View Point: Semaphorin-3E: An Emerging Modulator of Natural Killer Cell Functions?

{kind=link}
{kind=link}
Abstract
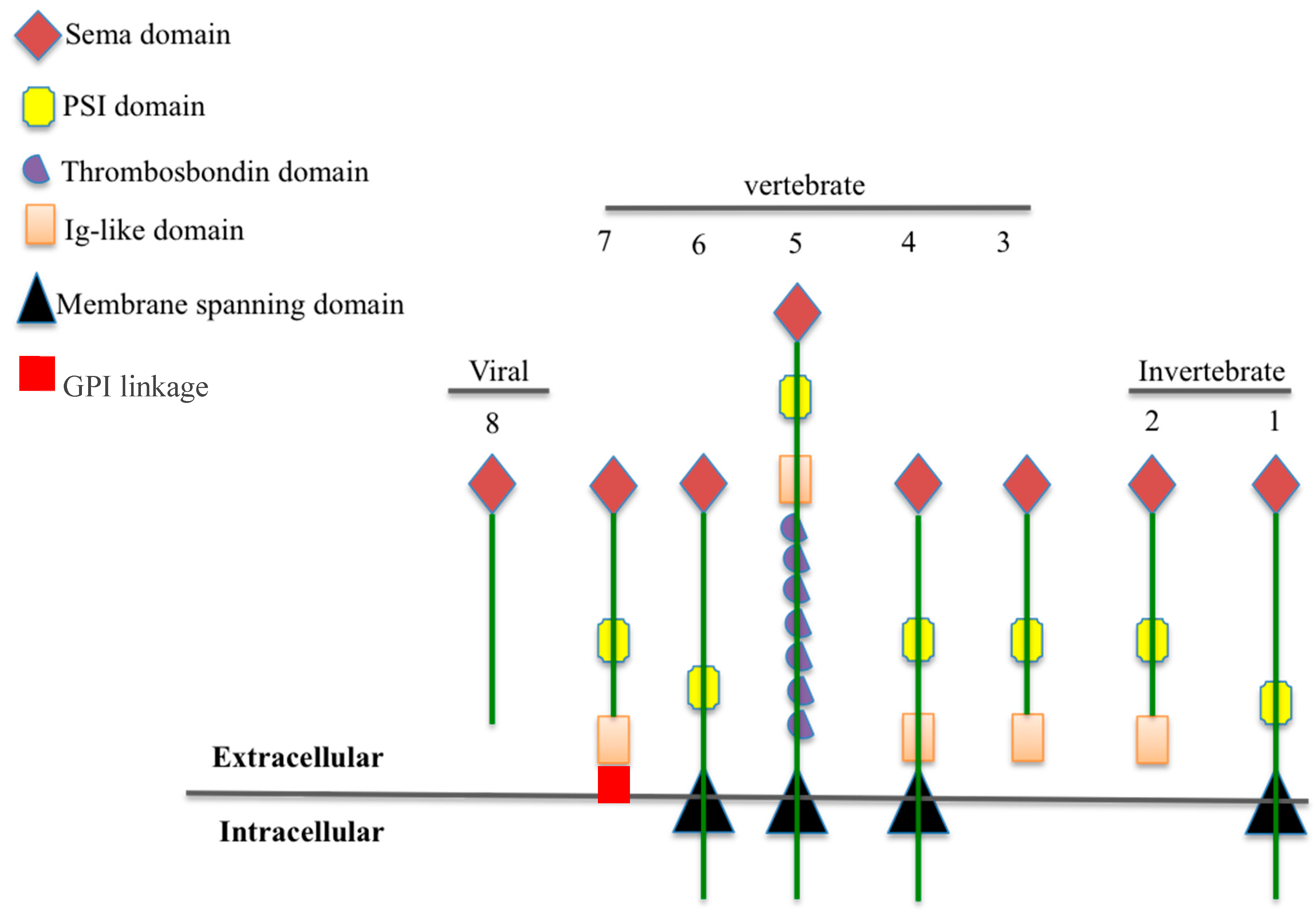
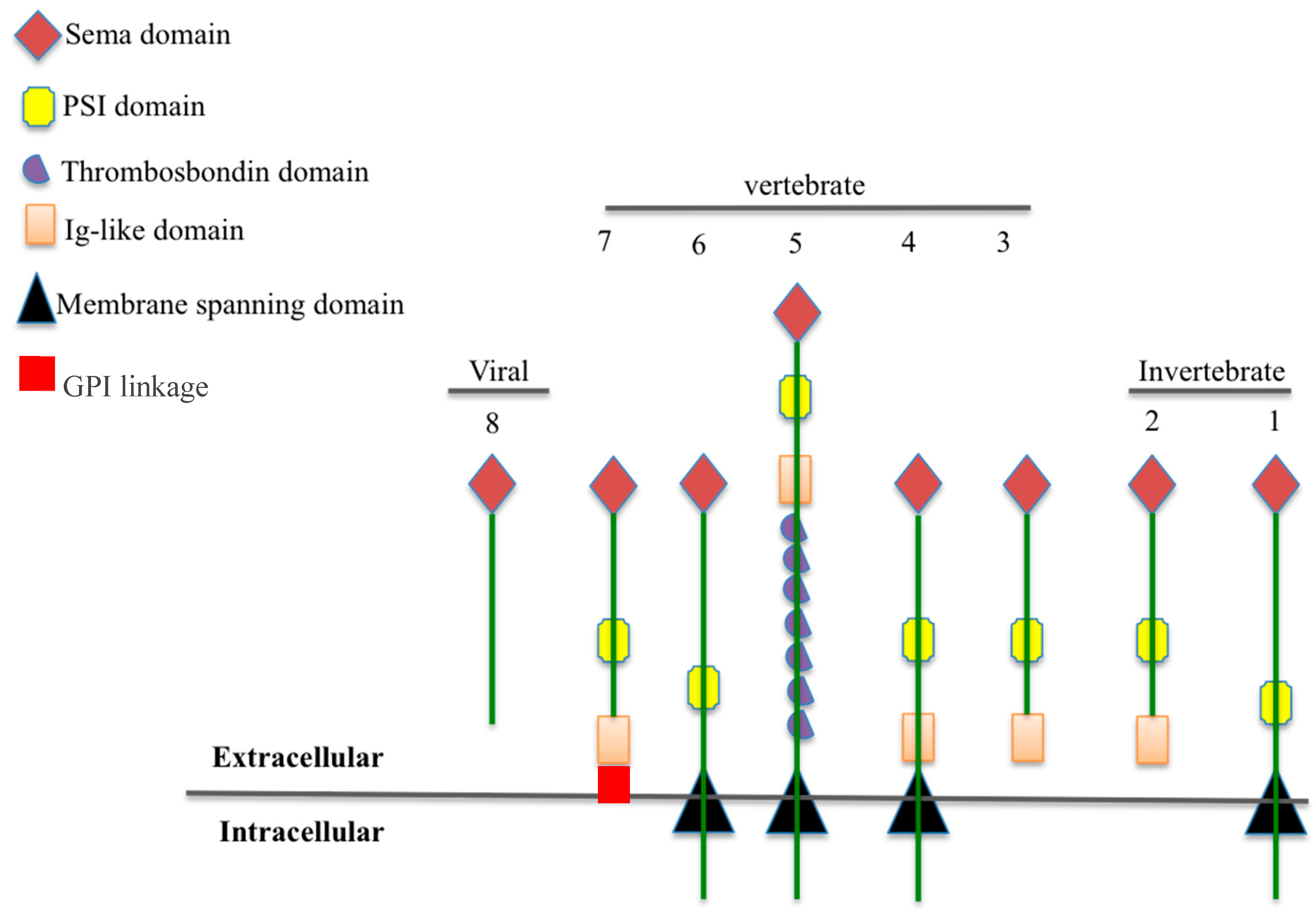
:1. The Semaphorin Family: Classification and Structure
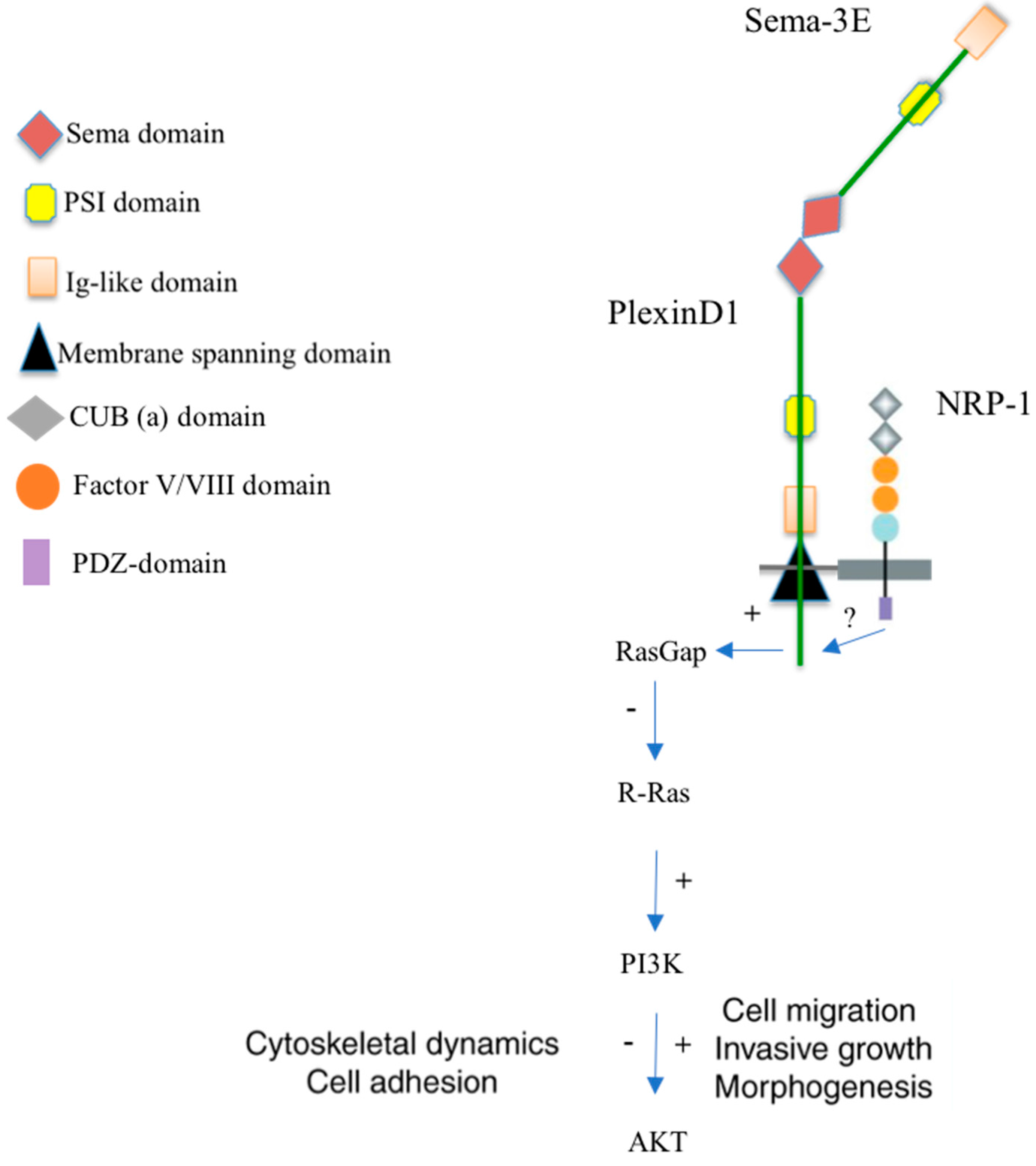
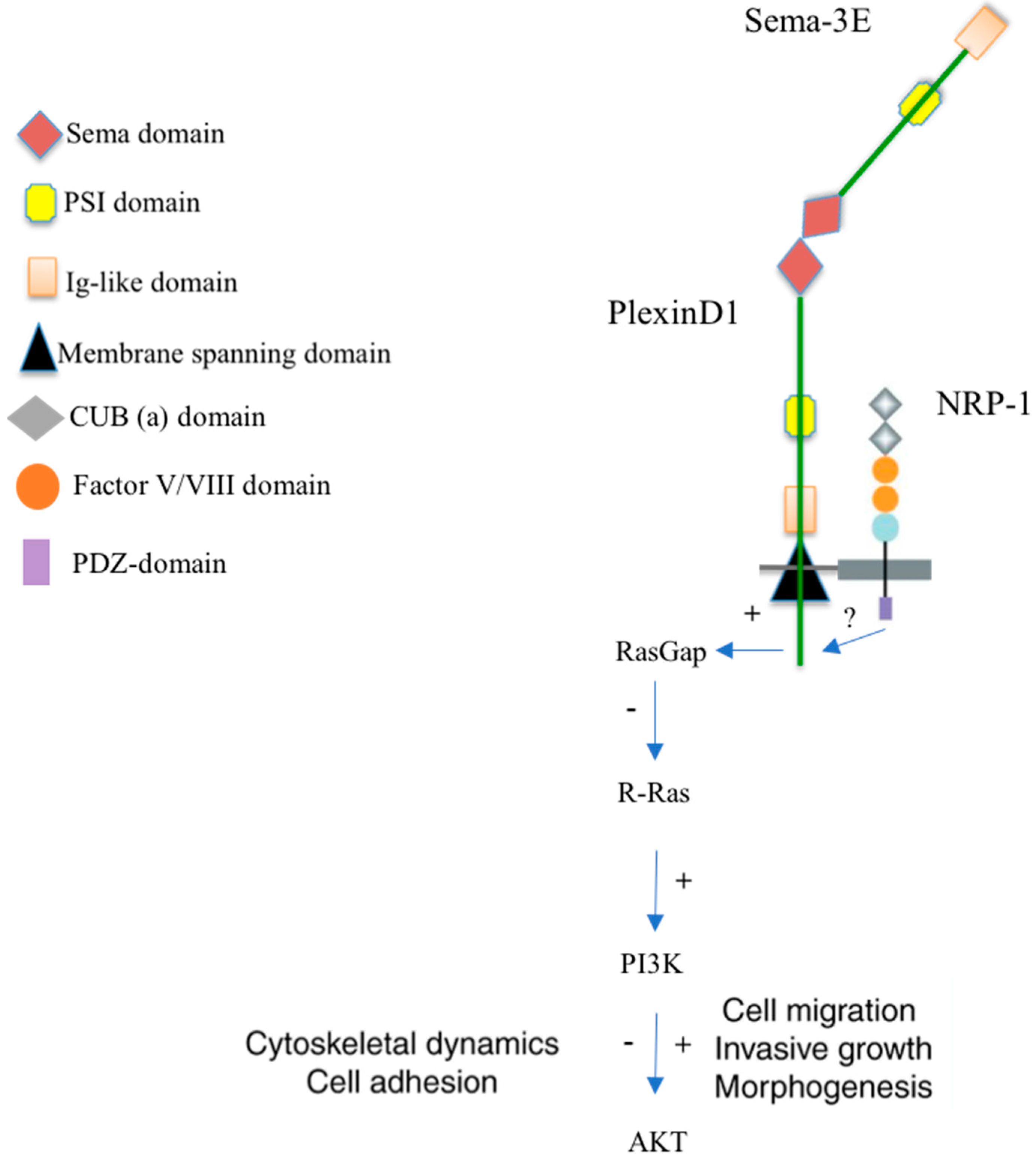
2. Receptors and Signaling of the Semaphorin-3E
3. Semaphorin-3E in the Nervous System
4. Semaphorin-3E in Cardiovascular Development
5. Semaphorin-3E and Cancers
6. Semaphorin-3E in an Allergic Asthma Model
7. The Prospective Role of Semaphorin-3E in Regulating Natural Killer (NK) Cell Functions
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Koncina, E.; Roth, L.; Gonthier, B.; Bagnard, D. Role of semaphorins during axon growth and guidance. In Axon Growth and Guidance; Bagnard, D., Ed.; Springer: New York, NY, USA, 2007; pp. 50–64. [Google Scholar]
- Wong, J.; Yu, W.; O’Connor, T.P. Transmembrane grasshopper Semaphorin I promotes axon outgrowth in vivo. Development 1997, 124, 3597–3607. [Google Scholar] [PubMed]
- Roney, K.; Holl, E.; Ting, J. Immune plexins and semaphorins: Old proteins, new immune functions. Protein Cell 2013, 4, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, U.; Terman, J.R. The semaphorins. Genome Biol. 2006, 7, 211. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A. Receptor complexes for each of the Class 3 Semaphorins. Front. Cell. Neurosci. 2012, 6. [Google Scholar] [CrossRef]
- Koppel, A.M.; Feiner, L.; Kobayashi, H.; Raper, J.A. A 70 Amino Acid Region within the Semaphorin Domain Activates Specific Cellular Response of Semaphorin Family Members. Neuron 1997, 19, 531–537. [Google Scholar] [CrossRef]
- Gu, C.; Yoshida, Y.; Livet, J.; Reimert, D.V.; Mann, F.; Merte, J.; Henderson, C.E.; Jessell, T.M.; Kolodkin, A.L.; Ginty, D.D. Semaphorin 3E and Plexin-D1 Control Vascular Pattern Independently of Neuropilins. Science 2005, 307, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Siebold, C.; Jones, E.Y. Structural insights into semaphorins and their receptors. Semin. Cell Dev. Biol. 2013, 24, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.P.; Shanks, K.; Lipsky, M.M.; DeTolla, L.J.; Keegan, A.D.; Chapoval, S.P. Expression of neuroimmune semaphorins 4A and 4D and their receptors in the lung is enhanced by allergen and vascular endothelial growth factor. BMC Immunol. 2011, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Nkyimbeng-Takwi, E.; Chapoval, S.P. Biology and function of neuroimmune semaphorins 4A and 4D. Immunol. Res. 2011, 50, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Lepelletier, Y.; Moura, I.C.; Hadj-Slimane, R.; Renand, A.; Fiorentino, S.; Baude, C.; Shirvan, A.; Barzilai, A.; Hermine, O. Immunosuppressive role of semaphorin-3A on T cell proliferation is mediated by inhibition of actin cytoskeleton reorganization. Eur. J. Immunol. 2006, 36, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Worzfeld, T.; Offermanns, S. Semaphorins and plexins as therapeutic targets. Nat. Rev. Drug Discov. 2014, 13, 603. [Google Scholar] [CrossRef] [PubMed]
- Kumanogoh, A.; Kikutani, H. Immunological functions of the neuropilins and plexins as receptors for semaphorins. Nat. Rev. Immunol. 2013, 13, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, M.V.; Hong, S.; Gu, C. Guidance from above: Common cues direct distinct signaling outcomes in vascular and neural patterning. Trends Cell Biol. 2009, 19, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.; Rougon, G. Mechanisms of axon guidance: Membrane dynamics and axonal transport in semaphorin signalling. J. Neurochem. 2007, 102, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, P.; Gemmill, R.M.; Drabkin, H.A. The emerging role of class-3 semaphorins and their neuropilin receptors in oncology. OncoTargets Ther. 2014, 7, 1663. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, F.; Goshima, Y. Structural and functional relation of neuropilins. In Neuropilin: From Nervous System to Vascular and Tumor Biology; Springer Science & Business Media: New York, NY, USA, 2002; pp. 55–69. [Google Scholar]
- Ito, D.; Kumanogoh, A. The role of Sema4A in angiogenesis, immune responses, carcinogenesis, and retinal systems. Cell Adhes. Migr. 2016, 10, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Taniguchi, M. Congenital diseases and semaphorin signaling: Overview to date of the evidence linking them. Congenit. Anom. 2015, 55, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-F.; Vander Kooi, C.W. Neuropilin Functions as an Essential Cell Surface Receptor. J. Biol. Chem. 2015, 290, 29120–29126. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chédotal, A.; He, Z.; Goodman, C.S.; Tessier-Lavigne, M. Neuropilin-2, a Novel Member of the Neuropilin Family, Is a High Affinity Receptor for the Semaphorins Sema E and Sema IV but Not Sema III. Neuron 1997, 19, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Nakamura, F.; Jin, Z.; Kalb, R.G.; Strittmatter, S.M. Semaphorins A and E act as antagonists of neuropilin-1 and agonists of neuropilin-2 receptors. Nat. Neurosci. 1998, 1, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Castellani, V.; Chédotal, A.; Schachner, M.; Faivre-Sarrailh, C.; Rougon, G. Analysis of the L1-Deficient Mouse Phenotype Reveals Cross-Talk between Sema3A and L1 Signaling Pathways in Axonal Guidance. Neuron 2000, 27, 237–249. [Google Scholar] [CrossRef]
- Gu, C.; Rodriguez, E.R.; Reimert, D.V.; Shu, T.; Fritzsch, B.; Richards, L.J.; Kolodkin, A.L.; Ginty, D.D. Neuropilin-1 Conveys Semaphorin and VEGF Signaling during Neural and Cardiovascular Development. Dev. Cell 2003, 5, 45–57. [Google Scholar] [CrossRef]
- Kolk, S.M.; Gunput, R.A.; Tran, T.S.; van den Heuvel, D.M.; Prasad, A.A.; Hellemons, A.J.; Adolfs, Y.; Ginty, D.D.; Kolodkin, A.L.; Burbach, J.P.; et al. Semaphorin 3F Is a Bifunctional Guidance Cue for Dopaminergic Axons and Controls Their Fasciculation, Channeling, Rostral Growth, and Intracortical Targeting. J. Neurosci. 2009, 29, 12542–12557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.H.; Lohrum, M.; Klostermann, A.; Betz, H.; Püschel, A.W. The chemorepulsive activity of secreted semaphorins is regulated by furin-dependent proteolytic processing. EMBO J. 1997, 16, 6077–6086. [Google Scholar] [CrossRef] [PubMed]
- Klostermann, A.; Lohrum, M.; Adams, R.H.; Püschel, A.W. The chemorepulsive activity of the axonal guidance signal semaphorin D requires dimerization. J. Biol. Chem. 1998, 273, 7326–7331. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, H.; Kumanogoh, A. Diverse roles for semaphorin-plexin signaling in the immune system. Trends Immunol. 2012, 33, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Capparuccia, L.; Tamagnone, L. Semaphorin signaling in cancer cells and in cells of the tumor microenvironment—Two sides of a coin. J. Cell Sci. 2009, 122, 1723–1736. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.S.; Kolodkin, A.L.; Bharadwaj, R. Semaphorin regulation of cellular morphology. Annu. Rev. Cell Dev. Biol. 2007, 23, 263–292. [Google Scholar] [CrossRef] [PubMed]
- Vanderhaeghen, P.; Cheng, H.-J. Guidance molecules in axon pruning and cell death. Cold Spring Harb. Perspect. Biol. 2010, 2, a001859. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, D.H.; Khadem, D.F.; Gounni, P.A.S. Semaphorins and Their Roles in Airway Biology: Potential as Therapeutic Targets. Am. J. Respir. Cell Mol. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Klagsbrun, M.; Shimizu, A. Semaphorin 3E, an exception to the rule. J. Clin. Investig. 2010, 120, 2658–2660. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.-J.; Gu, C. The Role and Mechanism-of-Action of Sema3E and Plexin-D1 in Vascular and Neural Development. Semin. Cell Dev. Biol. 2013, 24, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Tamagnone, L.; Artigiani, S.; Chen, H.; He, Z.; Ming, G.-L.; Song, H.-J.; Chedotal, A.; Winberg, M.L.; Goodman, C.S.; Poo, M.-M.; et al. Plexins Are a Large Family of Receptors for Transmembrane, Secreted, and GPI-Anchored Semaphorins in Vertebrates. Cell 1999, 99, 71–80. [Google Scholar] [CrossRef]
- Gay, C.M.; Zygmunt, T.; Torres-Vázquez, J. Diverse functions for the semaphorin receptor PlexinD1 in development and disease. Dev. Biol. 2011, 349, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, K.; Oinuma, I.; Katoh, H.; Negishi, M. Different Requirement for Rnd GTPases of R-Ras GAP Activity of Plexin-C1 and Plexin-D1. J. Biol. Chem. 2009, 284, 6743–6751. [Google Scholar] [CrossRef] [PubMed]
- Mecollari, V.; Nieuwenhuis, B.; Verhaagen, J. A perspective on the role of class III semaphorin signaling in central nervous system trauma. Front. Cell. Neurosci. 2014, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Chauvet, S.; Cohen, S.; Yoshida, Y.; Fekrane, L.; Livet, J.; Gayet, O.; Segu, L.; Buhot, M.-C.; Jessell, T.M.; Henderson, C.E.; et al. Gating of Sema3E/PlexinD1 Signaling by Neuropilin-1 Switches Axonal Repulsion to Attraction during Brain Development. Neuron 2007, 56, 807–822. [Google Scholar] [CrossRef] [PubMed]
- Pecho-Vrieseling, E.; Sigrist, M.; Yoshida, Y.; Jessell, T.M.; Arber, S. Specificity of Sensory-Motor Connections Encoded by Sema3e-PlexinD1 Recognition. Nature 2009, 459, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y. Semaphorin Signaling in Vertebrate Neural Circuit Assembly. Front. Mol. Neurosci. 2012, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.B.; Oh, W.-J.; Sabatini, B.L.; Gu, C. Semaphorin 3E–Plexin-D1 signaling controls pathway-specific synapse formation in the striatum. Nat. Neurosci. 2012, 15, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Valdembri, D.; Regano, D.; Maione, F.; Giraudo, E.; Serini, G. Class 3 semaphorins in cardiovascular development. Cell Adhes. Migr. 2016, 10, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.A.; Aghajanian, H.; Singh, M.K. Semaphorin Signaling in Cardiovascular Development. Cell Metab. 2015, 21, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Meadows, S.M.; Ratliff, L.A.; Singh, M.K.; Epstein, J.A.; Cleaver, O. Resolution of defective dorsal aortae patterning in Sema3E deficient mice occurs via angiogenic remodeling. Dev. Dynam. 2013, 242, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, T.; Gay, C.M.; Blondelle, J.; Singh, M.K.; Flaherty, K.M.; Means, P.C.; Herwig, L.; Krudewig, A.; Belting, H.G.; Affolter, M.; et al. Semaphorin-PlexinD1 Signaling Limits Angiogenic Potential via the VEGF Decoy Receptor sFlt1. Dev. Cell 2011, 21, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Oh, W.-J.; Gaiano, N.; Yoshida, Y.; Gu, C. Semaphorin 3E–Plexin-D1 signaling regulates VEGF function in developmental angiogenesis via a feedback mechanism. Genes Dev. 2011, 25, 1399–1411. [Google Scholar] [CrossRef] [PubMed]
- Luchino, J.; Hocine, M.; Amoureux, M.-C.; Gibert, B.; Bernet, A.; Royet, A.; Treilleux, I.; Lécine, P.; Borg, J.-P.; Mehlen, P. Semaphorin 3E suppresses tumor cell death triggered by the plexin D1 dependence receptor in metastatic breast cancers. Cancer Cell 2013, 24, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Casazza, A.; Kigel, B.; Maione, F.; Capparuccia, L.; Kessler, O.; Giraudo, E.; Mazzone, M.; Neufeld, G.; Tamagnone, L. Tumour growth inhibition and anti-metastatic activity of a mutated furin-resistant Semaphorin 3E isoform. EMBO Mol. Med. 2012, 4, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Nariculam, J.; Munson, P.; Freeman, A.; Klocker, H.; Masters, J.; Williamson, M. A role for class 3 semaphorins in prostate cancer. Prostate 2011, 71, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Casazza, A.; Finisguerra, V.; Capparuccia, L.; Camperi, A.; Swiercz, J.M.; Rizzolio, S.; Rolny, C.; Christensen, C.; Bertotti, A.; Sarotto, I. Sema3E–Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice. J. Clin. Investig. 2010, 120, 2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Movassagh, H.; Shan, L.; Chakir, J.; McConville, J.F.; Halayko, A.J.; Koussih, L.; Gounni, A.S. Expression of semaphorin 3E is suppressed in severe asthma. J. Allergy Clin. Immunol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Shan, L.; Mohammed, A.; Halayko, A.J.; Gounni, A.S. Semaphorin 3E Deficiency Exacerbates Airway Inflammation, Hyperresponsiveness, and Remodeling in a Mouse Model of Allergic Asthma. J. Immunol. 2017, 198, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Shan, L.; Halayko, A.J.; Roth, M.; Tamm, M.; Chakir, J.; Gounni, A.S. Neuronal chemorepellent Semaphorin 3E inhibits human airway smooth muscle cell proliferation and migration. J. Allergy Clin. Immunol. 2014, 133, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Saati, A.; Nandagopal, S.; Mohammed, A.; Tatari, N.; Shan, L.; Duke-Cohan, J.S.; Fowke, K.R.; Lin, F.; Gounni, A.S. Chemorepellent Semaphorin 3E Negatively Regulates Neutrophil Migration In Vitro and In Vivo. J. Immunol. 2017, 198, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Shan, L.; Duke-Cohan, J.S.; Halayko, A.J.; Uzonna, J.E.; Gounni, A.S. Semaphorin 3E Alleviates Hallmarks of House Dust Mite-Induced Allergic Airway Disease. Am. J. Pathol. 2017, 187, 1566–1576. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.S.; Hasegawa, J. Natural killer cell biology: An update and future directions. J. Allergy Clin. Immunol. 2013, 132, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Waldhauer, I.; Steinle, A. NK cells and cancer immunosurveillance. Oncogene 2008, 27, 5932–5943. [Google Scholar] [CrossRef] [PubMed]
- Cella, M.; Miller, H.; Song, C. Beyond NK Cells: The Expanding Universe of Innate Lymphoid Cells. Front. Immunol. 2014, 5, 282. [Google Scholar] [CrossRef] [PubMed]
- Spits, H.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells—A proposal for uniform nomenclature. Nat. Rev. Immunol. 2013, 13, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Colonna, M. Innate lymphoid cells in homeostasis, infection, chronic inflammation and tumors of the gastrointestinal tract. Curr. Opin. Gastroenterol. 2013, 29, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, G.; Pack, M.; Thomas, D.; Paludan, C.; Schmid, D.; Strowig, T.; Bougras, G.; Muller, W.A.; Moretta, L.; Münz, C. Distinct roles of IL-12 and IL-15 in human natural killer cell activation by dendritic cells from secondary lymphoid organs. Proc. Natl. Acad. Sci. USA 2004, 101, 16606–16611. [Google Scholar] [CrossRef] [PubMed]
- Wehner, R.; Dietze, K.; Bachmann, M.; Schmitz, M. The Bidirectional Crosstalk between Human Dendritic Cells and Natural Killer Cells. J. Innate Immun. 2011, 3, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, J.E.; Bonehill, A.; Thielemans, K.; Wan, Y. Engineering Dendritic Cells to Enhance Cancer Immunotherapy. Mol. Ther. 2011, 19, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Ferlazzo, G.; Morandi, B. Cross-Talks between Natural Killer Cells and Distinct Subsets of Dendritic Cells. Front. Immunol. 2014, 5, 159. [Google Scholar] [CrossRef] [PubMed]
- Laffont, S.; Seillet, C.; Ortaldo, J.; Coudert, J.D.; Guéry, J.-C. Natural killer cells recruited into lymph nodes inhibit alloreactive T-cell activation through perforin-mediated killing of donor allogeneic dendritic cells. Blood 2008, 112, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Morandi, B.; Mortara, L.; Chiossone, L.; Accolla, R.S.; Mingari, M.C.; Moretta, L.; Moretta, A.; Ferlazzo, G. Dendritic Cell Editing by Activated Natural Killer Cells Results in a More Protective Cancer-Specific Immune Response. PLoS ONE 2012, 7, e39170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, D.M.; Scalzo, A.A.; Yokoyama, W.M.; Smyth, M.J.; Degli-Esposti, M.A. Functional interactions between dendritic cells and NK cells during viral infection. Nat. Immunol. 2003, 4, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Karlhofer, F.M.; Ribaudo, R.K.; Yokoyama, W.M. MHC class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature 1992, 358, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Fuchs, A.; Colonna, M.; Caligiuri, M.A. NK cell and DC interactions. Trends Immunol. 2004, 25, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Cools, N.; Ponsaerts, P.; Van Tendeloo, V.F.I.; Berneman, Z.N. Balancing between immunity and tolerance: An interplay between dendritic cells, regulatory T cells, and effector T cells. J. Leukoc. Biol. 2007, 82, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Marcenaro, E.; Carlomagno, S.; Pesce, S.; Moretta, A.; Sivori, S. NK/DC Crosstalk in Anti-viral Response. In Current Topics in Innate Immunity II; Lambris, J.D., Hajishengallis, G., Eds.; Springer: New York, NY, USA, 2012; pp. 295–308. [Google Scholar]
- Nguyen-Pham, T.-N.; Yang, D.-H.; Nguyen, T.-A.T.; Lim, M.-S.; Hong, C.Y.; Kim, M.-H.; Lee, H.J.; Lee, Y.-K.; Cho, D.; Bae, S.-Y.; et al. Optimal culture conditions for the generation of natural killer cell-induced dendritic cells for cancer immunotherapy. Cell. Mol. Immunol. 2012, 9, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Gao, X.; Joyee, A.G.; Zhao, L.; Qiu, H.; Yang, M.; Fan, Y.; Wang, S.; Yang, X. NK Cells Promote Type 1 T Cell Immunity through Modulating the Function of Dendritic Cells during Intracellular Bacterial Infection. J. Immunol. 2011, 187, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, S.; Peng, Y.; Gao, X.; Joyee, A.G.; Wang, S.; Bai, H.; Zhao, L.; Yang, J.; Yang, X. NK cells modulate the lung dendritic cell-mediated Th1/Th17 immunity during intracellular bacterial infection. Eur. J. Immunol. 2015, 45, 2810–2820. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, S.; Yang, X. Natural killer cells in host defense against veterinary pathogens. Vet. Immunol. Immunopathol. 2015, 168, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Holl, E.K.; Roney, K.E.; Allen, I.C.; Steinbach, E.; Arthur, J.C.; Buntzman, A.; Plevy, S.; Frelinger, J.; Ting, J.P.Y. Plexin-B2 and Plexin-D1 in Dendritic Cells: Expression and IL-12/IL-23p40 Production. PLoS ONE 2012, 7, e43333. [Google Scholar] [CrossRef] [PubMed]
- Alamri, A.; Kung, S.; Department of Immunology, Max Rady College of Medicine, Rady Faculty of Health Sciences, University of Manitoba, Winnipeg, MB, Canada. Personal communication, 2017.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alamri, A.; Soussi Gounni, A.; Kung, S.K.P. View Point: Semaphorin-3E: An Emerging Modulator of Natural Killer Cell Functions? Int. J. Mol. Sci. 2017, 18, 2337. https://doi.org/10.3390/ijms18112337
Alamri A, Soussi Gounni A, Kung SKP. View Point: Semaphorin-3E: An Emerging Modulator of Natural Killer Cell Functions? International Journal of Molecular Sciences. 2017; 18(11):2337. https://doi.org/10.3390/ijms18112337
Chicago/Turabian StyleAlamri, Abdulaziz, Abdelilah Soussi Gounni, and Sam K. P. Kung. 2017. "View Point: Semaphorin-3E: An Emerging Modulator of Natural Killer Cell Functions?" International Journal of Molecular Sciences 18, no. 11: 2337. https://doi.org/10.3390/ijms18112337
APA StyleAlamri, A., Soussi Gounni, A., & Kung, S. K. P. (2017). View Point: Semaphorin-3E: An Emerging Modulator of Natural Killer Cell Functions? International Journal of Molecular Sciences, 18(11), 2337. https://doi.org/10.3390/ijms18112337