Metallothioneins: Emerging Modulators in Immunity and Infection



Abstract

{kind=link}
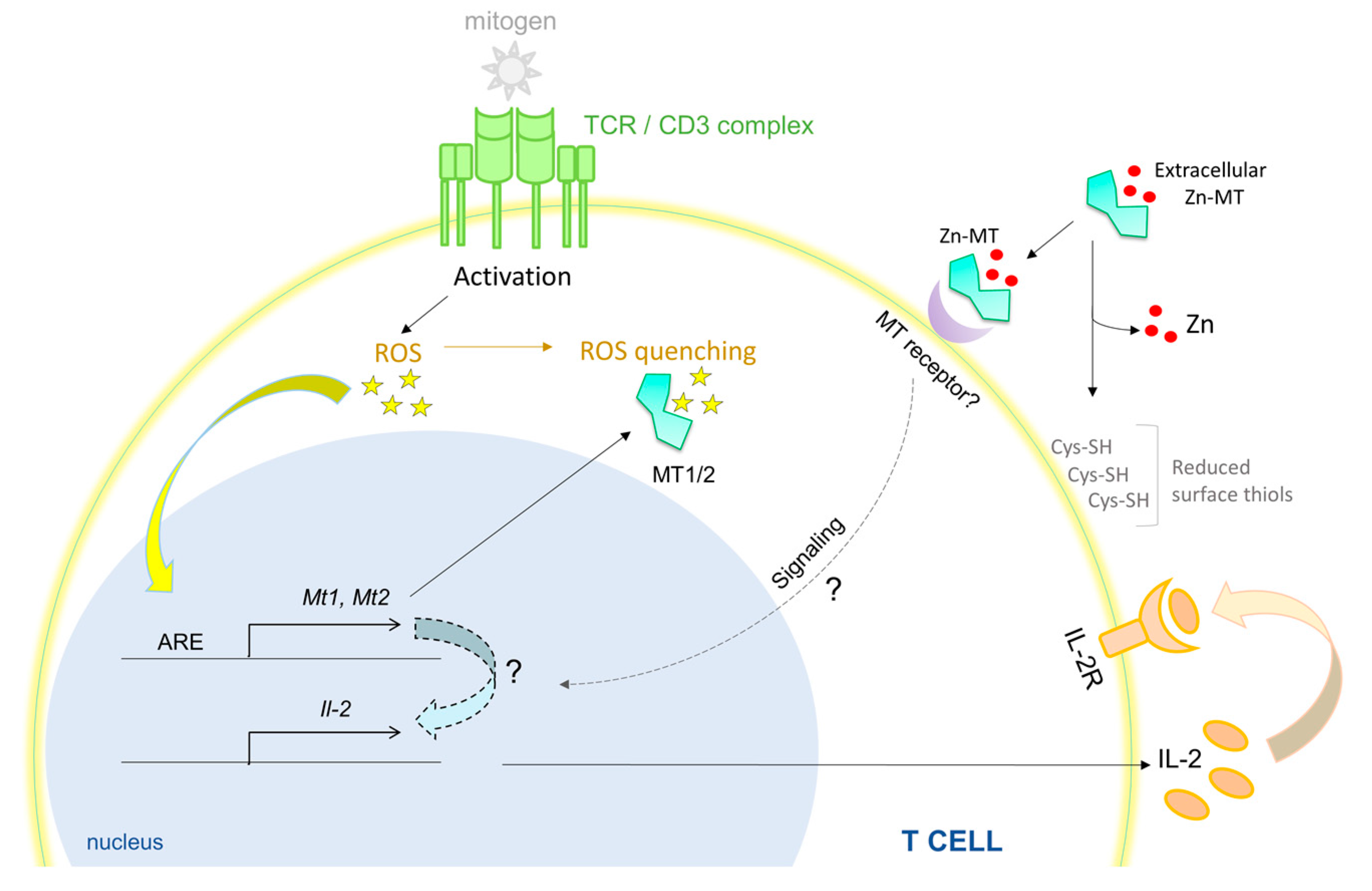{kind=link}
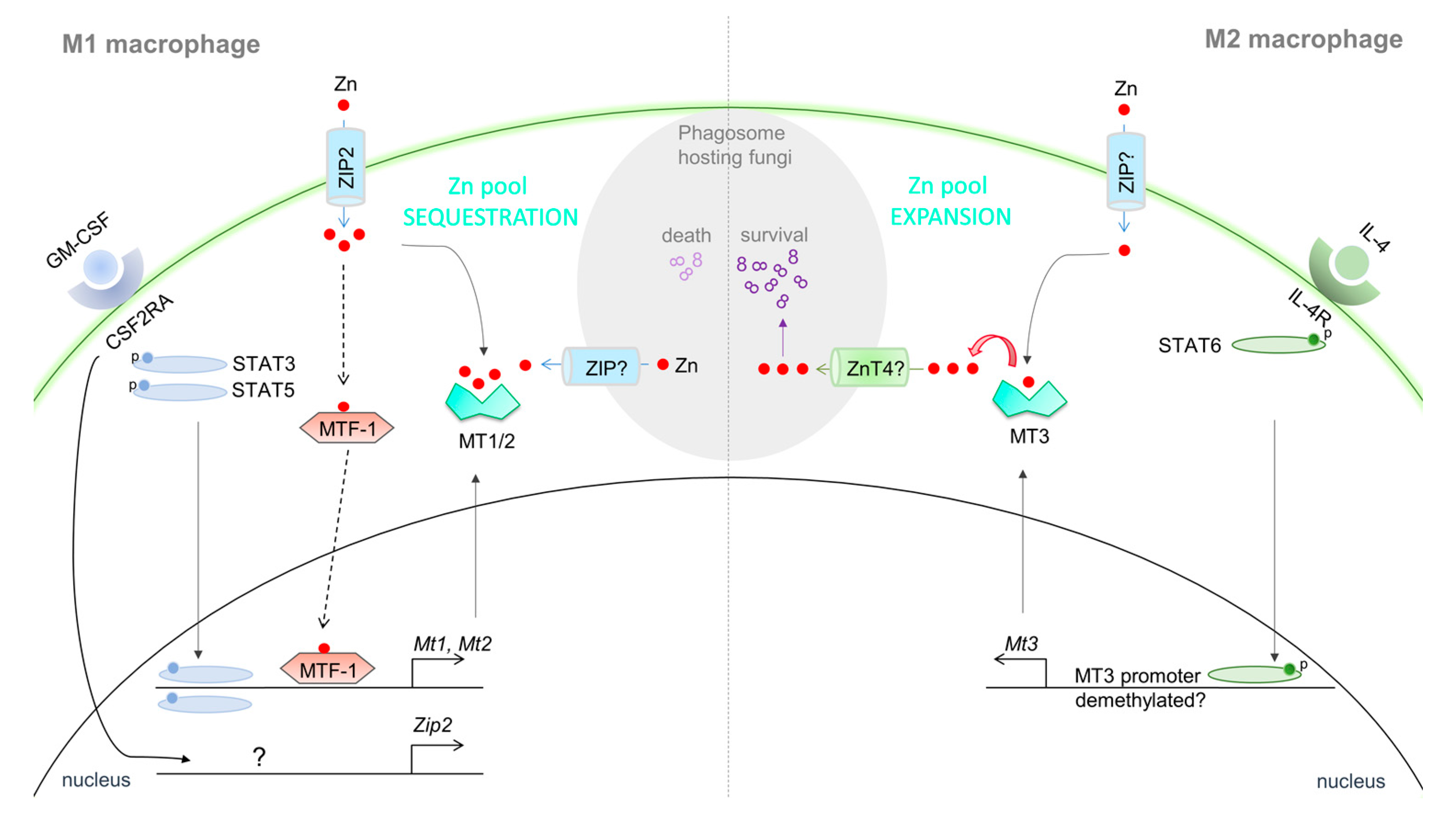{kind=link}
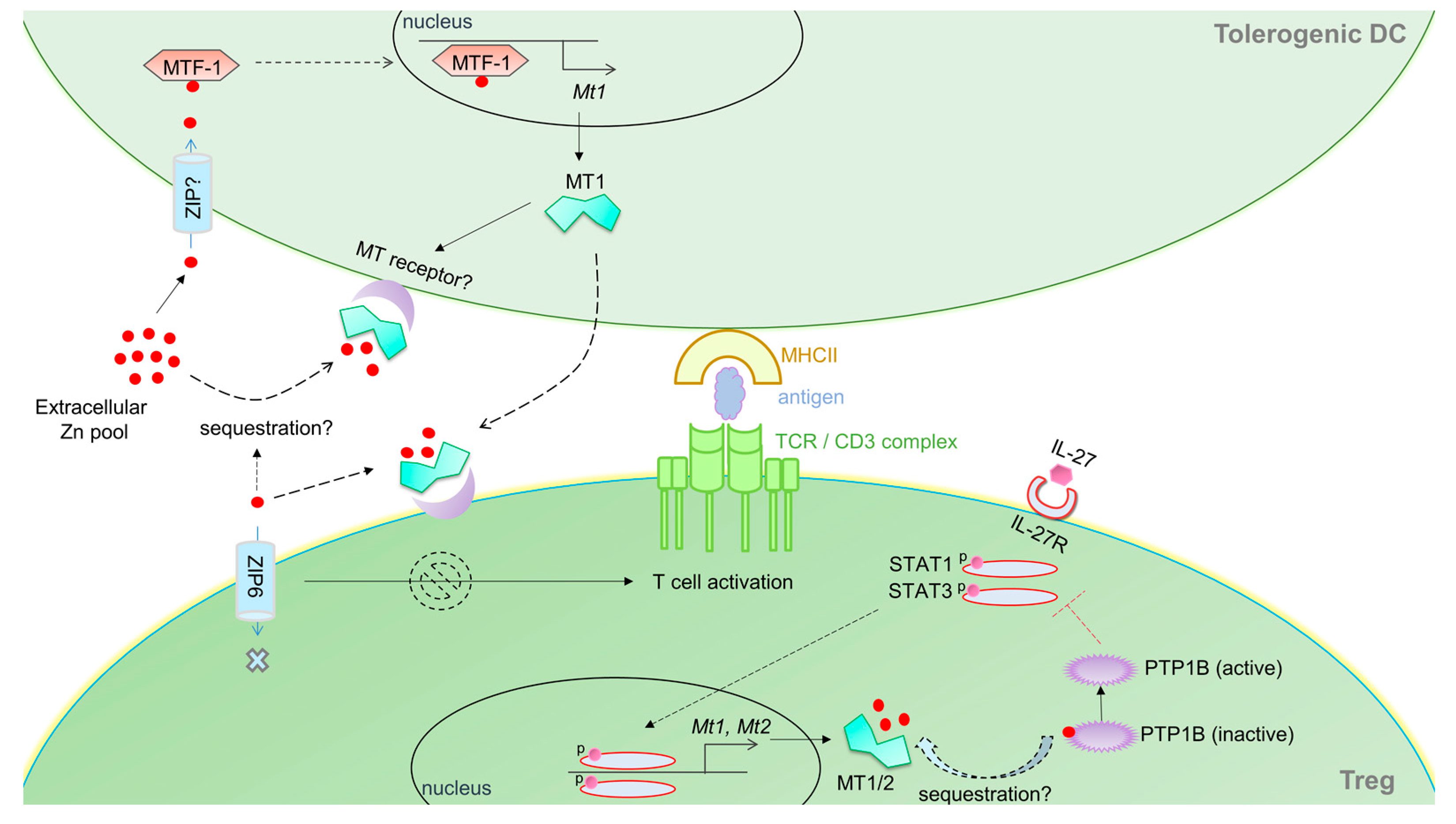{kind=link}
1. Introduction
2. The Metallothionein Family: Master Zinc Regulators
3. Immunity: Do Metallothioneins Take Center Stage?
4. Metallothionein Induction and Signaling in Immunity
5. Metallothioneins Respond to Microbial Stress
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| APC | antigen presenting cell |
| ARE | antioxidant response element |
| Ca | calcium |
| CCL | C-C motif ligand |
| Cd | cadmium |
| CTL | cytotoxic T lymphocyte |
| Cu | copper |
| Cys | cysteine |
| DC | dendritic cell |
| Egr-1 | early growth response-1 |
| ER | endoplasmic reticulum |
| Fe | iron |
| FoxP3 | Fork head box P3 |
| Gfi-1 | growth factor independent-1 |
| GM-CSF | granulocyte macrophage-colony stimulating factor |
| GRE | glucocorticoid response element |
| GSH | glutathione (reduced) |
| HCV | hepatitis C virus |
| Hg | mercury |
| ICAM-1 | intracellular adhesion molecule-1 |
| IFN | interferon |
| IL- | interleukin |
| IκB | inhibitor of κB |
| Iκκ | IκB kinase |
| LPS | lipopolysaccharide |
| MCP | monocyte chemotactic protein |
| Mg | magnesium |
| MHC | major histocompatibility class |
| MIP | macrophage inflammatory protein |
| Mn | manganese |
| MRE | metal response element |
| MT | Metallothionein |
| MT-null | MT1/2 deficient |
| MTF-1 | metal-response element-binding transcription factor-1 |
| NF-κB | nuclear factor-kappa B |
| NK | natural killer |
| NO | nitric oxide |
| PMA | phorbol myristate acetate |
| PTP | protein tyrosine phosphatase |
| ROS | reactive oxygen species |
| STAT | signal transducer and activator of transcription |
| Th | T helper cell |
| TNF | tumor necrosis factor, |
| Tr1 | Type-1 regulatory T cell |
| Treg | regulatory T cell |
| USF | upstream stimulatory factor |
| ZIP | zinc importer |
| Zn | zinc |
References
- Nelson, N. Metal ion transporters and homeostasis. EMBO J. 1999, 18, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Liu, J. Metallothionein transgenic and knock-out mouse models in the study of cadmium toxicity. J. Toxicol. Sci. 1998, 23 (Suppl. 2), 97–102. [Google Scholar] [CrossRef] [PubMed]
- Masters, B.A.; Kelly, E.J.; Quaife, C.J.; Brinster, R.L.; Palmiter, R.D. Targeted disruption of metallothionein i and ii genes increases sensitivity to cadmium. Proc. Natl. Acad. Sci. USA 1994, 91, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.R.; Cousins, R.J. Metallothionein expression in animals: A physiological perspective on function. J. Nutr. 2000, 130, 1085–1088. [Google Scholar] [PubMed]
- Davis, S.R.; McMahon, R.J.; Cousins, R.J. Metallothionein knockout and transgenic mice exhibit altered intestinal processing of zinc with uniform zinc-dependent zinc transporter-1 expression. J. Nutr. 1998, 128, 825–831. [Google Scholar] [PubMed]
- Liu, J.; Liu, Y.; Hartley, D.; Klaassen, C.D.; Shehin-Johnson, S.E.; Lucas, A.; Cohen, S.D. Metallothionein-i/ii knockout mice are sensitive to acetaminophen-induced hepatotoxicity. J. Pharmacol. Exp. Ther. 1999, 289, 580–586. [Google Scholar] [PubMed]
- Liu, Y.; Liu, J.; Habeebu, S.S.; Klaassen, C.D. Metallothionein protects against the nephrotoxicity produced by chronic cdmt exposure. Toxicol. Sci. 1999, 50, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Eckschlager, T.; Adam, V.; Hrabeta, J.; Figova, K.; Kizek, R. Metallothioneins and cancer. Curr. Protein Pept. Sci. 2009, 10, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Gobel, H.; van der Wal, A.C.; Teeling, P.; van der Loos, C.M.; Becker, A.E. Metallothionein in human atherosclerotic lesions: A scavenger mechanism for reactive oxygen species in the plaque? Virchows Arch. 2000, 437, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kawakami, T.; Kadota, Y.; Mori, M.; Suzuki, S. Obesity and metallothionein. Curr. Pharm. Biotechnol. 2013, 14, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Espejo, C.; Carrasco, J.; Hidalgo, J.; Penkowa, M.; Garcia, A.; Sáez-Torres, I.; Martı́nez-Cáceres, E.M. Differential expression of metallothioneins in the cns of mice with experimental autoimmune encephalomyelitis. Neuroscience 2001, 105, 1055–1065. [Google Scholar] [CrossRef]
- Pedersen, M.O.; Jensen, R.; Pedersen, D.S.; Skjolding, A.D.; Hempel, C.; Maretty, L.; Penkowa, M. Metallothionein-i+ii in neuroprotection. Biofactors 2009, 35, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Subramanian Vignesh, K.; Deepe, G.S., Jr. Immunological orchestration of zinc homeostasis: The battle between host mechanisms and pathogen defenses. Arch. Biochem. Biophys. 2016, 611, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Sherman, A.R. Zinc, copper, and iron nutriture and immunity. J. Nutr. 1992, 122, 604–609. [Google Scholar] [PubMed]
- Dziegiel, P.; Pula, B.; Kobierzycki, C.; Stasiolek, M.; Podhorska-Okolow, M. Metallothioneins and immune function. In Metallothioneins in Normal and Cancer Cells; Springer International Publishing: Cham, Switzerland, 2016; pp. 65–77. [Google Scholar]
- Bert, L.V. The function of metallothionein. Neurochem. Int. 1995, 27, 23–33. [Google Scholar]
- Blindauer, C.A.; Leszczyszyn, O.I. Metallothioneins: Unparalleled diversity in structures and functions for metal ion homeostasis and more. Nat. Prod. Rep. 2010, 27, 720–741. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Redox biochemistry of mammalian metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Vasak, M. Advances in metallothionein structure and functions. J. Trace Elem. Med. Biol. 2005, 19, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Mehus, A.A.; Muhonen, W.W.; Garrett, S.H.; Somji, S.; Sens, D.A.; Shabb, J.B. Quantitation of human metallothionein isoforms: A family of small, highly conserved, cysteine-rich proteins. Mol. Cell. Proteom. 2014, 13, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- National Library of Medicine (US); National Center for Biotechnology Information. Gene (Metallothionein) and “Homo Sapiens” [Porgn:__txid9606]; National Center for Biotechnology Information: Bethesda, MD, USA, 2004. Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 12 October 2017).
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- West, A.K.; Stallings, R.; Hildebrand, C.E.; Chiu, R.; Karin, M.; Richards, R.I. Human metallothionein genes: Structure of the functional locus at 16q13. Genomics 1990, 8, 513–518. [Google Scholar] [CrossRef]
- Piotrowski, J.K.; Trojanowska, B.; Sapota, A. Binding of cadmium and mercury by metallothionein in the kidneys and liver of rats following repeated administration. Arch. Toxicol. 1974, 32, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. The function of zinc metallothionein: A link between cellular zinc and redox state. J. Nutr. 2000, 130, 1455S–1458S. [Google Scholar] [PubMed]
- Eide, D.J. Zinc transporters and the cellular trafficking of zinc. Biochim. Biophys. Acta 2006, 1763, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian zinc transport, trafficking, and signals. J. Biol. Chem. 2006, 281, 24085–24089. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Vallee, B.L. Thiolate ligands in metallothionein confer redox activity on zinc clusters. Proc. Natl. Acad. Sci. USA 1998, 95, 3478–3482. [Google Scholar] [CrossRef] [PubMed]
- Krezel, A.; Maret, W. The functions of metamorphic metallothioneins in zinc and copper metabolism. Int. J. Mol. Sci. 2017, 18, 1237. [Google Scholar] [CrossRef] [PubMed]
- Rigby Duncan, K.E.; Stillman, M.J. Metal-dependent protein folding: Metallation of metallothionein. J. Inorg. Biochem. 2006, 100, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.J.; Maret, W.; Vallee, B.L. The ATP-metallothionein complex. Proc. Natl. Acad. Sci. USA 1998, 95, 9146–9149. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Oxidative metal release from metallothionein via zinc-thiol/disulfide interchange. Proc. Natl. Acad. Sci. USA 1994, 91, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.C.; Ni, F.Y.; Huang, Z.X. Neuronal growth-inhibitory factor (metallothionein-3): Structure-function relationships. FEBS J. 2010, 277, 2912–2920. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Q.; Cai, B.; Li, H.; Sze, K.H.; Huang, Z.X.; Wu, H.M.; Sun, H. Solution structure and dynamics of human metallothionein-3 (MT-3). FEBS Lett. 2006, 580, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Maret, W.; Vallee, B.L. Control of zinc transfer between thionein, metallothionein, and zinc proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 3489–3494. [Google Scholar] [CrossRef] [PubMed]
- Palumaa, P.; Eriste, E.; Njunkova, O.; Pokras, L.; Jornvall, H.; Sillard, R. Brain-specific metallothionein-3 has higher metal-binding capacity than ubiquitous metallothioneins and binds metals noncooperatively. Biochemistry 2002, 41, 6158–6163. [Google Scholar] [CrossRef] [PubMed]
- Tio, L.; Villarreal, L.; Atrian, S.; Capdevila, M. Functional differentiation in the mammalian metallothionein gene family: Metal binding features of mouse MT4 and comparison with its paralog MT1. J. Biol. Chem. 2004, 279, 24403–24413. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K. Cellular zinc sensors: MTF-1 regulation of gene expression. Biometals 2001, 14, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Gunther, V.; Lindert, U.; Schaffner, W. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta 2012, 1823, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Cherian, M.G.; Apostolova, M.D. Nuclear localization of metallothionein during cell proliferation and differentiation. Cell. Mol. Biol. 2000, 46, 347–356. [Google Scholar] [PubMed]
- Ye, B.; Maret, W.; Vallee, B.L. Zinc metallothionein imported into liver mitochondria modulates respiration. Proc. Natl. Acad. Sci. USA 2001, 98, 2317–2322. [Google Scholar] [CrossRef] [PubMed]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Caruso, J.A.; Deepe, G.S., Jr. Granulocyte macrophage-colony stimulating factor induced zn sequestration enhances macrophage superoxide and limits intracellular pathogen survival. Immunity 2013, 39, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Koh, J.Y. Roles of zinc and metallothionein-3 in oxidative stress-induced lysosomal dysfunction, cell death, and autophagy in neurons and astrocytes. Mol. Brain 2010, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Dittmer, P.J.; Park, J.G.; Jansen, K.B.; Palmer, A.E. Measuring steady-state and dynamic endoplasmic reticulum and golgi Zn2+ with genetically encoded sensors. Proc. Natl. Acad. Sci. USA 2011, 108, 7351–7356. [Google Scholar] [CrossRef] [PubMed]
- Wellenreuther, G.; Cianci, M.; Tucoulou, R.; Meyer-Klaucke, W.; Haase, H. The ligand environment of zinc stored in vesicles. Biochem. Biophys. Res. Commun. 2009, 380, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Levadoux, M.; Mahon, C.; Beattie, J.H.; Wallace, H.M.; Hesketh, J.E. Nuclear import of metallothionein requires its mrna to be associated with the perinuclear cytoskeleton. J. Biol. Chem. 1999, 274, 34961–34966. [Google Scholar] [CrossRef] [PubMed]
- El Ghazi, I.; Martin, B.L.; Armitage, I.M. New proteins found interacting with brain metallothionein-3 are linked to secretion. Int. J. Alzheimers’s Dis. 2010, 2011, 208634. [Google Scholar] [CrossRef] [PubMed]
- Lynes, M.A.; Zaffuto, K.; Unfricht, D.W.; Marusov, G.; Samson, J.S.; Yin, X. The physiological roles of extracellular metallothionein. Exp. Biol. Med. (Maywood) 2006, 231, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.; Borghesi, L.A.; Olson, E.A.; Lynes, M.A. Immunomodulatory activities of extracellular metallothionein. II. Effects on macrophage functions. J. Toxicol. Environ. Health 1995, 45, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Lynes, M.A.; Borghesi, L.A.; Youn, J.; Olson, E.A. Immunomodulatory activities of extracellular metallothionein. I. Metallothionein effects on antibody production. Toxicology 1993, 85, 161–177. [Google Scholar] [CrossRef]
- Lynes, M.A.; Garvey, J.S.; Lawrence, D.A. Extracellular metallothionein effects on lymphocyte activities. Mol. Immunol. 1990, 27, 211–219. [Google Scholar] [CrossRef]
- Yin, X.; Knecht, D.A.; Lynes, M.A. Metallothionein mediates leukocyte chemotaxis. BMC Immunol. 2005, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Fraker, P.J.; Gershwin, M.E.; Good, R.A.; Prasad, A. Interrelationships between zinc and immune function. Fed. Proc. 1986, 45, 1474–1479. [Google Scholar] [PubMed]
- Prasad, A.S. Effects of zinc deficiency on TH1 and TH2 cytokine shifts. J. Infect. Dis. 2000, 182 (Suppl. 1), S62–S68. [Google Scholar] [CrossRef] [PubMed]
- Shankar, A.H.; Prasad, A.S. Zinc and immune function: The biological basis of altered resistance to infection. Am. J. Clin. Nutr. 1998, 68, 447S–463S. [Google Scholar] [PubMed]
- Haase, H.; Rink, L. Zinc signals and immune function. Biofactors 2014, 40, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Rink, L.; Gabriel, P. Zinc and the immune system. Proc. Nutr. Soc. 2000, 59, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R. Excessive intake of zinc impairs immune responses. JAMA 1984, 252, 1443–1446. [Google Scholar] [CrossRef] [PubMed]
- Olafson, R.W. Thymus metallothionein: Regulation of zinc-thionein in the aging mouse. Can. J. Biochem. Cell Biol. 1985, 63, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Giacconi, R.; Cipriano, C.; Muti, E.; Gasparini, N.; Malavolta, M. Are zinc-bound metallothionein isoforms (i + ii and iii) involved in impaired thymulin production and thymic involution during ageing? Immun. Ageing 2004, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Bach, J.F.; Dardenne, M. Thymulin, a zinc-dependent hormone. Med. Oncol. Tumor Pharmacother. 1989, 6, 25–29. [Google Scholar] [PubMed]
- Savino, W.; Huang, P.C.; Corrigan, A.; Berrih, S.; Dardenne, M. Thymic hormone-containing cells. V. Immunohistological detection of metallothionein within the cells bearing thymulin (a zinc-containing hormone) in human and mouse thymuses. J. Histochem. Cytochem. 1984, 32, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; Lo, S.K. Immunohistochemical metallothionein expression in thymoma: Correlation with histological types and cellular origin. Histopathology 1997, 30, 243–248. [Google Scholar] [CrossRef] [PubMed]
- George, M.M.; Subramanian Vignesh, K.; Landero Figueroa, J.A.; Caruso, J.A.; Deepe, G.S. Zinc induces dendritic cell tolerogenic phenotype and skews regulatory T cell–TH17 balance. J. Immunol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, H.; Morikawa, H.; Kamon, H.; Iguchi, M.; Hojyo, S.; Fukada, T.; Yamashita, S.; Kaisho, T.; Akira, S.; Murakami, M.; et al. Toll-like receptor-mediated regulation of zinc homeostasis influences dendritic cell function. Nat. Immunol. 2006, 7, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Weinlich, G.; Topar, G.; Eisendle, K.; Fritsch, P.O.; Zelger, B. Comparison of metallothionein-overexpression with sentinel lymph node biopsy as prognostic factors in melanoma. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Haerslev, T.; Jacobsen, G.K.; Zedeler, K. The prognostic significance of immunohistochemically detectable metallothionein in primary breast carcinomas. APMIS 1995, 103, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M. Lymphoid and myeloid lineage commitment in multipotent hematopoietic progenitors. Immunol. Rev. 2010, 238, 37–46. [Google Scholar] [CrossRef] [PubMed]
- King, L.E.; Osati-Ashtiani, F.; Fraker, P.J. Depletion of cells of the b lineage in the bone marrow of zinc-deficient mice. Immunology 1995, 85, 69–73. [Google Scholar] [PubMed]
- Fraker, P.J.; King, L.E. Reprogramming of the immune system during zinc deficiency. Annu. Rev. Nutr. 2004, 24, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Huber, K.L.; Cousins, R.J. Metallothionein expression in rat bone marrow is dependent on dietary zinc but not dependent on interleukin-1 or interleukin-6. J. Nutr. 1993, 123, 642–648. [Google Scholar] [PubMed]
- Krishnaraju, K.; Nguyen, H.Q.; Liebermann, D.A.; Hoffman, B. The zinc finger transcription factor EGR-1 potentiates macrophage differentiation of hematopoietic cells. Mol. Cell. Biol. 1995, 15, 5499–5507. [Google Scholar] [CrossRef] [PubMed]
- Hock, H.; Hamblen, M.J.; Rooke, H.M.; Traver, D.; Bronson, R.T.; Cameron, S.; Orkin, S.H. Intrinsic requirement for zinc finger transcription factor GFI-1 in neutrophil differentiation. Immunity 2003, 18, 109–120. [Google Scholar] [CrossRef]
- Feng, W.; Cai, J.; Pierce, W.M.; Franklin, R.B.; Maret, W.; Benz, F.W.; Kang, Y.J. Metallothionein transfers zinc to mitochondrial aconitase through a direct interaction in mouse hearts. Biochem. Biophys. Res. Commun. 2005, 332, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.J.; Maret, W.; Vallee, B.L. The glutathione redox couple modulates zinc transfer from metallothionein to zinc-depleted sorbitol dehydrogenase. Proc. Natl. Acad. Sci. USA 1998, 95, 3483–3488. [Google Scholar] [CrossRef] [PubMed]
- Roesijadi, G.; Bogumil, R.; Vasak, M.; Kagi, J.H. Modulation of DNA binding of a tramtrack zinc finger peptide by the metallothionein-thionein conjugate pair. J. Biol. Chem. 1998, 273, 17425–17432. [Google Scholar] [CrossRef] [PubMed]
- McCormick, N.H.; Kelleher, S.L. Znt4 provides zinc to zinc-dependent proteins in the trans-golgi network critical for cell function and zn export in mammary epithelial cells. Am. J. Physiol. Cell Physiol. 2012, 303, C291–C297. [Google Scholar] [CrossRef] [PubMed]
- Cousins, R.J. A role of zinc in the regulation of gene expression. Proc. Nutr. Soc. 1998, 57, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Leonard, S.W.; Traber, M.G.; Ho, E. Zinc deficiency affects DNA damage, oxidative stress, antioxidant defenses, and DNA repair in rats. J. Nutr. 2009, 139, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Suzuki, S. Endoplasmic reticulum stress and metallothionein. Yakugaku Zasshi 2007, 127, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Crowthers, K.C.; Kline, V.; Giardina, C.; Lynes, M.A. Augmented humoral immune function in metallothionein-null mice. Toxicol. Appl. Pharmacol. 2000, 166, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Mita, M.; Imura, N.; Kumazawa, Y.; Himeno, S. Suppressed proliferative response of spleen T cells from metallothionein null mice. Microbiol. Immunol. 2002, 46, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.H.; Devadas, S.; Kwon, J.; Pinto, L.A.; Williams, M.S. T cells express a phagocyte-type nadph oxidase that is activated after t cell receptor stimulation. Nat. Immunol. 2004, 5, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, K.; Majumder, S.; Zhu, Q.; Hunzeker, J.; Datta, J.; Shah, M.; Sheridan, J.F.; Jacob, S.T. Influenza virus infection induces metallothionein gene expression in the mouse liver and lung by overlapping but distinct molecular mechanisms. Mol. Cell. Biol. 2001, 21, 8301–8317. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Macrophages and systemic iron homeostasis. J. Innate Immun. 2012, 4, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Kojima, N.; Young, C.R.; Bates, G.W. Failure of metallothionein to bind iron or act as an iron mobilizing agent. Biochim. Biophys. Acta 1982, 716, 273–275. [Google Scholar] [CrossRef]
- Robertson, A.; Morrison, J.N.; Wood, A.M.; Bremner, I. Effects of iron deficiency on metallothionein-I concentrations in blood and tissues of rats. J. Nutr. 1989, 119, 439–445. [Google Scholar] [PubMed]
- Lonnerdal, B. Dietary factors influencing zinc absorption. J. Nutr. 2000, 130, 1378S–1383S. [Google Scholar] [PubMed]
- Rossander-Hulten, L.; Brune, M.; Sandstrom, B.; Lonnerdal, B.; Hallberg, L. Competitive inhibition of iron absorption by manganese and zinc in humans. Am. J. Clin. Nutr. 1991, 54, 152–156. [Google Scholar] [PubMed]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. The role of metallothionein in oxidative stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Muraoka, S.; Ogiso, T. Antioxidant activity of metallothionein compared with reduced glutathione. Life Sci. 1997, 60, PL 301–PL 309. [Google Scholar] [CrossRef]
- Liu, M.J.; Bao, S.; Galvez-Peralta, M.; Pyle, C.J.; Rudawsky, A.C.; Pavlovicz, R.E.; Killilea, D.W.; Li, C.; Nebert, D.W.; Wewers, M.D.; et al. Zip8 regulates host defense through zinc-mediated inhibition of NF-κB. Cell Rep. 2013, 3, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Butcher, H.L.; Kennette, W.A.; Collins, O.; Zalups, R.K.; Koropatnick, J. Metallothionein mediates the level and activity of nuclear factor kappa b in murine fibroblasts. J. Pharmacol. Exp. Ther. 2004, 310, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, S.H.; Weissman, I.L. Lymphoid tissue architecture. II. Ontogeny of peripheral T and B cells in mice: Evidence against peyer’s patches as the site of generation of B cells. J. Immunol. 1974, 113, 1477–1492. [Google Scholar] [PubMed]
- Gutman, G.A.; Weissman, I.L. Lymphoid tissue architecture. Experimental analysis of the origin and distribution of T-cells and B-cells. Immunology 1972, 23, 465–479. [Google Scholar] [PubMed]
- Weissman, I.L.; Gutman, G.A.; Friedberg, S.H.; Jerabek, L. Lymphoid tissue architecture. III. Germinal centers, T cells, and thymus-dependent vs. thymus-independent antigens. Adv. Exp. Med. Biol. 1976, 66, 229–237. [Google Scholar] [PubMed]
- Kataru, R.P.; Lee, Y.G.; Koh, G.Y. Interactions of immune cells and lymphatic vessels. Adv. Anat. Embryol. Cell Biol. 2014, 214, 107–118. [Google Scholar] [PubMed]
- Vallee, B.L.; Gibson, J.G., II. The zinc content of normal human whole blood, plasma, leucocytes, and erythrocytes. J. Biol. Chem. 1948, 176, 445–457. [Google Scholar] [PubMed]
- Schrodt, G.R.; Hall, T.; Whitmore, W.F., Jr. The concentration of zinc in diseased human prostate glands. Cancer 1964, 17, 1555–1566. [Google Scholar] [CrossRef]
- De, S.K.; McMaster, M.T.; Andrews, G.K. Endotoxin induction of murine metallothionein gene expression. J. Biol. Chem. 1990, 265, 15267–15274. [Google Scholar] [PubMed]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Divanovic, S.; Caruso, J.A.; Deepe, G.S., Jr. Interleukin-4 induces metallothionein 3- and SLC30A4-dependent increase in intracellular Zn2+ that promotes pathogen persistence in macrophages. Cell Rep. 2016, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Pot, C.; Apetoh, L.; Thalhamer, T.; Zhu, B.; Murugaiyan, G.; Xiao, S.; Lee, Y.; Rangachari, M.; Yosef, N.; et al. Metallothioneins negatively regulate IL-27-induced type 1 regulatory T-cell differentiation. Proc. Natl. Acad. Sci. USA 2013, 110, 7802–7807. [Google Scholar] [CrossRef] [PubMed]
- Kanekiyo, M.; Itoh, N.; Kawasaki, A.; Matsuyama, A.; Matsuda, K.; Nakanishi, T.; Tanaka, K. Metallothionein modulates lipopolysaccharide-stimulated tumour necrosis factor expression in mouse peritoneal macrophages. Biochem. J. 2002, 361, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, T.; Kuroda, E.; Yamashita, U. Dysfunction of macrophages in metallothionein-knock out mice. J. UOEH 2004, 26, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Shortman, K.; Liu, Y.J. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002, 2, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, R.A.; von Andrian, U.H. How tolerogenic dendritic cells induce regulatory T cells. Adv. Immunol. 2010, 108, 111–165. [Google Scholar] [PubMed]
- Reis e Sousa, C. Dendritic cells in a mature age. Nat. Rev. Immunol. 2006, 6, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.I.; Suzuki, T.; Nagai, S.; Yamashita, T.; Toyoda, N.; Matsushima, K. Identification of genes specifically expressed in human activated and mature dendritic cells through serial analysis of gene expression. Blood 2000, 96, 2206–2214. [Google Scholar] [PubMed]
- Spiering, R.; Wagenaar-Hilbers, J.; Huijgen, V.; van der Zee, R.; van Kooten, P.J.; van Eden, W.; Broere, F. Membrane-bound metallothionein 1 of murine dendritic cells promotes the expansion of regulatory T cells in vitro. Toxicol. Sci. 2014, 138, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Spiering, R.; van der Zee, R.; Wagenaar, J.; Kapetis, D.; Zolezzi, F.; van Eden, W.; Broere, F. Tolerogenic dendritic cells that inhibit autoimmune arthritis can be induced by a combination of carvacrol and thermal stress. PLoS ONE 2012, 7, e46336. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Lee, W.W.; Tomar, D.; Pryshchep, S.; Czesnikiewicz-Guzik, M.; Lamar, D.L.; Li, G.; Singh, K.; Tian, L.; Weyand, C.M.; et al. Regulation of T cell receptor signaling by activation-induced zinc influx. J. Exp. Med. 2011, 208, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Borghesi, L.A.; Youn, J.; Olson, E.A.; Lynes, M.A. Interactions of metallothionein with murine lymphocytes: Plasma membrane binding and proliferation. Toxicology 1996, 108, 129–140. [Google Scholar] [CrossRef]
- Rosenkranz, E.; Metz, C.H.; Maywald, M.; Hilgers, R.D.; Wessels, I.; Senff, T.; Haase, H.; Jager, M.; Ott, M.; Aspinall, R.; et al. Zinc supplementation induces regulatory T cells by inhibition of SIRT-1 deacetylase in mixed lymphocyte cultures. Mol. Nutr. Food Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.M.; Zweifach, A.; Lynes, M.A. Metallothionein regulates intracellular zinc signaling during CD4(+) T cell activation. BMC Immunol. 2016, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.; Lee, K.; Yun, H.S.; Paik, D.J.; Kim, J.M.; Youn, J. Functions of metallothionein generating interleukin-10-producing regulatory CD4+ t cells potentiate suppression of collagen-induced arthritis. J. Microbiol. Biotechnol. 2007, 17, 348–358. [Google Scholar] [PubMed]
- Haase, H.; Rink, L. Functional significance of zinc-related signaling pathways in immune cells. Annu. Rev. Nutr. 2009, 29, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Brockmann, L.; Gagliani, N.; Steglich, B.; Giannou, A.D.; Kempski, J.; Pelczar, P.; Geffken, M.; Mfarrej, B.; Huber, F.; Herkel, J.; et al. IL-10 receptor signaling is essential for TR1 cell function in vivo. J. Immunol. 2017, 198, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.; Lynes, M.A. Metallothionein-induced suppression of cytotoxic T lymphocyte function: An important immunoregulatory control. Toxicol. Sci. 1999, 52, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Beck, F.W.; Prasad, A.S.; Kaplan, J.; Fitzgerald, J.T.; Brewer, G.J. Changes in cytokine production and T cell subpopulations in experimentally induced zinc-deficient humans. Am. J. Physiol. 1997, 272, E1002–E1007. [Google Scholar] [PubMed]
- Fernandes, G.; Nair, M.; Onoe, K.; Tanaka, T.; Floyd, R.; Good, R.A. Impairment of cell-mediated immunity functions by dietary zinc deficiency in mice. Proc. Natl. Acad. Sci. USA 1979, 76, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, K.S.; Kim, H.N.; Kim, H.J.; Koh, J.Y. Role of zinc metallothionein-3 (ZNMT3) in epidermal growth factor (EGF)-induced C-ABL protein activation and actin polymerization in cultured astrocytes. J. Biol. Chem. 2011, 286, 40847–40856. [Google Scholar] [CrossRef] [PubMed]
- El Refaey, H.; Ebadi, M.; Kuszynski, C.A.; Sweeney, J.; Hamada, F.M.; Hamed, A. Identification of metallothionein receptors in human astrocytes. Neurosci. Lett. 1997, 231, 131–134. [Google Scholar] [CrossRef]
- Stitt, M.S.; Wasserloos, K.J.; Tang, X.; Liu, X.; Pitt, B.R.; St Croix, C.M. Nitric oxide-induced nuclear translocation of the metal responsive transcription factor, MTF-1 is mediated by zinc release from metallothionein. Vasc. Pharmacol. 2006, 44, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Carrasco, J.; Hidalgo, J.; Andrews, G.K. Identification of a signal transducer and activator of transcription (STAT) binding site in the mouse metallothionein-i promoter involved in interleukin-6-induced gene expression. Biochem. J. 1999, 337 Pt 1, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Miyagi, C.; Fukada, T.; Kagara, N.; Che, Y.S.; Hirano, T. Zinc transporter livi controls epithelial-mesenchymal transition in zebrafish gastrula organizer. Nature 2004, 429, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Minakami, R.; Sumimotoa, H. Phagocytosis-coupled activation of the superoxide-producing phagocyte oxidase, a member of the nadph oxidase (NOX) family. Int. J. Hematol. 2006, 84, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Fernandez, M.A.; Fernandez, M.A.; Fresno, M. Activation of human macrophages for the killing of intracellular trypanosoma cruzi by TNF-α and IFN-γ through a nitric oxide-dependent mechanism. Immunol. Lett. 1992, 33, 35–40. [Google Scholar] [CrossRef]
- Wolf, J.E.; Massof, S.E. In vivo activation of macrophage oxidative burst activity by cytokines and amphotericin B. Infect. Immun. 1990, 58, 1296–1300. [Google Scholar] [PubMed]
- Segal, A.W. How superoxide production by neutrophil leukocytes kills microbes. Novartis Found. Symp. 2006, 279, 92–98. [Google Scholar] [PubMed]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Rink, L.; Haase, H. Zinc signals in neutrophil granulocytes are required for the formation of neutrophil extracellular traps. Innate Immun. 2013, 19, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K. Regulation of metallothionein gene expression by oxidative stress and metal ions. Biochem. Pharmacol. 2000, 59, 95–104. [Google Scholar] [CrossRef]
- Kelly, E.J.; Sandgren, E.P.; Brinster, R.L.; Palmiter, R.D. A pair of adjacent glucocorticoid response elements regulate expression of two mouse metallothionein genes. Proc. Natl. Acad. Sci. USA 1997, 94, 10045–10050. [Google Scholar] [CrossRef] [PubMed]
- Franchimont, D. Overview of the actions of glucocorticoids on the immune response: A good model to characterize new pathways of immunosuppression for new treatment strategies. Ann. N. Y. Acad. Sci. 2004, 1024, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Reeve, V.E.; Nishimura, N.; Bosnic, M.; Michalska, A.E.; Choo, K.H. Lack of metallothionein-I and -II exacerbates the immunosuppressive effect of ultraviolet B radiation and cis-urocanic acid in mice. Immunology 2000, 100, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D.; Whitmore, T.E.; Durnam, D.M. Mt-iii, a brain-specific member of the metallothionein gene family. Proc. Natl. Acad. Sci. USA 1992, 89, 6333–6337. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, I.; Suzuki, J.S.; Kanazawa, H.; Hara, A.; Saio, M.; Inuzuka, T.; Miyairi, S.; Naganuma, A.; Tohyama, C. Metallothionein-3 is expressed in the brain and various peripheral organs of the rat. Neurosci. Lett. 2008, 438, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Slusser, A.; Zheng, Y.; Zhou, X.D.; Somji, S.; Sens, D.A.; Sens, M.A.; Garrett, S.H. Metallothionein isoform 3 expression in human skin, related cancers and human skin derived cell cultures. Toxicol. Lett. 2014, 232, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.F.; Xu, L.X.; Lu, J.; Cao, L.; Li, Z.H.; Hu, S.Y.; Wang, N.N.; Du, X.J.; Sun, L.C.; Zhao, W.L.; et al. Metallothionein III (MT3) is a putative tumor suppressor gene that is frequently inactivated in pediatric acute myeloid leukemia by promoter hypermethylation. J. Transl. Med. 2014, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Somji, S.; Garrett, S.H.; Zhou, X.D.; Zheng, Y.; Sens, D.A.; Sens, M.A. Absence of metallothionein 3 expression in breast cancer is a rare, but favorable marker of outcome that is under epigenetic control. Toxicol. Environ. Chem. 2010, 92, 1673–1695. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Hu, T.L.; Jiang, A.; Washington, M.K.; Moskaluk, C.A.; Schneider-Stock, R.; El-Rifai, W. Location-specific epigenetic regulation of the metallothionein 3 gene in esophageal adenocarcinomas. PLoS ONE 2011, 6, e22009. [Google Scholar] [CrossRef] [PubMed]
- Haq, F.; Mahoney, M.; Koropatnick, J. Signaling events for metallothionein induction. Mutat. Res. 2003, 533, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Majumder, S.; Kutay, H.; Datta, J.; Summers, D.; Jacob, S.T.; Ghoshal, K. Epigenetic regulation of metallothionein-I gene expression: Differential regulation of methylated and unmethylated promoters by DNA methyltransferases and methyl cpg binding proteins. J. Cell. Biochem. 2006, 97, 1300–1316. [Google Scholar] [CrossRef] [PubMed]
- Busslinger, M.; Tarakhovsky, A. Epigenetic control of immunity. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Rada, B.K.; Geiszt, M.; Kaldi, K.; Timar, C.; Ligeti, E. Dual role of phagocytic nadph oxidase in bacterial killing. Blood 2004, 104, 2947–2953. [Google Scholar] [CrossRef] [PubMed]
- Botella, H.; Peyron, P.; Levillain, F.; Poincloux, R.; Poquet, Y.; Brandli, I.; Wang, C.; Tailleux, L.; Tilleul, S.; Charriere, G.M.; et al. Mycobacterial P(1)-type atpases mediate resistance to zinc poisoning in human macrophages. Cell Host Microbe 2011, 10, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T.; Suzuki, K.; Kondo, T.; Nakazato, K.; Kakizaki, S.; Takagi, H.; Nakajima, K. Interferon-α-induced changes in metallothionein expression in liver biopsies from patients with chronic hepatitis C. Can. J. Gastroenterol. 2005, 19, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.R.; Shanley, T.P.; Sakthivel, B.; Cvijanovich, N.; Lin, R.; Allen, G.L.; Thomas, N.J.; Doctor, A.; Kalyanaraman, M.; Tofil, N.M.; et al. Genome-level expression profiles in pediatric septic shock indicate a role for altered zinc homeostasis in poor outcome. Physiol. Genom. 2007, 30, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Kominsky, D.J.; Campbell, E.L.; Colgan, S.P. Metabolic shifts in immunity and inflammation. J. Immunol. 2010, 184, 4062–4068. [Google Scholar] [CrossRef] [PubMed]
- Sobocinski, P.Z.; Canterbury, W.J., Jr. Hepatic metallothionein induction in inflammation. Ann. N. Y. Acad. Sci. 1982, 389, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B.; Chang, S.M.; Guthrie, G.J.; Maki, A.B.; Ryu, M.S.; Karabiyik, A.; Cousins, R.J. Zinc transporter ZIP14 functions in hepatic zinc, iron and glucose homeostasis during the innate immune response (endotoxemia). PLoS ONE 2012, 7, e48679. [Google Scholar]
- Sobocinski, P.Z.; Canterbury, W.J., Jr.; Mapes, C.A.; Dinterman, R.E. Involvement of hepatic metallothioneins in hypozincemia associated with bacterial infection. Am. J. Physiol. 1978, 234, E399–E406. [Google Scholar] [PubMed]
- Knoell, D.L.; Julian, M.W.; Bao, S.; Besecker, B.; Macre, J.E.; Leikauf, G.D.; DiSilvestro, R.A.; Crouser, E.D. Zinc deficiency increases organ damage and mortality in a murine model of polymicrobial sepsis. Crit. Care Med. 2009, 37, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Kogan, S.; Sood, A.; Garnick, M.S. Zinc and wound healing: A review of zinc physiology and clinical applications. Wounds 2017, 29, 102–106. [Google Scholar] [PubMed]
- Mita, M.; Satoh, M.; Shimada, A.; Okajima, M.; Azuma, S.; Suzuki, J.S.; Sakabe, K.; Hara, S.; Himeno, S. Metallothionein is a crucial protective factor against helicobacter pylori-induced gastric erosive lesions in a mouse model. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G877–G884. [Google Scholar] [CrossRef] [PubMed]
- Emeny, R.T.; Marusov, G.; Lawrence, D.A.; Pederson-Lane, J.; Yin, X.; Lynes, M.A. Manipulations of metallothionein gene dose accelerate the response to Listeria monocytogenes. Chem. Biol. Interact. 2009, 181, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Carrera, G.; Paternain, J.L.; Carrere, N.; Folch, J.; Courtade-Saidi, M.; Orfila, C.; Vinel, J.P.; Alric, L.; Pipy, B. Hepatic metallothionein in patients with chronic hepatitis C: Relationship with severity of liver disease and response to treatment. Am. J. Gastroenterol. 2003, 98, 1142–1149. [Google Scholar] [PubMed]
- O’Connor, K.S.; Parnell, G.; Patrick, E.; Ahlenstiel, G.; Suppiah, V.; van der Poorten, D.; Read, S.A.; Leung, R.; Douglas, M.W.; Yang, J.Y.; et al. Hepatic metallothionein expression in chronic hepatitis C virus infection is IFNL3 genotype-dependent. Genes Immun. 2014, 15, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Ilback, N.G.; Glynn, A.W.; Wikberg, L.; Netzel, E.; Lindh, U. Metallothionein is induced and trace element balance changed in target organs of a common viral infection. Toxicology 2004, 199, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Lazarczyk, M.; Favre, M. Role of Zn2+ ions in host-virus interactions. J. Virol. 2008, 82, 11486–11494. [Google Scholar] [CrossRef] [PubMed]
- Suara, R.O.; Crowe, J.E., Jr. Effect of zinc salts on respiratory syncytial virus replication. Antimicrob. Agents Chemother. 2004, 48, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Cancellieri, M.; Bassetto, M.; Widjaja, I.; van Kuppeveld, F.; de Haan, C.A.; Brancale, A. In silico structure-based design and synthesis of novel anti-rsv compounds. Antivir. Res. 2015, 122, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Hulisz, D. Efficacy of zinc against common cold viruses: An overview. J. Am. Pharm. Assoc. 2004, 44, 594–603. [Google Scholar] [CrossRef]
- Krenn, B.M.; Gaudernak, E.; Holzer, B.; Lanke, K.; Van Kuppeveld, F.J.; Seipelt, J. Antiviral activity of the zinc ionophores pyrithione and hinokitiol against picornavirus infections. J. Virol. 2009, 83, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Sera, T. Inhibition of virus DNA replication by artificial zinc finger proteins. J. Virol. 2005, 79, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Allendoerfer, R.; Deepe, G.S., Jr. Intrapulmonary response to Histoplasma capsulatum in gamma interferon knockout mice. Infect. Immun. 1997, 65, 2564–2569. [Google Scholar] [PubMed]
- Haase, H. An element of life: Competition for zinc in host-pathogen interaction. Immunity 2013, 39, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Subramanian Vignesh, K.; Landero Figueroa, J.A.; Porollo, A.; Caruso, J.A.; Deepe, G.S., Jr. Zinc sequestration: Arming phagocyte defense against fungal attack. PLoS Pathog. 2013, 9, e1003815. [Google Scholar]
- Wu, A.; Tymoszuk, P.; Haschka, D.; Heeke, S.; Dichtl, S.; Petzer, V.; Seifert, M.; Hilbe, R.; Sopper, S.; Talasz, H.; et al. Salmonella utilizes zinc to subvert anti-microbial host defense of macrophages via modulation of nf-kappab signaling. Infect. Immun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Z.; Jellbauer, S.; Poe, A.J.; Ton, V.; Pesciaroli, M.; Kehl-Fie, T.E.; Restrepo, N.A.; Hosking, M.P.; Edwards, R.A.; Battistoni, A.; et al. Zinc sequestration by the neutrophil protein calprotectin enhances salmonella growth in the inflamed gut. Cell Host Microbe 2012, 11, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Kapetanovic, R.; Bokil, N.J.; Achard, M.E.; Ong, C.Y.; Peters, K.M.; Stocks, C.J.; Phan, M.D.; Monteleone, M.; Schroder, K.; Irvine, K.M.; et al. Salmonella employs multiple mechanisms to subvert the tlr-inducible zinc-mediated antimicrobial response of human macrophages. FASEB J. 2016, 30, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Festa, R.A.; Chen, Y.L.; Espart, A.; Palacios, O.; Espin, J.; Capdevila, M.; Atrian, S.; Heitman, J.; Thiele, D.J. Cryptococcus neoformans copper detoxification machinery is critical for fungal virulence. Cell Host Microbe 2013, 13, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Szymczak, W.A.; Deepe, G.S., Jr. The CCL7-CCL2-CCR2 axis regulates IL-4 production in lungs and fungal immunity. J. Immunol. 2009, 183, 1964–1974. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Kroetz, D.N.; Tweedle, J.L.; Deepe, G.S., Jr. Type II cytokines impair host defense against an intracellular fungal pathogen by amplifying macrophage generation of IL-33. Mucosal. Immunol. 2015, 8, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.E.; Koski, K.G. Zinc deficiency impairs immune responses against parasitic nematode infections at intestinal and systemic sites. J. Nutr. 2000, 130, 1412S–1420S. [Google Scholar] [PubMed]
- Ragusa, M.A.; Nicosia, A.; Costa, S.; Cuttitta, A.; Gianguzza, F. Metallothionein gene family in the sea urchin paracentrotus lividus: Gene structure, differential expression and phylogenetic analysis. Int. J. Mol. Sci. 2017, 18, 812. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F.; Kasahara, M. Origin and evolution of the adaptive immune system: Genetic events and selective pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.D.; Butler, R.N.; Philcox, J.C.; Rofe, A.M.; Howarth, G.S.; Coyle, P. Regional distribution of metallothionein and zinc in the mouse gut: Comparison with metallothionien-null mice. Biol. Trace Elem. Res. 1998, 63, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Waeytens, A.; de Vos, M.; Laukens, D. Evidence for a potential role of metallothioneins in inflammatory bowel diseases. Mediat. Inflamm. 2009, 2009, 729172. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramanian Vignesh, K.; Deepe Jr., G.S. Metallothioneins: Emerging Modulators in Immunity and Infection. Int. J. Mol. Sci. 2017, 18, 2197. https://doi.org/10.3390/ijms18102197
Subramanian Vignesh K, Deepe Jr. GS. Metallothioneins: Emerging Modulators in Immunity and Infection. International Journal of Molecular Sciences. 2017; 18(10):2197. https://doi.org/10.3390/ijms18102197
Chicago/Turabian StyleSubramanian Vignesh, Kavitha, and George S. Deepe Jr. 2017. "Metallothioneins: Emerging Modulators in Immunity and Infection" International Journal of Molecular Sciences 18, no. 10: 2197. https://doi.org/10.3390/ijms18102197
APA StyleSubramanian Vignesh, K., & Deepe Jr., G. S. (2017). Metallothioneins: Emerging Modulators in Immunity and Infection. International Journal of Molecular Sciences, 18(10), 2197. https://doi.org/10.3390/ijms18102197