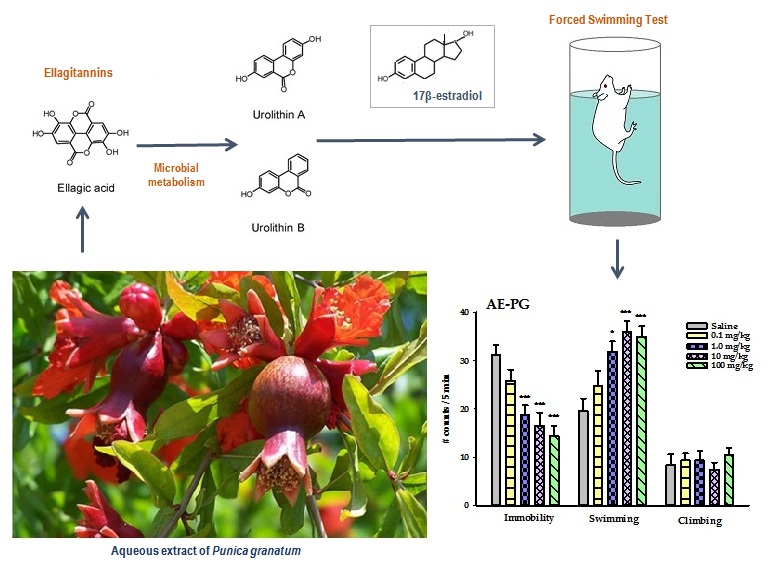
Aqueous Extract of Pomegranate Alone or in Combination with Citalopram Produces Antidepressant-Like Effects in an Animal Model of Menopause: Participation of Estrogen Receptors

 , and
, and 
Abstract
:
1. Introduction
2. Results
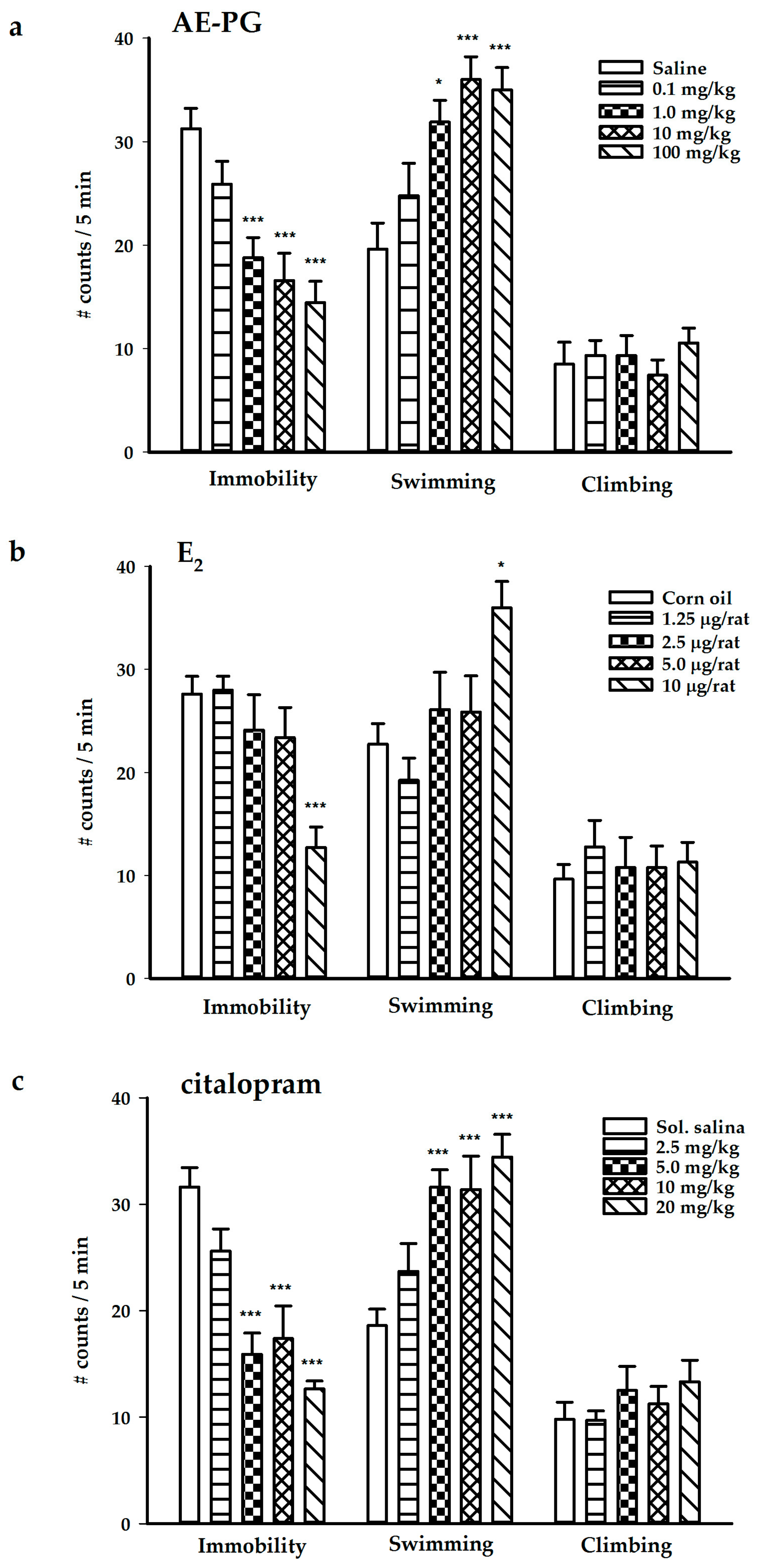
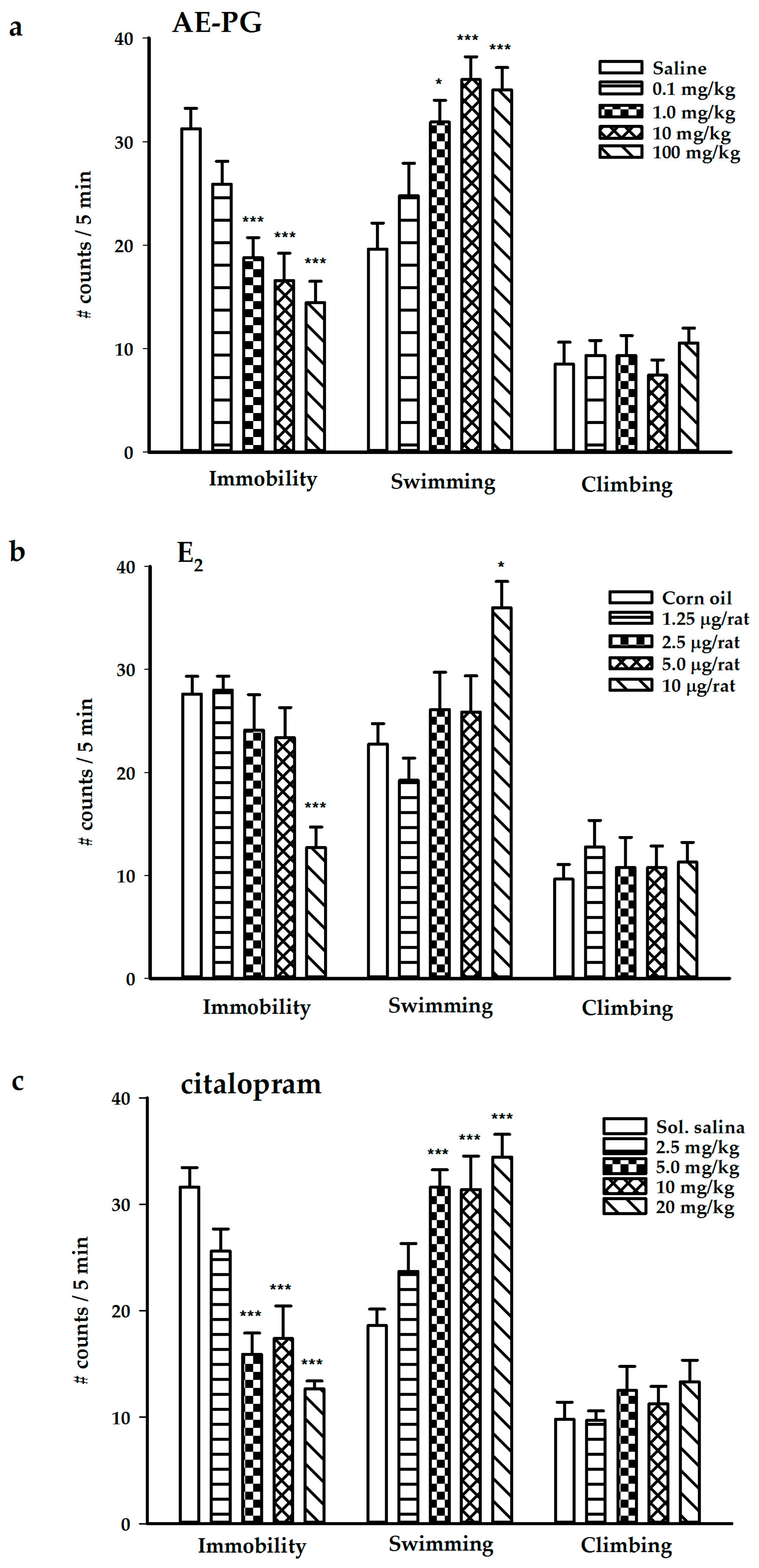
2.1. Antidepressant-Like Effect of the AE-PG, E2, and Citalopram
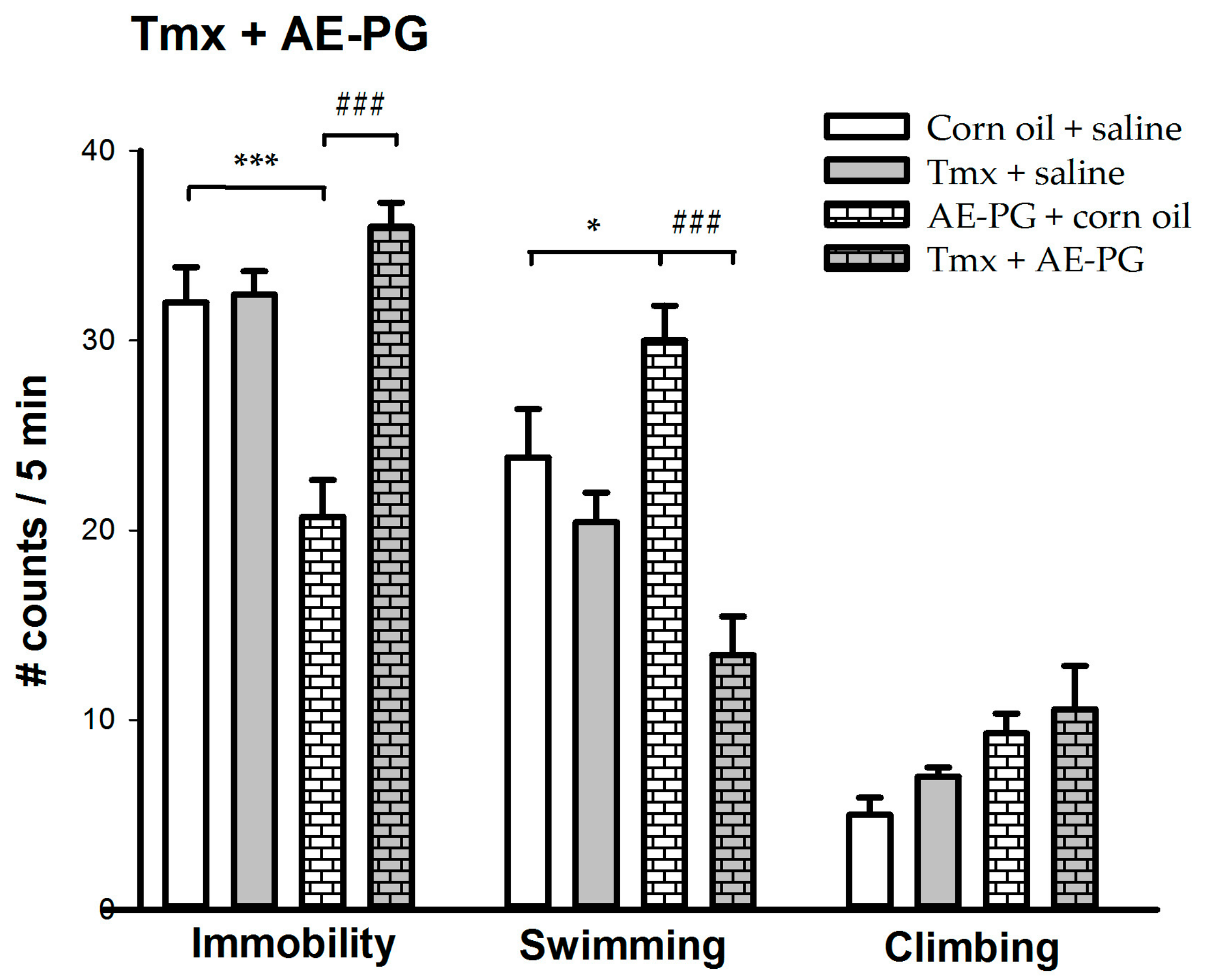
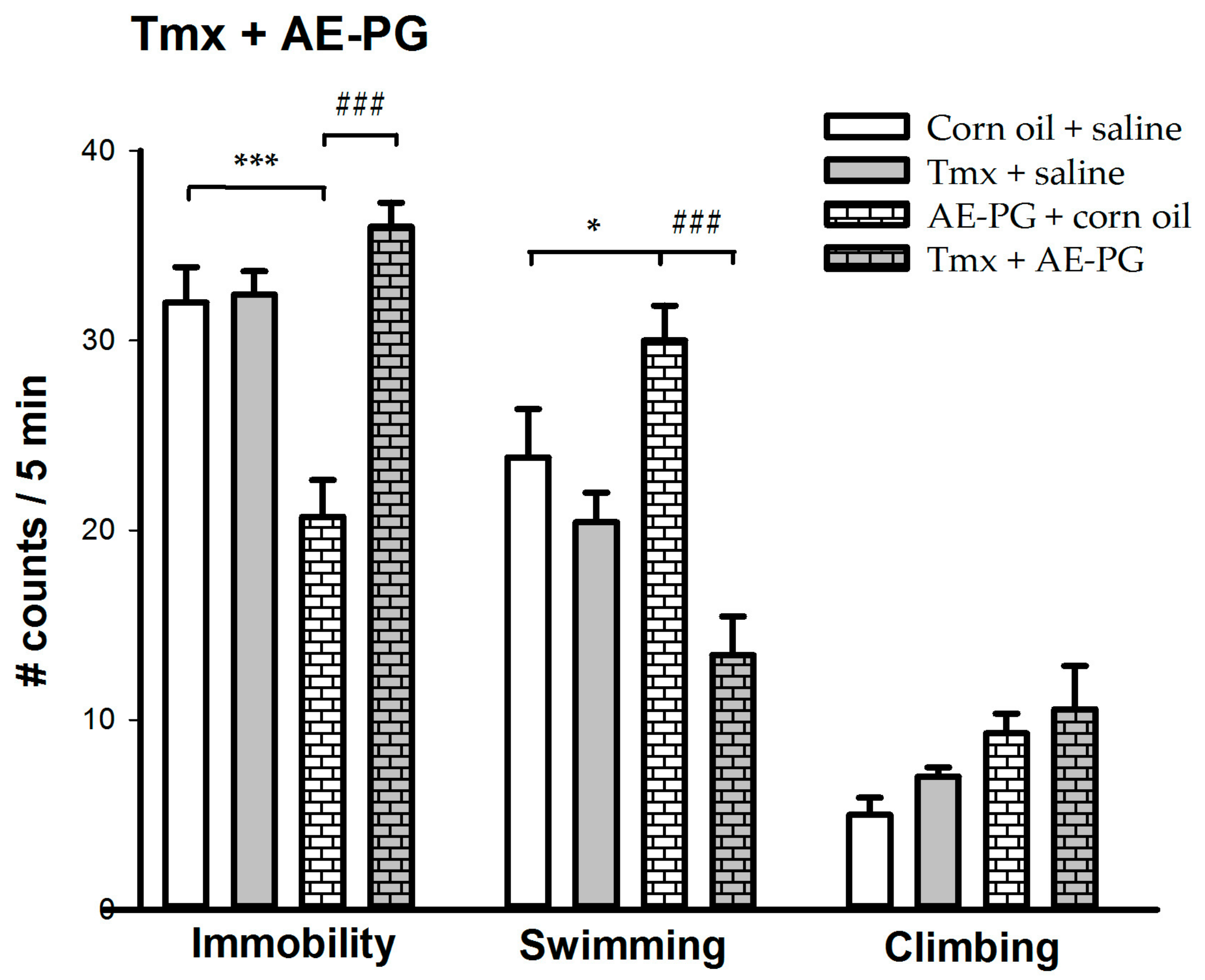
2.2. Participation of ER in the Antidepressant-Like Effect of AE-PG
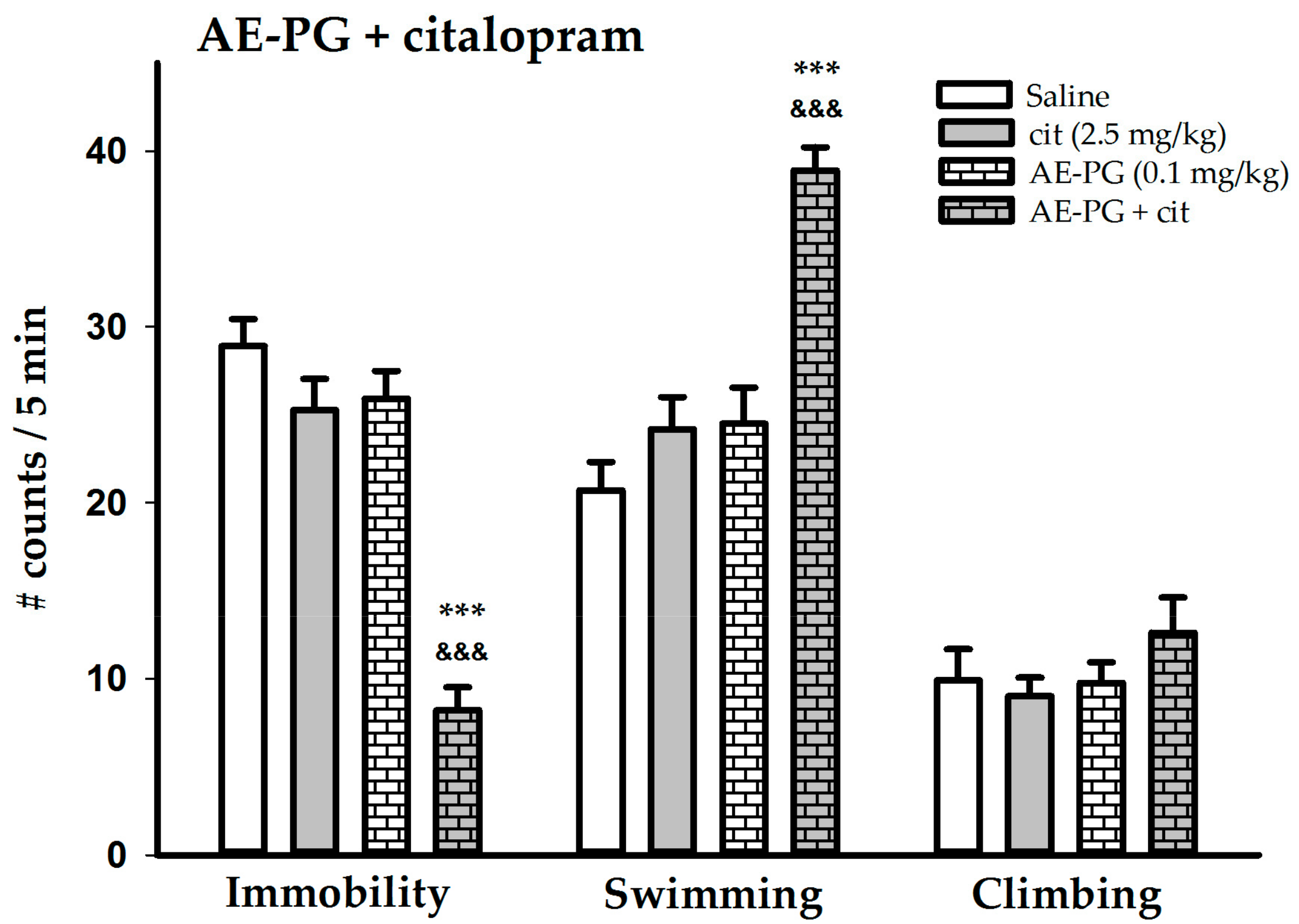
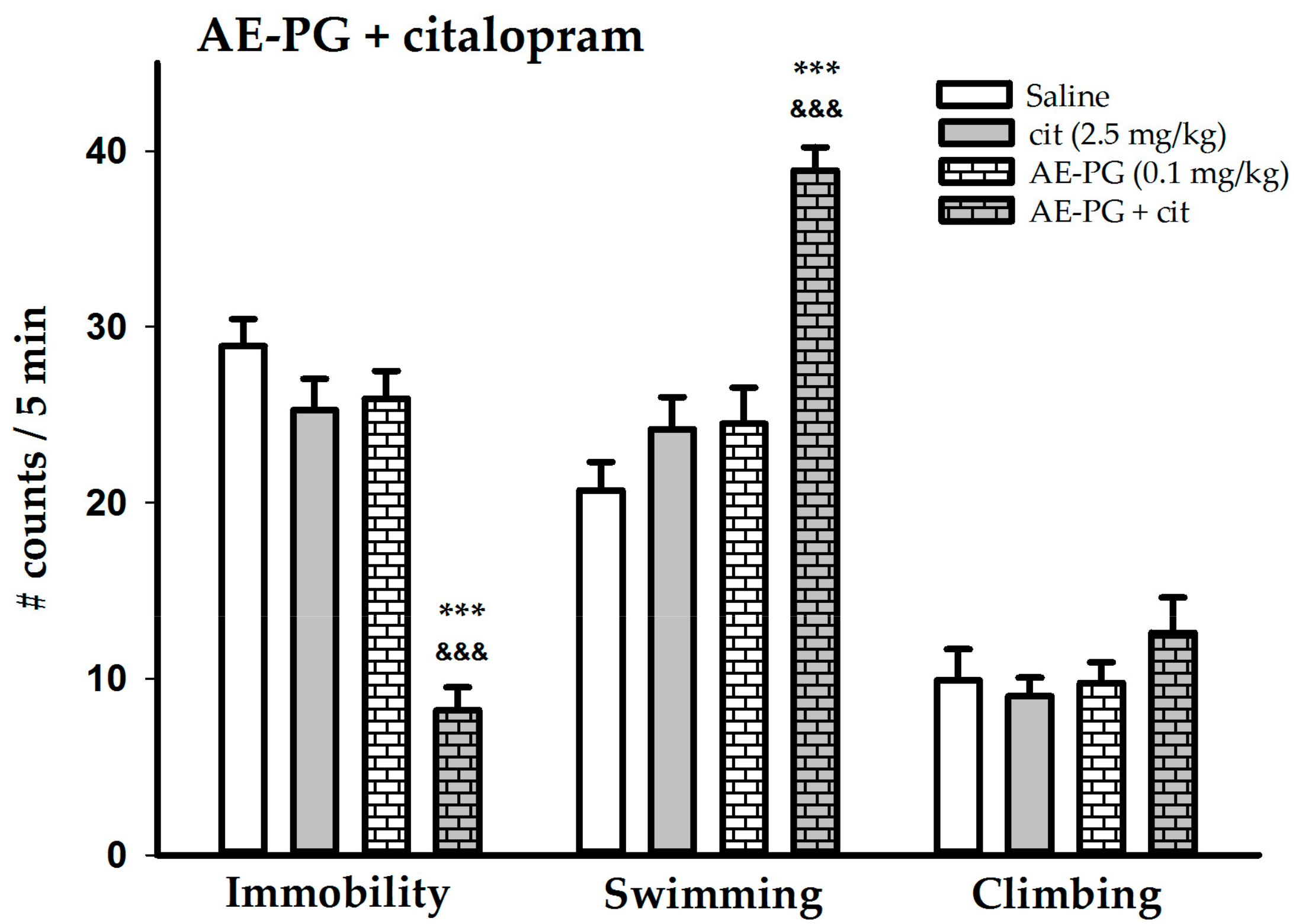
2.3. Effect of the Simultaneous Administration of Suboptimal Doses of AE-PG plus Citalopram
2.4. Effects of Treatments on the General Activity Test
3. Discussion
3.1. Antidepressant-Like Action Induced by Chronic Treatment of AE-PG
3.2. Combined Treatment of AE-PG plus Citalopram
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Ovariectomy
4.4. Forced Swimming Test (FST)
4.5. Behavioral Scoring
4.6. Activity Test
4.7. Statistics
4.8. Experimental Design
4.8.1. Experiment 1: Antidepressant-Like Effects of AE-PG, E2, and Citalopram
4.8.2. Experiment 2: Analysis of the Participation of ER in the Antidepressant-Like Effects of AE-PG
4.8.3. Experiment 3: Effect of the Simultaneous Administration of Suboptimal Doses of AE-PG Plus Citalopram
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Viladomiu, M.; Hontecillas, R.; Lu, P.; Bassaganya-Riera, J. Preventive and prophylactic mechanisms of action of pomegranate bioactive constituents. Evid. Based Complement. Alternat. Med. 2013, 2013, 789764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.H.; Fu, Q.J.; Zhang, Y.H. Composition of anthocyanins in pomegranate flowers and their antioxidant activity. Food Chem. 2011, 127, 1444–1449. [Google Scholar] [CrossRef]
- Al-Muammar, M.N.; Khan, F. Obesity: The preventive role of the pomegranate (Punica granatum). Nutrition 2012, 28, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Trujano, M.E.; Pellicer, F.; Mena, P.; Moreno, D.A.; Garcia-Viguera, C. Antinociceptive and anti-inflammatory activities of a pomegranate (Punica granatum L.) extract rich in ellagitannins. Int. J. Food Sci. Nutr. 2015, 66, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Ma, H.; Liu, W.; Niesen, D.B.; Shah, N.; Crews, R.; Rose, K.N.; Vattem, D.A.; Seeram, N.P. Pomegranate’s Neuroprotective Effects against Alzheimer’s Disease Are Mediated by Urolithins, Its Ellagitannin-Gut Microbial Derived Metabolites. ACS Chem. Neurosci. 2016, 7, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Viuda-Martos, M.; Fernandez-Lopez, J.; Perez-Alvarez, J.A. Pomegranate and its Many Functional Components as Related to Human Health: A Review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 635–654. [Google Scholar] [CrossRef]
- Choi, D.W.; Kim, J.Y.; Choi, S.H.; Jung, H.S.; Kim, H.J.; Cho, S.Y.; Kang, C.S.; Chang, S.Y. Identification of steroid hormones in pomegranate (Punica granatum) using HPLC and GC-mass spectrometry. Food Chem. 2006, 96, 562–571. [Google Scholar] [CrossRef]
- Mori-Okamoto, J.; Otawara-Hamamoto, Y.; Yamato, H.; Yoshimura, H. Pomegranate extract improves a depressive state and bone properties in menopausal syndrome model ovariectomized mice. J. Ethnopharmacol. 2004, 92, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Van Elswijk, D.A.; Schobel, U.P.; Lansky, E.P.; Irth, H.; van der Greef, J. Rapid dereplication of estrogenic compounds in pomegranate (Punica granatum) using on-line biochemical detection coupled to mass spectrometry. Phytochemistry 2004, 65, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Ellagitannins, ellagic acid and their derived metabolites: A review about source, metabolism, functions and health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Larrosa, M.; Gonzalez-Sarrias, A.; Garcia-Conesa, M.T.; Tomas-Barberan, F.A.; Espin, J.C. Urolithins, ellagic acid-derived metabolites produced by human colonic microflora, exhibit estrogenic and antiestrogenic activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Espin, J.C.; Larrosa, M.; Garcia-Conesa, M.T.; Tomas-Barberan, F. Biological significance of urolithins, the gut microbial ellagic acid-derived metabolites: The evidence so far. Evid. Based Complement. Alternat. Med. 2013, 2013, 270418. [Google Scholar] [CrossRef] [PubMed]
- Heftmann, E.; Ko, S.T.; Bennett, R.D. Identification of Estrone in Pomegranate Seeds. Phytochemistry 1966, 5, 1337–1339. [Google Scholar] [CrossRef]
- Handley, A.P.; Williams, M. The efficacy and tolerability of SSRI/SNRIs in the treatment of vasomotor symptoms in menopausal women: A systematic review. J. Am. Assoc. Nurse Pract. 2015, 27, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Roberts, H.; Hickey, M. Managing the menopause: An update. Maturitas 2016, 86, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Lethaby, A.; Marjoribanks, J.; Kronenberg, F.; Roberts, H.; Eden, J.; Brown, J. Phytoestrogens for menopausal vasomotor symptoms. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- Shuid, A.N.; Mohamed, I.N. Pomegranate Use to Attenuate Bone Loss in Major Musculoskeletal Diseases: An Evidence-Based Review. Curr. Drug Targets 2013, 14, 1565–1578. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, L.; Rakus, J.; Bauer, C.; Gerner, C.; Ullmann, R.; Wimmer, H.; Huber, J. Pomegranate seed oil in women with menopausal symptoms: A prospective randomized, placebo-controlled, double-blinded trial. Menopause 2012, 19, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Spilmont, M.; Leotoing, L.; Davicco, M.J.; Lebecque, P.; Mercier, S.; Miot-Noirault, E.; Pilet, P.; Rios, L.; Wittrant, Y.; Coxam, V. Pomegranate seed oil prevents bone loss in a mice model of osteoporosis, through osteoblastic stimulation, osteoclastic inhibition and decreased inflammatory status. J. Nutr. Biochem. 2013, 24, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Naveen, S.; Siddalingaswamy, M.; Singsit, D.; Khanum, F. Anti-depressive effect of polyphenols and omega-3 fatty acid from pomegranate peel and flax seed in mice exposed to chronic mild stress. Psychiatry Clin. Neurosci. 2013, 67, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Rodriguez, G.; Vega-Rivera, N.M.; Oikawa-Sala, J.; Gomez-Sanchez, A.; Ortiz-Lopez, L.; Estrada-Camarena, E. Melatonin synergizes with citalopram to induce antidepressant-like behavior and to promote hippocampal neurogenesis in adult mice. J. Pineal Res. 2014, 56, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Camarena, E.; Fernandez-Guasti, A.; Lopez-Rubalcava, C. Antidepressant-like effect of different estrogenic compounds in the forced swimming test. Neuropsychopharmacology 2003, 28, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Camarena, E.; Fernandez-Guasti, A.; Lopez-Rubalcava, C. Interaction between estrogens and antidepressants in the forced swimming test in rats. Psychopharmacology 2004, 173, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Valentino, R.J.; Lucki, I. Assessing substrates underlying the behavioral effects of antidepressants using the modified rat forced swimming test. Neurosci. Biobehav. Rev. 2005, 29, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Rygula, R.; Abumaria, N.; Flugge, G.; Fuchs, E.; Ruther, E.; Havemann-Reinecke, U. Anhedonia and motivational deficits in rats: Impact of chronic social stress. Behav. Brain Res. 2005, 162, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, O.V.; Kanekar, S.; D’Anci, K.E.; Renshaw, P.F. Factors influencing behavior in the forced swim test. Physiol. Behav. 2013, 118, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Reneric, J.P.; Lucki, I. Antidepressant behavioral effects by dual inhibition of monoamine reuptake in the rat forced swimming test. Psychopharmacology 1998, 136, 190–197. [Google Scholar] [PubMed]
- Detke, M.J.; Lucki, I. Detection of serotonergic and noradrenergic antidepressants in the rat forced swimming test: The effects of water depth. Behav. Brain Res. 1996, 73, 43–46. [Google Scholar] [CrossRef]
- Chaki, S.; Yoshikawa, R.; Hirota, S.; Shimazaki, T.; Maeda, M.; Kawashima, N.; Yoshimizu, T.; Yasuhara, A.; Sakagami, K.; Okuyama, S.; et al. MGS0039: A potent and selective group II metabotropic glutamate receptor antagonist with antidepressant-like activity. Neuropharmacology 2004, 46, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Reneric, J.P.; Bouvard, M.; Stinus, L. In the rat forced swimming test, NA-system mediated interactions may prevent the 5-HT properties of some subacute antidepressant treatments being expressed. Eur. Neuropsychopharm. 2002, 12, 159–171. [Google Scholar] [CrossRef]
- Estrada-Camarena, E.; Fernandez-Guasti, A.; Lopez-Rubalcava, C. Participation of the 5-HT1A receptor in the antidepressant-like effect of estrogens in the forced swimming test. Neuropsychopharmacology 2006, 31, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Mondragon-Jacobo, C.; Hernandez-Herrera, G.; Guzman-Maldonado, S.H. Agronomical, Physicochemical, and Functional Characterization of Mexican Pomegranates as Compared to Wonderful Pomegranate. Acta Hortic. 2015, 1089, 299–306. [Google Scholar] [CrossRef]
- Girish, C.; Raj, V.; Arya, J.; Balakrishnan, S. Evidence for the involvement of the monoaminergic system, but not the opioid system in the antidepressant-like activity of ellagic acid in mice. Eur. J. Pharmacol. 2012, 682, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Mena, P.; Cozzini, P.; Brighenti, F.; del Rio, D. Modelling the possible bioactivity of ellagitannin-derived metabolites. In silico tools to evaluate their potential xenoestrogenic behavior. Food Funct. 2013, 4, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Walf, A.A.; Rhodes, M.E.; Frye, C.A. Antidepressant effects of ERβ-selective estrogen receptor modulators in the forced swim test. Pharmacol. Biochem. Behav. 2004, 78, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.M.; Ni, G.L.; Shao, A.M.; Cui, R. Genistein alleviates anxiety-like behaviors in post-traumatic stress disorder model through enhancing serotonergic transmission in the amygdala. Psychiatry Res. 2017, 255, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Ma, L.; Wang, Y.G.; Ye, F.; Wang, C.; Zhou, W.H.; Zhao, X. Genistein, a dietary soy isoflavone, exerts antidepressant-like effects in mice: Involvement of serotonergic system. Neurochem. Int. 2017, 108, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Creech, R.D.; Li, Q.; Carrasco, G.A.; Van de Kar, L.D.; Muma, N.A. Estradiol induces partial desensitization of serotonin 1A receptor signaling in the paraventricular nucleus of the hypothalamus and alters expression and interaction of RGSZ1 and G alpha z. Neuropharmacology 2012, 62, 2040–2049. [Google Scholar] [CrossRef] [PubMed]
- López-Rubalcava, C.; Vega-Rivera, N.M.; Páez-Martínez, N.; Estrada-Camarena, E. Participation of the Monoaminergic System in the Antidepressant-Like Actions of Estrogens: A Review in Preclinical Studies. In Effects of Antidepressants; Ru-Bans, L., Ed.; InTech: London, UK, 2012. [Google Scholar]
- Lemini, C.; Cruz-Lopez, B.; Martinez-Mota, L. Participation of estrogen receptors in the antidepressant-like effect of prolame on the forced swimming test. Pharmacol. Biochem. Behav. 2013, 103, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Vega Rivera, N.M.; Tenorio, A.G.; Fernandez-Guasti, A.; Estrada-Camarena, E. The Post-Ovariectomy Interval Affects the Antidepressant-Like Action of Citalopram Combined with Ethynyl-Estradiol in the Forced Swim Test in Middle Aged Rats. Pharmaceuticals 2016, 9. [Google Scholar] [CrossRef] [PubMed]
- Flores-Serrano, A.G.; Vila-Luna, M.L.; Alvarez-Cervera, F.J.; Heredia-Lopez, F.J.; Gongora-Alfaro, J.L.; Pineda, J.C. Clinical doses of citalopram or reboxetine differentially modulate passive and active behaviors of female Wistar rats with high or low immobility time in the forced swimming test. Pharmacol. Biochem. Behav. 2013, 110, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Kusmider, M.; Solich, J.; Palach, P.; Dziedzicka-Wasylewska, M. Effect of citalopram in the modified forced swim test in rats. Pharmacol. Rep. 2007, 59, 785–788. [Google Scholar] [PubMed]
- Brosen, K.; Naranjo, C.A. Review of pharmacokinetic and pharmacodynamic interaction studies with citalopram. Eur. Neuropsychopharm. 2001, 11, 275–283. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Munoz, C.; Vaillant, F. Metabolic Fate of Ellagitannins: Implications for Health, and Research Perspectives for Innovative Functional Foods. Crit. Rev. Food Sci. 2014, 54, 1584–1598. [Google Scholar] [CrossRef] [PubMed]
- Basheer, L.; Kerem, Z. Interactions between CYP3A4 and Dietary Polyphenols. Oxid. Med. Cell. Longev. 2015, 2015, 854015. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.; Putt, D.; Kresty, L.; Stoner, G.D.; Fromm, D.; Hollenberg, P.F. The effects of dietary ellagic acid on rat hepatic and esophageal mucosal cytochromes P450 and phase II enzymes. Carcinogenesis 1996, 17, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Recamier-Carballo, S.; Estrada-Camarena, E.; Reyes, R.; Fernandez-Guasti, A. Synergistic effect of estradiol and fluoxetine in young adult and middle-aged female rats in two models of experimental depression. Behav. Brain Res. 2012, 233, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Tam, L.W.; Parry, B.L. Does estrogen enhance the antidepressant effects of fluoxetine? J. Affect. Disord. 2003, 77, 87–92. [Google Scholar]
- Soares, C.N.; Poitras, J.R.; Prouty, J.; Alexander, A.B.; Shifren, J.L.; Cohen, L.S. Efficacy of citalopram as a monotherapy or as an adjunctive treatment to estrogen therapy for perimenopausal and postmenopausal women with depression and vasomotor symptoms. J. Clin. Psychiat. 2003, 64, 473–479. [Google Scholar] [CrossRef]
- Contreras, C.M.; Rodríguez-Landa, J.F.; Gutiérrez-García, A.G. The lowest effective dose of fluoxetine in the forced swim test significantly affects the firing rate of lateral septal nucleus neurons in the rat. J. Psychopharmacol. 2001, 15, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Mostalac-Preciado, C.R.; de Gortari, P.; López-Rubalcava, C. Antidepressant-like effects of mineralocorticoid but not glucocorticoid antagonists in th elateral septum: Interactions with the serotonergic system. Behav. Brain Res. 2011, 223, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rivera, N.M.; Fernández-Guasti, A.; Ramírez-Rodríguez, G.; Estrada-Camarena, E. Acute stress further decreases the effect of ovariectomy on immobility behavior and hippocampal cell survival in rats. Psychoneuroendocrinology 2013, 38, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Detke, M.J.; Rickels, M.; Lucki, I. Active Behaviors in the Rat Forced Swimming Test Differentially Produced by Serotonergic and Noradrenergic Antidepressants. Psychopharmacology 1995, 121, 66–72. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dose | n | Mean ± SEM | One Way ANOVA (F) | p |
|---|---|---|---|---|---|
| AE-PG | Saline | 8 | 54.667 ± 5.632 | F(4,37) = 0.869 | 0.491 |
| 0.1 mg/kg | 9 | 53.667 ± 5.370 | |||
| 1 mg/kg | 9 | 59.778 ± 7.579 | |||
| 10 mg/kg | 7 | 60.556 ± 3.775 | |||
| 100 mg/kg | 9 | 67.333 ± 6.298 | |||
| 17β-estradiol | Corn oil | 8 | 79.375 ± 1.711 | F(4,34) = 0.628 | 0.646 |
| 1.25 μg/rat | 8 | 77.750 ± 3.299 | |||
| 2.5 μg/rat | 8 | 74.375 ± 3.615 | |||
| 5 μg/rat | 8 | 75.125 ± 2.702 | |||
| 10 μg/rat | 7 | 79.714 ± 3.656 | |||
| Citalopram | Saline | 10 | 77.500 ± 3.390 | F(4,42) = 8.197 | - |
| 2.5 mg/kg | 10 | 82.400 ± 2.864 | 0.618 | ||
| 5 mg/kg | 10 | 60.500 ± 2.051 | <0.05 | ||
| 10 mg/kg | 8 | 70.625 ± 2.179 | 0.376 | ||
| 20 mg/kg | 9 | 79.333 ± 4.444 | 0.982 |
| Experiment | Treatment | n | Mean ± SEM | One Way ANOVA (F) | p |
|---|---|---|---|---|---|
| Tmx + AE-PG | Corn oil + Saline | 6 | 74.000 ± 2.944 | F(3,23) = 0.644 | 0.594 |
| Corn oil + AE-PG | 7 | 75.000 ± 0.093 | |||
| Tmx + Saline | 7 | 78.143 ± 1.438 | |||
| Tmx + AE-PG | 7 | 74.429 ± 2.608 | |||
| AE-PG + Cit | Saline | 10 | 62.200 ± 6.277 | F(3,36) = 4.797 | 0.07 |
| Saline + AE-PG | 10 | 57.400 ± 6.848 | |||
| Saline + citalopram | 10 | 79.700 ± 2.745 | |||
| AE-PG + citalopram | 10 | 78.200 ± 3.408 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdés-Sustaita, B.; López-Rubalcava, C.; González-Trujano, M.E.; García-Viguera, C.; Estrada-Camarena, E. Aqueous Extract of Pomegranate Alone or in Combination with Citalopram Produces Antidepressant-Like Effects in an Animal Model of Menopause: Participation of Estrogen Receptors. Int. J. Mol. Sci. 2017, 18, 2643. https://doi.org/10.3390/ijms18122643
Valdés-Sustaita B, López-Rubalcava C, González-Trujano ME, García-Viguera C, Estrada-Camarena E. Aqueous Extract of Pomegranate Alone or in Combination with Citalopram Produces Antidepressant-Like Effects in an Animal Model of Menopause: Participation of Estrogen Receptors. International Journal of Molecular Sciences. 2017; 18(12):2643. https://doi.org/10.3390/ijms18122643
Chicago/Turabian StyleValdés-Sustaita, Brenda, Carolina López-Rubalcava, María Eva González-Trujano, Cristina García-Viguera, and Erika Estrada-Camarena. 2017. "Aqueous Extract of Pomegranate Alone or in Combination with Citalopram Produces Antidepressant-Like Effects in an Animal Model of Menopause: Participation of Estrogen Receptors" International Journal of Molecular Sciences 18, no. 12: 2643. https://doi.org/10.3390/ijms18122643
APA StyleValdés-Sustaita, B., López-Rubalcava, C., González-Trujano, M. E., García-Viguera, C., & Estrada-Camarena, E. (2017). Aqueous Extract of Pomegranate Alone or in Combination with Citalopram Produces Antidepressant-Like Effects in an Animal Model of Menopause: Participation of Estrogen Receptors. International Journal of Molecular Sciences, 18(12), 2643. https://doi.org/10.3390/ijms18122643