Understanding the Inguinal Sinus in Sheep (Ovis aries)—Morphology, Secretion, and Expression of Progesterone, Estrogens, and Prolactin Receptors

 , , ,
, , , 
Abstract
1. Introduction
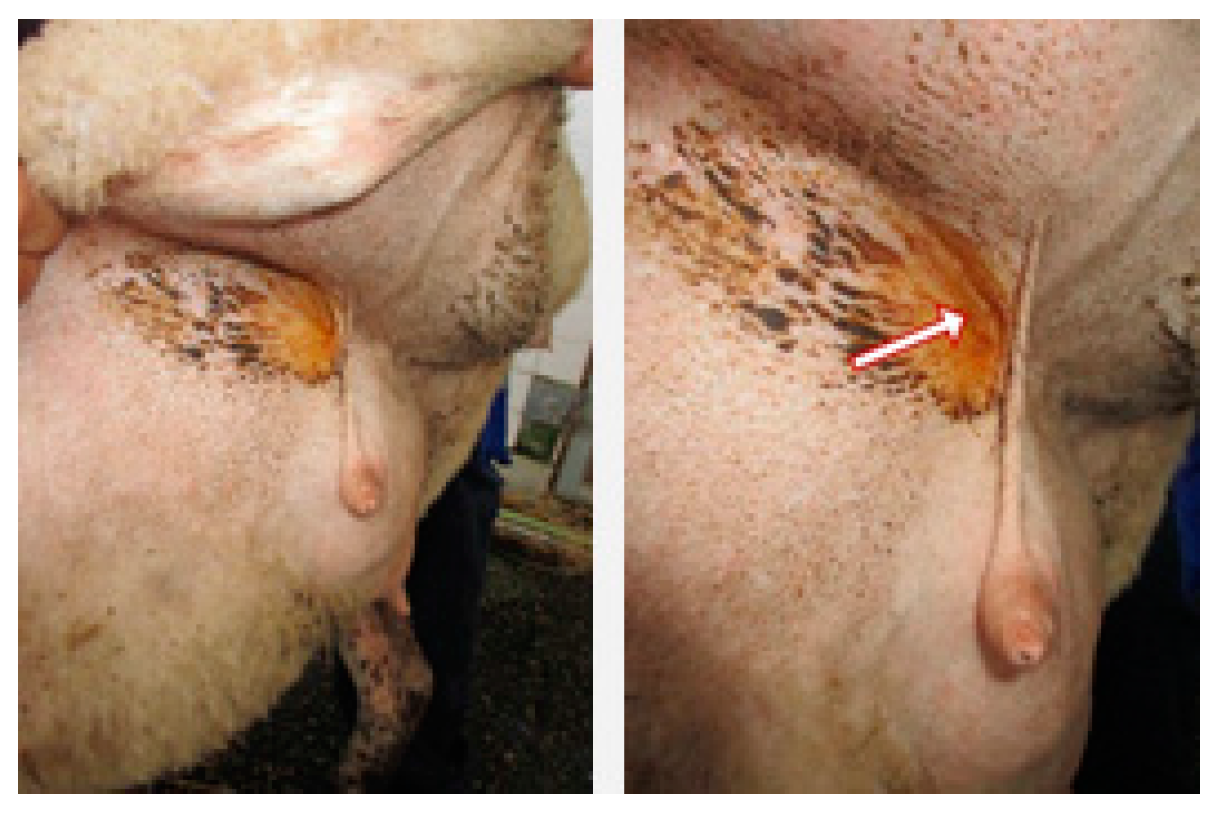
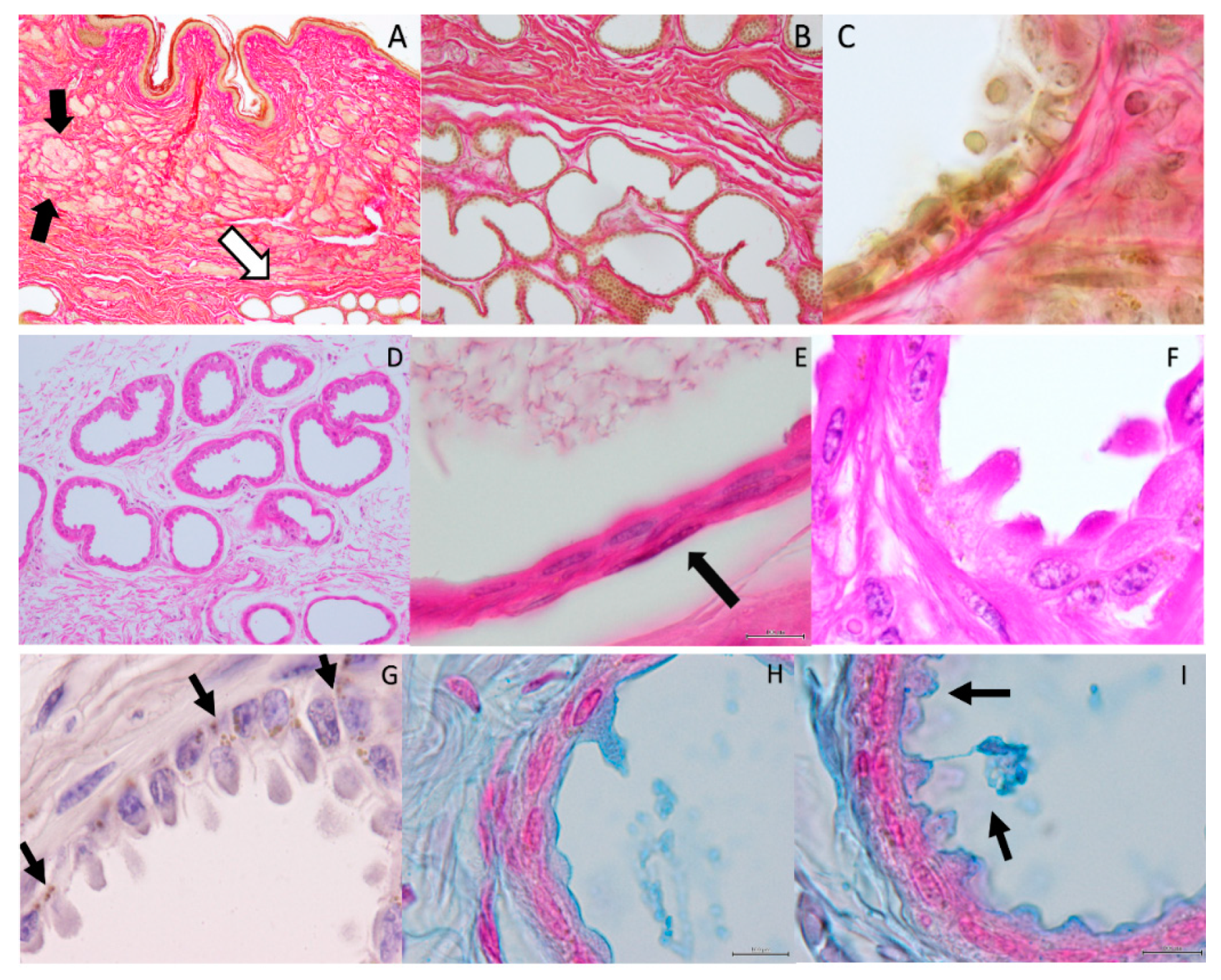
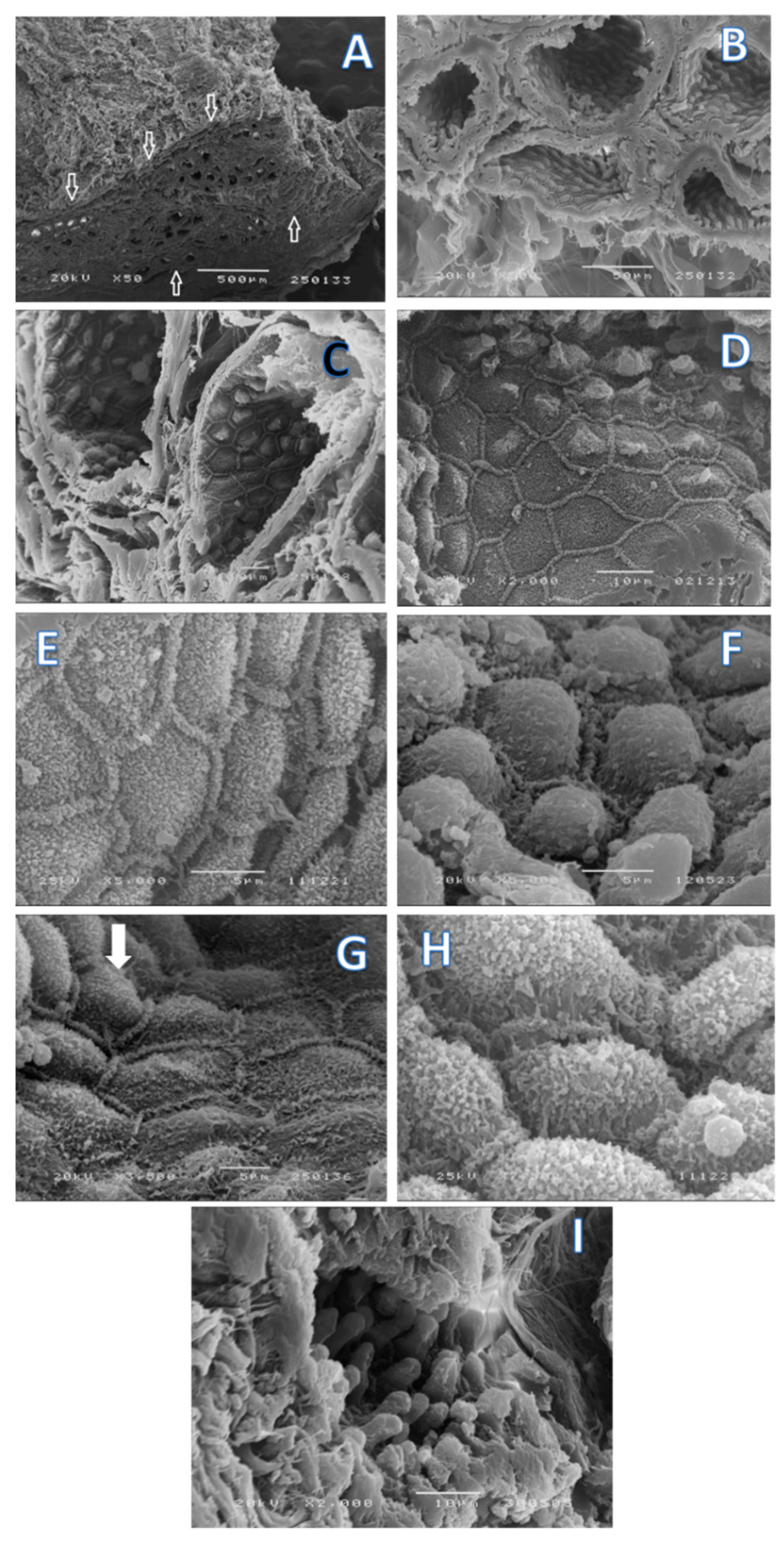
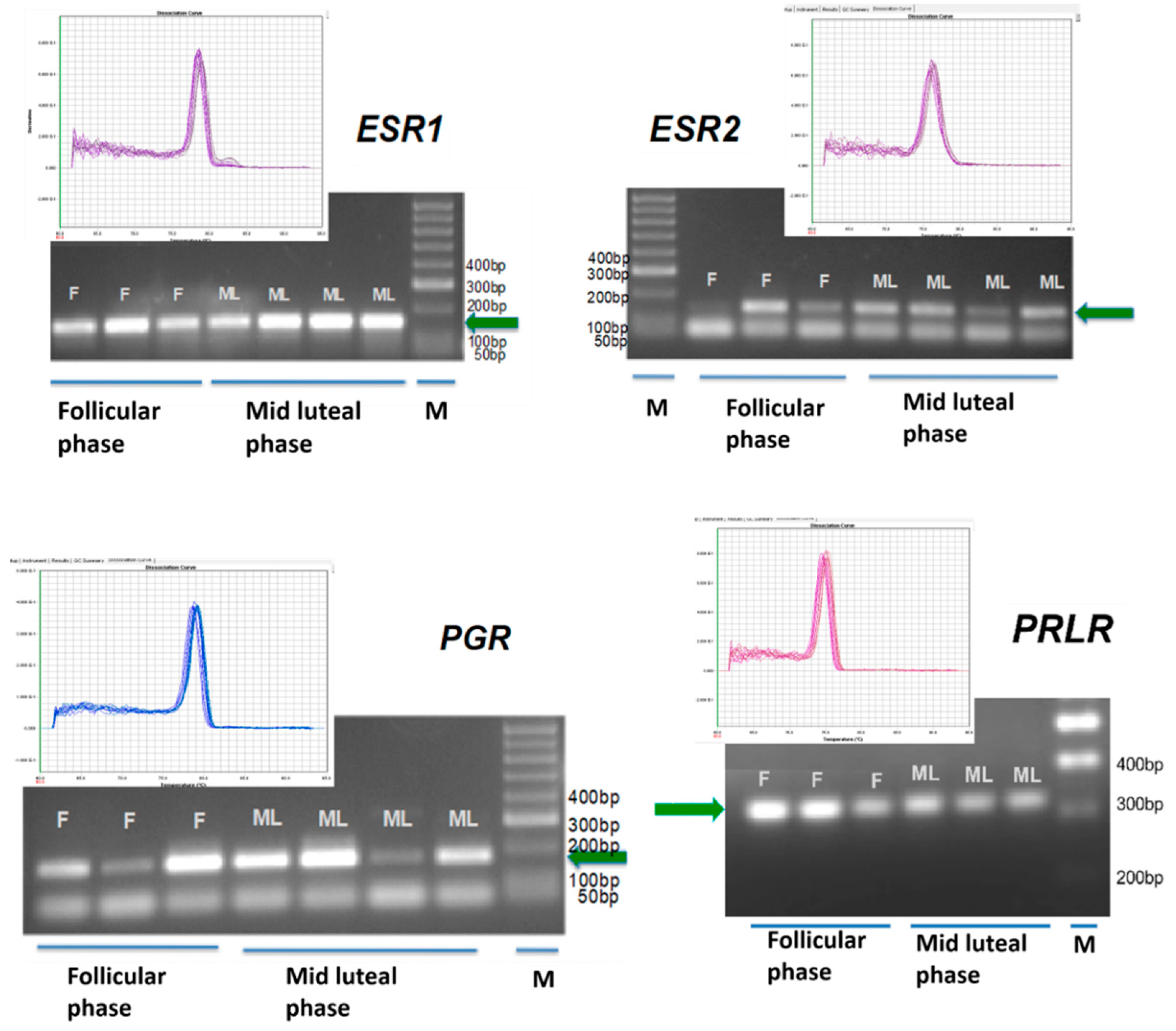
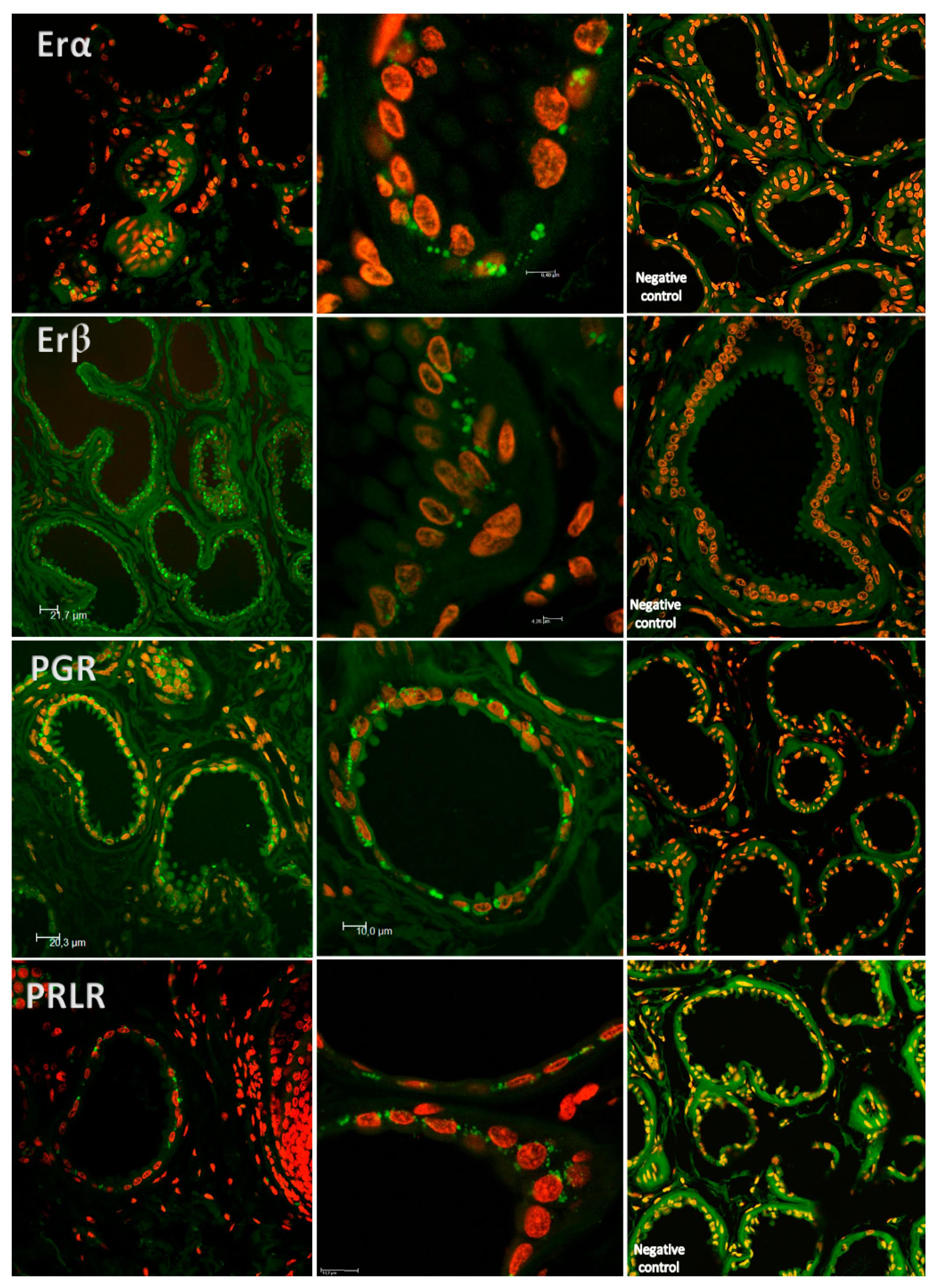
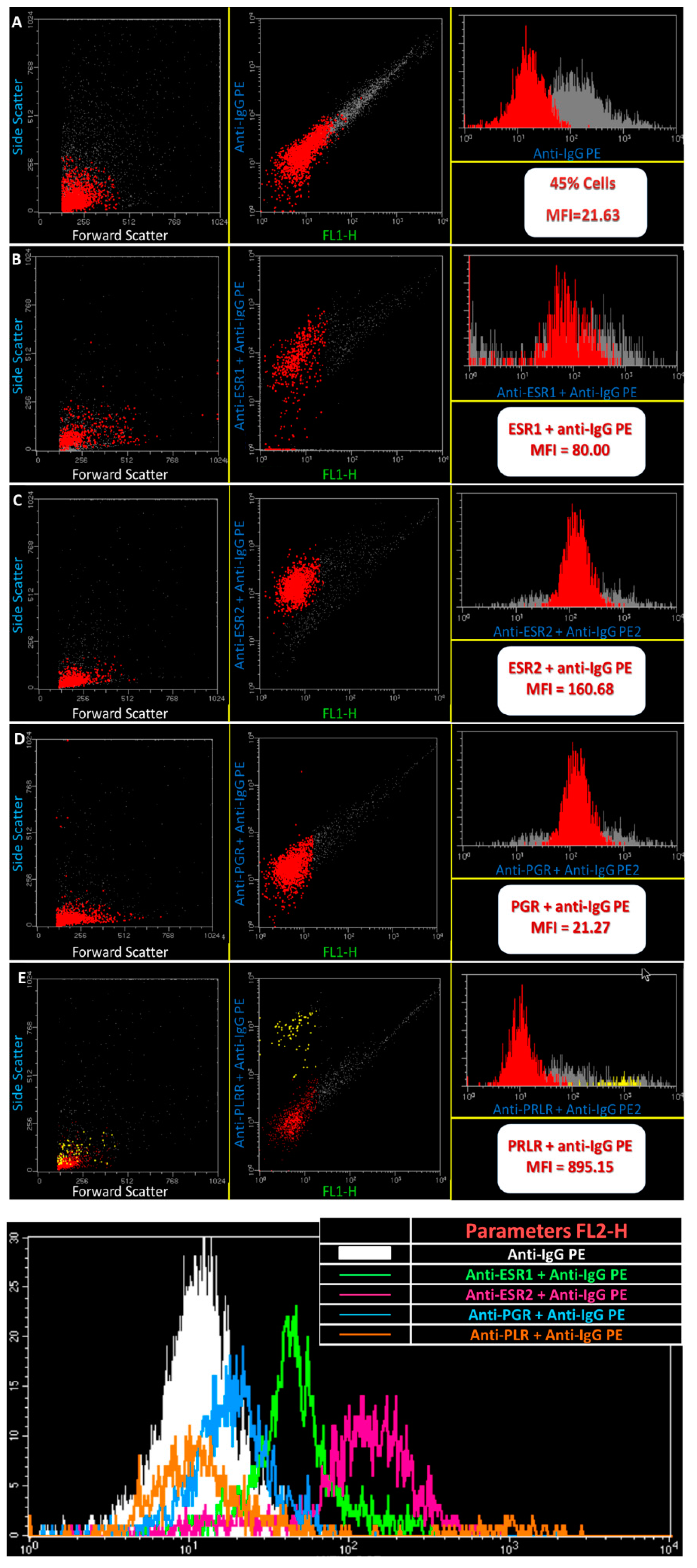
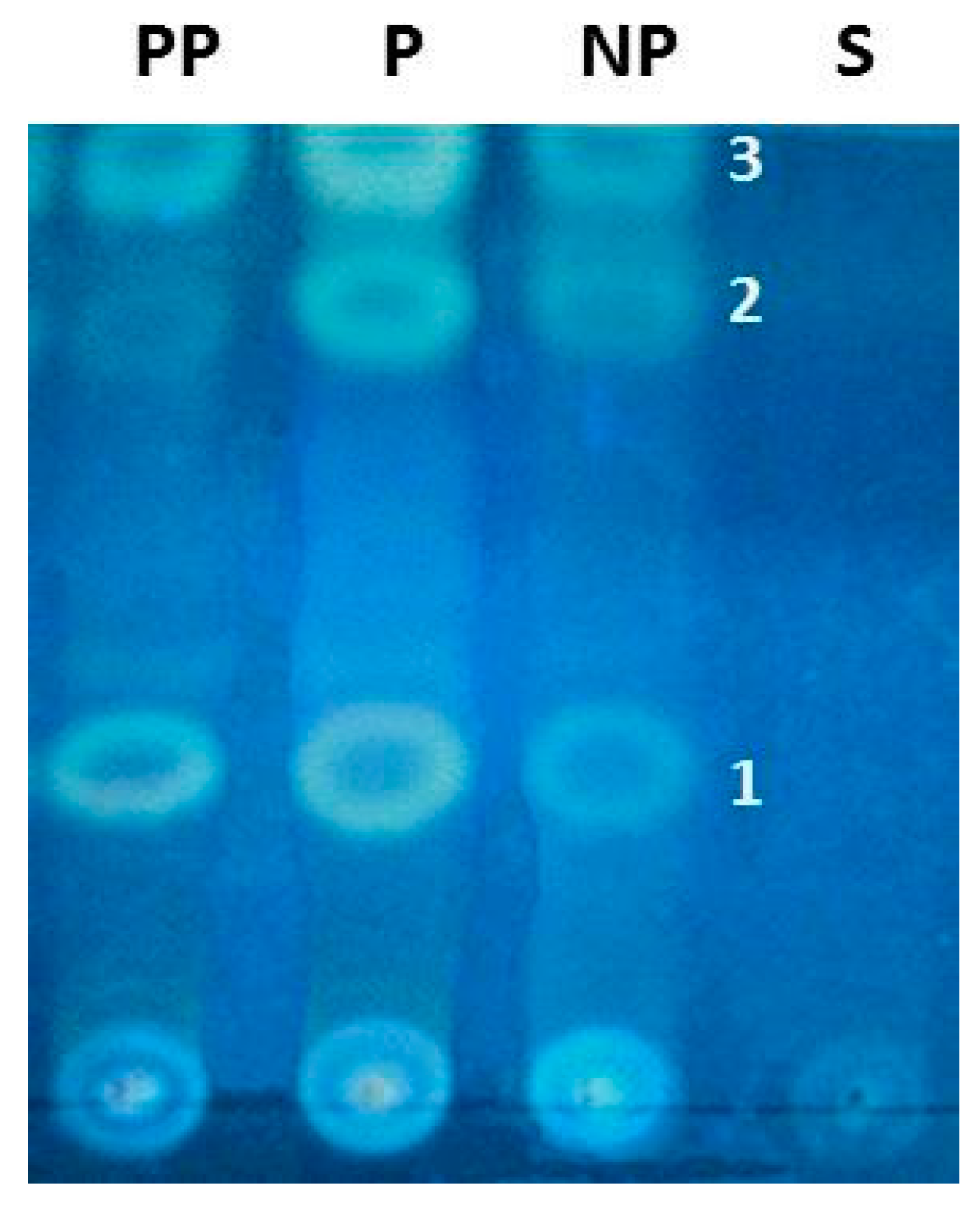
2. Results
3. Discussion
4. Materials and Methods
4.1. Histology Evaluation
4.2. Scanning Electron Microscopy
4.3. Flow Cytometry Analysis
4.4. Laser-Scanning Confocal Microscopy
4.5. Genomic Analysis
4.6. Progesterone Analysis
4.7. Chemical Studies
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brockman, J.H.; Snowdon, C.; Roper, T.; Naguib, M.; Wynne-Eduards, K. Advances in the Study of Behavior; Elsevier Inc.: San Diego, CA, USA, 2009; Volume 36, ISBN 978-0-12-374474-6. [Google Scholar]
- Fuchs, S. Optimality of parental investment: The influence of nursing on reproductive success of mother and female young house mice. Behav. Ecol. Sociobiol. 1982, 10, 39–51. [Google Scholar] [CrossRef]
- Voloschin, L.M.; Tramezzani, J.H. Milk Ejection Reflex Linked to Slow Wave Sleep in Nursing Rats. Endocrinology 1979, 105, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Marnila, P.; Gill, H.S. Milk immunoglobulins and complement factors. Br. J. Nutr. 2000, 84, 75–80. [Google Scholar] [CrossRef]
- Schaal, B. Pheromones for Newborns; Mucignat-Caretta, C., Raton, B., Eds.; Taylor & Francis: Abingdon, UK, 2014; Chapter 17; ISBN-13: 978-1-4665-5341. [Google Scholar]
- Nowak, R. Suckling, Milk, and the Development of Preferences toward Maternal Cues by Neonates: From Early Learning to Filial Attachment? Adv. Study Behav. 2006, 36, 1–58. [Google Scholar] [CrossRef]
- Serra, J.; Ferreira, G.; Mirabito, L.; Lévy, F.; Nowak, R. Post-oral and Perioral Stimulations during Nursing Enhance Appetitive Olfactory Memory in Neonatal rabbits. Chem. Senses 2009, 34, 405–413. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Litwack, G. Vitamines and Hormones in: Pheromones; Elsevier: San Diego, CA, USA, 2010; ISBN 978-0-12-381516-3. [Google Scholar]
- Mykytowycz, R. Skin Glands as Organs of Communication in Mammal. J. Investig. Dermatol. 1974, 62, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.; González-Mariscal, G.; Beyer, C. Chin marking behavior, sexual receptivity, and pheromone emission in steroid-treated, ovariectomized rabbits. Horm. Behav. 1990, 24, 1–13. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, G.; Chirino, R.; Hudson, R. Prolactin Stimulates Emission of Nipple Pheromone in Ovariectomized New Zealand White Rabbits. Biol. Reprod. 1994, 50, 373–376. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brennan, P.; Keverne, E.B. Biological complexity and adaptability of simple mammalian olfactory memory systems. Neurosci. Biobehav. Rev. 2015, 50, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Oboti, L.; Ibarra-Soria, X.; Pérez-Gómez, A.; Schmid, A.; Pyrski, M.; Paschek, N.; Kircher, S.; Logan, D.W.; Leinders-Zufall, T.; Zufall, F.; et al. Pregnancy and estrogen enhance neural progenitor-cell proliferation in the vomeronasal sensory epithelium. BMC Biol. 2015, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Rekwot, P.I.; Ogwu, D.; Oyedipe, E.O.; Sekoni, V.O. The role of pheromones and biostimulation in animal reproduction. Anim. Reprod. Sci. 2001, 65, 157–170. [Google Scholar] [CrossRef]
- Parillo, F.; Diverio, S. Glycocomposition of the apocrine interdigital gland secretions in the fallow deer (Dama dama). Res. Vet. Sci. 2009, 86, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R. Neonatal survival: Contributions from behavioural studies in sheep. Appl. Anim. Behav. Sci. 1996, 49, 61–72. [Google Scholar] [CrossRef]
- Nowak, R.; Porter, R.H.; Lévy, F.; Orgeur, P.; Schaal, B. Role of mother–young interactions in the survival of offspring in domestic mammals. Rev. Reprod. 2000, 5, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Vince, M.A.; Ward, T.M. The responsiveness of newly born Clun forest lambs to odour sources in the ewe. Behaviour 1984, 89, 117–127. [Google Scholar] [CrossRef]
- Schaal, B.; Orgeur, P.; Arnould, C. Olfactory preferences in newborn lambs: Possible influence of prenatal experience. Behaviour 1995, 132, 5351–5365. [Google Scholar] [CrossRef]
- Vince, M.A.; Lynch, J.J.; Mottershead, B.E.; Green, G.C. Interactions between normal ewes and newly born lambs deprived of visual, olfactory and tactile sensory information. Appl. Anim. Behav. Sci. 1987, 19, 119–136. [Google Scholar] [CrossRef]
- Vince, M.A.; Billing, A.E. Infancy in the sheep: The part played by sensory stimulation in bounding between ewe and lamb. In Advances in Infancy Research; Lipsitt, L.P., Rovee-Collier, C., Eds.; Ablex Noorwood: New York, NY, USA, 1986; Volume 4, pp. 1–37. [Google Scholar]
- Logan, D.W.; Brunet, L.J.; Webb, W.R.; Cutforth, T.; Ngai, J.; Stowers, L. Learned Recognition of Maternal Signature Odors Mediates the First Suckling Episode in Mice. Curr. Biol. 2012, 22, 1998–2007. [Google Scholar] [CrossRef] [PubMed]
- Schaal, B.; Coureaud, G.; Doucet, S.; Delaunay, E.l.; Allam, M.; Moncomble, A.-S.; Montigny, D.; Patris, B.; Holley, A. Mammary olfactory signalisation in females and odour pocessing in neonates: Ways evolved by rabbits and humans. Behav. Brain Res. 2009, 200, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Morrow-Tesch, J.; McGlone, J.J. Sources of maternal odors and the development of odor preferences in baby pigs. J. Anim. Sci. 1990, 68, 3563–3571. [Google Scholar] [CrossRef] [PubMed]
- Schaal, B. Mammary Odor Cues and Pheromones: Mammalian Infant-Directed Communication about Maternal State, Mammae, and Milk. In Vitamins and Hormones; Litwack, G., Ed.; Elsevier Inc.: San Diego, CA, USA, 2009; Chapter 4; Volume 83, pp. 83–136. [Google Scholar]
- Vaglio, S.; Minicozzi, P.; Bonometti, E.; Mello, G.; Chiarelli, B. Volatile signals during pregnancy: A possible chemical basis for mother-infant recognition. J. Chem. Ecol. 2009, 53, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Doucet, S.; Sooussignan, R.; Sagot, P.; Schaal, B. The secretion of areolar (Montgomery’s) glands from lactating women elicits selective, unconditional responses in neonates. PLoS ONE 2009, 4, e7579. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Legendre, A.; Faure, P.; Tiesset, H.; Potin, C.; Jakob, I.; Sicard, G.; Schaal, B.; Artur, Y.; Coureaud, G.; Heydel, J.M. When the Nose Must Remain Responsive: Glutathione Conjugation of the Mammary Pheromone in the Newborn Rabbit. Chem. Sens. 2014, 39, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.; Rojas, C.; Carolina Rojas, L.; Martínez-Gómez, M.; Distel, H. Rabbit Nipple-Search Pheromone versus Rabbit Mammary Pheromone Revisited. In Chemical Signals in Vertebrates; Muller-Schwarze, D., Ed.; Springer Link: London, UK, 2008; pp. 315–324. [Google Scholar]
- Coureaud, G.; Charra, R.; Datiche, F.; Sinding, C.; Thomas-Danguin, T.; Languille, S.; Hars, B.; Schaal, B. A pheromone to behave, a pheromone to learn: The rabbit mammary pheromone. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2010, 196, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Brus, M.; Meurisse, M.; Franceschini, I.; Keller, M.; Lévy, F. Evidence for cell proliferation in the sheep brain and its down-regulation by parturition and interactions with the young. Horm. Behav. 2010, 58, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, D. Breeding the Flock: Modern Research and Reproduction in Sheep; Inkata Pr.: Berlin, Germany, 1988; ISBN 0909605459. [Google Scholar]
- Vince, M.A. Newborn lambs and their dams: The interaction that leads to suckling. Adv. Study Behav. 1993, 22, 239–268. [Google Scholar]
- Sell, C. A Fragrant Introduction to Terpenoid Chemistry; The Royal Society of Chemistry: Cambridge, UK, 2003; ISBN 0-85404-681-X. [Google Scholar]
- Poindron, P.; Lévy, F. Physiological, sensory and experiential determinants of maternal behaviour in sheep. In Mammalian Parenting: Biochemical, Neurobiological and Behavioral Determinants; Krasnegor, N.A., Bridges, R.S., Eds.; Oxford University Press: New York, NY, USA, 1990; pp. 133–156, 485–488. ISBN1 0195056000. ISBN2 9780195056006. [Google Scholar]
- Poindron, P.; Nowak, R.; Lévy, F.; Porter, R.H.; Schaal, B. Development of exclusive mother-young bonding in sheep and goats. In Oxford Reviews of Reproductive Biology; Michigan, S.R., Ed.; Oxford University Press: Oxford, UK, 1993; Volume 15, pp. 311–364. [Google Scholar]
- Foitzik, K.; Krause, K.; Conrad, F.; Nakamura, M.; Funk, W.; Paus, R. Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine promoter of apoptosis-driven hair follicle regression. Am. J. Pathol. 2006, 168, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Egli, M.; Leeners, B.; Kruge, TH. Prolactin secretion patterns: Basic mechanisms and clinical implications for reproduction. Reproduction 2010, 140, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Foitzik, K.; Krause, K.; Nixon, A.J.; Ford, C.A.; Ohnemus, U.; Pearson, A.J.; Paus, R. Prolactin and its receptor are expressed in murine hair follicle epithelium, show hair cycle-dependent expression, and induce catagen. Am. J. Pathol. 2003, 162, 1611–1621. [Google Scholar] [CrossRef]
- Foitzik, K.; Langan, E.A.; Paus, R. Prolactin and the skin: A dermatological perspective on an ancient pleiotropic peptide hormone. J. Investig. Dermatol. 2009, 129, 1071–1087. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [PubMed]
- Kikuyama, S.; Yamamoto, K.; Iwata, T.; Toyoda, F. Peptide and protein pheromones in amphibians. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 132, 69–74. [Google Scholar] [CrossRef]
- González-Mariscal, G. Neuroendocrinology of Maternal Behaviour in the Rabbit. Horm. Behav. 2001, 40, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, A.M.; Green, A.R.; Karthik, S.; Alizadeh, Y.; Hughes, T.A.; Harkins, L.; Ellis, I.O.; Robertson, J.F.; Paish, E.C.; Saunders, P.T.; et al. Nuclear and cytoplasmic expression of ERbeta1, ERbeta2, and ERbeta5 identifies distinct prognostic outcome for breast cancer patients. Clin. Cancer. Res. 2008, 14, 5228–5235. [Google Scholar] [CrossRef] [PubMed]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A Transmembrane Intracellular Estrogen Receptor Mediates Rapid Cell Signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Robert, X.-D.; Song, R.X.-D.; Zhang, Z.; Santen, R.J. Estrogen Rapid Action via Protein Complex Formation Involving ERα and Src; Cell Press: Cambridge, MA, USA, 2005; Volume 16, pp. 347–353. [Google Scholar]
- Simoncini, T.; Rabkin, E.; Liao, J.K. Membrane estrogen receptor interaction with ph osphatidylinositol 3-kinase in endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Hammes, S.R. The further redefining of steroid-mediated signalling. Proc. Nat. Acad. Sci. USA 2003, 100, 2168–2170. [Google Scholar] [CrossRef] [PubMed]
- Cato, A.; Nestl, A.; Mink, S. Rapid actions of steroid receptors in cellular signaling pathways. Science’s STKE 2002. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R. Cell localization, physiology and nongenomic actions of estrogen receptors. J. Appl. Physiol. 2001, 91, 1860–1867. [Google Scholar] [PubMed]
- Wittliff, J.L.; Hilf, R.; Brooks, W.F., Jr.; Savlov, E.D.; Hall, T.; Robert, A.O. Specific estrogen-binding capacity of the cytoplasmic receptor in normal and neoplastic breast tissues of humans. J. Cancer Res. 1972, 11, 1–73. [Google Scholar]
- Welsh, A.W.; Lannin, D.R.; Gregory, S.; Young, G.S.; Mark, E.; Sherman, M.E.; Jonine, D.; Figueroa, J.D.; Lynn, N.; Henry, N.L.; et al. Cytoplasmic Estrogen Receptor in breast cancer. Clin. Cancer Res. 2012, 18, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. Estrogen receptor interaction with estrogen response elements. Nucleic Acids Res. 2001, 29, 2905–2919. [Google Scholar] [CrossRef] [PubMed]
- McKenna, N.J.; O’Malley, B.W. Combinatorial control of gene expression by nuclear receptors and coregulators. Cell 2002, 108, 465–474. [Google Scholar] [CrossRef]
- Stossi, F.; Likhite, V.S.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Estrogen-occupied estrogen receptor represses cyclin G2 gene expression and recruits a repressor complex at the cyclin G2 promoter. J. Biol. Chem. 2006, 281, 16272–16278. [Google Scholar] [CrossRef] [PubMed]
- Farach-Carson, M.C.; Davis, P.J. Steroid hormone interactions with target cells: Cross talk between membrane and nuclear pathways. J. Pharmacol. Exper. Therap. 2003, 30, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Alexandre-Pires, G.; Martins, C.; Miguel Galvão, A.; Correia, M.; Ramilo, D.; Quaresma, M.; Ligeiro, D.; Nunes, T.; Caldeira, R.M.; Ferreira-Dias, G. Morphological Aspects and Expression of Estrogen and Progesterone Receptors in the Interdigital Sinus in Cyclic Ewes. Microsc. Res. Tech. 2014, 77, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Zaviacic, M.; Zajickova, M.; Blazekova, J.; Donarov, L.; Stvrtina, S.; Mikuleck, M.; Zaviacic, T.; Holom, A.N.K.; Breza, J. Size, macroanatomy and histology of the normal prostate in the adult human female: A minireview. J. Histotechnol. 2000, 21, 61–69. [Google Scholar] [CrossRef]
- Gesase, A.P.; Satoh, Y.; Ono, K. Secretagogue-induced apocrine secretion in the rat Harderian gland of the rat. Cell Tissue Res. 1996, 285, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Gesase, A.P.; Satoh, Y. Apocrine secretory mechanism: Recent findings and unresolved problems. Histol. Histopathol. 2003, 18, 597–608. [Google Scholar] [PubMed]
- Jarret, A. Physiology and Pathophysiology of the Skin: The Sweat Gland, Skin Permeation, Lymphatics, the Nails; Academic Press: Salt Lake, UT, USA, 1978; ISBN1 0123806054. ISBN2 9780123806055. [Google Scholar]
- Pourlis, A.F. Functional morphological characteristics of the interdigital sinus in the sheep. Folia Morphol. 2010, 69, 107–111. [Google Scholar]
- Cristofoletti, P.T.; Ribeiro, A.F.; Terra, W.R. Apocrine secretion of amylase and exocytosis of trypsin along the midgut of Tenebrio molitor larvae. J. Insect. Physiol. 2001, 47, 143–155. [Google Scholar] [CrossRef]
- Craven, A.J.; Ormandy, C.J.; Robertson, F.G.; Wilkins, R.J.; Kelly, P.A.; Nixon, A.J.; Pearson, A.J.; Craven, T. Prolactin signaling influences the timing mechanism of the hair follicle: Analysis of hair growth cycles in prolactin receptor knockout mice. Endocrinology 2001, 142, 2533–2539. [Google Scholar] [CrossRef] [PubMed]
- Craven, A.J.; Nixon, A.J.; Ashby, M.G.; Ormandy, C.J.; Blazek, K.; Wilkins, R.J.; Pearson, A.J. Prolactin delays hair regrowth in mice. J. Endocrinol. 2006, 19, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Goffin, V.; Bernichtein, S.; Touraine, P.; Kelly, P.A. Development and potential clinical uses of human prolactin receptor antagonists. Endocr. Rev. 2005, 26, 400–422. [Google Scholar] [CrossRef] [PubMed]
- Horseman, N.D.; Gregerson, K.A. Prolactin actions. Soc. Endocrinol. 2014, 52, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Oftedal, O.T. The origin of lactation as a water source for parchment-shelled eggs. J. Mammary Gland Biol. Neoplasia 2002, 7, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Oftedal, O.T. The mammary gland and its origin during synapsid evolution. J. Mammary Gland Biol. Neoplasia 2002, 7, 225–252. [Google Scholar] [CrossRef] [PubMed]
- Oftedal, O.T. The evolution of milk secretion and its ancient origins. Animal 2012, 6, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Nicoll, C.S. Prolactin: Ontogeny and evolution of prolactin’s functions. Fed. Proc. 1980, 39, 2561–2566. [Google Scholar]
- Srivastava, S.; Matsuda, M.; Hou, Z.; Bailey, J.P.; Kitazawa, R.; Herbst, M.P.; Horseman, N.D. Receptor activator of NF-κB ligand induction via Jak2 and Stat5a in mammary epithelial cells. J. Biol. Chem. 2003, 278, 46171–46178. [Google Scholar] [CrossRef] [PubMed]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Horseman, N.D.; Yu-Lee, L.Y. Transcriptional regulation by the helix bundle peptide hormones: Growth hormone, prolactin, and hematopoietic cytokines. Endocr. Rev. 1994, 15, 627–649. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.L. Molecular mechanisms of prolactin and its receptor. Endocr. Rev. 2012, 33, 504–525. [Google Scholar] [CrossRef] [PubMed]
- Sidis, Y.; Horseman, N.D. Prolactin induces rapid p95/p70 tyrosine phosphorylation, and protein binding to GAS-like sites in the anx Icp35 and c-fos genes. Endocrinology 1994, 134, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Standke, G.J.R.; Meier, V.S.; Groner, B. Mammary gland factor activated by prolactin in mammary epithelial cells and acute-phase response factor activated by interleukin-6 in liver cells share DNA binding and transactivation potential. Molecul. Endocrinol. 1994, 8, 469–477. [Google Scholar] [CrossRef]
- Schramek, D.A.; Sigl, V.; Kenner, L.; Pospisilik, J.A.; Lee, H.J.; Hanada, R.; Joshi, P.A.; Aliprantis Glimcher, L.; Pasparakis, M.; Khokha, R.; et al. Osteoclast differentiation factor RANKL controls development Leibbrandt of progestin-driven mammary cancer. Nature 2010, 468, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Bonizzi, G.; Seagroves, T.N.; Greten, F.R.; Johnson, R.; Schmidt, E.V.; Karin, M. IKKα provides an essential link between RANK signaling and cyclin D1 expression during mammary gland development. Cell 2001, 107, 763–775. [Google Scholar] [CrossRef]
- Baxter, F.O.; Came, P.J.; Abell, K.; Kedjouar, B.; Huth, M.; Rajewsky, K.; Pasparakis, M.; Watson, C.J. IKKβ/2 induces TWEAK and apoptosis in mammary epithelial cells. Development 2006, 133, 3485–3494. [Google Scholar] [CrossRef] [PubMed]
- Müller-Schwarze, D. The chemical ecology of ungulates. Appl. Anim. Sci. 1991, 29, 389–402. [Google Scholar] [CrossRef]
- Luna, LG. Histopathologic Methods and Color Atlas of Special Stains and Tissue Artifacts, 1st ed.; Johnson Printers: Downers Grove, IL, USA, 1992. [Google Scholar]
- Bruno, T.J.; Svoronos, P.D.N. Handbook of Basic Tables for Chemical Analysis, 2nd ed.; Taylor & Francis Group: New York, NY, USA, 2004; Chapter 3; pp. 177–212. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Acession Number) | Sequence 5′–3′ Amplicon (Base Pairs) |
|---|---|
| ESR1 (XM_015097472.1) | Forward: CCATGGAATCTGCCAAGGAG (167 bp) Reverse: ATCAATTGTGCACTGGTTGGT |
| ESR2 (NM_001009737.1) | Forward: TGGAGTCTGGTCATGTGAAGGA (150 bp) Reverse: TCATAGCACTTCCGCAGTCG |
| PGR (XM_015100878.1) | Forward: CAGCCAGAGCCCACAGTACA (176 bp) Reverse: TGCAATCGTTTCTTCCAGCA |
| PRLR (NM_001009204.1) | Forward: GTCTCCACCCACCCTGACTG (320 bp) Reverse: AAGCCACTGCCCAGACCATA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandre-Pires, G.; Martins, C.; Galvão, A.M.; Miranda, M.; Silva, O.; Ligeiro, D.; Nunes, T.; Ferreira-Dias, G. Understanding the Inguinal Sinus in Sheep (Ovis aries)—Morphology, Secretion, and Expression of Progesterone, Estrogens, and Prolactin Receptors. Int. J. Mol. Sci. 2017, 18, 1516. https://doi.org/10.3390/ijms18071516
Alexandre-Pires G, Martins C, Galvão AM, Miranda M, Silva O, Ligeiro D, Nunes T, Ferreira-Dias G. Understanding the Inguinal Sinus in Sheep (Ovis aries)—Morphology, Secretion, and Expression of Progesterone, Estrogens, and Prolactin Receptors. International Journal of Molecular Sciences. 2017; 18(7):1516. https://doi.org/10.3390/ijms18071516
Chicago/Turabian StyleAlexandre-Pires, Graça, Catarina Martins, António M. Galvão, Margarida Miranda, Olga Silva, Dário Ligeiro, Telmo Nunes, and Graça Ferreira-Dias. 2017. "Understanding the Inguinal Sinus in Sheep (Ovis aries)—Morphology, Secretion, and Expression of Progesterone, Estrogens, and Prolactin Receptors" International Journal of Molecular Sciences 18, no. 7: 1516. https://doi.org/10.3390/ijms18071516
APA StyleAlexandre-Pires, G., Martins, C., Galvão, A. M., Miranda, M., Silva, O., Ligeiro, D., Nunes, T., & Ferreira-Dias, G. (2017). Understanding the Inguinal Sinus in Sheep (Ovis aries)—Morphology, Secretion, and Expression of Progesterone, Estrogens, and Prolactin Receptors. International Journal of Molecular Sciences, 18(7), 1516. https://doi.org/10.3390/ijms18071516