Praeruptorin A Inhibits Human Cervical Cancer Cell Growth and Invasion by Suppressing MMP-2 Expression and ERK1/2 Signaling


 ,
,  and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
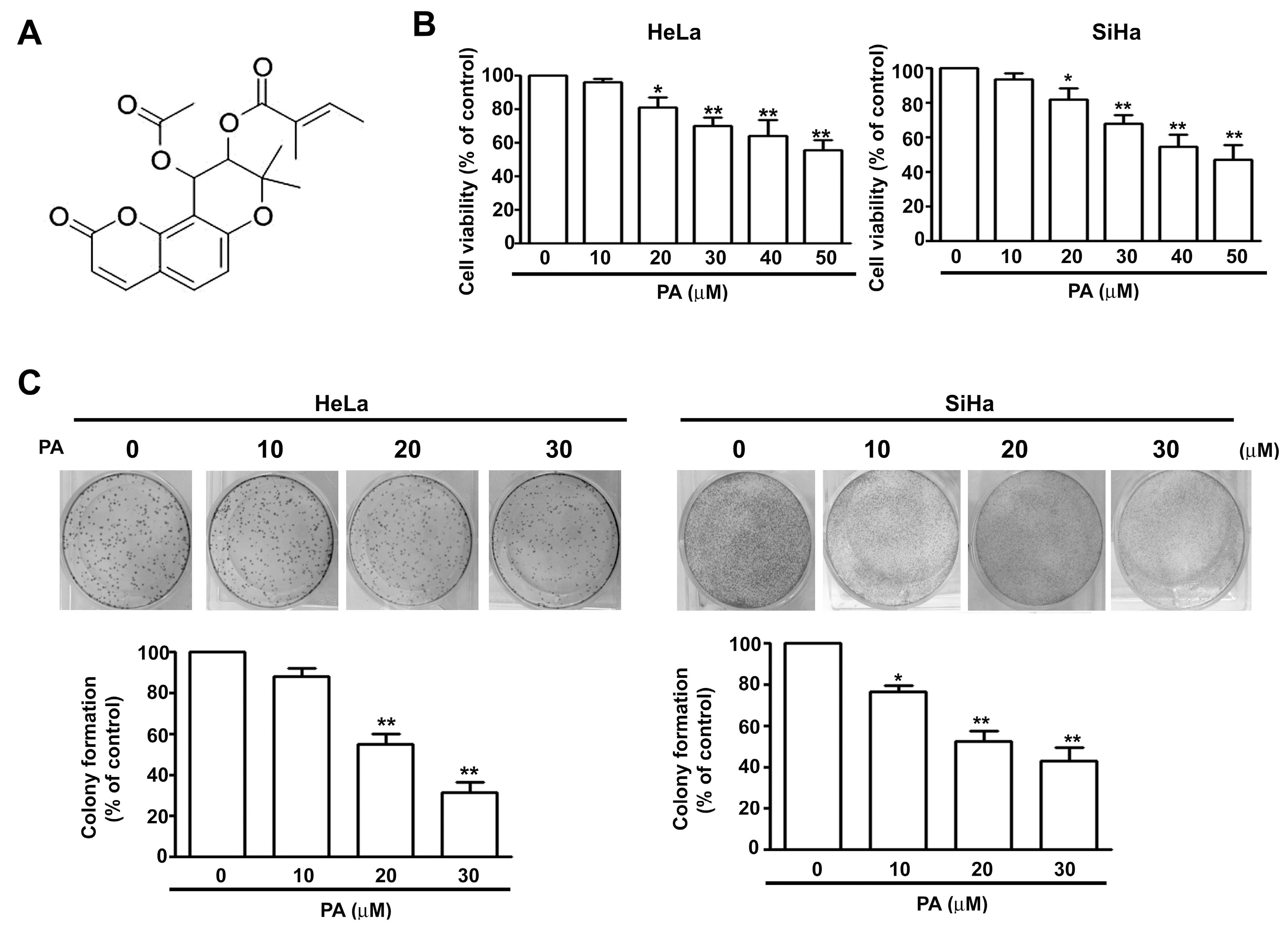
2.1. PA Reduces Viability and Colony Formation of HeLa and SiHa Cells
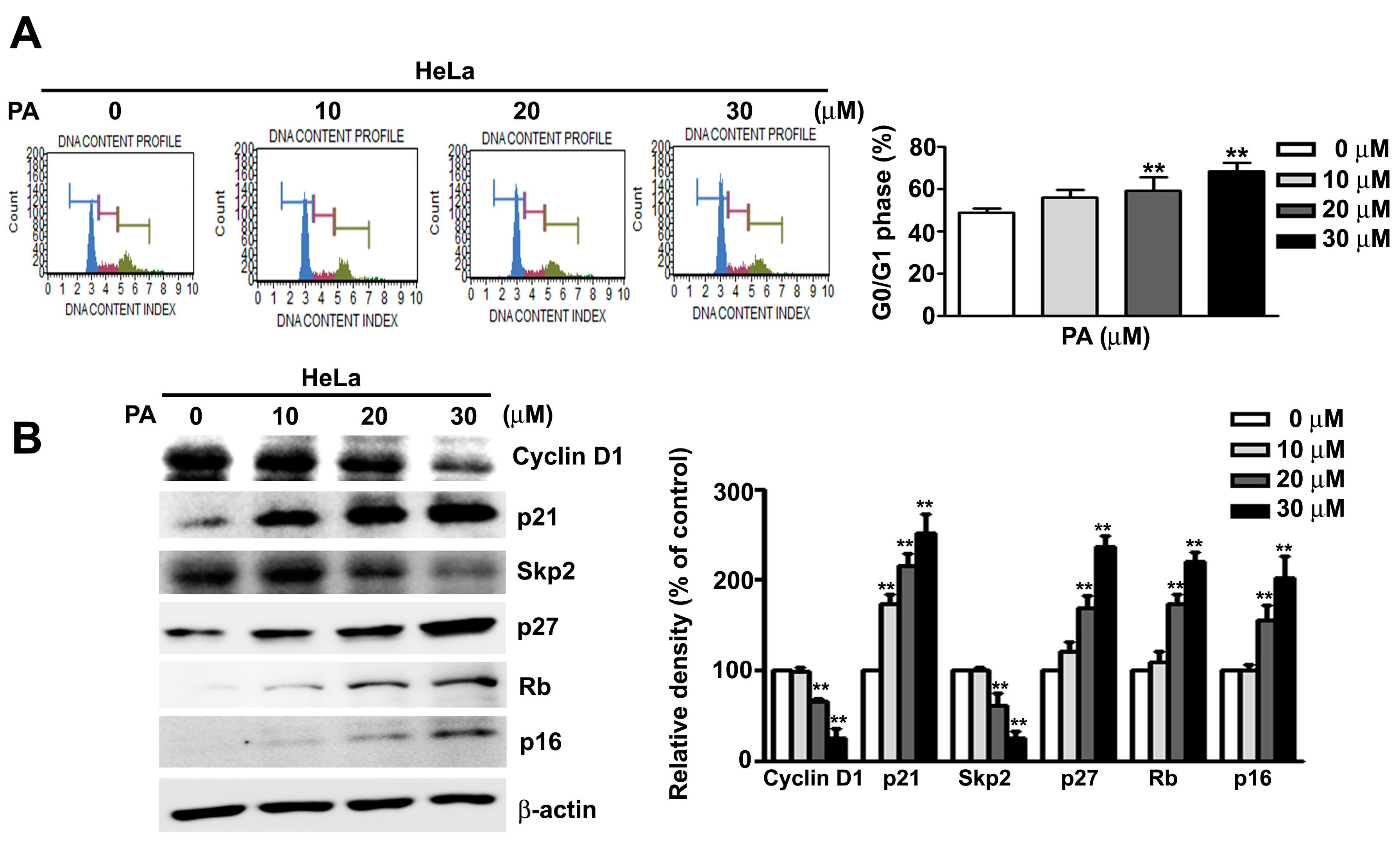
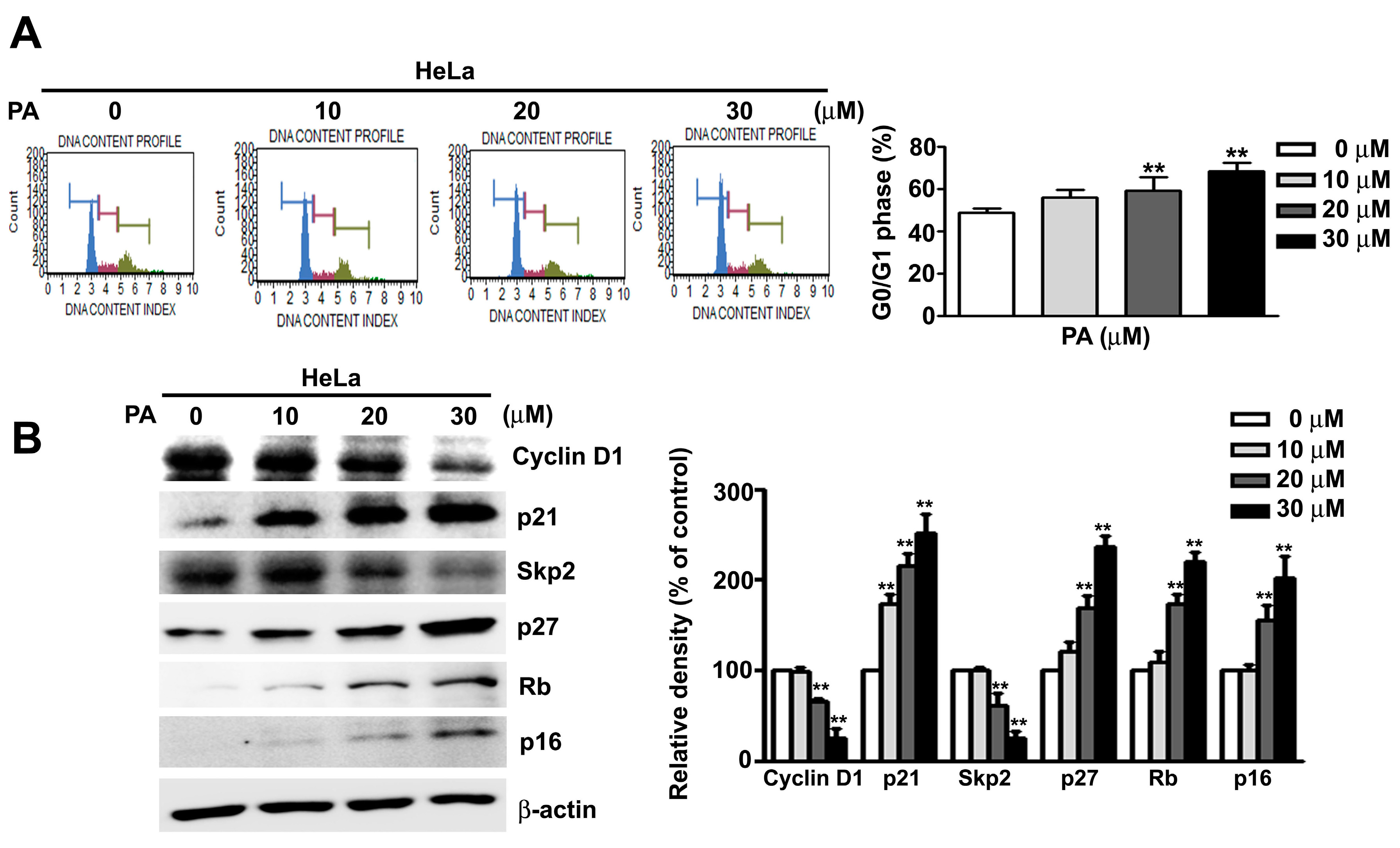
2.2. PA Induces Cell Cycle Arrest at G0/G1 Phase in HeLa Cells
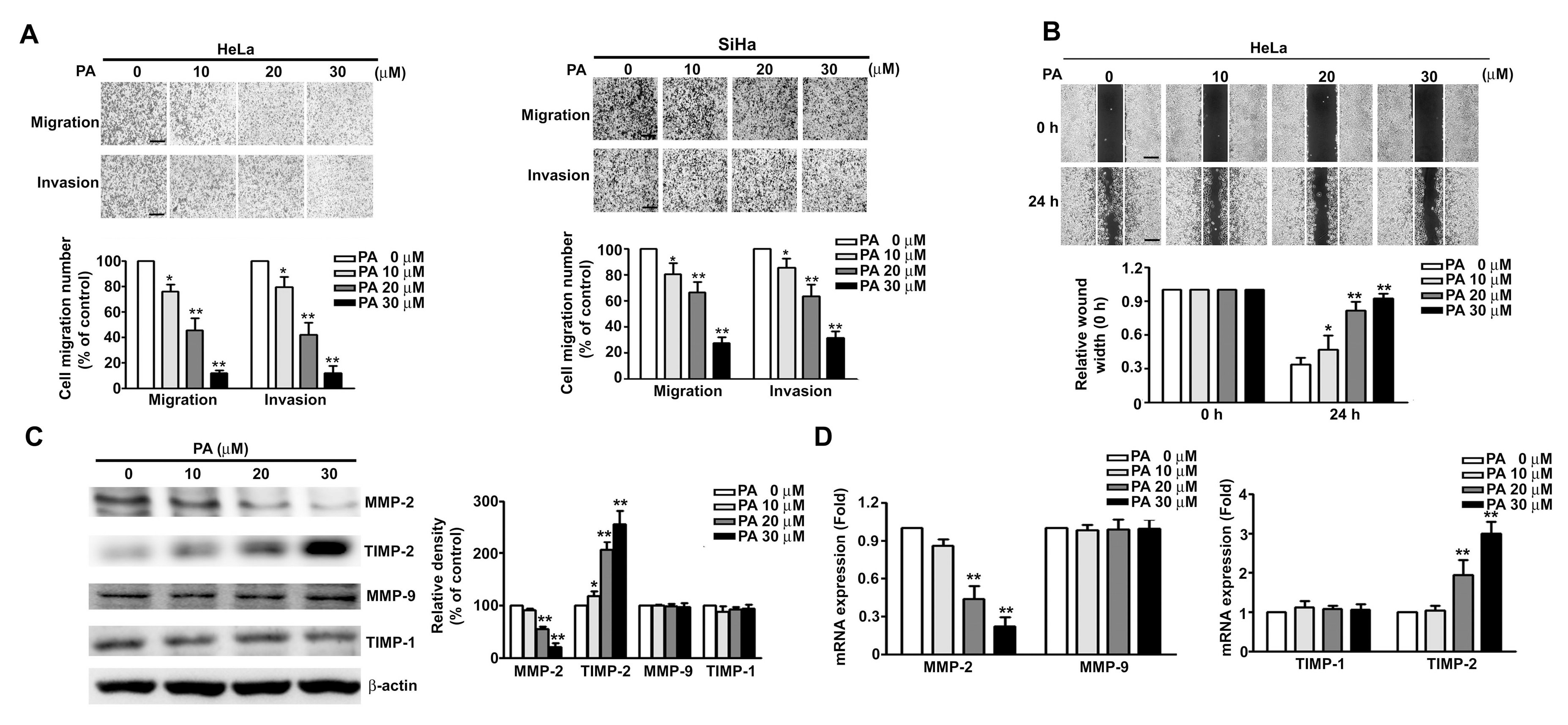
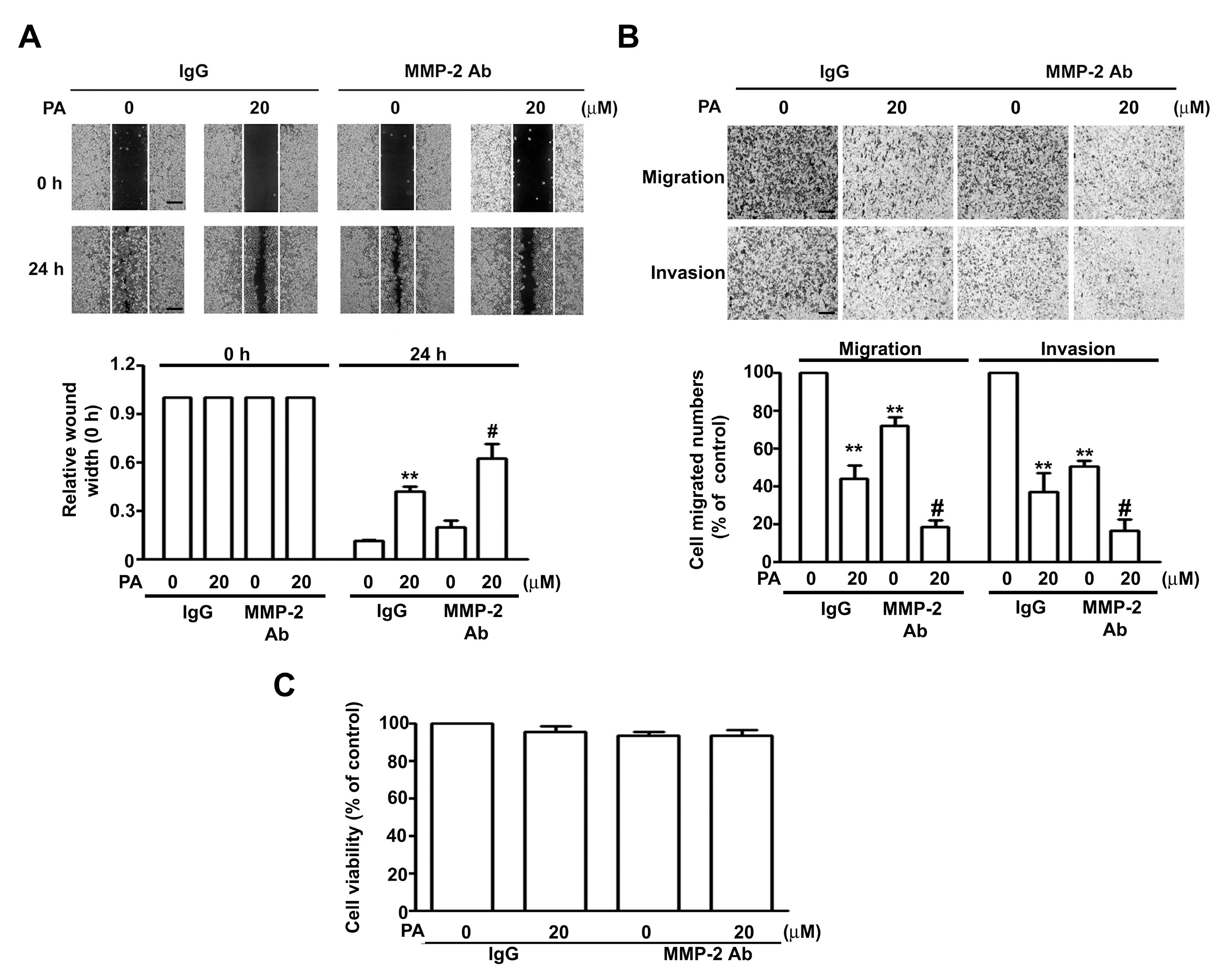
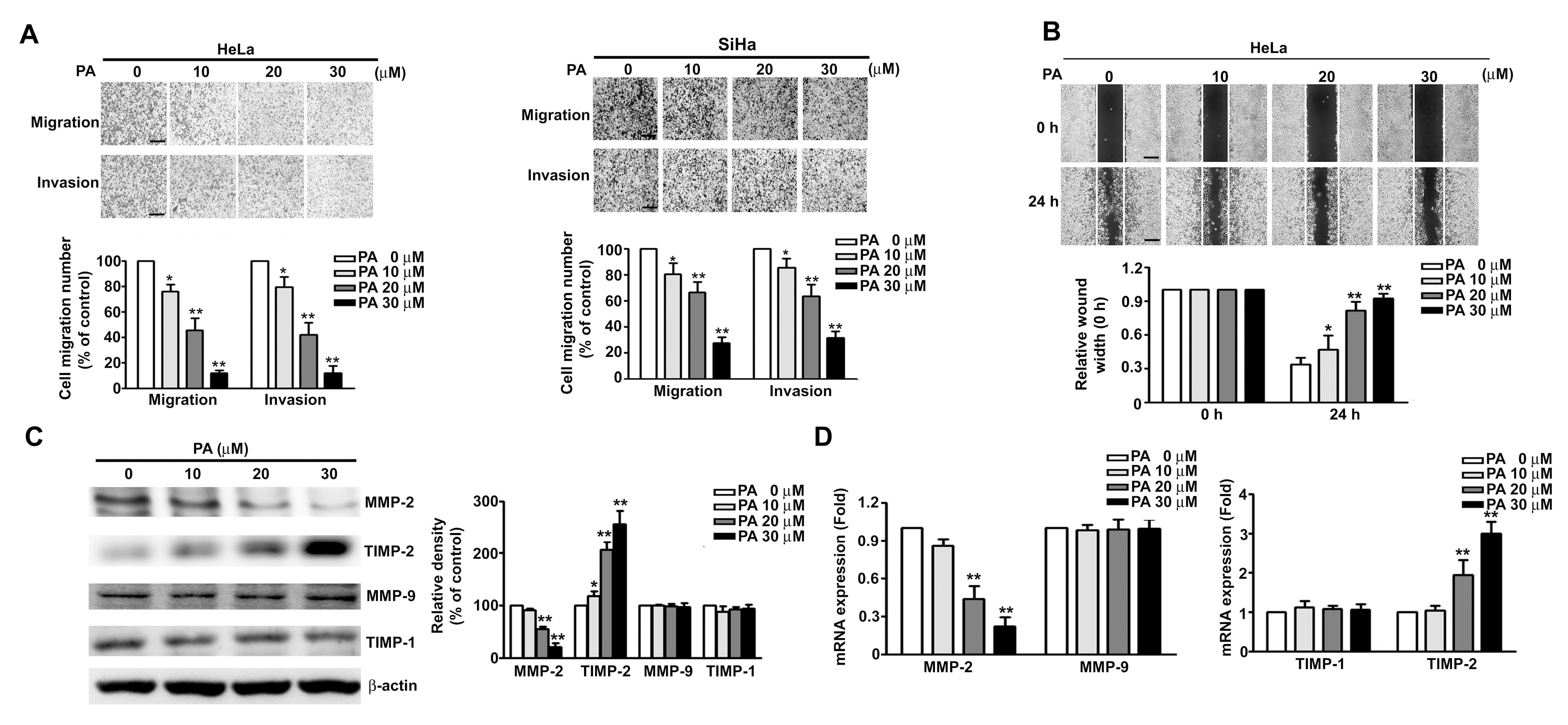
2.3. PA Inhibits Cell Migration/Invasion and MMP-2 Expression in HeLa and SiHa Cells
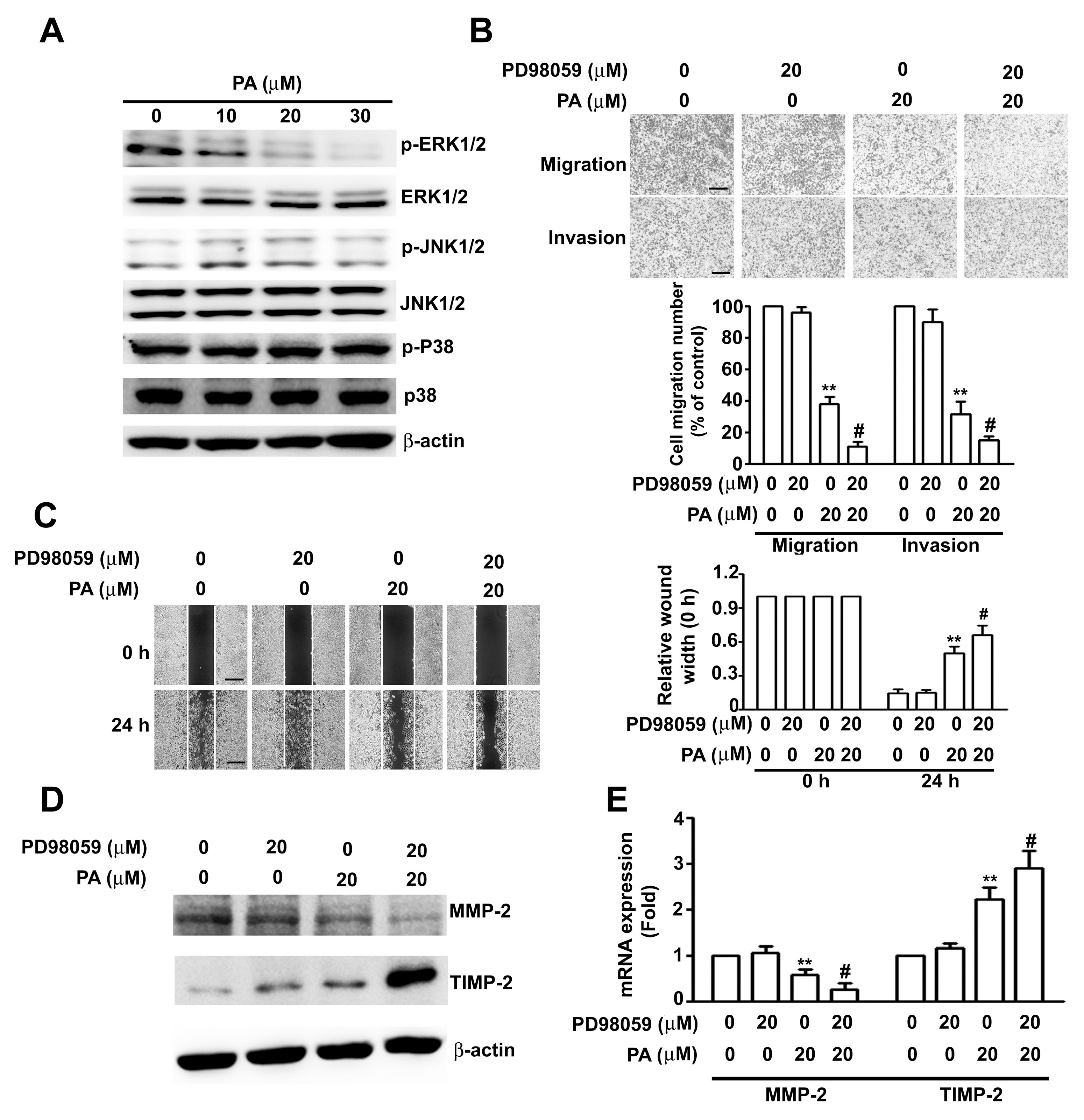
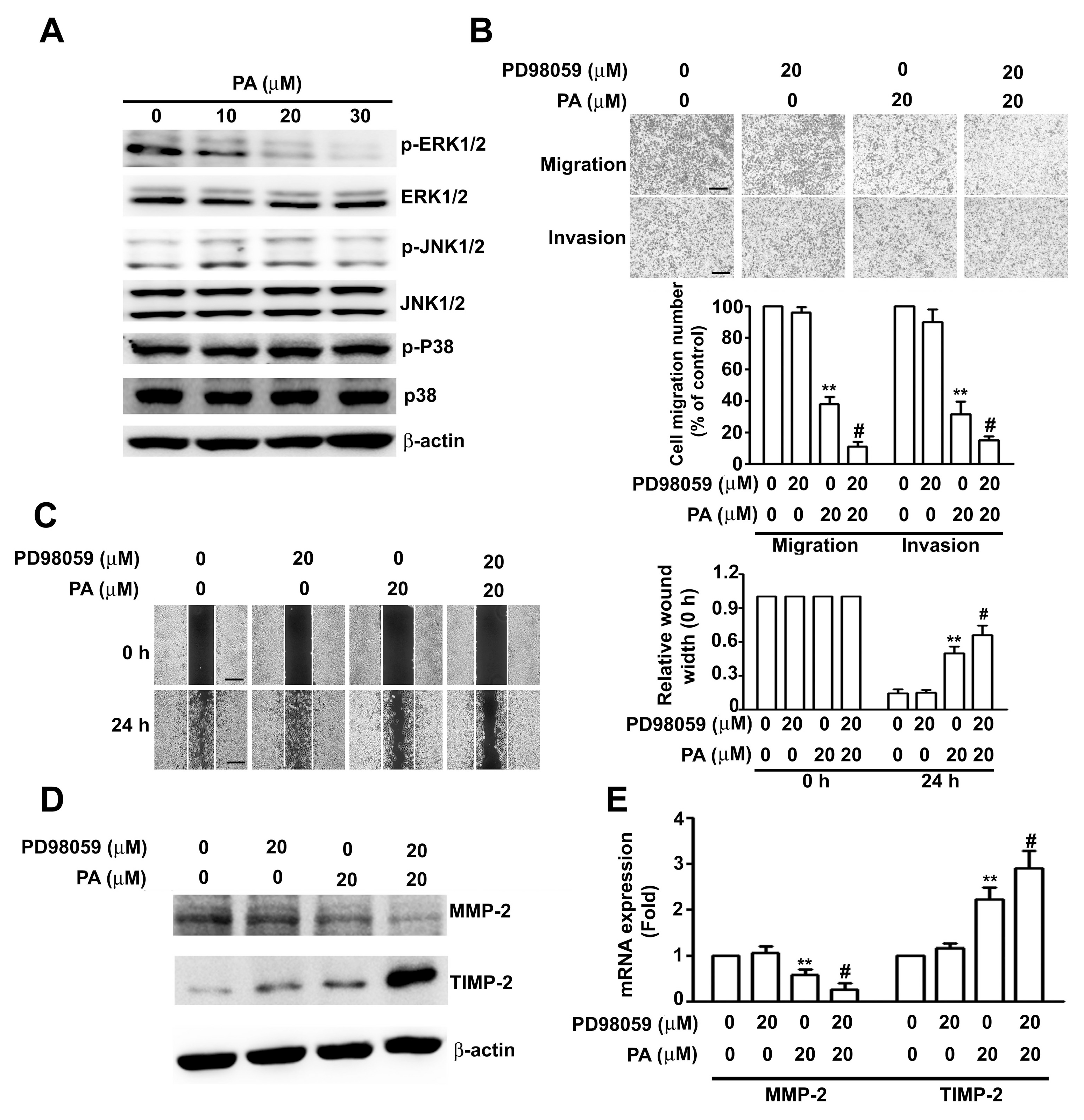
2.4. Role of ERK1/2 in PA-Modified MMP-2 and TIMP-2 Expression in HeLa Cells
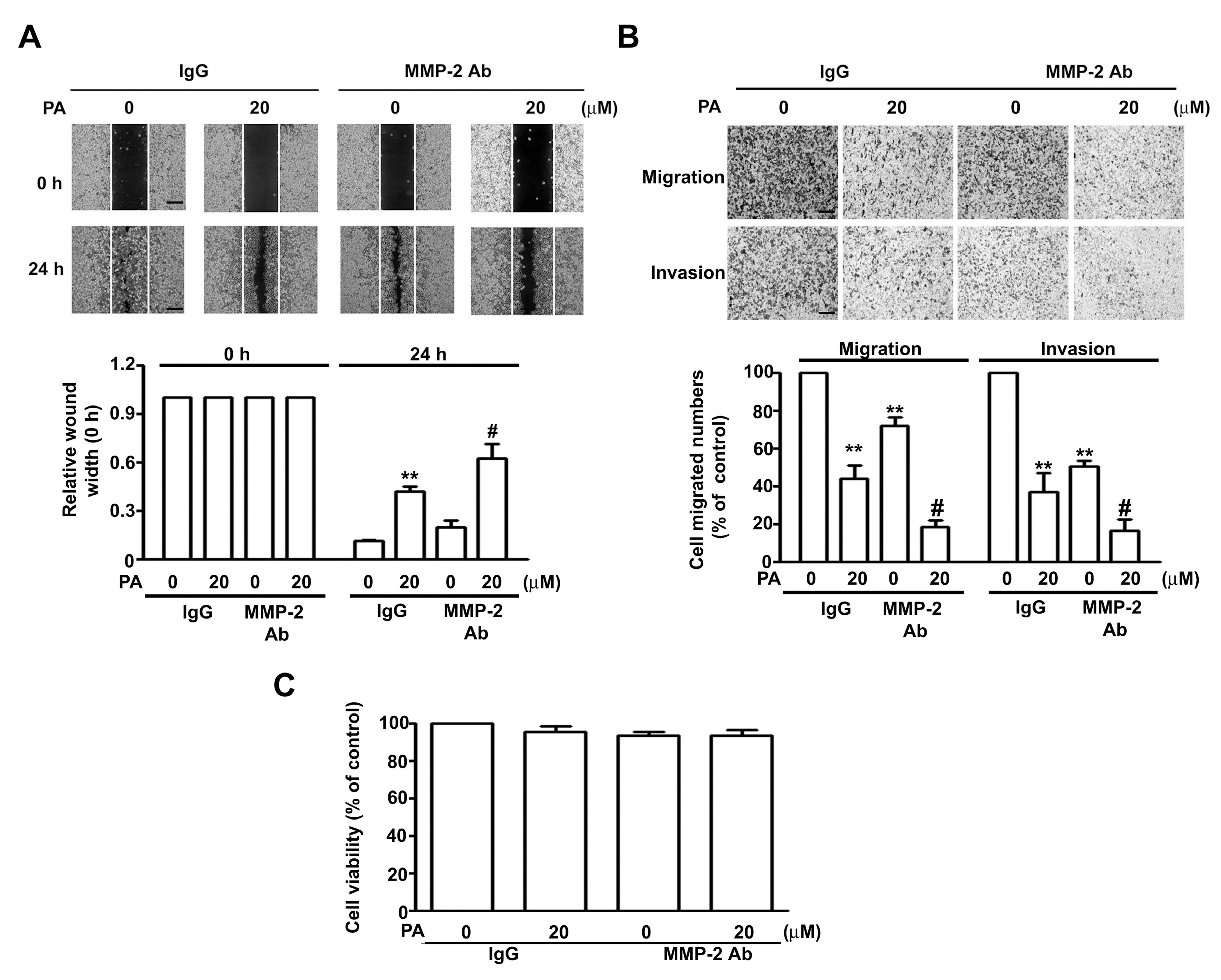
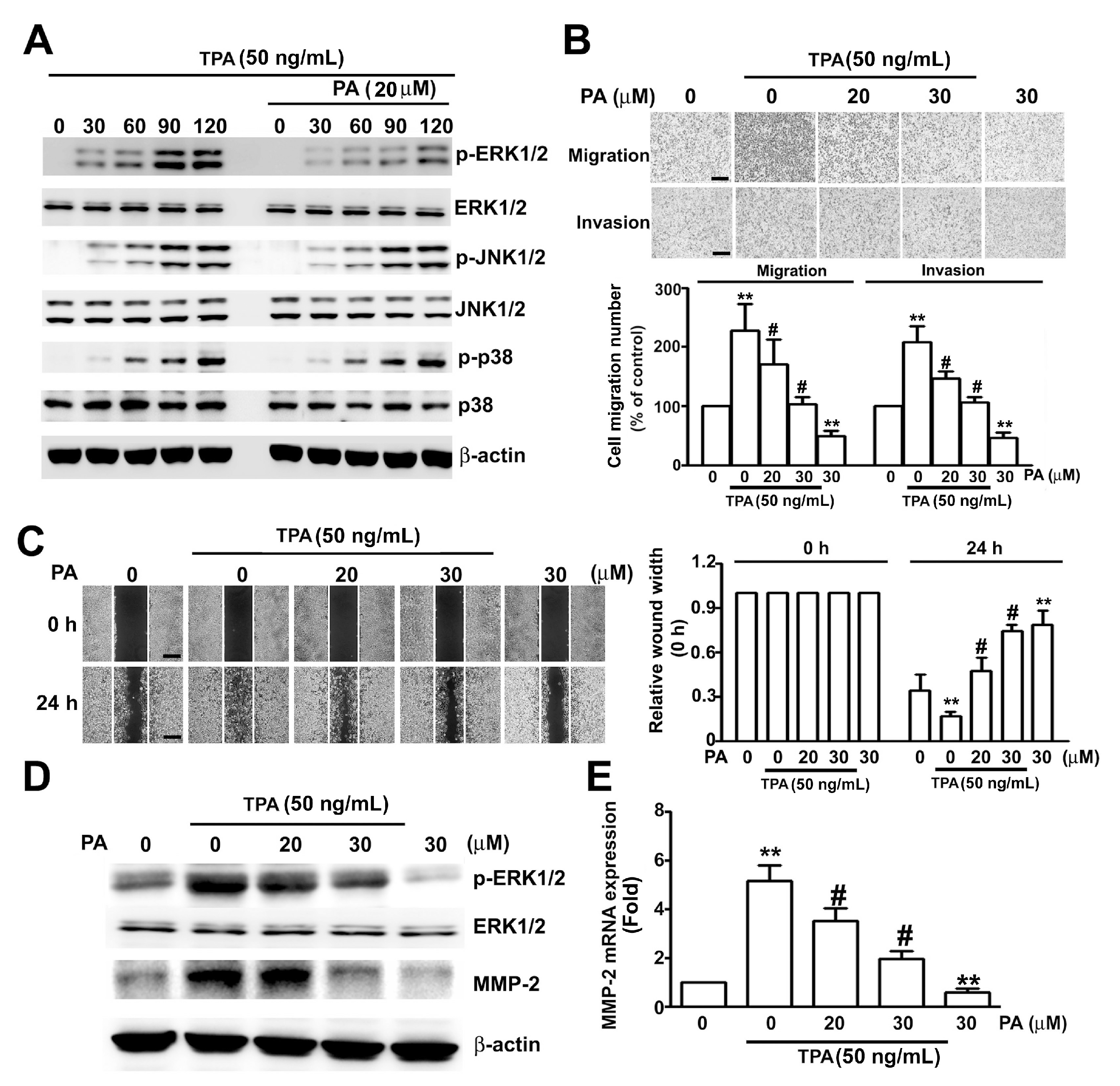
2.5. PA Inhibits TPA-Induced ERK1/2 Activation, MMP-2 Expression, and Migration/Invasion in HeLa Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Culture Condition
4.3. Immunoblotting
4.4. Cell Viability Assay
4.5. Colony Formation Assay
4.6. Flow Cytometric Analysis
4.7. Migration and Invasion Assay
4.8. Wound Healing Assay
4.9. Reverse Transcription and Real-Time PCR Assay
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eskander, R.N.; Tewari, K.S. Chemotherapy in the treatment of metastatic, persistent, and recurrent cervical cancer. Curr. Opin. Obstet. Gynecol. 2014, 26, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Hu, Y.; Jin, P.; Meng, X.; Chen, Y.; Zhang, H. Intensity-modulated radiotherapy combined with iodine-125 seed implantation in non-central recurrence of cervical cancer: A case report and literature review. Oncol. Lett. 2017, 14, 4085–4091. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Lim, M.C.; Lim, S.Y.; Bae, J.M.; Yoo, C.W.; Seo, S.S.; Kang, S.; Park, S.Y. Port-site and liver metastases after laparoscopic pelvic and para-aortic lymph node dissection for surgical staging of locally advanced cervical cancer. Int. J. Gynecol. Cancer 2008, 18, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Thanapprapasr, D.; Nartthanarung, A.; Likittanasombut, P.; Na Ayudhya, N.I.; Charakorn, C.; Udomsubpayakul, U.; Subhadarbandhu, T.; Wilailak, S. Bone metastasis in cervical cancer patients over a 10-year period. Int. J. Gynecol. Cancer 2010, 20, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Liotta, L.A.; Stetler-Stevenson, W.G. Tumor invasion and metastasis: An imbalance of positive and negative regulation. Cancer Res. 1991, 51 (Suppl. S18), 5054s–5059s. [Google Scholar] [PubMed]
- Yoon, W.H.; Jung, Y.J.; Kim, T.D.; Li, G.; Park, B.J.; Kim, J.Y.; Lee, Y.C.; Kim, J.M.; Park, J.I.; Park, H.D.; et al. Gabexatemesilate inhibits colon cancer growth, invasion, and metastasis by reducing matrix metalloproteinases and angiogenesis. Clin. Cancer Res. 2004, 10, 4517–4526. [Google Scholar] [CrossRef] [PubMed]
- Ellerbroek, S.M.; Stack, M.S. Membrane associated matrix metalloproteinases in metastasis. BioEssays 1999, 21, 940–949. [Google Scholar] [CrossRef]
- Zhu, D.; Ye, M.; Zhang, W. E6/E7 oncoproteins of high risk HPV-16 upregulate MT1-MMP, MMP-2 and MMP-9 and promote the migration of cervical cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 4981–4989. [Google Scholar] [PubMed]
- Meng, F.; Chen, X.; Song, H.; Lou, G. LAPTM4B down regulation inhibits the proliferation, invasion and angiogenesis of HeLa cells in vitro. Cell. Physiol. Biochem. 2015, 37, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.E.; Alonso, D.F.; Yoshiji, H.; Thorgeirsson, U.P. Tissue inhibitors of metalloproteinases: Structure, regulation and biological functions. Eur. J. Cell Biol. 1997, 74, 111–122. [Google Scholar] [PubMed]
- Kma, L. Roles of plant extracts and constituents in cervical cancer therapy. Asian Pac. J. Cancer Prev. 2013, 14, 3429–3436. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Nicoletti, M.; Battinelli, L.; Mazzanti, G. Isolation of praeruptorins A and B from Peucedanumpraeruptorum Dunn and their general pharmacological evaluation in comparison with extracts of the drug. Farmaco 2001, 56, 417–420. [Google Scholar] [CrossRef]
- Yu, P.J.; Ci, W.; Wang, G.F.; Zhang, J.Y.; Wu, S.Y.; Xu, W.; Jin, H.; Zhu, Z.G.; Zhang, J.J.; Pang, J.X.; et al. Praeruptorin A inhibits lipopolysaccharide-induced inflammatory response in murine macrophages through inhibition of NF-kappaB pathway activation. Phytother. Res. 2011, 25, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, J.; Yang, L.; Zhou, S.M.; Guan, S.Y.; Yang, L.K.; Shi, Q.X.; Zhao, M.G.; Yang, Q. Effect of Praeruptorin C on 3-nitropropionic acid induced Huntington’s disease-like symptoms in mice. Biomed. Pharmacother. 2017, 86, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Yue, W.; Li, Q. Chemopreventive effects of Peucedanumpraeruptorum Dunn and its major constituents on SGC7901 gastric cancer cells. Molecules 2010, 15, 8060–8071. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, Y.J.; Kim, J.; Bang, O.S. Pyranocoumarins from Root Extracts of Peucedanumpraeruptorum Dunn with Multidrug Resistance Reversal and Anti-Inflammatory Activities. Molecules 2015, 20, 20967–20978. [Google Scholar] [CrossRef] [PubMed]
- Overall, C.M.; Dean, R.A. Degradomics: Systems biology of the protease web. Pleiotropic roles of MMPs in cancer. Cancer Metastasis Rev. 2006, 25, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Jiang, X.J.; Yang, K.; Wang, L.X.; Wen, S.Z.; Wang, F. Prenylated Coumarins from Heracleumstenopterum, Peucedanumpraeruptorum, Clausenalansium, and Murrayapaniculata. Nat. Prod. Bioprospect. 2016, 6, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Ma, L.; Shen, Y.; Zhai, W.; Zhou, Y. Effect of angular pyranocoumarin isolated from Peucedanumpraeruptorum on the proliferation and apoptosis of U266 cells. Zhonghua Xue Ye Xue Za Zhi 2015, 36, 937–941. [Google Scholar]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, N.; Bager, C.L.; Kehlet, S.N.; Dragsbaek, K.; Neergaard, J.S.; Hansen, H.B.; Bay-Jensen, A.C.; Leeming, D.J.; Lipton, A.; Christiansen, C.; et al. Excessive matrix metalloprotease-mediated degradation of interstitial tissue (type I collagen) independently predicts short-term survival in an observational study of postmenopausal women diagnosed with cancer. Oncotarget 2017, 8, 52501–52510. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Wu, D.; Zou, J.; Chen, J.; Chen, L.; Chen, Y.; Ni, C.; Yuan, H. Prognostic impact of serum and tissue MMP-9 in non-small cell lung cancer: A systematic review and meta-analysis. Oncotarget 2016, 7, 18458–18468. [Google Scholar] [CrossRef] [PubMed]
- Tokito, A.; Jougasaki, M. Matrix Metalloproteinases in Non-Neoplastic Disorders. Int. J. Mol. Sci. 2016, 17, 1178. [Google Scholar] [CrossRef] [PubMed]
- Solovyeva, N.I.; Timoshenko, O.S.; Gureeva, T.A.; Kugaevskaya, E.V. Matrix metalloproteinases and their endogenous regulators in squamous cervical carcinoma (review of the own data). Biomed. Khim. 2015, 61, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.S.; Prasad, C.B.; Prasad, S.B.; Pandey, L.K.; Singh, S.; Pradhan, S.; Narayan, G. Anti-tumor activity of staurosporine in the tumor microenvironment of cervical cancer: An in vitro study. Life Sci. 2015, 133, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kaewprag, J.; Umnajvijit, W.; Ngamkham, J.; Ponglikitmongkol, M. HPV16 oncoproteins promote cervical cancer invasiveness by upregulating specific matrix metalloproteinases. PLoS ONE 2013, 8, e71611. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Baksi, R.; Agarwal, M. Repositioning of anti-viral drugs as therapy for cervical cancer. Pharmacol. Rep. 2016, 68, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Moirangthem, A.; Dalui, R.; Ghosh, T.; Bandyopadhyay, A.; Dasgupta, A.; Banerjee, U.; Jana, N.; Basu, A. Expression of matrix metalloproteinase-2 and 9 in cervical intraepithelial neoplasia and cervical carcinoma among different age groups of premenopausal and postmenopausal women. J. Cancer Res. Clin. Oncol. 2014, 140, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.J.; Cho, H.J.; Chung, F.L.; Wang, X.; Hoe, H.S.; Park, K.K.; Kim, C.H.; Chang, H.W.; Lee, S.R.; Chang, Y.C. Isothiocyanates suppress the invasion and metastasis of tumors by targeting FAK/MMP-9 activity. Oncotarget 2017, 8, 63949–63962. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.S.; Chu, S.C.; Yang, S.F.; Chen, P.N.; Liu, Y.C.; Lu, K.H. Silibinin suppresses human osteosarcoma MG-63 cell invasion by inhibiting the ERK-dependent c-Jun/AP-1 induction of MMP-2. Carcinogenesis 2007, 28, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.C.; Shih, Y.W.; Chao, C.H.; Lee, X.Y.; Chiang, T.A. Involvement of the ERK signaling pathway in fisetin reduces invasion and migration in the human lung cancer cell line A549. J. Agric. Food Chem. 2009, 57, 8933–8941. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.; Shen, K.H.; Chiang, T.A.; Shih, Y.W. Acacetin inhibits TPA-induced MMP-2 and u-PA expressions of human lung cancer cells through inactivating JNK signaling pathway and reducing binding activities of NF-kappaB and AP-1. J. Food Sci. 2010, 75, H30–H38. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.R.; Liu, C.J.; Yeh, C.C. Naringenin suppresses TPA-induced tumor invasion by suppressing multiple signal transduction pathways in human hepatocellular carcinoma cells. Chem. Biol. Interact. 2015, 235, 1–9. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.-H.; Lin, C.-L.; Chiou, H.-L.; Yang, S.-F.; Lin, C.-Y.; Liu, C.-J.; Hsieh, Y.-H. Praeruptorin A Inhibits Human Cervical Cancer Cell Growth and Invasion by Suppressing MMP-2 Expression and ERK1/2 Signaling. Int. J. Mol. Sci. 2018, 19, 10. https://doi.org/10.3390/ijms19010010
Wu M-H, Lin C-L, Chiou H-L, Yang S-F, Lin C-Y, Liu C-J, Hsieh Y-H. Praeruptorin A Inhibits Human Cervical Cancer Cell Growth and Invasion by Suppressing MMP-2 Expression and ERK1/2 Signaling. International Journal of Molecular Sciences. 2018; 19(1):10. https://doi.org/10.3390/ijms19010010
Chicago/Turabian StyleWu, Min-Hua, Chia-Liang Lin, Hui-Ling Chiou, Shun-Fa Yang, Ching-Yi Lin, Chung-Jung Liu, and Yi-Hsien Hsieh. 2018. "Praeruptorin A Inhibits Human Cervical Cancer Cell Growth and Invasion by Suppressing MMP-2 Expression and ERK1/2 Signaling" International Journal of Molecular Sciences 19, no. 1: 10. https://doi.org/10.3390/ijms19010010
APA StyleWu, M.-H., Lin, C.-L., Chiou, H.-L., Yang, S.-F., Lin, C.-Y., Liu, C.-J., & Hsieh, Y.-H. (2018). Praeruptorin A Inhibits Human Cervical Cancer Cell Growth and Invasion by Suppressing MMP-2 Expression and ERK1/2 Signaling. International Journal of Molecular Sciences, 19(1), 10. https://doi.org/10.3390/ijms19010010