
Major Ampullate Spider Silk with Indistinguishable Spidroin Dope Conformations Leads to Different Fiber Molecular Structures


Abstract
:
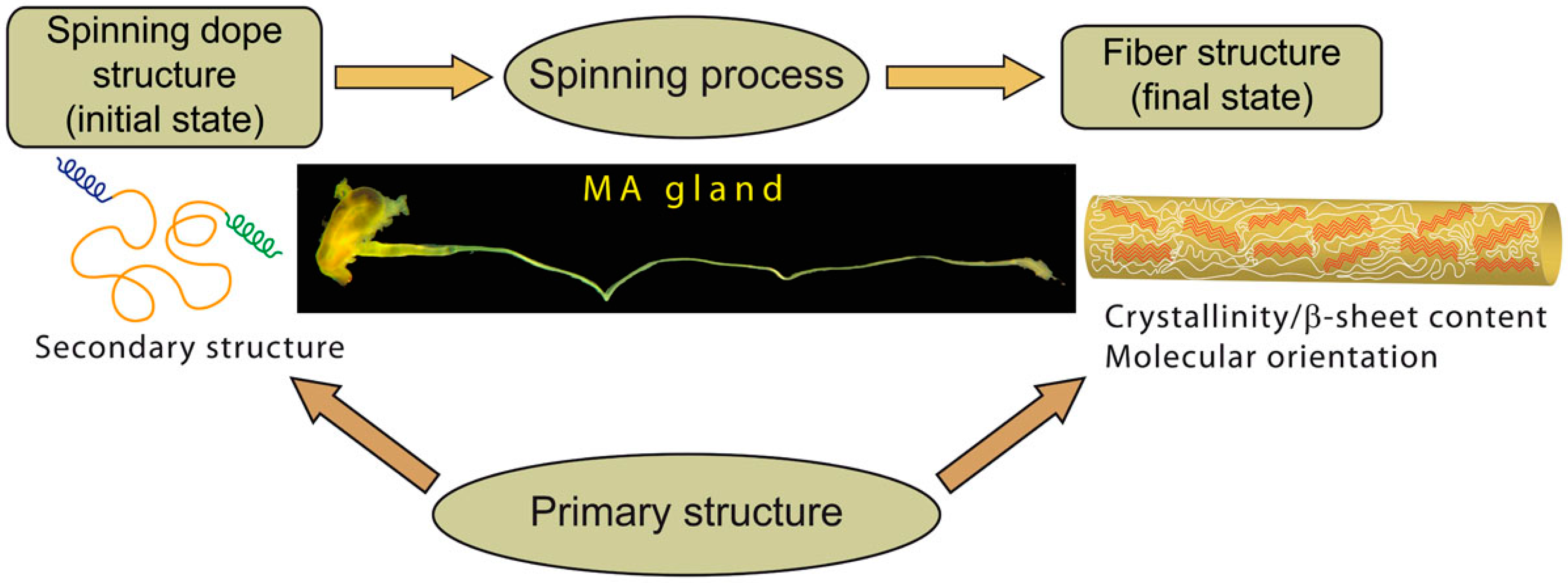
1. Introduction
2. Results
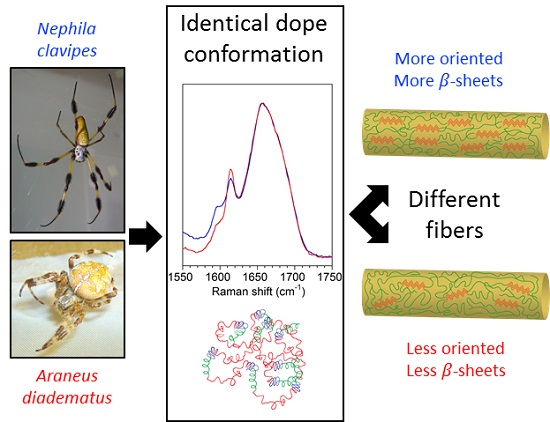
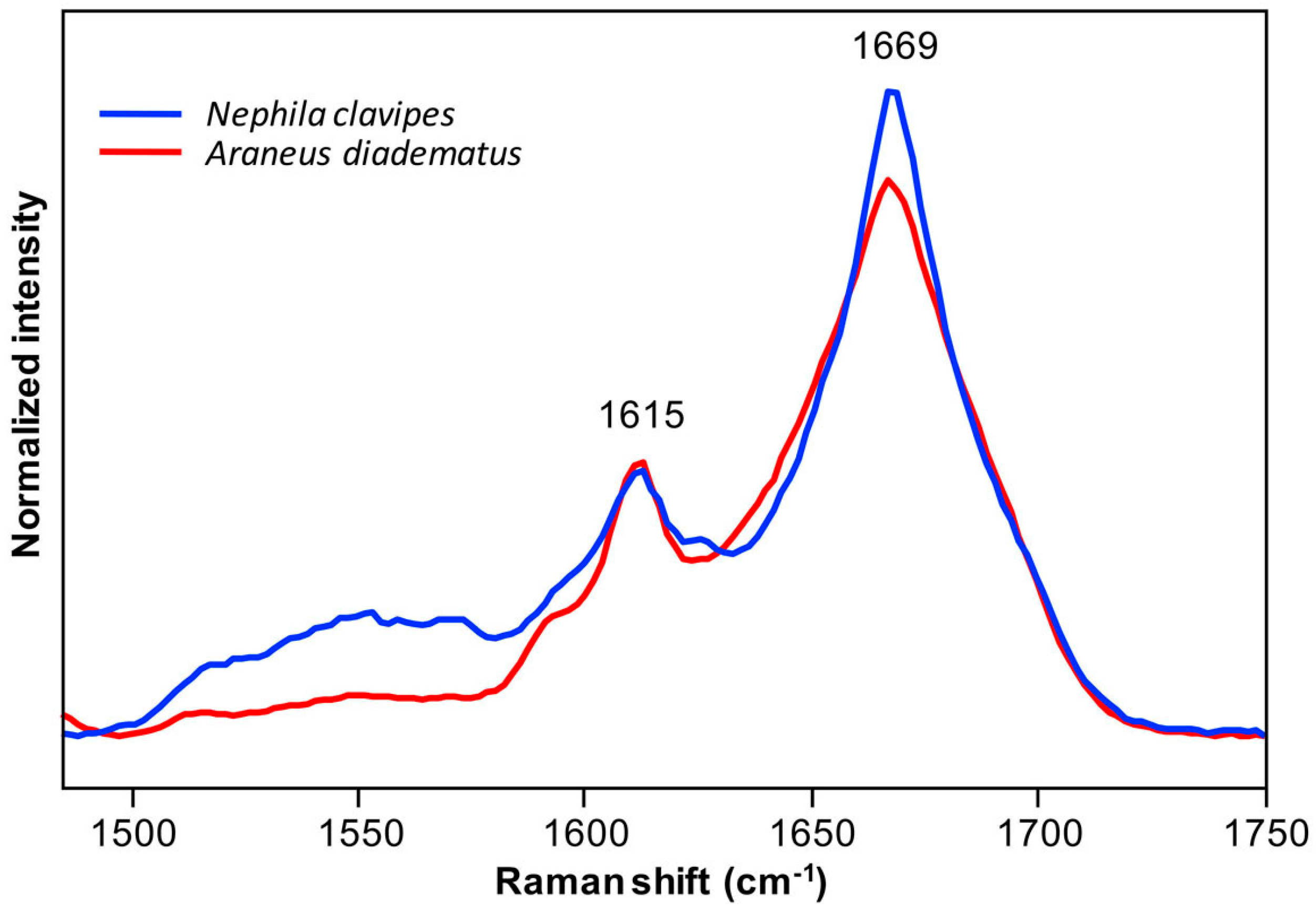
2.1. Spidroins Conformation in the Major Ampullate (MA) Dope
2.2. Molecular Structure of the MA Fibers
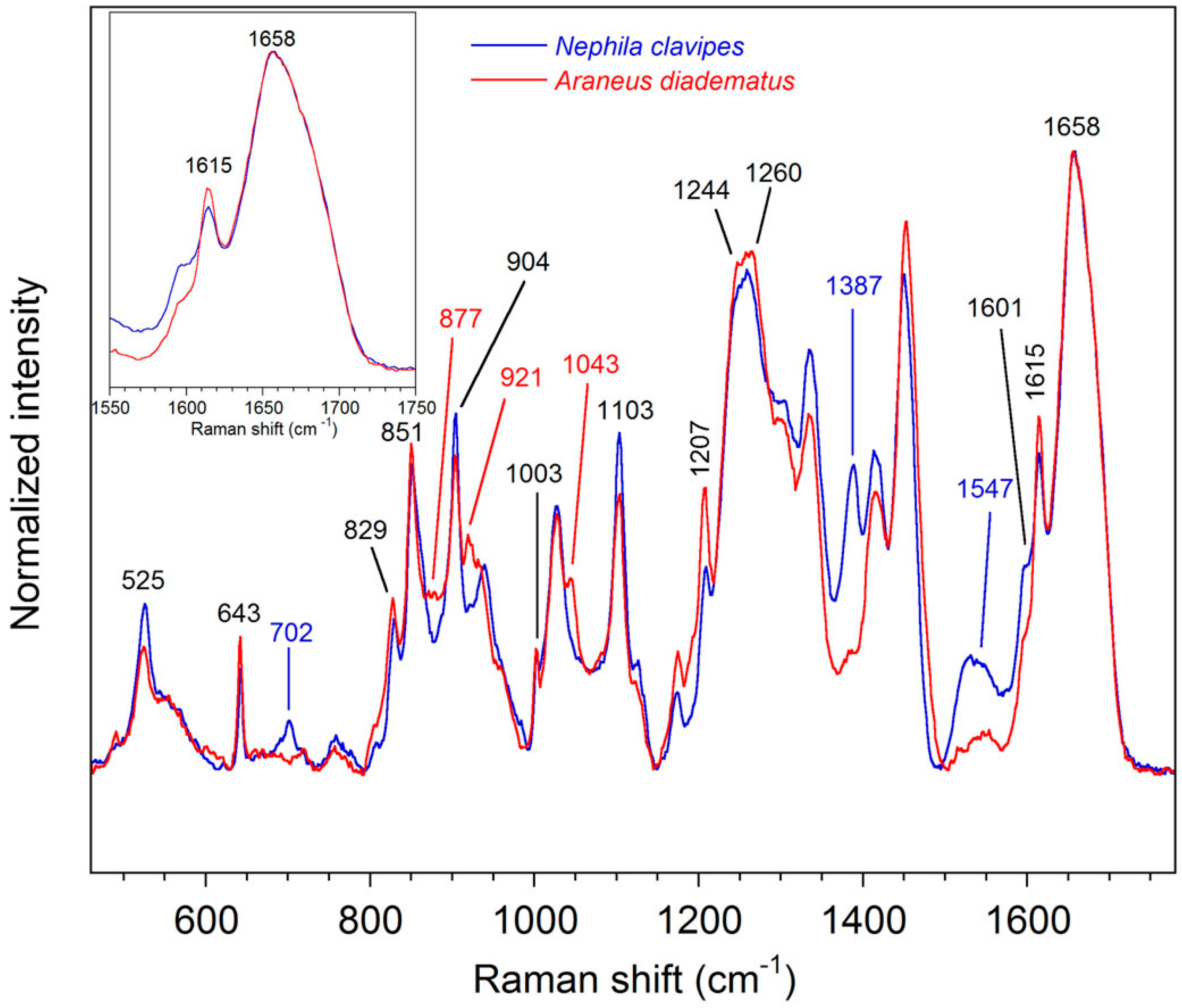
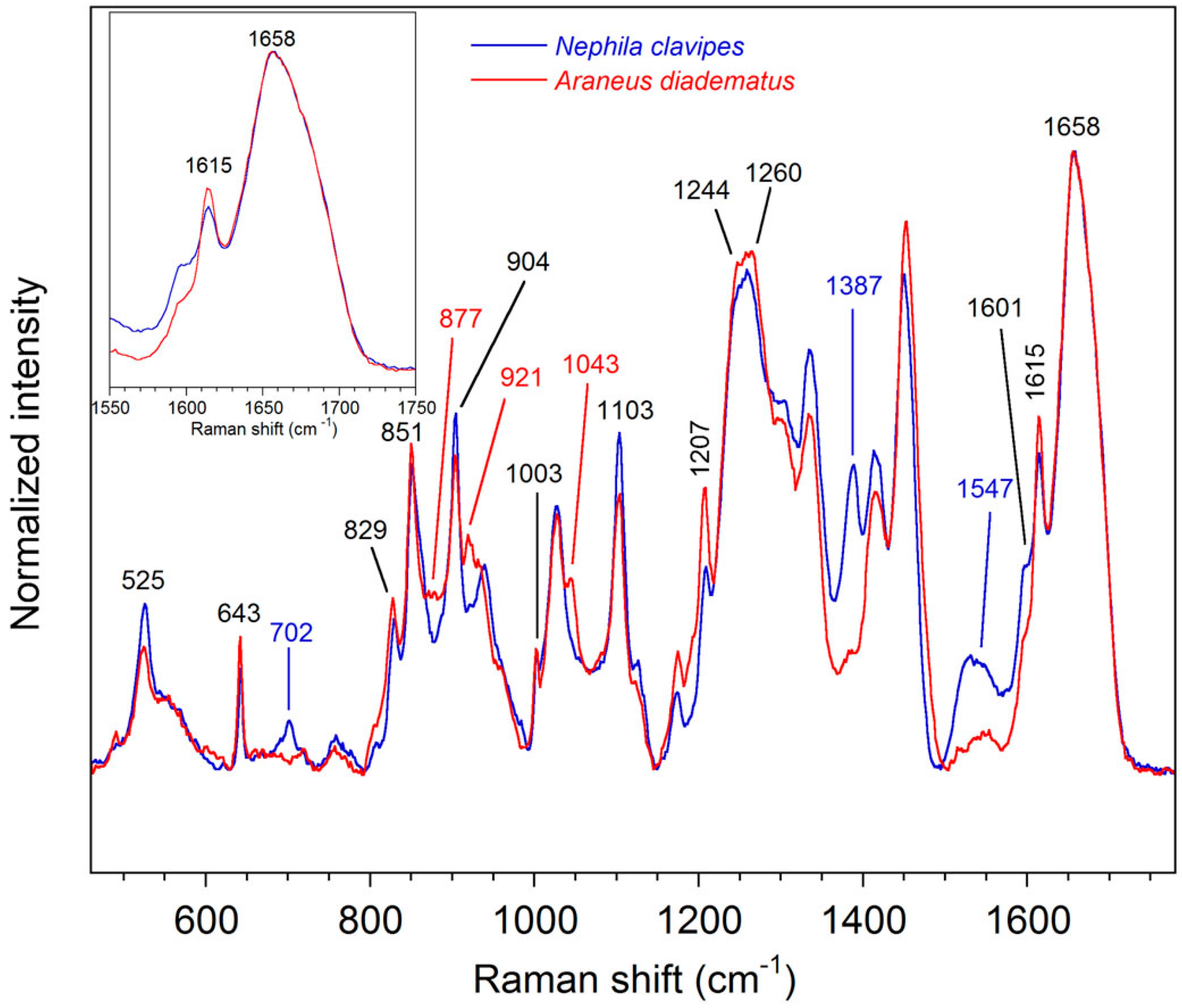
2.2.1. Qualitative Analysis
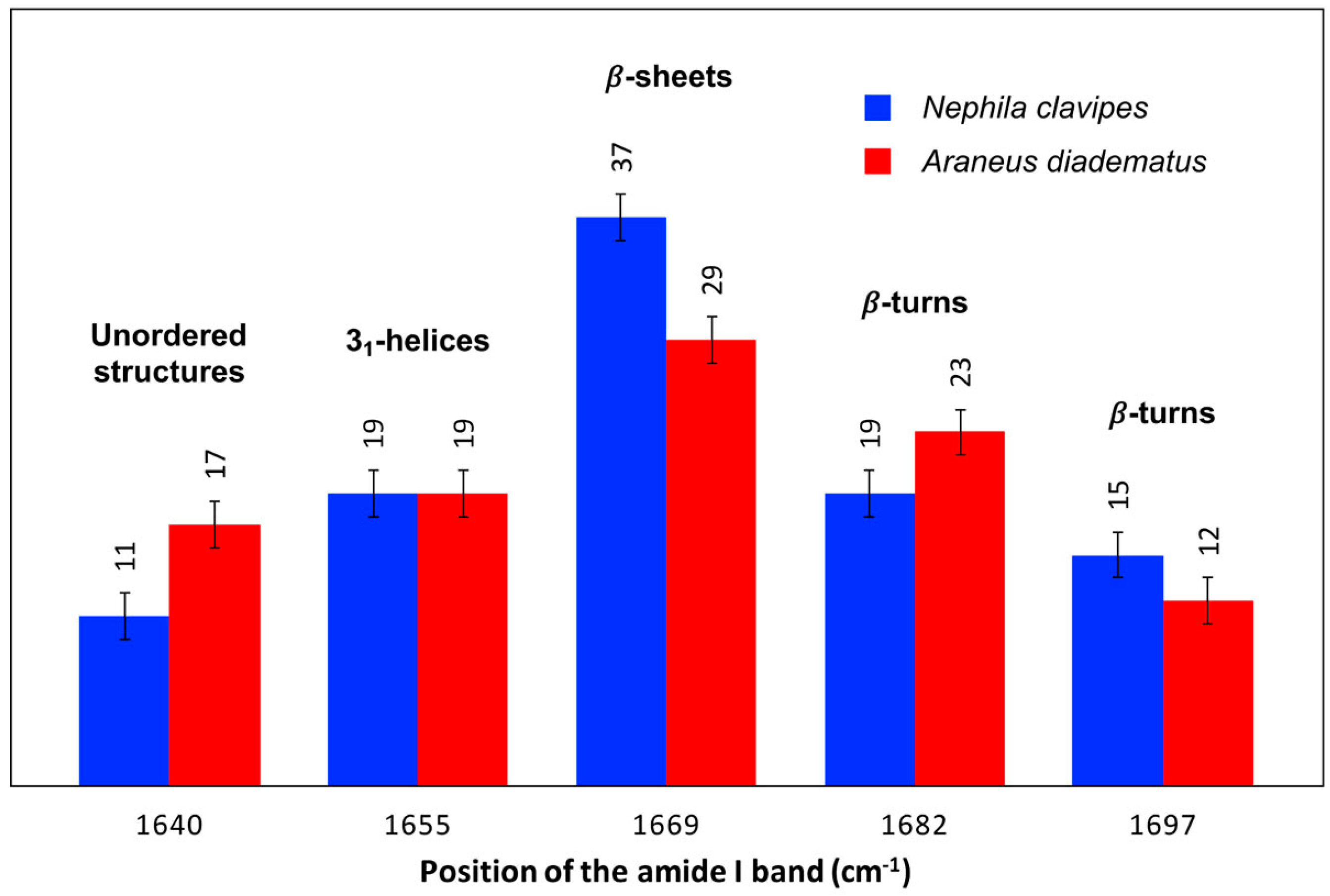
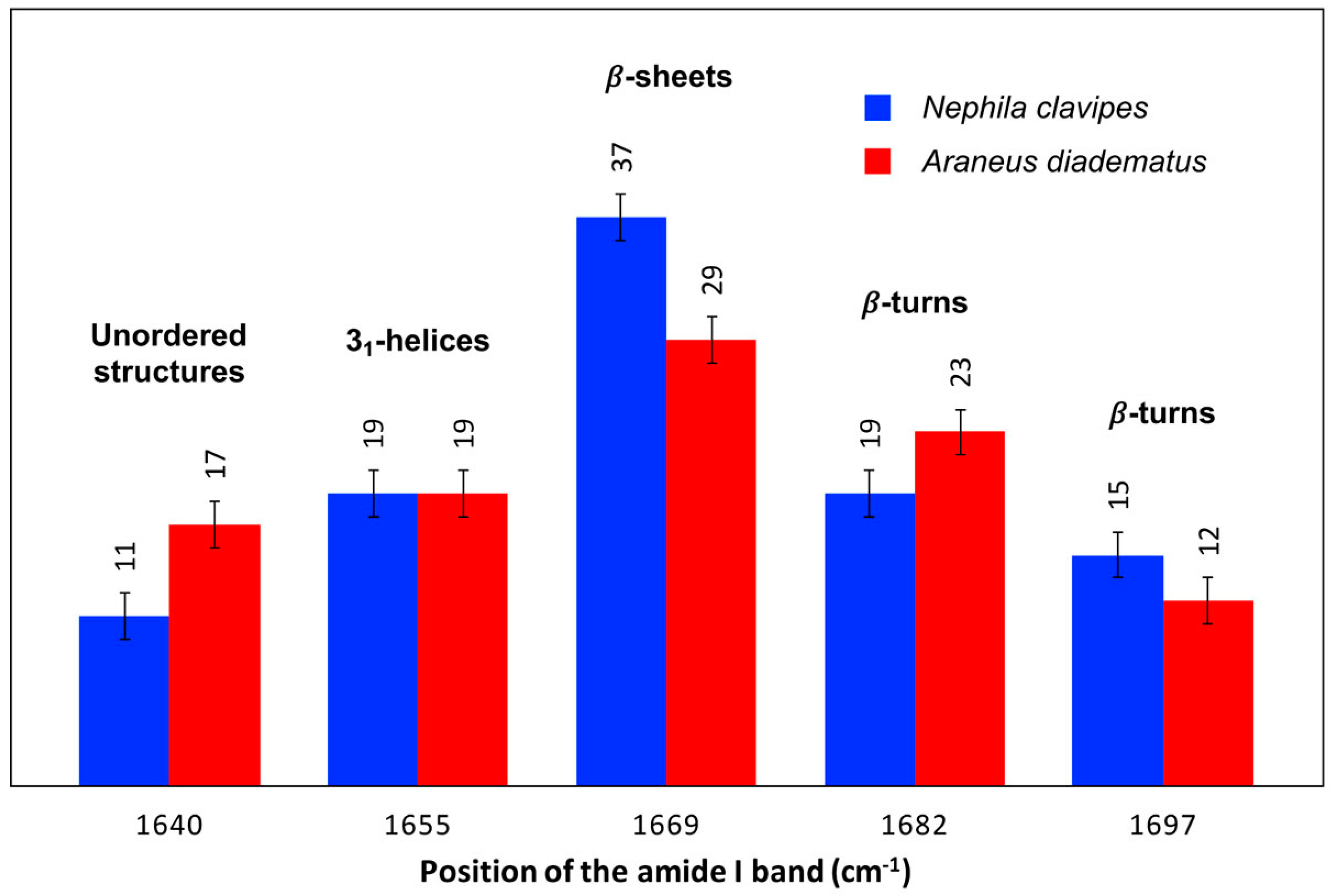
2.2.2. Secondary Structure Content
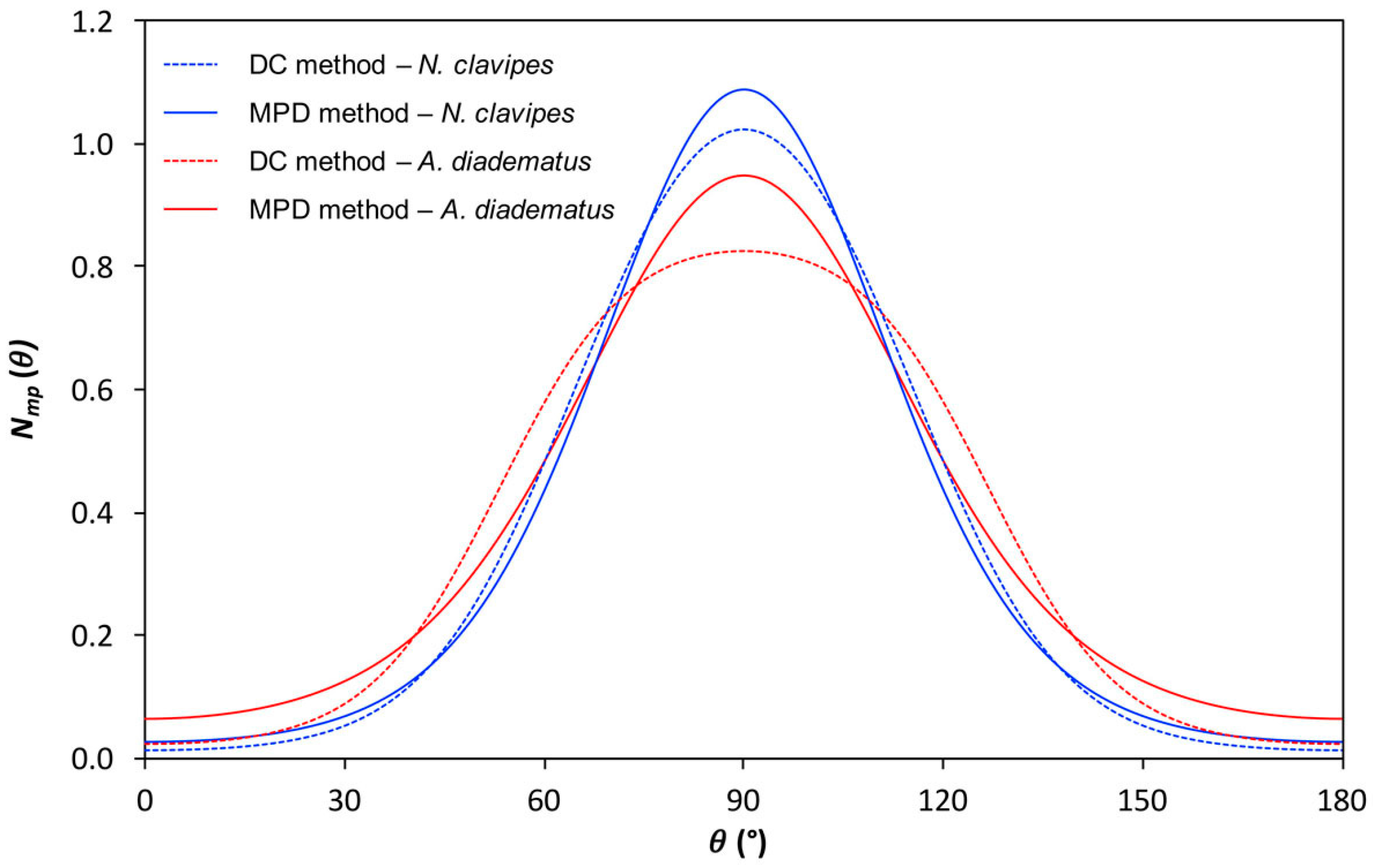
2.2.3. Quantitative Orientation Analysis
2.3. Sequences Analysis
3. Discussion
3.1. The Conformation of the Spidroins in the Dope Is Not Critical for the Fiber Structure
3.2. Molecular Orientation and β-Sheet Content Influence Silk Mechanical Properties
4. Materials and Methods
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hardy, J.G.; Römer, L.M.; Scheibel, T.R. Polymeric materials based on silk proteins. Polymer 2008, 49, 4309–4327. [Google Scholar] [CrossRef]
- Kundu, B.; Kurland, N.E.; Bano, S.; Patra, C.; Engel, F.B.; Yadavalli, V.K.; Kundu, S.C. Silk proteins for biomedical applications: Bioengineering perspectives. Prog. Polym. Sci. 2014, 39, 251–267. [Google Scholar] [CrossRef]
- Lefèvre, T.; Auger, M. Spider silk as a blueprint for greener materials: A review. Int. Mater. Rev. 2016, 61, 127–153. [Google Scholar] [CrossRef]
- Lefèvre, T.; Auger, M. Spider silk inspired materials and sustainability: Perspective. Mater. Technol. 2016, 31, 384–399. [Google Scholar] [CrossRef]
- Omenetto, F.G.; Kaplan, D.L. New opportunities for an ancient material. Science 2010, 329, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Warwicker, J.O. Comparative studies of fibroins. II. The crystal structures of various fibroins. J. Mol. Biol. 1960, 2, 350–362. [Google Scholar] [CrossRef]
- Grubb, D.T.; Jelinski, L.W. Fiber morphology of spider silk: The effects of tensile deformation. Macromolecules 1997, 30, 2860–2867. [Google Scholar] [CrossRef]
- Riekel, C.; Bräden, C.; Craig, C.L.; Ferrero, C.; Heidelbach, F.; Müller, M. Aspects of X-ray diffraction on single spider fibers. Int. J. Biol. Macromol. 1999, 24, 179–186. [Google Scholar] [CrossRef]
- Termonia, Y. Molecular modeling of spider silk elasticity. Macromolecules 1994, 27, 7378–7381. [Google Scholar] [CrossRef]
- Porter, D.; Vollrath, F.; Shao, Z. Predicting the mechanical properties of spider silk as a model nanostructured polymer. Eur. Phys. J. E 2005, 16, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Keten, S.; Buehler, M.J. Geometric confinement governs the rupture strength of H-bond assemblies at a critical length scale. Nanoletters 2008, 8, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Keten, S.; Xu, Z.; Ihle, B.; Buehler, M.J. Nanoconfinement controls stiffness, strength and mechanical toughness of β-sheet crystals in silk. Nat. Mater. 2010, 9, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Savage, K.N.; Gosline, J.M. The effect of proline on the network structure of major ampullate silks as inferred from their mechanical and optical properties. J. Exp. Biol. 2008, 211, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.P.; Fahnestock, S.R.; Yves, T.; Gardner, K.H. Nylons from nature: Synthetic analogs to spider silk. Adv. Mater. 1998, 10, 1185–1195. [Google Scholar] [CrossRef]
- Simmons, A.; Ray, E.; Jelinski, L.W. Solid-State 13C NMR of Nephila clavipes dragline silk establishes structure and identity of crystalline regions. Macromolecules 1994, 27, 5235–5237. [Google Scholar] [CrossRef]
- Lefèvre, T.; Paquet-Mercier, F.; Rioux-Dubé, J.-F.; Pézolet, M. Structure of silk by Raman spectromicroscopy: From the spinning glands to the fibers. Biopolymers 2012, 97, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Kummerlen, J.; van Beek, J.D.; Vollrath, F.; Meier, B.H. Local structure in spider dragline silk investigated by two-dimensional spin-diffusion nuclear magnetic resonance. Macromolecules 1996, 29, 2920–2928. [Google Scholar] [CrossRef]
- Van Beek, J.D.; Hess, S.; Vollrath, F.; Meier, B.H. The molecular structure of spider dragline silk: Folding and orientation of the protein backbone. Proc. Natl. Acad. Sci. USA 2002, 99, 10266–10271. [Google Scholar] [CrossRef] [PubMed]
- Valluzzi, R.; Szela, S.; Avtges, P.; Kirschner, D.; Kaplan, D. Methionine redox-controlled crystallization of biosynthetic silk spidroin. J. Phys. Chem. B 1999, 103, 11382–11392. [Google Scholar] [CrossRef]
- Eles, P.T.; Michal, C.A. A DECODER NMR study of backbone orientation in Nephila clavipes dragline silk under varying strain and draw rate. Biomacromolecules 2004, 5, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lewis, R.V. Structure of a protein superfiber—Spider dragline silk. Proc. Natl. Acad. Sci. USA 1990, 87, 7120–7124. [Google Scholar] [CrossRef] [PubMed]
- Hinnan, M.B.; Lewis, R.V. Isolation of a clone encoding a second dragline silk fibroin—Nephila clavipes dragline silk is a two-protein fiber. J. Biol. Chem. 1992, 267, 19320–19324. [Google Scholar]
- Guerette, P.A.; Ginzinger, D.G.; Weber, B.H.F.; Gosline, J.M. Silk properties determined by gland-specific expression of a spider fibroin gene family. Science 1996, 272, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.O. Amino acid composition of spider silks. Comp. Biochem. Physiol. 1970, 35, 705–711. [Google Scholar] [CrossRef]
- Work, R.W.; Young, C.T. The amino acid compositions of major and minor ampullate silks of certain orb-web-building spiders (Araneae, Araneidae). J. Arachnol. 1987, 15, 65–80. [Google Scholar]
- Hayashi, C.Y.; Shipley, N.H.; Lewis, R.V. Hypotheses that correlate the sequence, structure, and mechanical properties of spider silk proteins. Int. J. Biol. Macromol. 1999, 24, 271–275. [Google Scholar] [CrossRef]
- Simmons, A.H.; Michal, C.A.; Jelinski, L.W. Molecular orientation and two-component nature of the crystalline fraction of spider dragline silk. Science 1996, 271, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Hinman, M.B.; Jones, J.A.; Lewis, R.V. Synthetic spider silk: A modular fiber. Trends Biotechnol. 2000, 18, 374–379. [Google Scholar] [CrossRef]
- Tokareva, O.; Jacobsen, M.; Buehler, M.; Wong, J.; Kaplan, D.L. Structure-function-property-design interplay in biopolymers: Spider silk. Acta Biomater. 2014, 10, 1612–1626. [Google Scholar] [CrossRef] [PubMed]
- Askarieh, G.; Hedhammar, M.; Nordling, K.; Saenz, A.; Casals, C.; Rising, A.; Johansson, J.; Knight, S.D. Self-assembly of spider silk proteins is controlled by a pH-sensitive relay. Nature 2010, 465, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Hagn, F.; Eisoldt, L.; Hardy, J.G.; Vendrely, C.; Coles, M.; Scheibel, T.; Kessler, H. A conserved spider silk domain acts as a molecular switch that controls fibre assembly. Nature 2010, 465, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.; Terry, A.E.; Porter, D.; Vollrath, F. Comparing the rheology of native spider and silkwormspinning dope. Nat. Mater. 2006, 5, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Kojić, N.; Bico, J.; Clasen, C.; McKinley, G.H. Ex vivo rheology of spiders. J. Exp. Biol. 2006, 209, 4355–4362. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.P.; Vollrath, F. Liquid crystals and flow elongation in a spider’s silk production line. Proc. R. Soc. Lond. B 1999, 266, 519–523. [Google Scholar] [CrossRef]
- Vollrath, F.; Knight, D.P. Structure and function of the silk production pathway in the spider Nephila edulis. Int. J. Biol. Macromol. 1999, 24, 243–249. [Google Scholar] [CrossRef]
- Vollrath, F.; Knight, D.P.; Hu, X.W. Silk production in a spider involves acid bath treatment. Proc. R. Soc. Lond. B 1998, 265, 817–820. [Google Scholar] [CrossRef]
- Knight, D.P.; Vollrath, F. Changes in element composition along the spinning duct in a Nephila spider. Naturwissenschaften 2001, 88, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Chen, G.; Otikovs, M.; Landreh, M.; Nordling, K.; Kronqvist, N.; Westermark, P.; Joörnvall, H.; Knight, S.; Ridderstråle, Y.; et al. Carbonic anhydrase generates CO2 and H+ that drive spider silk formation via opposite effects on the terminal domains. PLoS Biol. 2014, 12, e1001921. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sponner, A.; Porter, D.; Vollrath, F. Proline and processing of spider silks. Biomacromolecules 2008, 9, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Vollrath, F.; Porter, D. Spider silk as archetypal protein elastomer. Soft Matter 2006, 2, 377–385. [Google Scholar] [CrossRef]
- Savage, K.N.; Gosline, J.M. The role of proline in the elastic mechanism of hydrated spider silks. J. Exp. Biol. 2008, 211, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Rauscher, S.; Baud, S.; Miao, M.; Keeley, F.W.; Pomès, R. Proline and glycine control protein self-organization into elastomeric or amyloid fibrils. Structure 2006, 14, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, T.A.; Kuntner, M.; Marhabaie, M.; Leeper, T.C.; Agnarsson, I. Biomaterial evolution parallels behavioral innovation in the origin of orb-like spider webs. Sci. Rep. 2012, 2, 833. [Google Scholar] [CrossRef] [PubMed]
- Gatesy, J.; Hayashi, C.; Motriuk, D.; Woods, J.; Lewis, R.V. Extreme diversity, conservation, and convergence of spider silk fibroin sequences. Science 2001, 291, 2603–2605. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Vollrath, F. Surprising strength of silkworm silk. Nature 2002, 418, 741. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.-X.; Qiana, Z.-G.; Kib, C.S.; Park, Y.H.; Kaplan, D.; Lee, S.Y. Native-sized recombinant spider silk protein produced in metabolically engineered Escherichia coli results in a strong fiber. Proc. NatL. Acad. Sci. USA 2010, 107, 14059–14063. [Google Scholar] [CrossRef] [PubMed]
- Marhabaie, M.; Leeper, T.C.; Blackledge, T.A. Protein Composition Correlates with the Mechanical Properties of Spider (Argiope trifasciata) Dragline Silk. Biomacromolecules 2014, 15, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Plaza, G.R.; Corsini, P.; Marsano, E.; Perez-Rigueiro, J.; Biancotto, L.; Elices, M.; Riekel, C.; Agullo-Rueda, F.; Gallardo, E.; Calleja, J.M.; et al. Old Silks Endowed with New Properties. Macromolecules 2009, 42, 8977–8982. [Google Scholar] [CrossRef]
- Blamires, S.J.; Wu, C.L.; Blackledge, T.A.; Tso, I.M. Post-secretion processing influences spider silk performance. J. R. Soc. Interface 2012, 9, 2479–2487. [Google Scholar] [CrossRef] [PubMed]
- Denny, M. The physical properties of spider’s silk and their role in the design of orb-webs. J. Exp. Biol. 1976, 65, 483–506. [Google Scholar]
- Madurga, R.; Blackledge, T.A.; Perea, B.; Plaza, G.R.; Riekel, C.; Burghammer, M.; Elices, M.; Guinea, G.; Perez-Rigueiro, J. Persistence and variation in microstructural design during the evolution of spider silk. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Stauffer, S.L.; Coguil, S.L.; Lewis, R.V. Comparison of physical properties of three silks from Nephila clavipes and Araneus diadematus. J. Arachnol. 1994, 22, 1–5. [Google Scholar]
- Cunniff, P.M.; Fossey, S.A.; Auerbach, M.A.; Song, J.W.; Kaplan, D.L.; Adams, W.W.; Eby, R.K.; Mahoney, D.; Vezie, D.L. Mechanical and thermal properties of dragline silk from the spider Nephila clavipes. Polym. Adv. Technol. 1994, 5, 401–410. [Google Scholar] [CrossRef]
- Gosline, J.M.; Guerette, P.A.; Ortlepp, C.S.; Savage, K.N. Mechanical design of spider silk: From fibroin sequence to mechanical function. J. Exp. Biol. 1999, 202, 3295–3303. [Google Scholar] [PubMed]
- Swanson, B.O.; Blackledge, T.A.; Beltrán, J.; Hayashi, C.Y. Variation in the material properties of spider dragline silk across species. Appl. Phys. A 2006, 82, 213–218. [Google Scholar] [CrossRef]
- Swanson, B.O.; Blackledge, T.A.; Summers, A.P.; Hayashi, C.Y. Spider dragline silk: Correlated and mosaic evolution in high-performance biological materials. Evolution 2006, 60, 2539–2551. [Google Scholar] [CrossRef] [PubMed]
- Swanson, B.O.; Blackledge, T.A.; Hayashi, C.Y. Spider capture silk: Performance implications of variation in an exeptional biomaterial. J. Exp. Biol. 2007, 307A, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Vollrath, F.; Sirichaisit, J.; Young, R.J. Analysis of spider silk in native and supercontracted states using Raman spectroscopy. Polymer 1999, 40, 2493–2500. [Google Scholar] [CrossRef]
- Lefèvre, T.; Rousseau, M.-E.; Pézolet, M. Protein secondary structure and orientation in silk as revealed by Raman spectromicroscopy. Biophys. J. 2007, 92, 2885–2895. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yarger, J.L.; Holland, G.P. Exploring the backbone dynamics of native spider silk proteins in Black Widow silk glands with solution-state NMR spectroscopy. Polymer 2014, 55, 3879–3885. [Google Scholar] [CrossRef]
- Wait, S.C.; McNerney, J.C. Vibrational spectra and assignments for quinoline and isoquinoline. J. Mol. Spectrosc. 1970, 34, 56–77. [Google Scholar] [CrossRef]
- Lefèvre, T.; Boudreault, S.; Cloutier, C.; Pézolet, M. Conformational and orientational transformation of silk proteins in the major ampullate gland of Nephila clavipes spiders. Biomacromolecules 2008, 9, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, M.-E.; Lefèvre, T.; Pézolet, M. Conformation and orientation of proteins in various types of silk fibers produced by Nephila clavipes spiders. Biomacromolecules 2009, 10, 2945–2953. [Google Scholar] [CrossRef] [PubMed]
- Denny, M.W. Silks—Their properties and functions. In The Mechanical Properties of Biological Materials; Society for Experimental Biology, Ed.; Cambridge University Press: Cambridge, UK, 1980; Volume XXXIV, pp. 246–272. [Google Scholar]
- Craig, C.L. Evolution of arthropod silks. Annu. Rev. Entomol. 1997, 42, 231–267. [Google Scholar] [CrossRef] [PubMed]
- Plaza, G.R.; Perez-Rigueiro, J.; Riekel, C.; Perea, G.B.; Agullo-Rueda, F.; Burghammer, M.; Guinea, G.V.; Elices, M. Relationship between microstructure and mechanical properties in spider silk fibers: Identification of two regimes in the microstructural changes. Soft Matter 2012, 8, 6015–6026. [Google Scholar] [CrossRef]
- Richard-Lacroix, M.; Pellerin, C. Novel method for quantifying molecular orientation by polarized raman spectroscopy: A comparative simulations study. Appl. Spectrosc. 2013, 67, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Jelinski, L.W.; Blye, A.; Liivak, O.; Michal, C.; LaVerde, G.; Seidel, A.; Shah, N.; Yang, Z. Orientation, structure, wet-spinning, and molecular basis for supercontraction of spider dragline silk. Int. J. Biol. Macromol. 1999, 24, 197–201. [Google Scholar] [CrossRef]
- Lefèvre, T.; Leclerc, J.; Rioux-Dubé, J.-F.; Buffeteau, T.; Paquin, M.-C.; Rousseau, M.-E.; Cloutier, I.; Auger, M.; Gagné, S.M.; Boudreault, S.; et al. Conformation of spider silk proteins in situ in the intact major ampullate gland and in solution. Biomacromolecules 2007, 8, 2342–2344. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Vollrath, F. Thread biomechanics in the two orb-weaving spiders Araneus diadematus (Araneae, Araneidae) and Uloborus walckenaerius (Araneae, Uloboridae). J. Exp. Zool. 1995, 271, 1–17. [Google Scholar] [CrossRef]
- Sensenig, A.; Agnarsson, I.; Blackledge, T.A. Behavioural and biomaterial coevolution in spider orb webs. J. Evol. Biol. 2010, 23, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, C.Y.; Lewis, R.V. Molecular architecture and evolution of a modular spider silk protein gene. Science 2000, 287, 1477–1479. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, C.Y.; Lewis, R.V. Evidence from flagelliform silk cDNA for the structural basis of elasticity and modular nature of spider silks. J. Biol. Mol. 1998, 275, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, T.; Pézolet, M. Unexpected β-sheets and molecular orientation in flagelliform spider silk as revealed by Raman spectromicroscopy. Soft Matter 2012, 8, 6350–6357. [Google Scholar] [CrossRef]
- Lefevre, T.; Rousseau, M.E.; Pézolet, M. Orientation-insensitive spectra for Raman microspectroscopy. Appl. Spectrosc. 2006, 60, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, T.; Pellerin, C.; Pézolet, M. Characterization of molecular orientation. In Molecular Characterization and Analysis of Polymers; Chalmers, J., Meier, R.J., Eds.; Elsevier: Amsterdam, The Newtherlands, 2008; Volume 53, pp. 295–335. [Google Scholar]
- Bower, D.I. Investigation of molecular orientation distributions by polarized Raman scattering and polarized fluorescence. J. Polym. Sci. 1972, 10, 2135–2153. [Google Scholar] [CrossRef]
- Turrell, G. Analysis of polarization measurements in Raman microspectroscopy. J. Raman Spectrosc. 1984, 15, 103–108. [Google Scholar] [CrossRef]
- Turrell, G. Raman sampling. In Practical Raman Spectroscopy; Gardiner, D.J., Graves, P.R., Eds.; Spriner-Verlag: Berlin, Germany, 1989; pp. 13–54. [Google Scholar]
- Rousseau, M.-E.; Lefèvre, T.; Beaulieu, L.; Asakura, T.; Pézolet, M. Study of protein conformation and orientation in silkworm and spider silk fibers using Raman microspectroscopy. Biomacromolecules 2004, 5, 2247–2257. [Google Scholar] [CrossRef] [PubMed]
- Lagugné-Labarthet, F.; Buffeteau, T.; Sourisseau, C. Orientation distribution functions in uniaxial systems centered perpendicularly to a constraint direction. Appl. Spectrosc. 2000, 54, 699–705. [Google Scholar] [CrossRef]
- Pottel, H.; Herreman, W.; ven der Meer, B.W.; Ameloot, M. On the significance of the fourth-rank orientational order parameter of fluorophores in membranes. Chem. Phys. 1986, 102, 37–44. [Google Scholar] [CrossRef]
- Berne, B.J.; Pechukas, P.; Harp, G.D. Molecular reorientation in liquids and gases. J. Chem. Phys. 1968, 49, 3125–3129. [Google Scholar] [CrossRef]
- Bower, D.I. Orientation distribution functions for uniaxially oriented polymers. J. Polym. Sci. 1981, 19, 93–107. [Google Scholar] [CrossRef]
- Elices, M.; Plaza, G.R.; Arnedo, M.A.; Pérez-Rigueiro, J.; Torres, F.G.; Guinea, G.V. Mechanical behavior of silk during the evolution of orb-web spinning spiders. Biomacromolecules 2009, 10, 1904–1910. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | R′ * | P2 * | P4 * | ||
|---|---|---|---|---|---|
| DC Method | MPD Method | DC Method | MPD Method | ||
| N. clavipes | −1.63 ± 0.008 | −0.306 ± 0.005 | −0.307 ± 0.005 | 0.075 ± 0.005 | 0.089 ± 0.004 |
| A. diadematus | −1.24 ± 0.008 | −0.256 ± 0.007 | −0.258 ± 0.006 | 0.02 ± 0.02 | 0.059 ± 0.003 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dionne, J.; Lefèvre, T.; Auger, M. Major Ampullate Spider Silk with Indistinguishable Spidroin Dope Conformations Leads to Different Fiber Molecular Structures. Int. J. Mol. Sci. 2016, 17, 1353. https://doi.org/10.3390/ijms17081353
Dionne J, Lefèvre T, Auger M. Major Ampullate Spider Silk with Indistinguishable Spidroin Dope Conformations Leads to Different Fiber Molecular Structures. International Journal of Molecular Sciences. 2016; 17(8):1353. https://doi.org/10.3390/ijms17081353
Chicago/Turabian StyleDionne, Justine, Thierry Lefèvre, and Michèle Auger. 2016. "Major Ampullate Spider Silk with Indistinguishable Spidroin Dope Conformations Leads to Different Fiber Molecular Structures" International Journal of Molecular Sciences 17, no. 8: 1353. https://doi.org/10.3390/ijms17081353
APA StyleDionne, J., Lefèvre, T., & Auger, M. (2016). Major Ampullate Spider Silk with Indistinguishable Spidroin Dope Conformations Leads to Different Fiber Molecular Structures. International Journal of Molecular Sciences, 17(8), 1353. https://doi.org/10.3390/ijms17081353