The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan

 and
and
Abstract
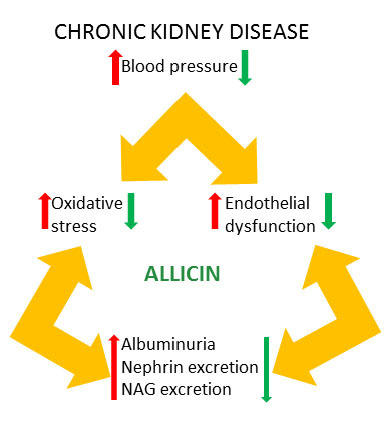
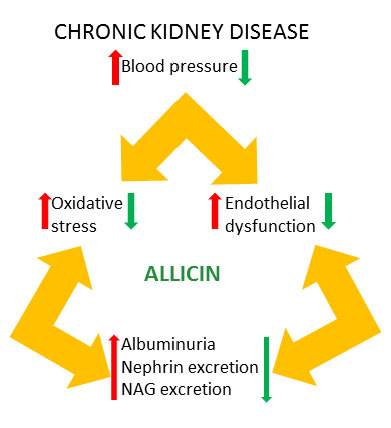
1. Introduction
2. Results
2.1. Physiological Data
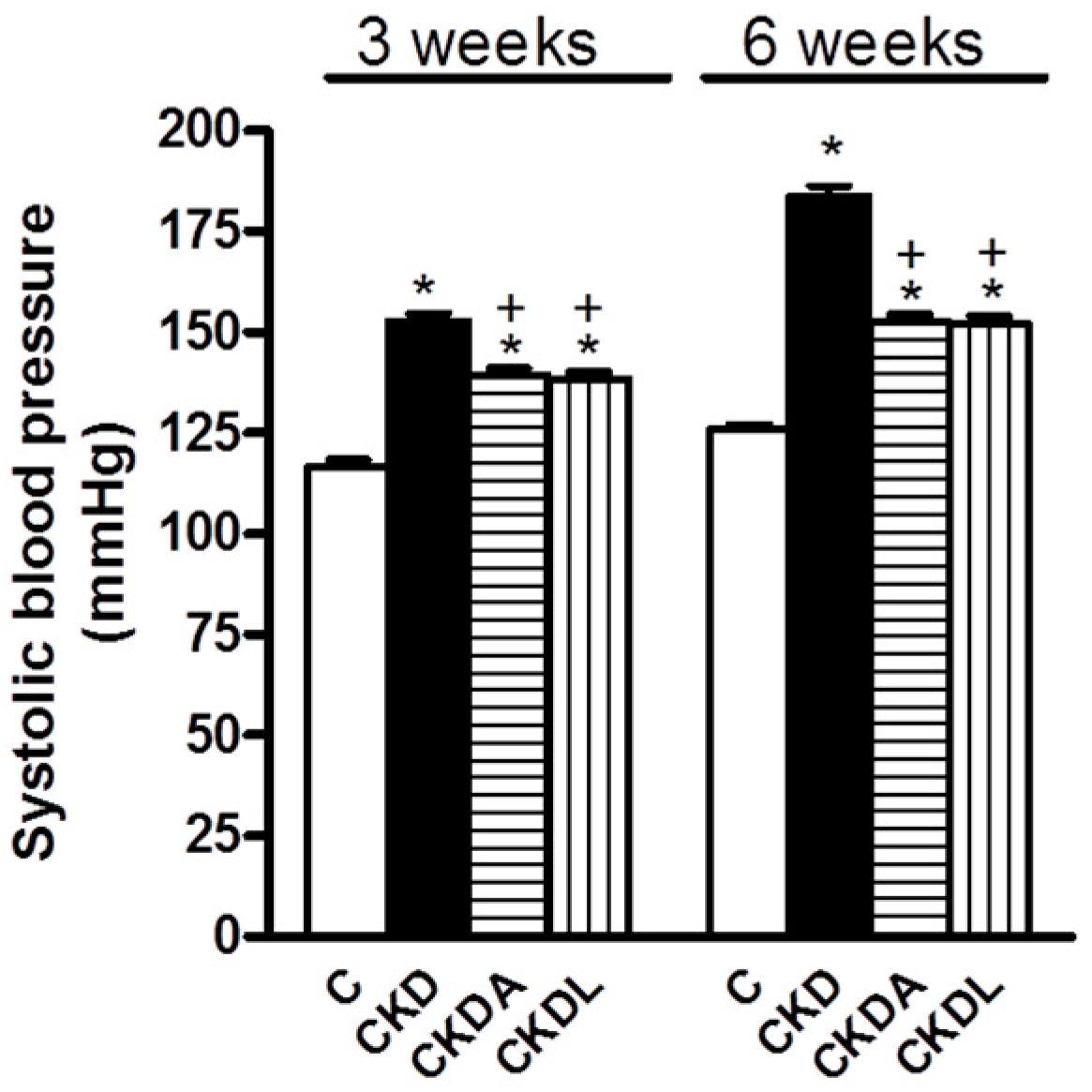
2.2. Systolic Blood Pressure (SBP)
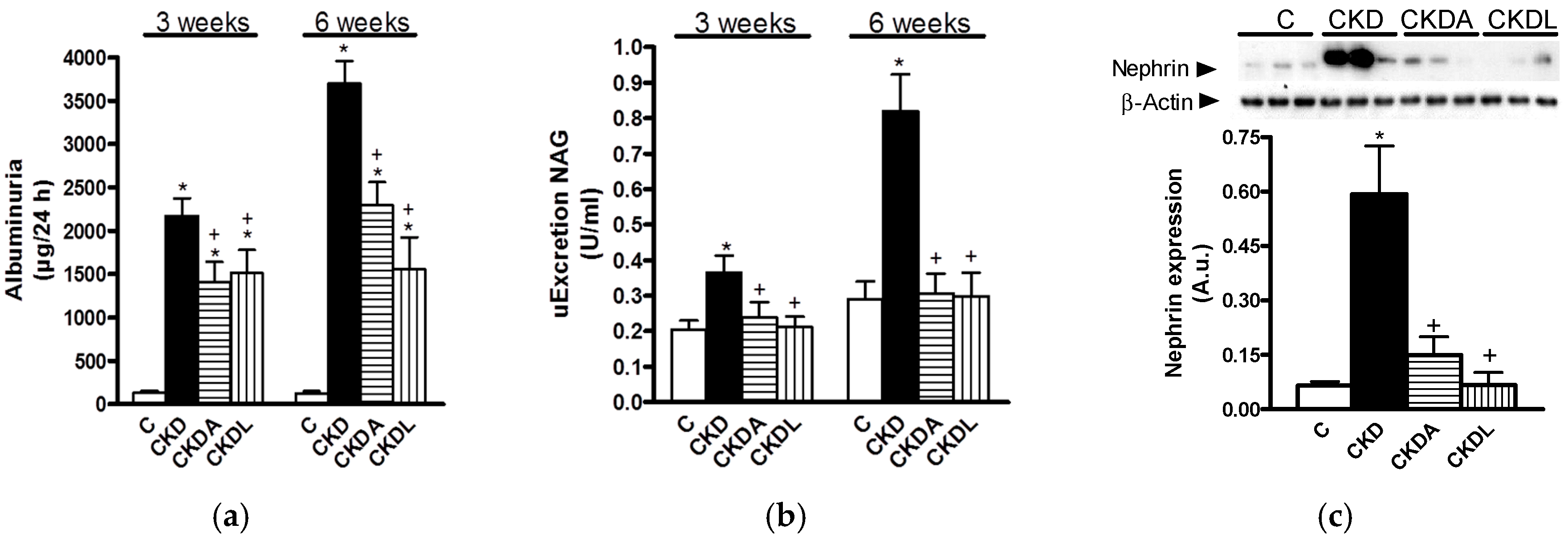
2.3. Markers of CKD Progression
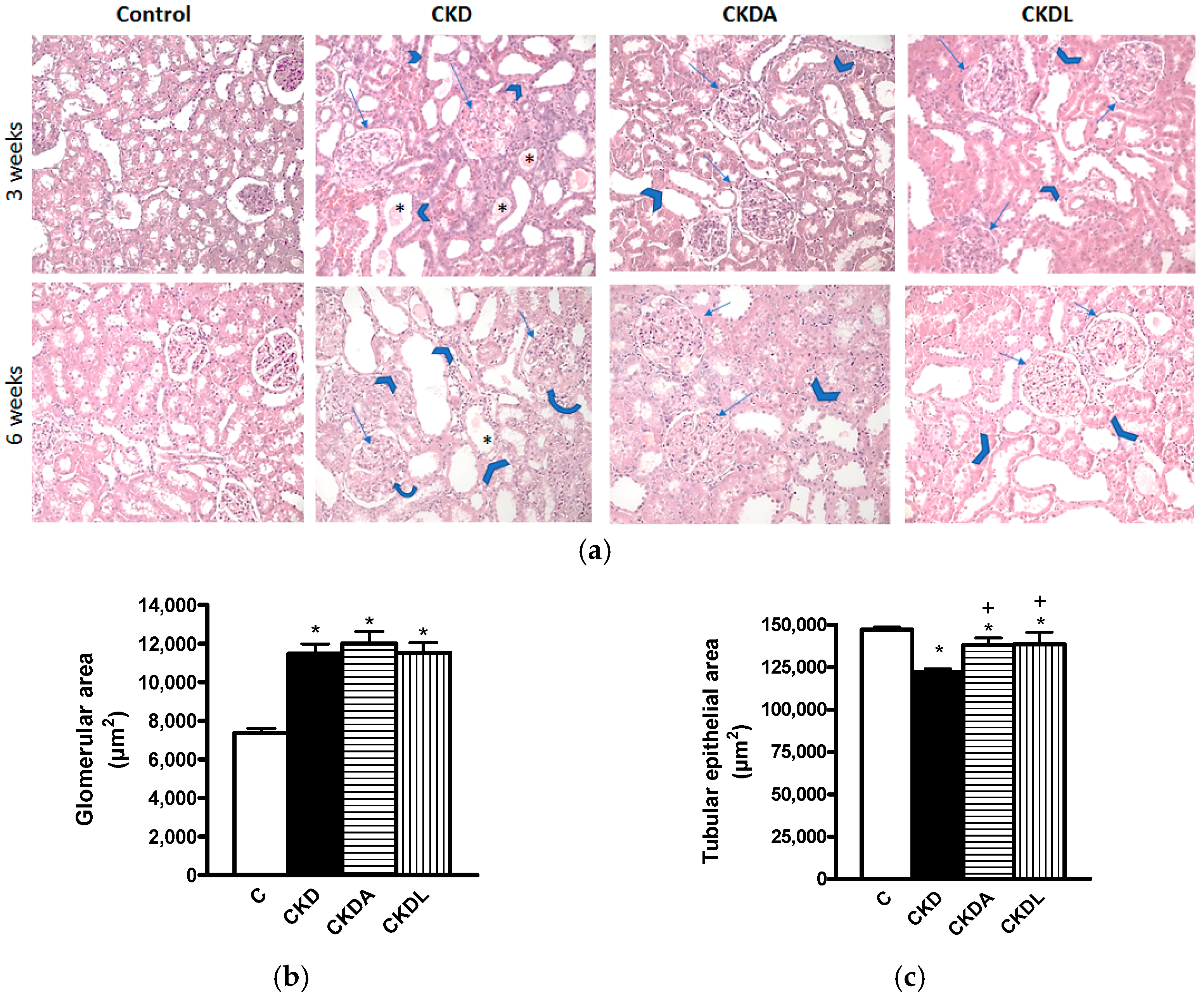
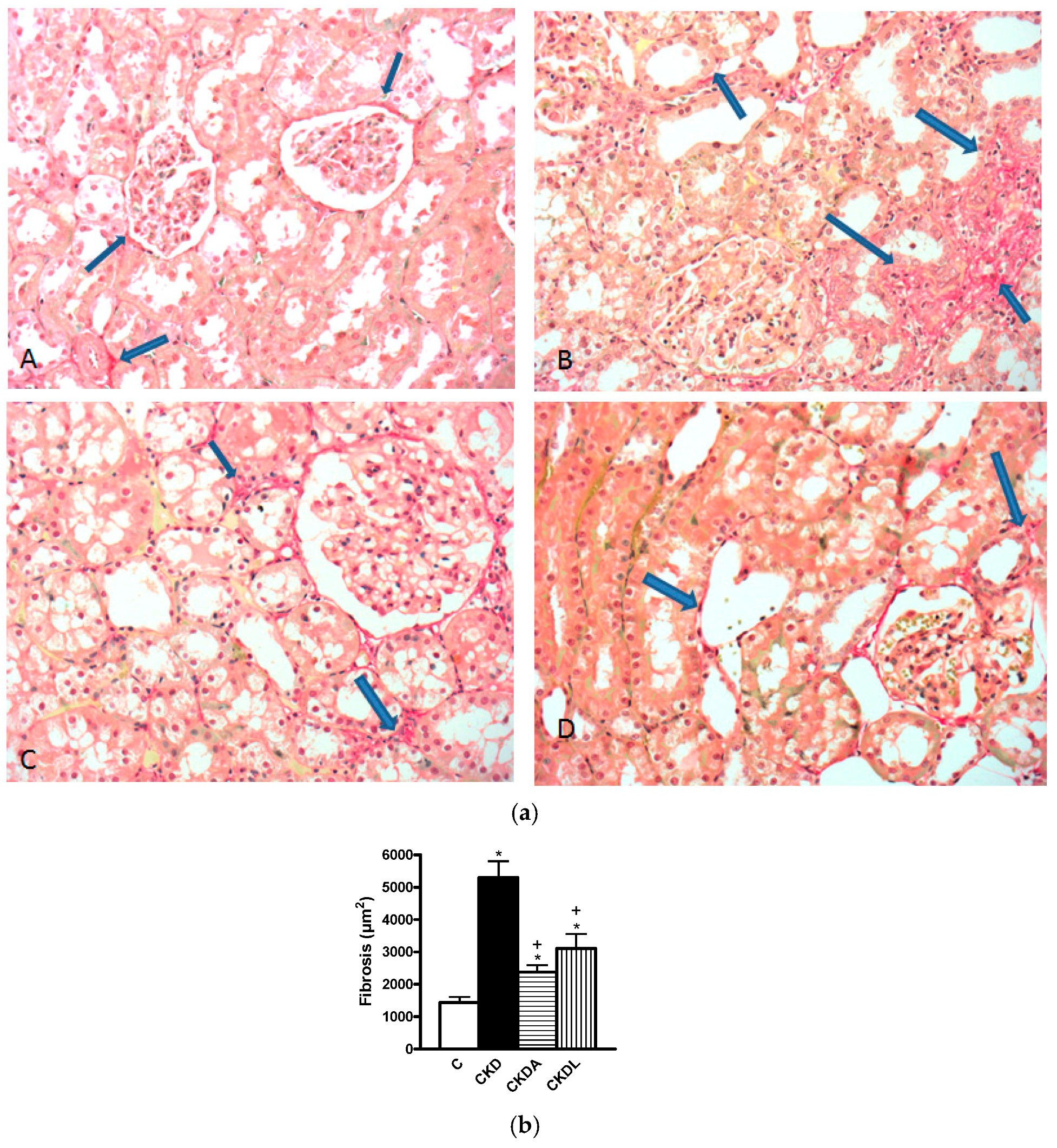
2.4. Histopathological Study
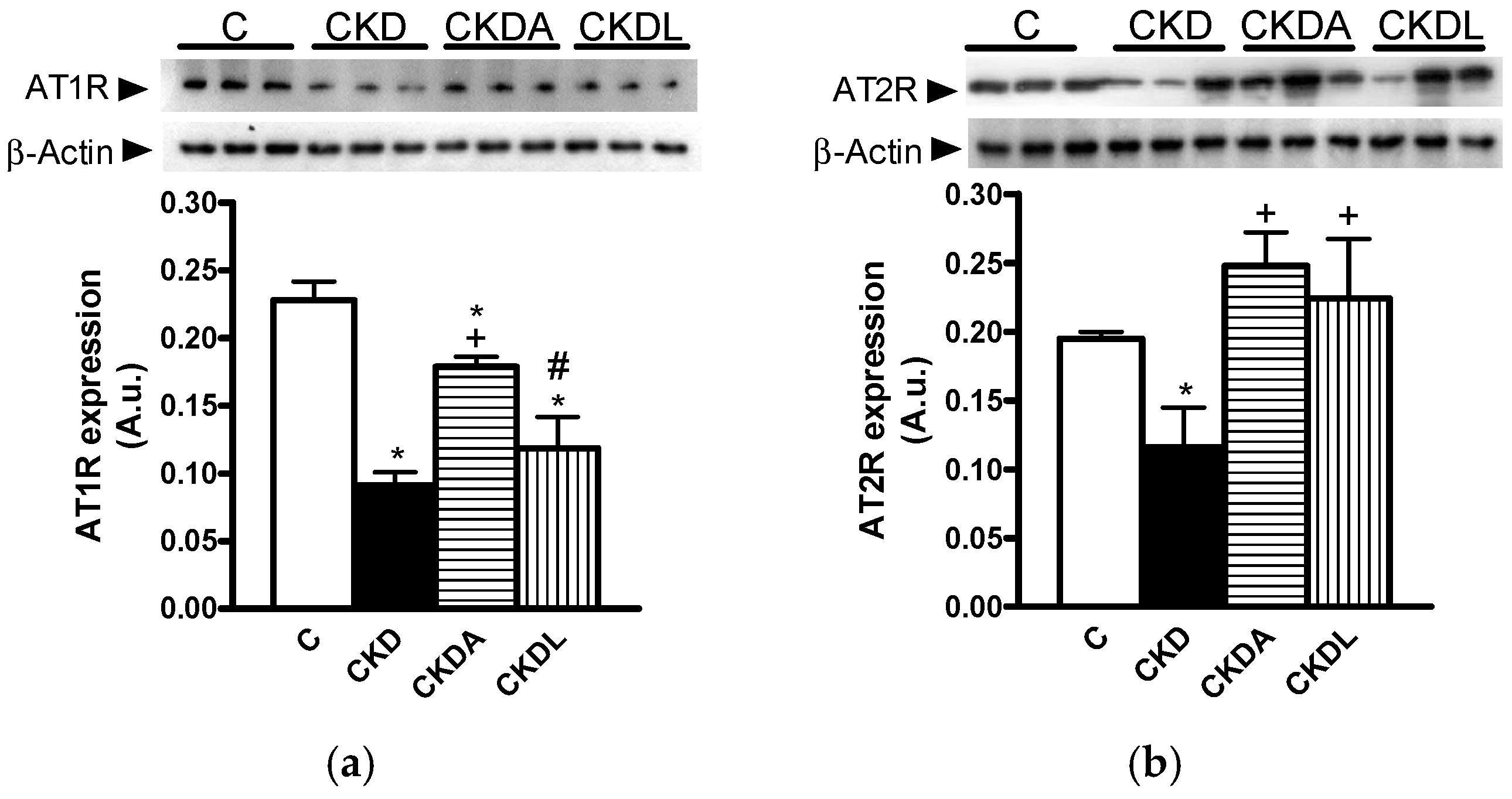
2.5. Expression of Angiotensin II Receptors in Kidney Cortex
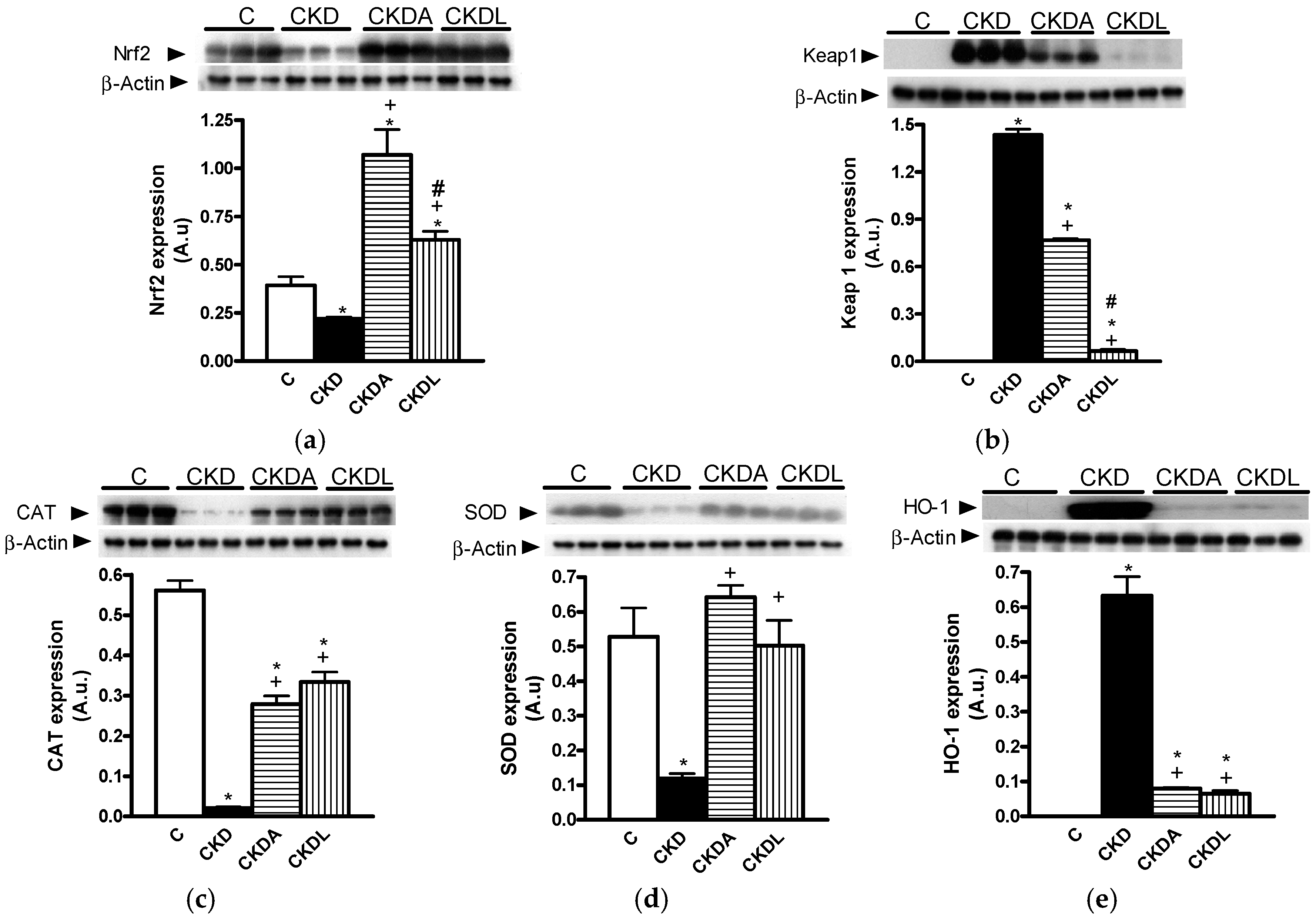
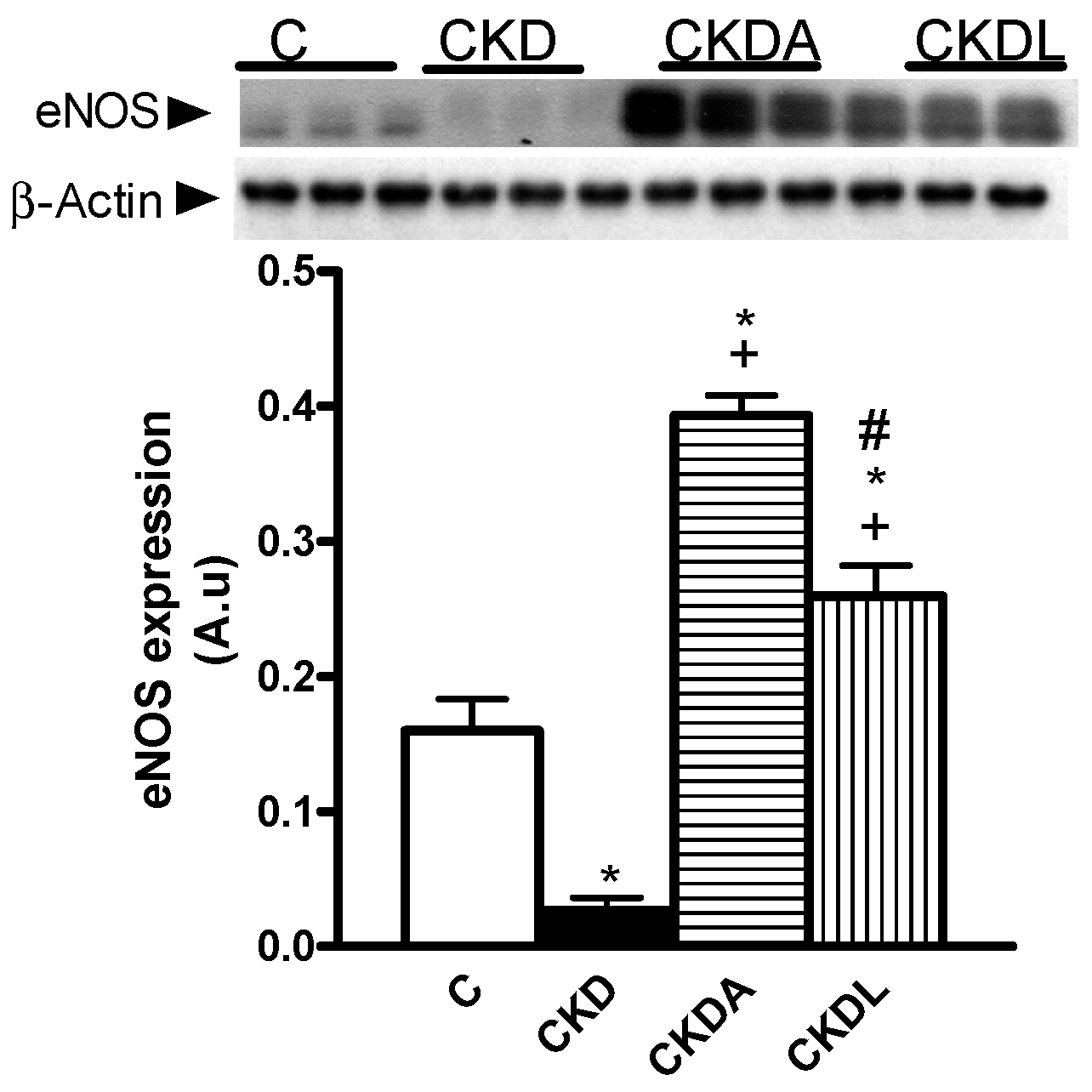
2.6. Evaluation of Renal Oxidative Stress
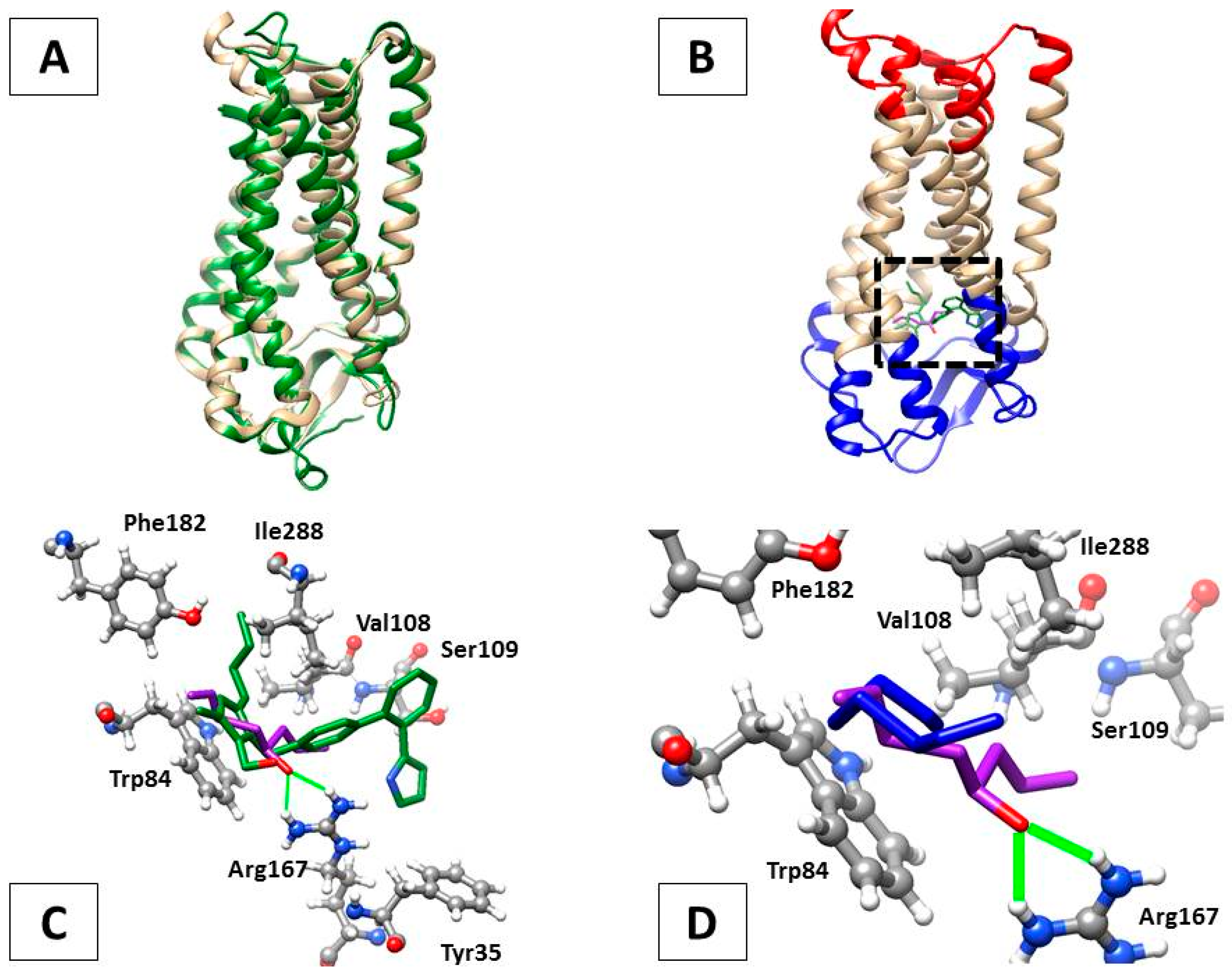
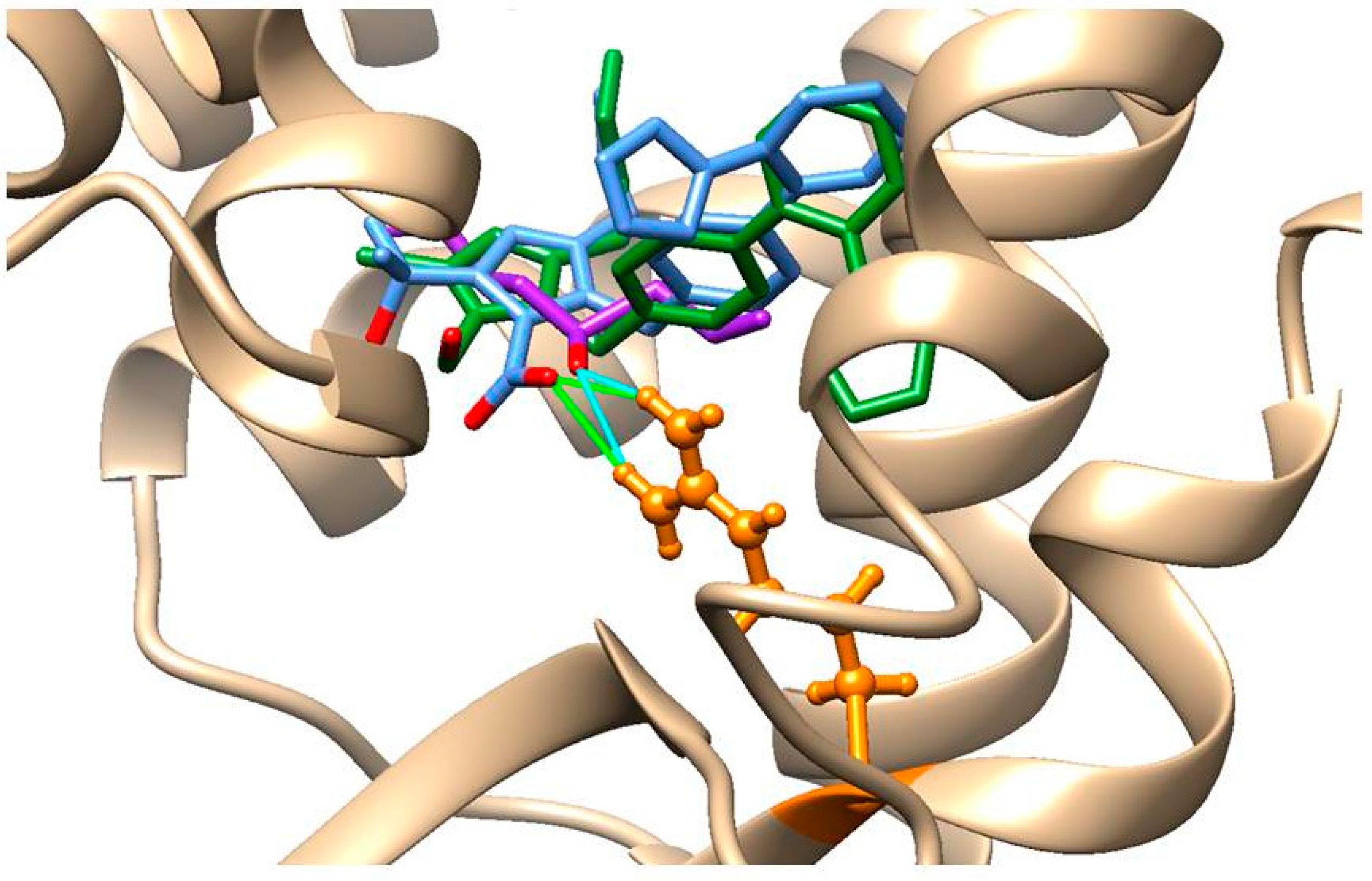
2.7. In Silico Analysis of AT1-Losartan and -Allicin Interactions
3. Discussion
4. Methods
4.1. Reagents
4.2. Experimental Design
4.3. Ethics Statement
4.4. Allicin
4.5. Induction of Experimental Model of Chronic Kidney Disease
4.6. Systolic Blood Pressure (SBP)
4.7. Renal Function
4.8. Measurement of NAG Activity
4.9. Histopathological Study
4.10. Evaluation of Renal Oxidative Stress
4.10.1. Determination of Lipid Peroxidation
4.10.2. Measurement of Oxidized Proteins
4.10.3. Expression of Nephrin, Nrf2/Keap1 Pathway, Endothelial Nitric Oxide Synthase (eNOS), and Angiotensin II Receptors in Renal Cortex
4.11. Bioinformatics Analyses
4.12. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sarafidis, P.A.; Bakris, G.L. Kidney disease and hypertension. In Comprehensive Hypertension; Lip, G.Y., Hall, J.E., Eds.; Elsevier Inc.: New York, NY, USA, 2007; pp. 607–619. [Google Scholar]
- The United States Renal Data System (USRDS). Atlas of Chronic Kidney Disease and End-Stage Renal Disease in the United States; National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2008.
- Sinclair, A.M.; Isles, C.G.; Brown, I.; Cameron, H.; Murray, G.D.; Robertson, J.W. Secondary hypertension in a blood pressure clinic. Arch. Int. Med. 1987, 147, 1289–1293. [Google Scholar] [CrossRef]
- Go, A.S.; Chertow, G.M.; Fan, D.; McCulloch, C.E.; Hsu, C.Y. Chronic kidney disease and the risks of death, cardiovascular events, and hospitalization. N. Engl. J. Med. 2004, 351, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Anavekar, N.S.; McMurray, J.J.; Velazquez, E.J.; Solomon, S.D.; Kober, L.; Rouleau, J.L.; White, H.D.; Nordlander, R.; Maggioni, A.; Dickstein, K.; et al. Relation between renal dysfunction and cardiovascular outcomes after myocardial infarction. N. Engl. J. Med. 2004, 351, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Sarafidis, P.A.; Bakris, G.L. Renin-angiotensin blockade and kidney disease. Lancet 2008, 372, 511–512. [Google Scholar] [CrossRef]
- Ravera, M.; Re, M.; Deferrari, L.; Vettoretti, S.; Deferrari, G. Importance of blood pressure control in chronic kidney disease. J. Am. Soc. Nephrol. 2006, 4, S98–S103. [Google Scholar] [CrossRef] [PubMed]
- Kidney Disease Outcomes Quality Initiative (K/DOQI). K/DOQI clinical practice guidelines on hypertension and antihypertensive agents in chronic kidney disease. Am. J. Kidney Dis. 2004, 43, 11–13. [Google Scholar]
- Nwokocha, C.R.; Owu, D.U.; Ufearo, C.; Iwuala, M.O. Comparative study on the efficacy of Allium sativum (garlic) in reducing some heavy metal accumulation in liver of wistar rats. Food Chem. Toxicol. 2011, 50, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Reinhart, K.M.; Coleman, C.I.; Teevan, C.; Vachhani, P.; White, C.M. Effects of garlic on blood pressure in patients with and without systolic hypertension: A meta-analysis. Ann. Pharmacother. 2008, 42, 1766–1771. [Google Scholar] [CrossRef] [PubMed]
- Ried, K.; Frank, O.R.; Stocks, N.P.; Fakler, P.; Sullivan, T. Effect of garlic on blood pressure: A systematic review and meta-analysis. BMC Cardiovasc. Disord. 2008, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Ried, K.; Frank, O.R.; Stocks, N.P. Aged garlic extract reduces blood pressure in hypertensives: A dose-response trial. Eur. J. Clin. Nutr. 2013, 67, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Tanaka, K.; Sato, E.; Okajima, H. Kinetic and mechanistic studies of allicin as an antioxidant. Org. Biomol. Chem. 2006, 4, 4113–4117. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, K.; Verma, M.K.; Verma, N.; Bhardwaj, S.; Mishra, S. Effect of long term supplementation of active garlic allicin in reducing blood pressure in hypertensive subjects. Int. J. Adv. Med. 2015, 2, 231–234. [Google Scholar] [CrossRef]
- Elkayam, A.; Peleg, E.; Grossman, E.; Shabtay, Z.; Sharabi, Y. Effects of allicin on cardiovascular risk factors in spontaneously hypertensive rats. Isr. Med. Assoc. J. 2013, 15, 170–173. [Google Scholar] [PubMed]
- Ali, M.; Al-Qattan, K.K.; Al-Enezi, F.; Khanafer, R.M.; Mustafa, T. Effect of allicin from garlic powder on serum lipids and blood pressure in rats fed with a high cholesterol diet. Prostaglandins Leukot. Essent. Fatty Acids 2000, 62, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Elkayam, A.; Mirelman, D.; Peleg, E.; Wilchek, M.; Miron, T.; Rabinkov, A.; Sadetzki, S.; Rosenthal, T. The effects of allicin and enalapril in fructose-induced hyperinsulinemic hyperlipidemic hypertensive rats. Am. J. Hypertens. 2001, 14, 377–381. [Google Scholar] [CrossRef]
- García-Trejo, E.M.A.; Arellano-Buendía, A.S.; Argüello-García, R.; Loredo-Mendoza, M.L.; García-Arroyo, F.E.; Arellano-Mendoza, M.G.; Castillo-Hernández, M.C.; Guevara-Balcázar, G.; Tapia, E.; Sánchez-Lozada, L.G.; et al. Effects of allicin on hypertension and cardiac function in chronic kidney disease. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Lisowska-Myjak, B.; Krich, A.; Kołodziejczyk, A.; Pachecka, J.; Gaciong, Z. Urinary proteins, N-acetyl-β-d-glucosaminidase activity and estimated glomerular filtration rate in hypertensive patients with normoalbuminuria and microalbuminuria. Nephrology 2011, 16, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Bonnet, F.; Candido, R.; Nesteroff, S.P.; Burns, W.C.; Kawachi, H.; Shimuzu, F.; Carey, R.M.; De Gasparo, M.; Cooper, M.E. Angiotensin type 2 receptor antagonism confers renal protection in a rat model of progressive renal injury. J. Am. Soc. Nephrol. 2002, 13, 1773–1787. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, V.; Ljungberg, P.; Wartiovaara, J.; Lenkkeri, U.; Kestilä, M.; Jalanko, H.; Holmberg, C.; Tryggvason, K. Nephrin is specifically located at the slit diaphragm of glomerular podocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 7962–7967. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, H.; Kioko, H.; Shimizu, F. Molecular structure and function of the slit diaphragm: Expression of nephrin in proteinuric states and in developing glomeruli. Nephrol. Dial. Transplant. 2002, 17, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Koop, K.; Eikmans, M.; Baelde, H.J.; Kawachi, H.; De Heer, E.; Paul, L.C.; Bruijn, J.A. Expression of podocyte-associated molecules in acquired human kidney diseases. J. Am. Soc. Nephrol. 2003, 14, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Ding, G.; Zhu, J.; Chen, C.; Liang, W.; Franki, N.; Singhal, P.C. Angiotensin II infusion induces nephrin expression changes and podocyte apoptosis. Am. J. Nephrol. 2008, 28, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.M.; Bonnet, F.; Russo, L.M.; Burns, W.C.; Cao, Z.; Candido, R.; Kawachi, H.; Allen, T.J.; Cooper, M.E.; Jerums, G.; et al. Modulation of nephrin in the diabetic kidney: Association with systemic hypertension and increasing albuminuria. J. Hypertens. 2002, 20, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Langham, R.G.; Kelly, D.J.; Cox, A.J.; Gow, R.M.; Holthofer, H.; Gilbert, R.E. Angiotensin II-induced proteinuria and expression of the podocyte slit pore membrane protein, nephrin. Nephrol. Dial. Transplant. 2004, 19, 262–263. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Zhao, H.; Fan, R.R.; Guan, J.; He, L.; Lee, H.M.; Chan, J.C.; Tong, P.C. Renin-angiotensin system activation in renal adipogenesis. Am. J. Physiol. Renal Physiol. 2010, 298, F391–F400. [Google Scholar] [CrossRef] [PubMed]
- Joly, E.; Nonclercq, D.; Caron, N.; Mertens, J.; Flamion, B.; Toubeau, G.; Kramp, R.; Bouby, N. Differential regulation of angiotensin II receptors during renal injury and compensatory hypertrophy in the rat. Clin. Exp. Pharmacol. Physiol. 2005, 32, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Millett, L.J.; Heiderstadt, N.T.; Siragy, H.M.; Johns, R.A.; Carey, R.M. Differential regulation of renal angiotensin subtype AT1A and AT2 receptor protein in rats with angiotensin-dependent hypertension. Hypertension 1999, 33, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Della Bruna, R.; Bernhard, I.; Gess, B.; Schricker, K.; Kurtz, A. Renin gene and angiotensin II AT1 receptor gene expression in the kidneys of normal and of two-kidney/one-clip rats. Pflug. Arch. 1995, 430, 265–272. [Google Scholar] [CrossRef]
- Mehta, P.K.; Griendling, K.K. Angiotensin II cell signaling: Physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 2007, 292, C82–C97. [Google Scholar] [CrossRef] [PubMed]
- Argüello-García, R.; Medina-Campos, O.N.; Pérez-Hernández, N.; Pedraza-Chaverrí, J.; Ortega-Pierres, G. Hypoclorous acid scavenging activities of thioallyl compound from garlic. J. Agric. Food Chem. 2010, 58, 11226–11233. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Shafiee, S.M.; Baluchnejadmojarad, T. Garlic extract reduces serum angiotensin converting enzyme (ACE) activity in nondiabetic and streptozotocin-diabetic rats. Pathophysiology 2007, 14, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Lever, J.M.; Boddu, R.; George, J.F.; Agarwal, A. Heme oxygenase-1 in kidney health and disease. Antioxid. Redox Signal. 2016, 25, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Zarjou, A.; Agarwal, A. Heme oxygenase-1 as a target for TGF-β in kidney disease. Semin. Nephrol. 2012, 32, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, C.; Xiang, Z.G.; Hu, J.J.; Lu, J.M.; Tian, R.B.; Jia, W. Allicin ameliorates cardiac hypertrophy and fibrosis through enhancing of Nrf2 antioxidant signaling pathways. Cardiovasc. Drugs Ther. 2012, 26, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Mendoza, M.G.; Vargas-Robles, H.; Del Valle-Mondragon, L.; Rios, A.; Escalante, B. Prevention of renal injury and endothelial dysfunction by chronic L-arginine and antioxidant treatment. Ren. Fail. 2011, 33, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ma, L.; Xie, R.J.; Xie, Y.S.; Wei, R.B.; Yin, M.; Wang, J.Z.; Chen, X.M. Treatment of 5/6 nephrectomy rats with sulodexide: A novel therapy for chronic renal failure. Acta Pharmacol. Sin. 2012, 33, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Tang, Q.; Li, Z.; Zhao, Z.; Zhang, Z.; Lu, L.; Zhu, T.; Vanhoutte, P.M.; Leung, S.W.; Tu, R.; et al. Prostaglandin E2 regulates renal function in C57/BL6 mouse with 5/6 nephrectomy. Life Sci. 2017, 174, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Varma, P.V.; Chakravarthi, M.; Jyothsna, G. Hypertension in patients with chronic kidney disease. Hypertens. J. 2016, 2, 28–34. [Google Scholar] [CrossRef]
- Vallance, P.; Leone, A.; Calver, A.; Collier, J.; Moncada, S. Accumulation of an endogenous inhibitor of nitric oxide synthesis in chronic renal failure. Lancet 1992, 339, 572–575. [Google Scholar] [PubMed]
- Rabinkov, A.; Miron, T.; Konstantinovski, L.; Wilchek, M.; Mirelman, D.; Weiner, L. The mode of action of allicin: Trapping of radicals and interaction with thiol containing proteins. Biochim. Biophys. Acta 1998, 1379, 233–244. [Google Scholar] [CrossRef]
- Prasad, K.; Laxdal, V.A.; Yu, M.; Raney, B.L. Antioxidant activity of allicin, an active principle in garlic. Mol. Cell. Biochem. 1995, 148, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cao, F.; Tang, Q.Z.; Yan, L.; Dong, Y.G.; Zhu, L.H.; Wang, L.; Bian, Z.Y.; Li, H. Allicin protects against cardiac hypertrophy and fibrosis via attenuating reactive oxygen species-dependent signaling pathways. J. Nutr. Biochem. 2010, 21, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Horev-Azaria, L.; Eliav, S.; Izigov, N.; Pri-Chen, S.; Mirelman, D.; Miron, T.; Rabinkov, A.; Wilcheck, M.; Jacob-Hirsch, J.; Amariglio, N.; et al. Allicin up-regulates cellular glutathione level in vascular endothelial cells. Eur. J. Nutr. 2009, 48, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [PubMed]
- Kaye, A.D.; Nossaman, B.D.; Ibrahim, I.N.; Feng, C.J.; McNamara, D.B.; Agrawal, K.C.; Kadowitz, P.J. Analysis of responses of allicin, a compound from garlic, in the pulmonary vascular bed of the cat and in the rat. Eur. J. Pharmacol. 1995, 276, 21–26. [Google Scholar] [CrossRef]
- Freeman, F.; Kodera, Y. Garlic chemistry: Stability of S-(2-propenyl)-2-propene-1-sulfinothioate (allicin) in blood, solvents, and simulated physiological fluids. Agric. Food Chem. 1995, 43, 2332–2338. [Google Scholar] [CrossRef]
- SwissDock. Available online: http://swissdock.ch (accessed on 12–18 November 2016).
- SWISS-MODEL. Available online: https://swissmodel.expasy.org/ (accessed on 11 November 2016).
- I-TASSER. Available online: http://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 12 June 2017).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | CKD | CKDA | CKDL | |
|---|---|---|---|---|
| Body weight (g) | 376.53 ± 2.38 | 331.69 ± 30.81 * | 364.45 ± 16.94 + | 370.6 ± 6.05 + |
| Serum creatinine (mg/dL) | 0.42 ± 0.02 | 1.43 ± 0.17 * | 1.122 ± 0.10 *,+ | 0.825 ± 0.28 *,+,# |
| Creatinine clearance (mL/min) | 1.42 ± 0.12 | 0.751 ± 0.09 * | 1.05 ± 0.10 *,+ | 1.19 ± 0.09 *,+,# |
| Blood urea nitrogen (mg/dL) | 4.16 ± 0.40 | 63.16 ± 10.96 * | 42.18 ± 6.21 *,+ | 38.66 ± 3.77 *,+ |
| C | CKD | CKDA | CKDL | |
|---|---|---|---|---|
| Oxidized proteins in the cortex (DNPH nmol/mg protein) | 0.90 ± 0.30 | 122.2 ± 20.99 * | 59.87 ± 4.06 *,+ | 76.7 ± 6.29 *,+ |
| Oxidized proteins in the medulla (DNPH nmol/mg protein) | 1.09 ± 0.30 | 74.42 ± 1.82 * | 36.27 ± 5.19 *,+ | 63.04 ± 3.46 *,+ |
| Lipid peroxidation in the cortex (4HNE nmol/mg protein) | 1.27 ± 0.28 | 6.18 ± 0.32 * | 2.26 ± 0.4 + | 2.04 ± 0.88 + |
| Lipid peroxidation in the medulla (4HNE nmol/mg protein) | 1.06 ± 0.36 | 6.13 ± 1.12 * | 1.88 ± 0.28 + | 0.92 ± 0.21 + |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García Trejo, E.M.Á.; Arellano Buendía, A.S.; Sánchez Reyes, O.; García Arroyo, F.E.; Arguello García, R.; Loredo Mendoza, M.L.; Tapia, E.; Sánchez Lozada, L.G.; Osorio Alonso, H. The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan. Int. J. Mol. Sci. 2017, 18, 1980. https://doi.org/10.3390/ijms18091980
García Trejo EMÁ, Arellano Buendía AS, Sánchez Reyes O, García Arroyo FE, Arguello García R, Loredo Mendoza ML, Tapia E, Sánchez Lozada LG, Osorio Alonso H. The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan. International Journal of Molecular Sciences. 2017; 18(9):1980. https://doi.org/10.3390/ijms18091980
Chicago/Turabian StyleGarcía Trejo, Ehécatl Miguel Ángel, Abraham Said Arellano Buendía, Omegar Sánchez Reyes, Fernando Enrique García Arroyo, Raúl Arguello García, María Lilia Loredo Mendoza, Edilia Tapia, Laura Gabriela Sánchez Lozada, and Horacio Osorio Alonso. 2017. "The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan" International Journal of Molecular Sciences 18, no. 9: 1980. https://doi.org/10.3390/ijms18091980
APA StyleGarcía Trejo, E. M. Á., Arellano Buendía, A. S., Sánchez Reyes, O., García Arroyo, F. E., Arguello García, R., Loredo Mendoza, M. L., Tapia, E., Sánchez Lozada, L. G., & Osorio Alonso, H. (2017). The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan. International Journal of Molecular Sciences, 18(9), 1980. https://doi.org/10.3390/ijms18091980