ± 7.41 × 100 | 2.12 × 102 ± 1.97 × 101 *** | 2.07 × 103 ± 6.61 × 102 | 2.11 × 103 ± 5.93 × 102 | |
| 2.83 × 102 ± 3.23 × 101 | 5.34 × 102 ± 6.54 × 10−1 | 9.93 × 103 ± 1.81 × 103 | 1.20 × 104 ± 3.63 × 103 * | |
| 4.58 × 100 ± 6.12 × 10−1 | 5.95 × 100 ± 7.30 × 10−1 | 1.48 × 101 ± 5.97 × 10−1 | 2.11 × 101 ± 7.01 × 10−1 ** | |
| 2.72 × 101 ± 8.73 × 10−1 | 2.17 × 101 ± 9.19 × 10−1 ** | 2.06 × 100 ± 8.87 × 10−1 | 2.49 × 100 ± 4.49 × 10−1 * | |
| 2.91 × 100 ± 5.65 × 10−1 | 1.54 × 100 ± 1.86 × 10−1 | 8.30 × 10−2 ± 7.32 × 10−3 | 6.89 × 10−2 ± 7.17 × 10−3 ** | |
| Sample | Protein Name | Accession No | Homology Model | Score | Identity | Gene Name |
|---|---|---|---|---|---|---|
| 3307↑ | Tubulin α-1A chain | gi|55977479 |  | 779 | 99.54 | Tuba1b |
| 3315↓ | Tubulin β-2A chain | gi|81885934 | 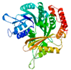 | 1020 | 100 | Tubb2a |
| 4422↑ | γ-actin | gi|809561 |  | 912 | 100 | Actg1 |
| 8116↓ | 14-3-3 protein ζ (Homo sapiens) | gi|148676868 |  | 644 | 100 | Ywhaz |
| 7119↓ | 14-3-3 protein ξ (Homosapiens) | gi|26344914 |  | 516 | 99.61 | Ywhae |
| 0314↑ | Heat shock protein HSP 90-β | gi|341941065 |  | 685 | 100 | Hsp90ab1 |
| 2814↓ | dihydropyrimidinase-related protein 2 | gi|568986628 |  | 1150 | 98.6 | Dpysl2 |
| 3311↓ | enolase 2, γ neuronal | gi|148667340 |  | 595 | 100 | Eno2 |
| Sample | Protein Name | KEGG Pathways |
|---|---|---|
| 3307 | α-tubulin | mmu04540, Gap junction|mmu04210, Apoptosis|mmu04145, Phagosome |
| 3155 | β-tubulin | mmu04540, Gap junction|mmu04145, Phagosome |
| 4422 | γ-actin | mmu04530, Tight junction|mmu04670, Leukocyte transendothelial migration|mmu05414, Dilated cardiomyopathy|mmu04015, Rap1 signaling pathway|mmu05416, Viral myocarditis|mmu05410, Hypertrophic cardiomyopathy (HCM)|mmu04510, Focal adhesion|mmu05205, Proteoglycans in cancer|mmu04520, Adherens junction|mmu04611, Platelet activation|mmu05100, Bacterial invasion of epithelial cells|mmu04210, Apoptosis|mmu04145, Phagosome|mmu05412, Arrhythmogenic right ventricular cardiomyopathy (ARVC)|mmu05132, Salmonella infection|mmu04390, Hippo signaling pathway|mmu04810, Regulation of actin cytoskeleton|mmu04921, Oxytocin signaling pathway|mmu04919, Thyroid hormone signaling pathway|mmu05164, Influenza A |
| 8116 | 14-3-3ζ | mmu05203, Viral carcinogenesis|mmu05161, Hepatitis B|mmu04114, Oocyte meiosis|mmu05169, Epstein-Barr virus infection|mmu04110, Cell cycle|mmu04151, PI3K-Akt signaling pathway|mmu04390, Hippo signaling pathway |
| 7119 | 14-3-3ε | mmu05203, Viral carcinogenesis|mmu04722, Neurotrophin signaling pathway|mmu04114, Oocyte meiosis|mmu05169, Epstein-Barr virus infection|mmu04110, Cell cycle|mmu04151, PI3K-Akt signaling pathway|mmu04390, Hippo signaling pathway |
| 0314 | HSP 90-β | mmu04915, Estrogen signaling pathway|mmu04914, Progesterone-mediated oocyte maturation|mmu04621, NOD-like receptor signaling pathway|mmu04612, Antigen processing and presentation|mmu05215, Prostate cancer|mmu04141, Protein processing in endoplasmic reticulum|mmu04151, PI3K-Akt signaling pathway|mmu05200, Pathways in cancer |
| 2814 | CRMP2 | mmu04360, Axon guidance |
| 3311 | ENO2 | mmu01100, Metabolic pathways|mmu01200, Carbon metabolism|mmu04066, HIF-1 signaling pathway|mmu01230, Biosynthesis of amino acids|mmu00010, Glycolysis/Gluconeogenesis|mmu03018, RNA degradation |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Cheng, C.; Baranenko, D.; Wang, J.; Li, Y.; Lu, W. Effects of Acanthopanax senticosus on Brain Injury Induced by Simulated Spatial Radiation in Mouse Model Based on Pharmacokinetics and Comparative Proteomics. Int. J. Mol. Sci. 2018, 19, 159. https://doi.org/10.3390/ijms19010159
Zhou Y, Cheng C, Baranenko D, Wang J, Li Y, Lu W. Effects of Acanthopanax senticosus on Brain Injury Induced by Simulated Spatial Radiation in Mouse Model Based on Pharmacokinetics and Comparative Proteomics. International Journal of Molecular Sciences. 2018; 19(1):159. https://doi.org/10.3390/ijms19010159
Chicago/Turabian StyleZhou, Yingyu, Cuilin Cheng, Denis Baranenko, Jiaping Wang, Yongzhi Li, and Weihong Lu. 2018. "Effects of Acanthopanax senticosus on Brain Injury Induced by Simulated Spatial Radiation in Mouse Model Based on Pharmacokinetics and Comparative Proteomics" International Journal of Molecular Sciences 19, no. 1: 159. https://doi.org/10.3390/ijms19010159
APA StyleZhou, Y., Cheng, C., Baranenko, D., Wang, J., Li, Y., & Lu, W. (2018). Effects of Acanthopanax senticosus on Brain Injury Induced by Simulated Spatial Radiation in Mouse Model Based on Pharmacokinetics and Comparative Proteomics. International Journal of Molecular Sciences, 19(1), 159. https://doi.org/10.3390/ijms19010159