Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds




Abstract

1. Introduction
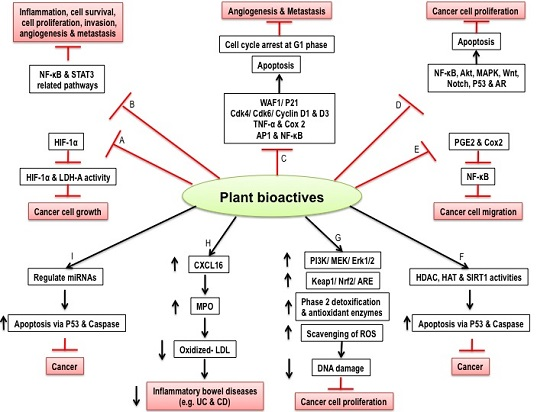
2. Inflammation, Cancer, and Regulation by Dietary Intakes
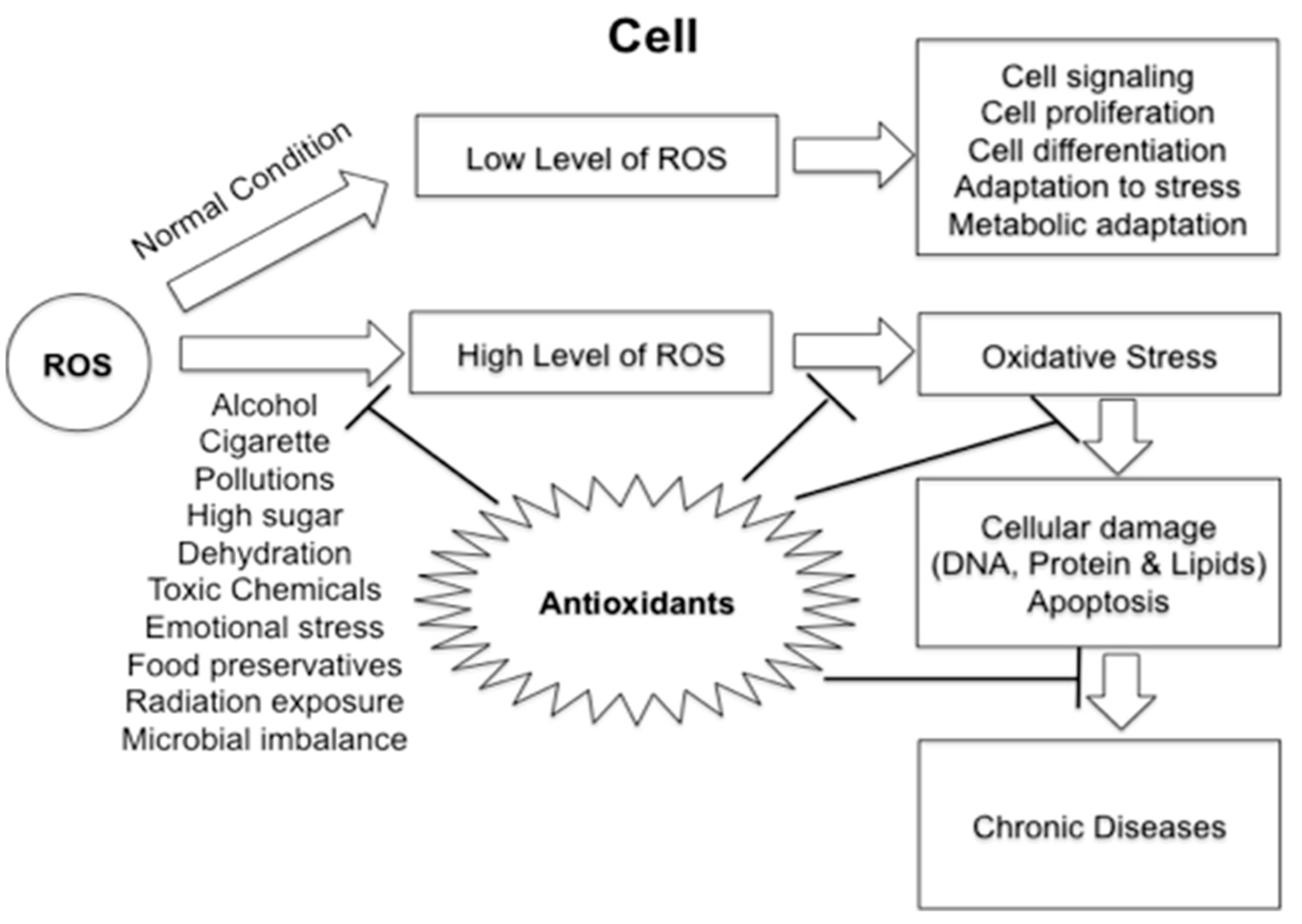
3. Plant Bioactives and Targeting Antioxidant Pathways
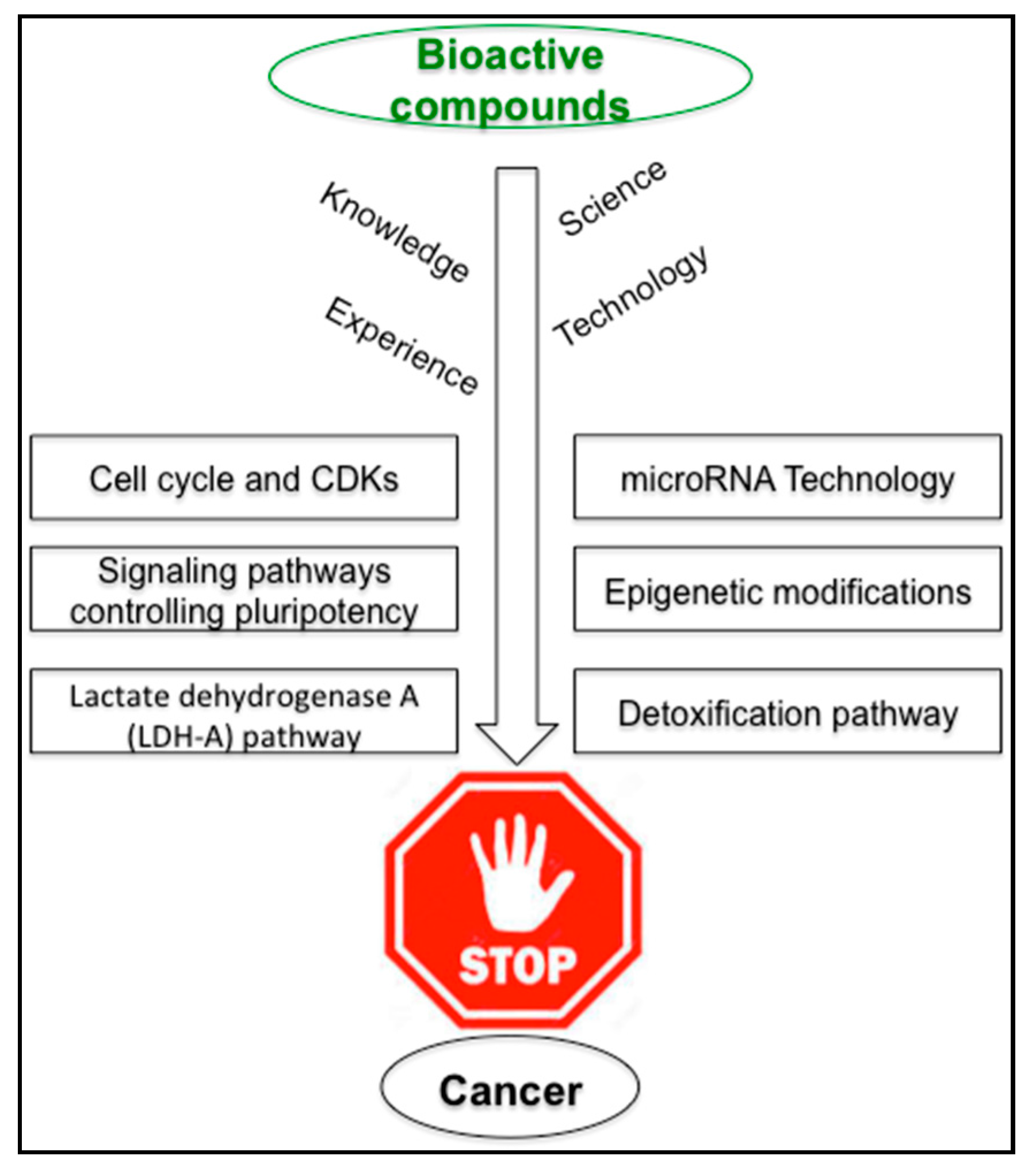
4. Arrest of Cancer Cell Cycle by Bioactive Compounds
5. Inhibition of Cancer Cell Proliferation and Migration by Plant Bioactives
6. Deregulation of Hypoxia and Glucose Metabolism in Cancer
7. Inflammatory Bowel Diseases and down Regulation of Immune System by Dietary Components
7.1. Inflammatory Bowel Diseases and Antioxidative Capacity of Carotenoids to Reduce Oxidative Stress and Inflammation
7.2. IBD and Immune-Modulating Activity of Carotenoids
8. Epigenetics and Cancer
9. MicroRNA, Nutrition and Cancer
10. Metabolic Stability of Plant Bioactives
11. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| NF-κB | Nuclear factor kappa B |
| IBD | Inflammatory bowel diseases |
| STAT3 | Signal transducer and activator of transcription 3 |
| MCF-7 | Michigan cancer foundation-7 |
| MCF-10A | Michigan cancer foundation-10A |
| LEF1 | Lymphoid enhancer-binding factor 1 |
| PTGS2 | Prostaglandin-endoperoxide synthase 2 (prostaglandin G/H synthase and cyclooxygenase) |
| PRKCE | Protein kinase C epsilon |
| ROS | Reactive oxygen species |
| TNF-α | Tumor necrosis factor alpha |
| Ap-1 | Activator protein-1 |
| CDKs | Cyclin-dependent kinases |
| PEITC | Phenethyl isothiocyanate |
| ATM | Ataxia telangiectasia mutated |
| ChK1 | Checkpoint kinase 1 |
| AR | Androgen receptor |
| MMPs | Matrix metalloproteinases (MMP-2 and MMP-9) |
| CK1α | Casein kinase-1a |
| SK3 β | Glycogen synthase kinase-3 β |
| LDH-A | Lactate dehydrogenase A LDH-A |
| HIF | Hypoxia inducible factor |
| PDH | Pyruvate dehydrogenase complex |
| GST | Glutathione S-transferase |
| UDP-GT | UDP-glucuronyl transferase |
| Nrf2 | Nuclear factor F-related factor 2 |
| Keap1 | Kelch-like ECH-associated protein 1 |
| ARE | Antioxidant responsive element |
| Keap1/Nrf2/ARE | Kelch ECH associating protein 1/NF-E2-related factor 2/Antioxidant Response Elements |
| I3C | Indole 3-carbinol |
| CD | Crohn’s disease |
| UC | Ulcerative colitis |
| GPR43 | G-protein coupled receptor 43 |
| CRP | C-Reactive protein |
| ILs | Interleukins |
| LDL | Low-density lipoprotein |
| MPO | Myeloperoxidase |
| ox-LDL | Oxidized-LDL |
| CXCL16 | Chemokine (C-X-C motif) ligand 16-a |
| IFNs | Interferons |
| GALT | Gut-associated lymphoid tissue |
| TLRs | Toll-like receptors |
| NOD2 | Nucleotide-binding oligomerization domain-containing protein 2 |
| COX | Cyclooxygenase |
| LP | Lamina propria |
| OS | Oxidative stress |
| HATs | Histone acetyl transferases |
| HDACs | Histone deacetylases |
| IP-10 | Inducible protein 10 |
| MIP-2 | Macrophage inflammatory protein 2 |
| DIM | Diindolylmethane |
| DADS | Diallyl disulfide |
| SAMC | S-allyl mercaptocysteine |
References
- Boerma, T.; Mathers, C.; AbouZahr, C.; Chatterji, S.; Hogan, D.; Stevens, D.; Mahanani, W.R.; Ho, J.; Rusciano, F.; Humphreys, G. Non communicable diseases Chapter 6. In Health in 2015: From MDGs, Millennium Development Goals to SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2015; pp. 142–143. [Google Scholar]
- Cooper, G.M. Cancer. In The Cell: A Molecular Approach, 2nd ed.; Sinauer Association: Sunderland, MA, USA, 2000. [Google Scholar]
- Cairns, J. The epidemiology of cancer. In Cancer: Science and Society; W.H. Freeman and Company: San Francisco, CA, USA, 1978; p. 41. [Google Scholar]
- National Research Council (US) Committee on Diet, Nutrition, and Cancer. Diet, Nutrition, and Cancer; National Academies Press: Washington, DC, USA, 1982. [Google Scholar]
- Xiao, J.; Capanoglu, E.; Jassbi, A.R.; Miron, A. The paradox of natural flavonoid C-glycosides and health benefits: When more occurrence is less research. Biotechnol. Adv. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; van Camp, J.; Capanoglu, E. Influence of different processing and storage conditions on in vitro bioaccessibility of polyphenols in black carrot jams and marmalades. Food Chem. 2015, 186, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113, 71S–88S. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Phytochemicals in cancer prevention and therapy: Truth or dare? Toxins 2010, 2, 517–551. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hosseini, A.; Ghorbani, A. Cancer therapy with phytochemicals: Evidence from clinical studies. Avicenna J. Phytomed. 2015, 5, 84–97. [Google Scholar] [PubMed] [PubMed Central]
- Hodges, R.E.; Minich, D.M. Modulation of metabolic detoxification pathways using foods and food-derived components: A scientific review with clinical application. J. Nutr. Metab. 2015, 2015, 760689. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Steinkellner, H.; Rabot, S.; Freywald, C.; Nobis, E.; Scharf, G.; Chabicovsky, M.; Knasmüller, S.; Kassie, F. Effects of cruciferous vegetables and their constituents on drug metabolizing enzymes involved in the bioactivation of DNA-reactive dietary carcinogens. Mutat. Res. 2001, 480–481, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jiang, S.; He, C.; Kimura, Y.; Yamashita, Y.; Ashida, H. Black soybean seed coat polyphenols prevent B (a) P-induced DNA damage through modulating drug-metabolizing enzymes in HepG2 cells and ICR mice. Mutat. Res. 2013, 752, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Devassy, J.G.; Nwachukwu, I.D.; Jones, P.J. Curcumin and cancer: Barriers to obtaining a health claim. Nutr. Rev. 2015, 73, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Sung, B.; Prasad, S.; Yadav, V.R.; Aggarwal, B.B. Cancer cell signaling pathways targeted by spice-derived nutraceuticals. Nutr. Cancer 2012, 64, 173–197. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gupta, S.C.; Kim, J.H.; Prasad, S.; Aggarwal, B.B. Regulation of survival, proliferation, invasion, angiogenesis, and metastasis of tumor cells through modulation of inflammatory pathways by nutraceuticals. Cancer Metastasis Rev. 2010, 29, 405–434. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Meeran, S.M.; Ahmed, A.; Tollefsbol, T.O. Epigenetic targets of bioactive dietary components for cancer prevention and therapy. Clin. Epigenet. 2010, 1, 101–116. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Aggarwal, B.B.; Vijayalekshmi, R.V.; Sung, B. Targeting inflammatory pathways for prevention and therapy of cancer: Short-Term Friend, Long-Term Foe. Clin. Cancer Res. 2009, 15, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Mao, R.; Yang, J. NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell 2013, 4, 176–185. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pavlick, K.P.; Laroux, F.S.; Fuseler, J.; Wolf, R.E.; Gray, L.; Hoffman, J.; Grisham, M.B. Role of reactive metabolites of oxygen and nitrogen in inflammatory bowel disease. Free Radic. Biol. Med. 2002, 33, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Rogler, G.; Brand, K.; Vogl, D.; Page, S.; Hofmeister, R.; Andus, T.; Knuechel, R.; Baeuerle, P.A.; Schölmerich, J.; Gross, V. Nuclear factor kappa-B is activated in macrophages and epithelial cells of inflamed intestinal mucosa. Gastroenterology 1998, 115, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Paliyath, G.; Bakovic, M.; Shetty, K. Functional Foods, Nutraceuticals and Degenerative Disease Prevention; Wiley-Blackwell: Oxford, UK, 2011; p. 392. [Google Scholar]
- Hakimuddin, F.; Paliyath, G.; Meckling, K. Selective cytotoxicity of a red grape wine flavonoid fraction against MCF-7 cells. Breast Cancer Res. Treat. 2004, 85, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Hakimuddin, F.; Paliyath, G.; Meckling, K. Treatment of mcf-7 breast cancer cells with a red grape wine polyphenol fraction results in disruption of calcium homeostasis and cell cycle arrest causing selective cytotoxicity. J. Agric. Food Chem. 2006, 54, 7912–7923. [Google Scholar] [CrossRef] [PubMed]
- Hakimuddin, F.; Tiwari, K.; Paliyath, G.; Meckling, K. Grape and wine polyphenols down-regulate the expression of signal transduction genes and inhibit the growth of estrogen receptor-negative mda-mb231 tumors in nu/nu mouse xenografts. Nutr. Res. 2008, 28, 702713. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.K.; Tiwari, K.; Correa-Betanzo, J.; Misran, A.; Chandrasekaran, R.; Paliyath, G. Biochemical basis for functional ingredient design from fruits. Annu. Rev. Food Sci. Technol. 2012, 3, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, V.; Ananga, A.; Tsolova, V. Recent advances and uses of grape flavonoids as nutraceuticals. Nutrients 2014, 6, 391–415. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Seymour, C.B.; Motherstill, C. Breast cancer causes and treatment: Where are we going wrong? Breast Cancer 2013, 5, 111–119. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Diplock, A.T.; Charleux, J.L.; Crozier-Willi, G.; Kok, F.J.; Rice-Evans, C.; Roberfroid, M.; Stahl, W.; Viña-Ribes, J. Functional food science and defence against reactive oxygen species. Br. J. Nutr. 1998, 80, S77–S112. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Strawberry as a health promoter: An evidence based review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, A.; Szostak-Wegierek, D. Flavonoids-food sources and health benefits. Rocz. Panstw. Zakl. Hig. 2014, 65, 79–85. [Google Scholar] [PubMed]
- Copple, I.M. The Keap1–Nrf2 cell defense pathway—A promising therapeutic target? Adv. Pharmacol. 2012, 63, 43–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Shan, Y. Sulforaphane retards the growth of UM-UC-3 xenographs, induces apoptosis, and reduces survivin in athymic mice. Nutr. Res. 2012, 32, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, S.H. Sulforaphane induces antioxidative and antiproliferative responses by generating reactive oxygen species in human bronchial epithelial BEAS-2B cells. J. Korean Med. Sci. 2011, 26, 1474–1482. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dinkova-Kostova, A.T.; Talalay, P. Relation of structure of curcumin analogs to their potencies as inducers of Phase 2 detoxification enzymes. Carcinogen 1999, 20, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Phamacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Wang, X.J. Induction of the Keap1/Nrf2/ARE pathway by oxidizable diphenols. Chem. Biol. Interact. 2011, 192, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, D.T.; Goldbohm, R.A.; van Poppel, G.; Verhagen, H.; van den Brandt, P.A. Epidemiological studies on Brassica vegetables and cancer risk. Cancer Epidemiol. Biomark. Prev. 1996, 5, 733–748. [Google Scholar] [PubMed]
- Jackson, S.J.T.; Singletary, K.W. Sulforaphane Inhibits human MCF-7 mammary cancer cell mitotic progression and tubulin polymerization. J. Nutr. 2004, 134, 2229–2236. [Google Scholar] [PubMed]
- Choi, S.; Singh, S.V. Bax and Bak are required for apoptosis induction by sulforaphane, a cruciferous vegetable–derived cancer chemopreventive agent. Cancer Res. 2005, 65, 2035–2043. [Google Scholar] [CrossRef] [PubMed]
- Myzak, M.M.; Dashwood, R.H. Chemoprotection by sulforaphane: Keep one eye beyond Keap1. Cancer Lett. 2006, 233, 208–218. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Finley, J.W.; Ip, C.; Lisk, D.J.; Davis, C.D.; Hintze, K.J.; Whanger, P.D. Cancer-protective properties of high-selenium broccoli. J. Agric. Food Chem. 2001, 49, 2679–2683. [Google Scholar] [CrossRef] [PubMed]
- Bostwick, D.G.; Alexander, E.E.; Singh, R.; Shan, A.; Qian, J.; Santella, R.M.; Oberley, L.W.; Yan, T.; Zhong, W.; Jiang, X.; et al. Antioxidant enzyme expression and reactive oxygen species damage in prostatic intraepithelial neoplasia and cancer. Cancer 2000, 89, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, Z.; Akhtar, N.; Anbazhagan, A.N.; Ramamurthy, S.; Shukla, M.; Haqqi, T.M. Polyphenol-rich pomegranate fruit extract (POMx) suppresses PMACI-induced expression of pro-inflammatory cytokinesby inhibiting the activation of MAP Kinases and NF-kappaB in human KU812 cells. J. Inflamm. 2009. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as cytokine modulators: A possible therapy for inflammation-related diseases. Int. J. Mol. Sci. 2016, 17, 921. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Danovi, S. Feeling the Heat-the Link between Inflammation and Cancer. Cancer Research UK, 1 February 2013. Available online: http://scienceblog.cancerresearchuk.org/2013/02/01/feeling-the-heat-the-link-between-inflammation-and-cancer/ (accessed on 25 September 2017).
- Jacob, J.K.; Paliyath, G. Composition and chemistry of functional foods and nutraceuticals: Influence on bioaccessibility and bioavailability. In Functional Foods, Nutraceuticals and Degenerative Disease Prevention; Paliyath, G., Bakovic, M., Shetty, K., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 113–144. [Google Scholar]
- Young, D.; Tsao, R.; Mine, Y. Nutraceuticals and antioxidant function. In Functional Foods, Nutraceuticals and Degenerative Disease Prevention; Paliyath, G., Bakovic, M., Shetty, K., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 75–112. [Google Scholar]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Takei, M.; Kobayashi, M.; Herndon, D.N.; Pollard, R.B.; Suzuki, F. Glycyrrhizin inhibits the manifestations of anti-inflammatory responses that appear in association with systemic inflammatory response syndrome (SIRS)-like reactions. Cytokine 2006, 35, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Y.; Khor, T.O.; Saw, C.L.; Loh, S.C.; Chen, A.I.; Lim, S.S.; Park, J.H.; Cai, L.; Kong, A.N. Anti-inflammatory/Anti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemum zawadskii and licorice Glycyrrhiza uralensis. AAPS J. 2011, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim, J.Y.; Park, S.J.; Yun, K.J.; Cho, Y.W.; Park, H.J.; Lee, K.T. Isoliquiritigenin isolated from the roots of Glycyrrhiza uralensis inhibits LPS-induced iNOS and COX-2 expression via the attenuation of NF-kappaB in RAW 264.7 macrophages. Eur. J. Pharmacol. 2008, 584, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Prawan, A.; Saw, C.L.; Khor, T.O.; Keum, Y.S.; Yu, S.; Hu, L. Anti-NF-kappaB and anti-inflammatory activities of synthetic isothio- cyanates: Effect of chemical structures and cellular signaling. Chem. Biol. Interact. 2009, 179, 202–211. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jeong, W.S.; Keum, Y.S.; Chen, C.; Jain, M.R.; Shen, G.; Kim, J.H.; Li, W.; Kong, A.N. Differential expression and stability of endogenous nuclear factor E2-related factor 2 (Nrf2) by natural chemopreventive compounds in HepG2 human hepatoma cells. J. Biochem. Mol. Biol. 2005, 38, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.; Huang, M.T.; Kwon, K.H.; Chan, J.Y.; Reddy, B.S.; Kong, A.N. Nrf2-deficient mice have an increased susceptibility to dextran sulfate sodium-induced colitis. Cancer Res. 2006, 66, 11580–11584. [Google Scholar] [CrossRef] [PubMed]
- Kawamori, T.; Lubet, R.; Steele, V.E.; Kelloff, G.J.; Kaskey, R.B.; Rao, C.V.; Reddy, B.S. Chemopreventive effect of curcumin, a naturally occurring anti-inflammatory agent, during the promotion/progression stages of colon cancer. Cancer Res. 1999, 59, 597–601. [Google Scholar] [PubMed]
- Aggarwal, B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef] [PubMed]
- Cen, L.; Hutzen, B.; Ball, S.; DeAngelis, S.; Chen, C.L.; Fuchs, J.R.; Li, C.; Li, P.K.; Lin, J. New structural analogues of curcumin exhibit potent growth suppressive activity in human colorectal carcinoma cells. BMC Cancer 2009, 9, 99. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Das, L.; Vinayak, M. Long term effect of curcumin in restoration of tumour suppressor p53 and phase-II antioxidant enzymes via activation of NRF2 signalling and modulation of inflammation in prevention of cancer. PLoS ONE 2015, 10, e0124000. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Khan, M.A.; Gahlot, S.; Majumdar, S. Oxidative stress induced by curcumin promotes the death of cutaneous T-cell lymphoma (HuT-78) by disrupting the function of several molecular targets. Mol. Cancer Ther. 2012, 11, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.Y.; Bradlow, H.L.; Sepkovic, D.; Mehl, S.; Mailman, J.; Osborne, M.P. Dose-ranging study of indole-3-carbinol for breast cancer prevention. J. Cell. Biochem. Suppl. 1997, 28–29, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Sowers, M.R.; Crawford, S.; McConnell, D.S.; Randolph, J.S.; Gold, E.B.; Wilkins, M.K.; Lasley, B. Selected diet and lifestyle factors are associated with estrogen metabolites in a multiracial/ethnic population of women. J. Nutr. 2006, 136, 1588–1595. [Google Scholar] [PubMed]
- Ashok, B.T.; Chen, Y.; Liu, X.; Bradlow, H.L.; Mittelman, A.; Tiwari, R.K. Abrogation of estrogen-mediated cellular and biochemical effects by indole-3-carbinol. Nutr. Cancer 2001, 41, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Chinni, S.R.; Li, Y.; Upadhyay, S.; Koppolu, P.K.; Sarkar, F.H. Indole-3-carbinol (i3c) induced cell growth inhibition, G1 cell cycle arrest and apoptosis in prostate cancer cells. Oncogene 2001, 20, 2927–2936. [Google Scholar] [CrossRef] [PubMed]
- Rosen, C.A.; Woodson, G.E.; Thompson, J.W.; Hengesteg, A.P.; Bradlow, H.L. Preliminary results of the use of indole-3-carbinol for recurrent respiratory papillomatosis. Otolaryngol. Head Neck Surg. 1998, 118, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.; Blanchette, M.; Barrette, S.; Moghrabi, A.; Béliveau, R. Inhibition of cancer cell proliferation and suppression of TNF-induced activation of NFkappaB by edible berry juice. Anticancer Res. 2007, 27, 937–948. [Google Scholar] [PubMed]
- Huang, C.; Zhang, D.; Li, J.; Tong, Q.; Stoner, G.D. Differential inhibition of UV-induced activation of NF kappa B and AP-1 by extracts from black raspberries, strawberries, and blueberries. Nutr. Cancer 2007, 58, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Li, L.; Celver, J.; Killian, C.; Kovoor, A.; Seeram, N.P. Effects of fruit ellagitannin extracts, ellagic acid, and their colonic metabolite, urolithin A, on Wnt signaling. J. Agric. Food Chem. 2010, 58, 3965–3969. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Poulose, S.M.; Harris, E.D.; Patil, B.S. Antiproliferative effects of citrus limonoids against human neuroblastoma and colonic adenocarcinoma cells. Nutr. Cancer 2006, 56, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasha, G.K.; Girennavar, B.; Patil, B.S. Radical scavenging activities of Rio Red grapefruits and Sour orange fruit extracts in different in vitro model systems. Bioresour. Technol. 2008, 99, 4484–4494. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasha, G.K.; Jadegoud, Y.; Nagana Gowda, G.A.; Patil, B.S. Bioactive compounds from sour orange inhibit colon cancer cell proliferation and induce cell cycle arrest. J. Agric. Food Chem. 2010, 58, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, F.; Darbon, J.M. Effects of structurally related flavonoids on cell cycle progression of human melanoma cells: Regulation of cyclin-dependent kinases CDK2 and CDK1. Biochem. Pharmacol. 2001, 61, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Chen, W.J.; Lin-Shiau, S.Y.; Ho, C.T.; Lin, J.K. Tangeretin induces cell-cycle G1 arrest through inhibiting cyclin-dependent kinases 2 and 4 activities as well as elevating Cdk inhibitors p21 and p27 in human colorectal carcinoma cells. Carcinogenesis 2002, 23, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Siess, M.H.; Le Bon, A.M.; Canivenc-Lavier, M.C.; Suschetet, M. Mechanisms involved in the chemoprevention of flavonoids. Biofactors 2000, 12, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.; Morrice, P. Antioxidant capacity of flavonoids in hepatic microsomes is not reflected by antioxidant effects in vivo. Oxid. Med. Cell. Longev. 2012, 2012, 165127. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tan, B.; Liu, Y.; Chang, K.; Lim, B.K.; Chiu, G.N. Perorally active nanomicellar formulation of quercetin in the treatment of lung cancer. Int. J. Nanomed. 2012, 7, 651–661. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jakubikova, J.; Cervi, D.; Ooi, M.; Kim, K.; Nahar, S.; Klippel, S.; Cholujova, D.; Leiba, M.; Daley, J.F.; Delmore, J.; et al. Anti-tumor activity and signaling events triggered by the isothiocyanates, sulforaphane and PEITC in multiple myeloma. Haematologica 2011, 96, 1170–1179. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yan, H.; Zhu, Y.; Liu, B.; Wu, H.; Li, Y.; Wu, X.; Zhou, Q.; Xu, K. Mitogen-activated protein kinase mediates the apoptosis of highly metastatic human non-small cell lung cancer cells induced by isothiocyanates. Br. J. Nutr. 2011, 106, 1779–1791. [Google Scholar] [CrossRef] [PubMed]
- Sa, G.; Das, T. Anti cancer effects of curcumin: Cycle of life and death. Cell Div. 2008, 3, 14. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sahu, R.P.; Batra, S.; Srivastava, S.K. Activation of ATM/Chk1 by curcumin causes cell cycle arrest and apoptosis in human pancreatic cancer cells. Br. J. Cancer 2009, 100, 1425–1433. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. NF-kappaB in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Birchmeier, W. Wnt signalling and its impact on development and cancer. Nat. Rev. Cancer 2008, 8, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Sebolt-Leopold, J.S.; Herrera, R. Targeting the mitogen-activated protein kinase cascade to treat cancer. Nat. Rev. Cancer 2004, 4, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Stiewe, T. The p53 family in differentiation and tumorigenesis. Nat. Rev. Cancer 2007, 7, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Bao, B.; Ahmad, A.; Sarkar, F.H. Induction of cancer cell death by isoflavone: The role of multiple signaling pathways. Nutrients 2011, 3, 877–896. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Singh, T.; Katiyar, S.K. Honokiol inhibits non-small cell lung cancer cell migration by targeting PGE2-mediated activation of β-catenin signaling. PLoS ONE 2013, 8, e60749. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, Y.; Wang, Z.; Kong, D.; Li, R.; Sarkar, S.H.; Sarkar, F.H. Regulation of Akt/FOXO3a/GSK-3beta/AR signaling network by isoflavone in prostate cancer cells. J. Biol. Chem. 2008, 283, 27707–27716. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Sarkar, F.H.; Li, Y.; Wang, Z.; Kong, D. Cellular signaling perturbation by natural products. Cell Signal. 2009, 21, 1541–1547. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tarapore, R.S.; Siddiqui, I.A.; Mukhtar, H. Modulation of Wnt/β-catenin signaling pathway by bioactive food components. Carcinogenesis 2012, 33, 483–491. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Amado, N.G.; Fonseca, B.F.; Cerqueira, D.M.; Neto, V.M.; Abreu, J.G. Flavonoids: Potential Wnt/beta-catenin signaling modulators in cancer. Life Sci. 2011, 89, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Kolev, Y.; Uetake, H.; Takagi, Y.; Sugihara, K. Lactate dehydrogenase-5 (LDH-5) expression in human gastric cancer: Association with hypoxia-inducible factor (HIF-1alpha) pathway, angiogenic factors production and poor prognosis. Ann. Surg. Oncol. 2008, 15, 2336–2344. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Valera, V.A.; Merino, M.J.; Amato, A.M.; Signoretti, S.; Linehan, W.M.; Sukhatme, V.P.; Seth, P. LDH-A inhibition, a therapeutic strategy for treatment of hereditary leiomyomatosis and renal cell cancer. Mol. Cancer Ther. 2009, 8, 626–635. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McFate, T.; Mohyeldin, A.; Lu, H.; Thakar, J.; Henriques, J.; Halim, N.D.; Wu, H.; Schell, M.J.; Tsang, T.M.; Teahan, O.; et al. Pyruvate dehydrogenase complex activity controls metabolic and malignant phenotype in cancer cells. J. Biol. Chem. 2008, 283, 22700–22708. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, Z.; Wang, D.; Han, S.; Wang, N.; Mo, F.; Loo, T.Y.; Shen, J.; Huang, H.; Chen, J. Bioactivity-guided identification and cell signaling technology to delineate the lactate dehydrogenase A inhibition effects of Spatholobus suberectus on breast cancer. PLoS ONE 2013, 8, e56631. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cerella, C.; Radogna, F.; Dicato, M.; Diederich, M. Natural compounds as regulators of the cancer cell metabolism. Int. J. Cell Biol. 2013, 2013, 16. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Guo, L.D.; Chen, X.J.; Hu, Y.H.; Yu, Z.J.; Wang, D.; Liu, J.Z. Curcumin inhibits proliferation and induces apoptosis of human colorectal cancer cells by activating the mitochondria apoptotic pathway. Phytother. Res. 2012, 27, 422–430. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kaser, A.; Zeissig, S.; Blumberg, R.S. Inflammatory bowel disease. Annu. Rev. Immunol. 2010, 28, 573–621. [Google Scholar] [CrossRef] [PubMed]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, C.N.; Nabalamba, A. Hospitalization, surgery, and readmission rates of IBD in Canada: A population-based study. Am. J. Gastroenterol. 2006, 101, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Baniyash, M. Chronic inflammation, immunosuppression and cancer: New insights and outlook. Semin. Cancer Biol. 2006, 16, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Head, K.A.; Jurenka, J.S. Inflammatory bowel disease Part 1: Ulcerative colitis—Pathophysiology and conventional and alternative treatment options. Altern. Med. Rev. 2003, 8, 247–283. [Google Scholar] [PubMed]
- Head, K.; Jurenka, J.S. Inflammatory bowel disease. Part II: Crohn’s disease—Pathophysiology and conventional and alternative treatment options. Altern. Med. Rev. 2004, 9, 360–401. [Google Scholar] [PubMed]
- Shapiro, H.; Singer, P.; Halpern, Z.; Bruck, R. Polyphenols in the treatment of inflammatory bowel disease and acute pancreatitis. Gut 2007, 56, 426–435. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Aghdassi, E.; Wendland, B.E.; Stapleton, M.; Raman, M.; Allard, J.P. Adequacy of nutritional intake in a Canadian population of patients with Crohn’s disease. J. Am. Diet. Assoc. 2007, 107, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- González-Gallego, J.; García-Mediavilla, M.V.; Sánchez-Campos, S.; Tuñón, M.J. Fruit polyphenols, immunity and inflammation. Br. J. Nutr. 2010, 104, S15–S27. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alonso, M.; Jacobs, E.; Raybould, A.; Nickson, T.E.; Sowig, P.; Willekens, H.; Van der Kouwe, P.; Layton, R.; Amijee, F.; Fuentes, A.M.; et al. A tiered system for assessing the risk of genetically modified plants to non-target organisms. Environ. Biosaf. Res. 2006, 5, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Wu, X.; Patterson, K.M.; Barnes, J.; Carter, S.G.; Scherwitz, L.; Beaman, R.; Endres, J.R.; Schauss, A.G. In vitro and in vivo antioxidant and anti-inflammatory capacities of an antioxidant-rich fruit and berry juice blend. Results of a pilot and randomized, double-blinded, placebo-controlled, crossover study. J. Agric. Food Chem. 2008, 56, 8326–8333. [Google Scholar] [CrossRef] [PubMed]
- Holt, E.M.; Steffen, L.M.; Moran, A.; Basu, S.; Steinberger, J.; Ross, J.A.; Hong, C.P.; Sinaiko, A.R. Fruit and vegetable consumption and its relation to markers of inflammation and oxidative stress in adolescents. J. Am. Diet. Assoc. 2009, 109, 414–421. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Karlsen, A.; Retterstøl, L.; Laake, P.; Paur, I.; Kjolsrud-Bohn, S.; Sandvik, L.; Blomhoff, R. Anthocyanins inhibit, Nuclear Factor kappa B-activation in monocytes and reduce plasma concentration of pro-inflammatory mediators in healthy adults. J. Nutr. 2007, 137, 1951–1954. [Google Scholar] [PubMed]
- Kelley, D.S.; Rasooly, R.; Jacob, R.A.; Kader, A.A.; Mackey, B.E. Consumption of Bing sweet cherries lowers circulating concentrations of inflammation markers in healthy men and women. J. Nutr. 2006, 136, 981–986. [Google Scholar] [PubMed]
- Helmersson, J.; Arnlöv, J.; Larsson, A.; Basu, S. Low dietary intake of beta-carotene, alpha-tocopherol and ascorbic acid is associated with increased inflammatory and oxidative stress status in a Swedish cohort. Br. J. Nutr. 2009, 101, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.R.; Katz, S.; Kirshoff, R. Curcumin therapy in inflammatory bowel disease: A pilot study. Dig. Dis. Sci. 2005, 50, 2191–2193. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Iida, T.; Takeuchi, K.; Watanabe, F.; Maruyama, Y.; Andoh, A.; Tsujikawa, T.; Fujiyama, Y.; Mitsuyama, K.; Sata, M.; et al. Curcumin maintenance therapy for ulcerative colitis: Randomized, multicenter, double-blind, placebo-controlled trial. Clin. Gastroenterol. Hepatol. 2006, 4, 1502–1506. [Google Scholar] [CrossRef] [PubMed]
- Lih-Brody, L.; Powell, S.R.; Collier, K.P.; Reddy, G.M.; Cerchia, R.; Kahn, E.; Weissman, G.S.; Katz, S.; Floyd, R.A.; McKinley, M.J.; et al. Increased oxidative stress and decreased antioxidant defenses in mucosa of inflammatory bowel disease. Dig. Dis. Sci. 1996, 41, 2078–2086. [Google Scholar] [CrossRef] [PubMed]
- Vandana, S.; Ram, S.; Ilavazhagan, M.; Kumar, G.D.; Banerjee, P.K. Comparative cytoprotective activity of vitamin C, E and beta-carotene against chromium induced oxidative stress in murine macrophages. Biomed. Pharmacother. 2006, 60, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Castelluccio, C.; Tijburg, L.; Rice-Evans, C. The antioxidant properties of theaflavins and their gallate esters--radical scavengers or metal chelators? FEBS Lett. 1996, 392, 40–44. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, T.M.; Tchao, R.; Harrison, E.H.; Morel, D.W. Carotenoids normally present in serum inhibit proliferation and induce differentiation of a human monocyte/macrophage cell line (U937). J. Nutr. 2005, 135, 160–164. [Google Scholar] [PubMed]
- Kawakami, Y.; Okada, H.; Murakami, Y.; Kawakami, T.; Ueda, Y.; Kunii, D.; Sakamoto, Y.; Shiratori, Y.; Okita, M. Dietary intake, neutrophil fatty acid profile, serum antioxidant vitamins and oxygen radical absorbance capacity in patients with ulcerative colitis. J. Nutr. Sci. Vitaminol. 2007, 53, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Adhami, V.M.; Syed, D.N.; Khan, N.; Afaq, F. Phytochemicals for prevention of solar ultraviolet radiation-induced damages. Photochem. Photobiol. 2008, 84, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Agarwal, S. Role of antioxidant lycopene in cancer and heart disease. J. Am. Coll. Nutr. 2000, 19, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Jialal, I.; Grundy, S.M. Influence of antioxidant vitamins on LDL oxidation. Ann. N. Y. Acad. Sci. 1992, 669, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Itabe, H.; Mori, M.; Fujimoto, Y.; Higashi, Y.; Takano, T. Minimally modified LDL is an oxidized LDL enriched with oxidized phosphatidylcholines. J. Biochem. 2003, 134, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.D.; Ball, J.M.; Sundar, S.; Coyle, P.; Howarth, G.S. The role of zinc and metallothionein in the dextran sulfate sodium-induced colitis mouse model. Dig. Dis. Sci. 2007, 52, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Lehrke, M.; Konrad, A.; Schachinger, V.; Tillack, C.; Seibold, F.; Stark, R.; Parhofer, I.G.; Broedl, U.C. CXCL16 is a surrogate marker of inflammatory bowel disease. Scand. J. Gastroenterol. 2008, 43, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, A.; Skibsted, L.H. Relative stability of carotenoid radical cations and homologue tocopheroxyl radicals. A real time kinetic study of antioxidant hierarchy. FEBS Lett. 1997, 417, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Sommerburg, O.; Langhans, C.D.; Arnhold, J.; Leichsenring, M.; Salerno, C.; Crifò, C.; Hoffmann, G.F.; Debatin, K.M.; Siems, W.G. Beta-carotene cleavage products after oxidation mediated by hypochlorous acid—A model for neutrophil-derived degradation. Free Radic. Biol. Med. 2003, 35, 1480–1490. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Stahl, W.; Sundquist, A.R. Antioxidant functions of vitamins. Vitamins E and C, beta-carotene, and other carotenoids. Ann. N. Y. Acad. Sci. 1992, 368, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E. Tomatoes, tomato-based products, lycopene, and cancer: Review of the epidemiologic literature. J. Natl. Cancer Inst. 1999, 91, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.; Periago, M.J.; Böhm, V.; Berruezo, G.R. Influence of lycopene and vitamin C from tomato juice on biomarkers of oxidative stress and inflammation. Br. J. Nutr. 2008, 99, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The role of carotenoids in the prevention of human pathologies. Biomed. Pharmacother. 2004, 58, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Haller, D. Intestinal epithelial cell signalling and chronic inflammation: From the proteome to specific molecular mechanisms. Mutat. Res. 2007, 622, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Schottelius, A.J.; Baldwin, A.S., Jr. A role for transcription factor NF-kappa B in intestinal inflammation. Int. J. Colorectal Dis. 1999, 14, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.; Nikolaus, S.; Hampe, J. Activation of nuclear factor kappa B inflammatory bowel disease. Gut 1998, 42, 477–484. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rogler, G.; Hausmann, M.; Vogl, D.; Aschenbrenner, E.; Andus, T.; Falk, W.; Andreesen, R.; Schölmerich, J.; Gross, V. Isolation and phenotypic characterization of colonic macrophages. Clin. Exp. Immunol. 1998, 112, 205–215. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Stronati, L.; Negroni, A.; Merola, P.; Pannone, V.; Borrelli, O.; Cirulli, M.; Annese, V.; Cucchiara, S. Mucosal NOD2 expression and NF-kappaB activation in pediatric Crohn’s disease. Inflamm. Bowel Dis. 2008, 14, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Mannick, E.E.; Bravo, L.E.; Zarama, G.; Realpe, J.L.; Zhang, X.J.; Ruiz, B.; Fontham, E.T.; Mera, R.; Miller, M.J.; Correa, P. Inducible nitric oxide synthase, nitrotyrosine, and apoptosis in Helicobacter pylori gastritis: Effect of antibiotics and antioxidants. Cancer Res. 1996, 56, 3238–3243. [Google Scholar] [PubMed]
- Bai, S.K.; Lee, S.J.; Na, H.J.; Ha, K.S.; Han, J.A.; Lee, H.; Kwon, Y.G.; Chung, C.K.; Kim, Y.M. beta-Carotene inhibits inflammatory gene expression in ipopolysaccharide-stimulated macrophages by suppressing redox-based NF-kappaB activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Park, B.; Goel, A.; Aggarwal, B.B. Epigenetic changes induced by curcumin and other natural compounds. Genes. Nutr. 2011, 6, 93–108. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Choi, S.W.; Friso, S. Epigenetics: A New Bridge between Nutrition and Health. Adv. Nutr. 2010, 1, 8–16. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Myzak, M.C.; Karplus, P.A.; Chung, F.L.; Dashwood, R.H. A novel mechanism of chemoprotection by sulforaphane: Inhibition of histone deacetylase. Cancer Res. 2004, 64, 5767–5774. [Google Scholar] [CrossRef] [PubMed]
- Myzak, M.C.; Dashwood, W.M.; Orner, G.A.; Ho, E.; Dashwood, R.H. Sulforaphane inhibits histone deacetylase in vivo and suppresses tumorigenesis in Apc-minus mice. FASEB J. 2006, 20, 506–508. [Google Scholar] [PubMed] [PubMed Central]
- Vahid, F.; Zand, H.; Nosrat-Mirshekarlou, E.; Najafi, R.; Hekmatdoost, A. The role dietary of bioactive compounds on the regulation of histone acetylases and deacetylases. Gene 2015, 562, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.K.; Chen, Q.; Fu, J.; Shankar, S.; Srivastava, R.K. Resveratrol inhibits growth of orthotopic pancreatic tumors through activation of FOXO transcription factors. PLoS ONE 2011, 6, e25166. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols causes cell cycle arrest and apoptosis in prostate cancer cells by suppressing class I histone deacetylases. Carcinogenesis 2012, 33, 377–384. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ruiz, P.A.; Braune, A.; Holzlwimmer, G.; Quintanilla-Fend, L.; Haller, D. Quercetin inhibits TNF-induced NF-kappaB transcription factor recruitment to proinflammatory gene promoters in murine intestinal epithelial cells. J. Nutr. 2007, 137, 1208–1215. [Google Scholar] [PubMed]
- Priyadarsini, R.V.; Vinothini, G.; Murugan, R.S.; Manikandan, P.; Nagini, S. The flavonoid quercetin modulates the hallmark capabilities of hamster buccal pouch tumors. Nutr. Cancer 2011, 63, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Guo, B. Chemopreventive agent 3,3′-diindolylmethane selectively induces proteasomal degradation of class I histone deacetylases. Cancer Res. 2010, 70, 646–654. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Druesne, N.; Pagniez, A.; Mayeur, C.; Thomas, M.; Cherbuy, C.; Duee, P.H.; Martel, P.; Chaumontet, C. Repetitive treatments of colon HT-29 cells with diallyl disulfide induce a prolonged hyperacetylation of histone H3 K14. Ann. N. Y. Acad. Sci. 2004, 1030, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of posttranscriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Vislovukh, A.; Vargas, T.R.; Polesskaya, A.; Groisman, I. Role of 3’-untranslated region translational control in cancer development, diagnostics and treatment. World J. Biol. Chem. 2014, 26, 40–57. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.; Srivastava, A. Comprehensive review of genetic association studies and meta-analyses on miRNA polymorphisms and cancer risk. PLoS ONE 2012, 7, e50966. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ferdin, J.; Kunej, T.; Calin, G.A. Non-coding RNAs: Identification of cancer- associated microRNAs by gene profiling. Technol. Cancer Res. Treat. 2010, 9, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Ferracin, M.; Pedriali, M.; Veronese, A.; Zagatti, B.; Gafà, R.; Magri, E.; Lunardi, M.; Munerato, G.; Querzoli, G.; Maestri, I.; et al. MicroRNA profiling for the identification of cancers with unknown primary tissue-of-origin. J. Pathol. 2011, 225, 43–53. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ross, S.A.; Davis, C.D. MicroRNA, nutrition, and cancer prevention. Adv. Nutr. 2011, 2, 472–485. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kent, O.A.; Mendell, J.T. A small piece in the cancer puzzle: MicroRNAs as tumor suppressors and oncogenes. Oncogene 2006, 25, 6188–6196. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Ruan, K.; Fang, X.; Ouyang, G. MicroRNAs: Novel regulators in the hallmarks of human cancer. Cancer Lett. 2009, 285, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Tsang, W.P.; Kwok, T.T. Epigallocatechin gallate up-regulation of miR-16 and induction of apoptosis in human cancer cells. J. Nutr. Biochem. 2010, 21, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Estrov, Z.; Ji, Y.; Coombes, K.R.; Harris, D.H.; Kurzrock, R. Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Mol. Cancer Ther. 2008, 7, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cao, Y.; Sun, J.; Zhang, Y. Curcumin reduces the expression of Bcl-2 by upregulating miR-15a and miR-16 in MCF-7 cells. Med. Oncol. 2010, 27, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Mudduluru, G.; George-William, J.N.; Muppala, S.; Asangani, I.A.; Regalla, K.; Nelson, L.D.; Allgayer, H. Curcumin regulates miR-21 expression and inhibits invasion and metastasis in colorectal cancer. Biosci. Rep. 2011, 31, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Alder, H.; Volinia, S.; Delmas, D.; Latruffe, N.; Croce, C.M. Resveratrol modulates the levels of microRNAs targeting genes encoding tumor suppressors and effectors of TGF𝛽 signaling pathway in SW480 cells. Biochem. Pharmacol. 2010, 80, 2057–2065. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, P.; Liang, H.; Xia, Q.; Li, P.; Kong, H.; Lei, P.; Wang, S.; Tu, Z. Resveratrol induces apoptosis of pancreatic cancers cells by inhibiting miR-21 regulation of BCL-2 expression. Clin. Transl. Oncol. 2013, 15, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Kumazaki, M.; Noguchi, S.; Yasui, Y.; Iwasaki, J.; Shinohara, H.; Yamada, N.; Akao, Y. Anti-cancer effects of naturally occurring compounds through modulation of signal transduction and miRNA expression in human colon cancer cells. J. Nutr. Biochem. 2013, 24, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Vandenboom, T.G., II; Kong, D.; Wang, Z.; Ali, S.; Philip, P.A.; Sarkar, F.H. Up-regulation of miR-200 and let-7 by natural agents leads to the reversal of epithelial-to-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells. Cancer Res. 2009, 69, 6704–6712. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jin, Y. 3,3’-Diindolylmethane inhibits breast cancer cell growth via miR-21-mediated Cdc25A degradation. Mol. Cell. Biochem. 2011, 358, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.H.; Wang, X.; Feng, Q. EGCG enhances the efficacy of cisplatin by downregulating hsa-miR-98-5p in NSCLC A549 cells. Nutr. Cancer 2014, 66, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.K.; Arora, S.; Averett, C.; Singh, S.; Singh, A.P. Modulation of microRNAs by phytochemicals in cancer: Underlying mechanisms and translational significance. Biomed. Res. Int. 2015, 2015, 848710. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, Y.; Zaman, M.S.; Deng, G.; Majid, S.; Saini, S.; Liu, J.; Tanaka, Y.; Dahiya, R. MicroRNAs 221/222 and genistein-mediated regulation of ARHI tumor suppressor gene in prostate cancer. Cancer Prev. Res. 2011, 4, 76–86. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xu, L.; Xiang, J.; Shen, J.; Zou, X.; Zhai, S.; Yin, Y.; Li, P.; Wang, X.; Sun, Q. Oncogenic microRNA-27a is a target for genistein in ovarian cancer cells. Anticancer Agents Med. Chem. 2013, 13, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Del Follo-Martinez, A.; Banerjee, N.; Li, X.; Safe, S.; Mertens-Talcott, S. Resveratrol and quercetin in combination have anticancer activity in colon cancer cells and repress oncogenic microRNA-27a. Nutr. Cancer 2013, 65, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Appari, M.; Babu, K.R.; Kaczorowski, A.; Gross, W.; Herr, I. Sulforaphane, quercetin and catechins complement each other in elimination of advanced pancreatic cancer by miRlet-7 induction and K-ras inhibition. Int. J. Oncol. 2014, 45, 1391–1400. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Correa-Betanzo, J.; Padmanabhan, P.; Corredig, M.; Subramanian, J.; Paliyath, G. Complex formation of blueberry (Vaccinium angustifolium) anthocyanins during freeze-drying and its influence on their biological activity. J. Agric. Food Chem. 2015, 25, 2935–2946. [Google Scholar] [CrossRef] [PubMed]
- Abuajah, C.I.; Ogbonna, A.C.; Osuji, C.M. Functional components and medicinal properties of food. J. Food Sci. Technol. 2015, 52, 2522–2529. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Biesalski, H.K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive compounds: Definition and assessment of activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | miRNA | Cancer | References |
|---|---|---|---|
| Curcumin | miR-22, miR-15a, miR-16, miR-21 | Pancreatic cancer, Breast cancer, Colon cancer | [162,163,164] |
| Diindolylmethane (DIM) | miR-200, let-7, miR-21 | Pancreatic cancer, Breast cancer | [168,169] |
| EGCG | miR-98-5p, miR-13, miR-48, miR-16, miR-21 | Lung cancer, HCC, Prostate cancer, Pancreatic cancer | [161,167,170,171] |
| Genistein | miR-221, miR-222, miR-27a | Prostate cancer, Ovarian cancer | [172,173] |
| Quercetin | miR-27a, Let-7 | Colorectal cancer, Pancreatic cancer | [174,175] |
| Resveratrol | miR-17, miR-21, miR-25, miR-92a-2, miR-103-1 and miR-103-2, miR-663, miR-34a | Colon cancer, Pancreatic cancer | [165,166,167] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nosrati, N.; Bakovic, M.; Paliyath, G. Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds. Int. J. Mol. Sci. 2017, 18, 2050. https://doi.org/10.3390/ijms18102050
Nosrati N, Bakovic M, Paliyath G. Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds. International Journal of Molecular Sciences. 2017; 18(10):2050. https://doi.org/10.3390/ijms18102050
Chicago/Turabian StyleNosrati, Nagisa, Marica Bakovic, and Gopinadhan Paliyath. 2017. "Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds" International Journal of Molecular Sciences 18, no. 10: 2050. https://doi.org/10.3390/ijms18102050
APA StyleNosrati, N., Bakovic, M., & Paliyath, G. (2017). Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds. International Journal of Molecular Sciences, 18(10), 2050. https://doi.org/10.3390/ijms18102050