
A Female-Biased Odorant Receptor from Apolygus lucorum (Meyer-Dür) Tuned to Some Plant Odors

Abstract
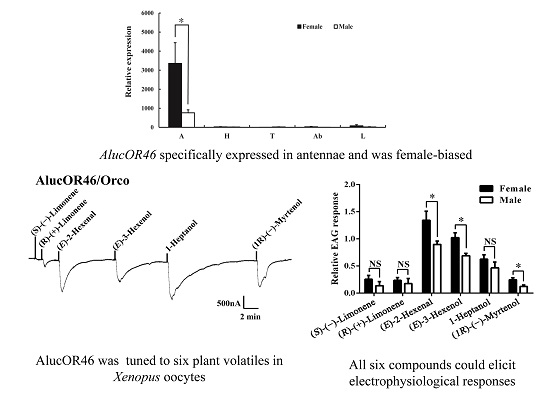
:
1. Introduction
2. Results
2.1. Gene Cloning and Sequence Analysis
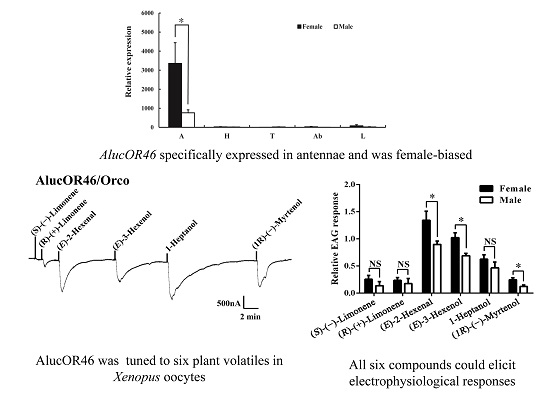
2.2. Tissue Expression Pattern of AlucOR46
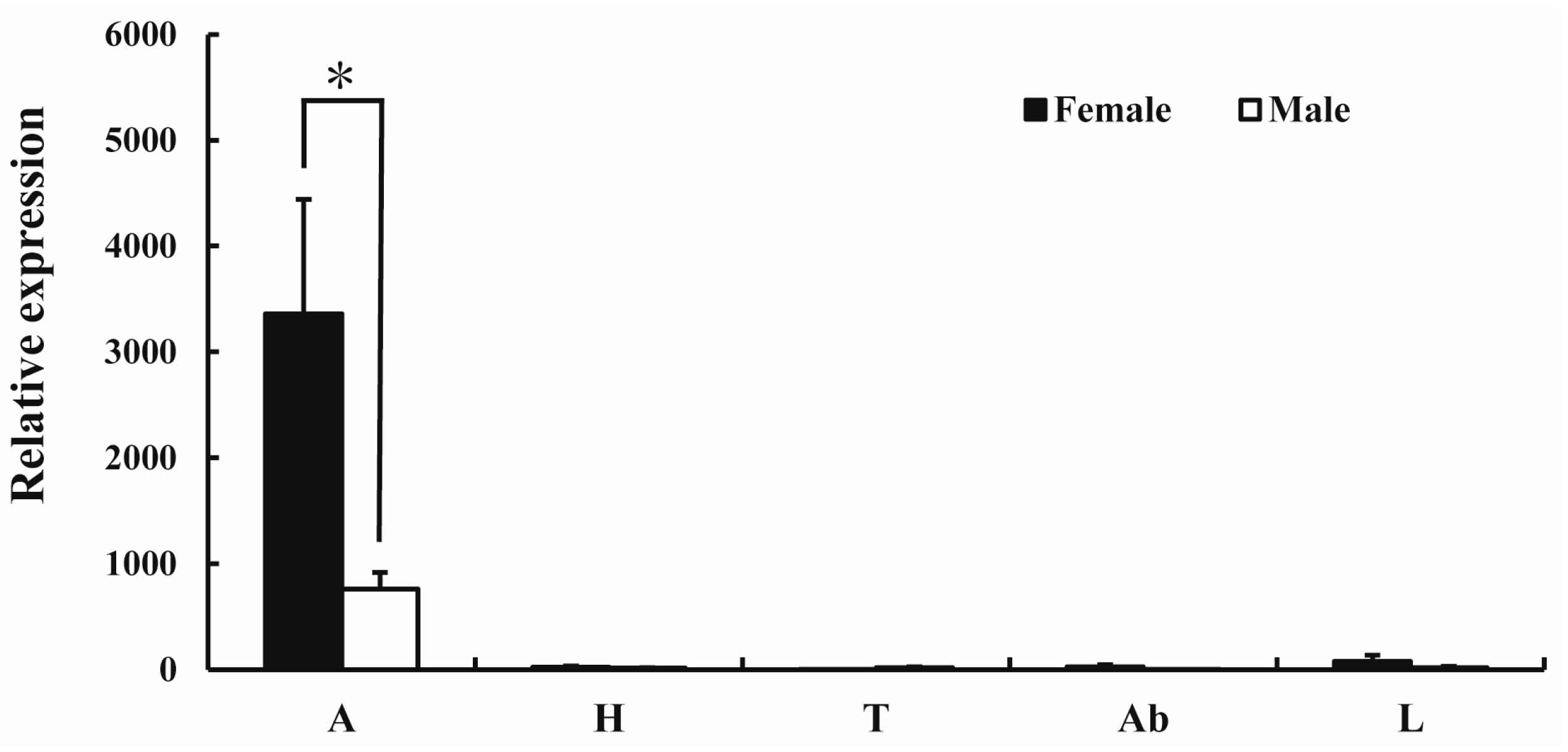
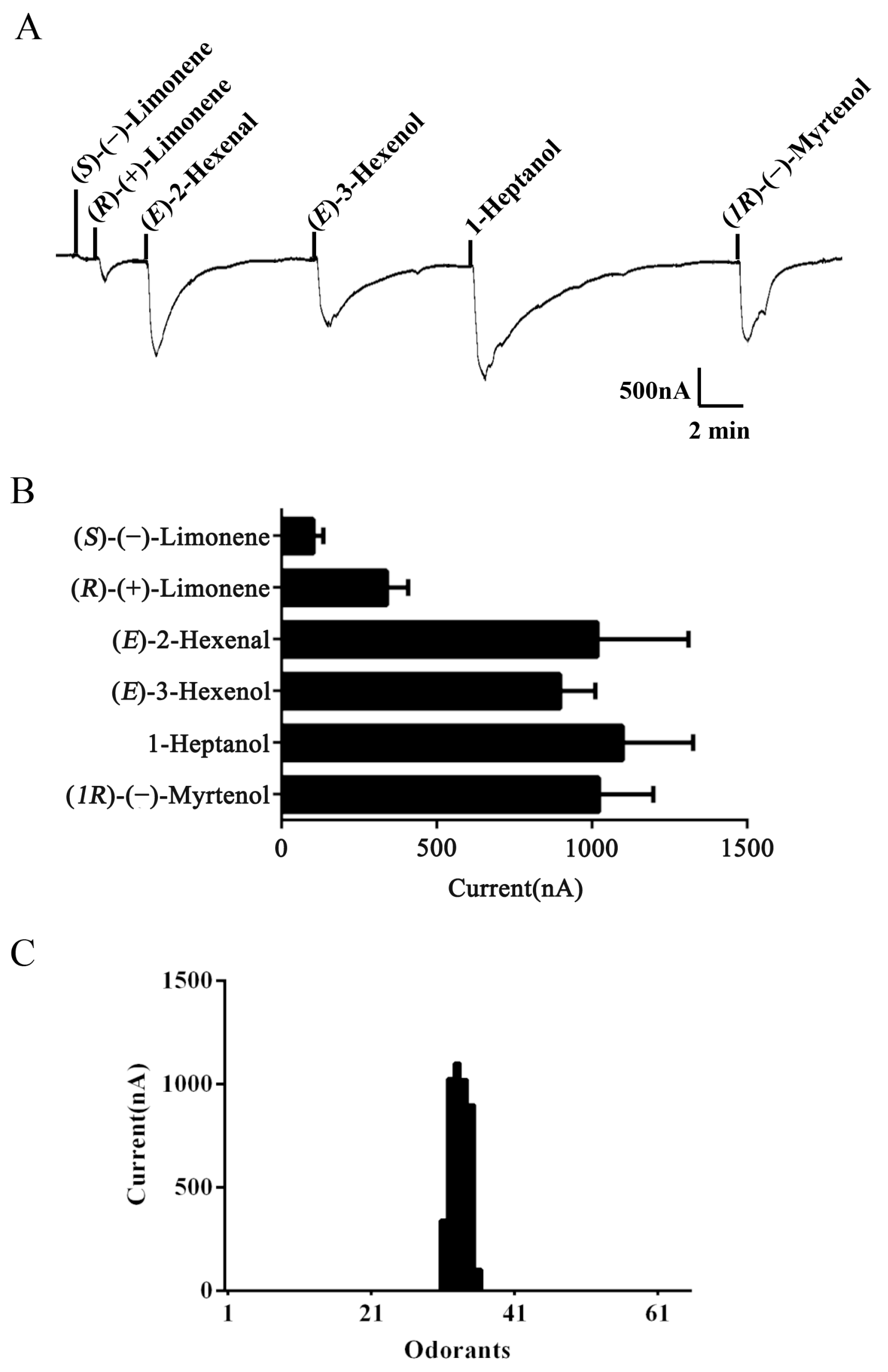
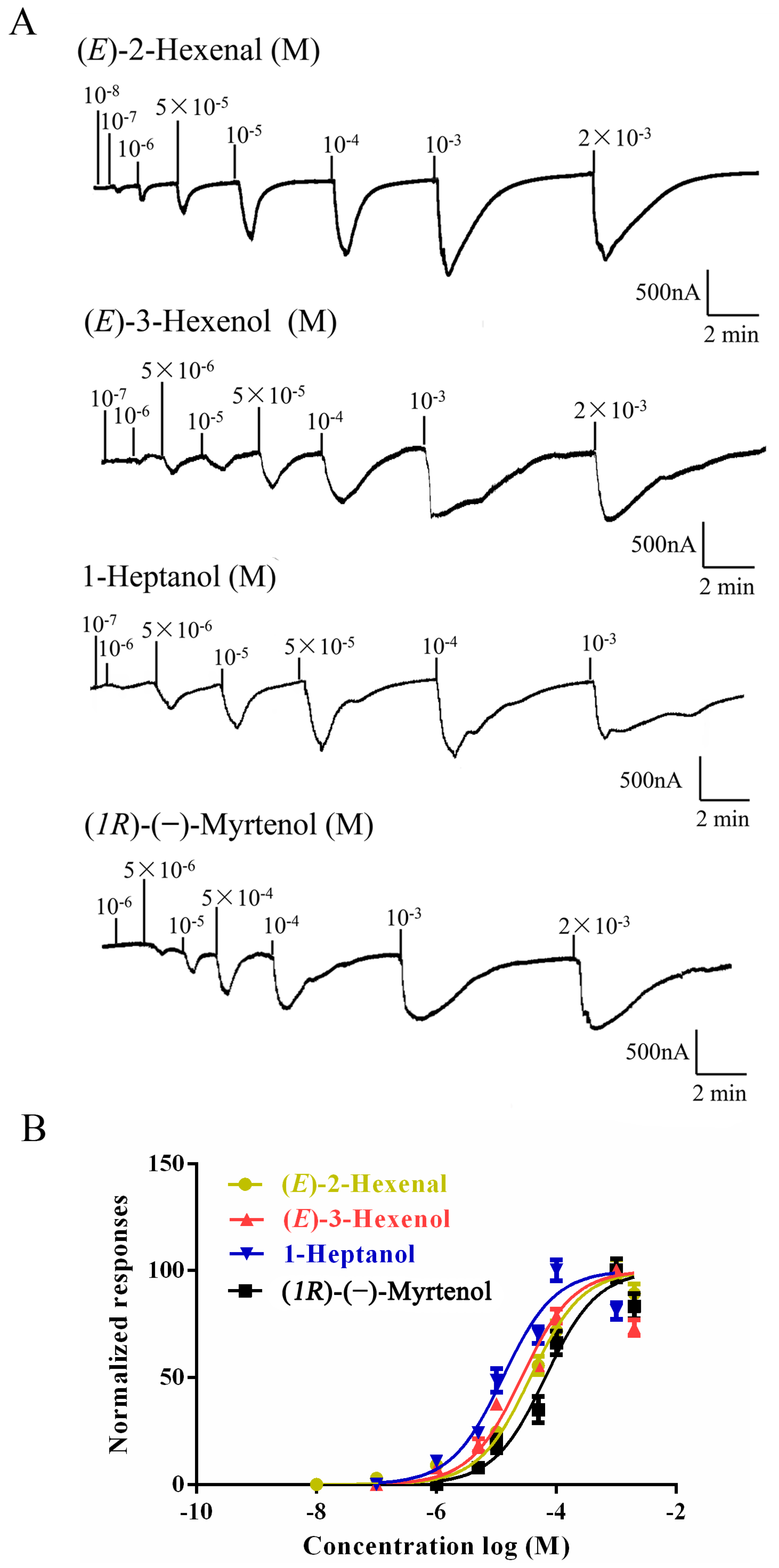
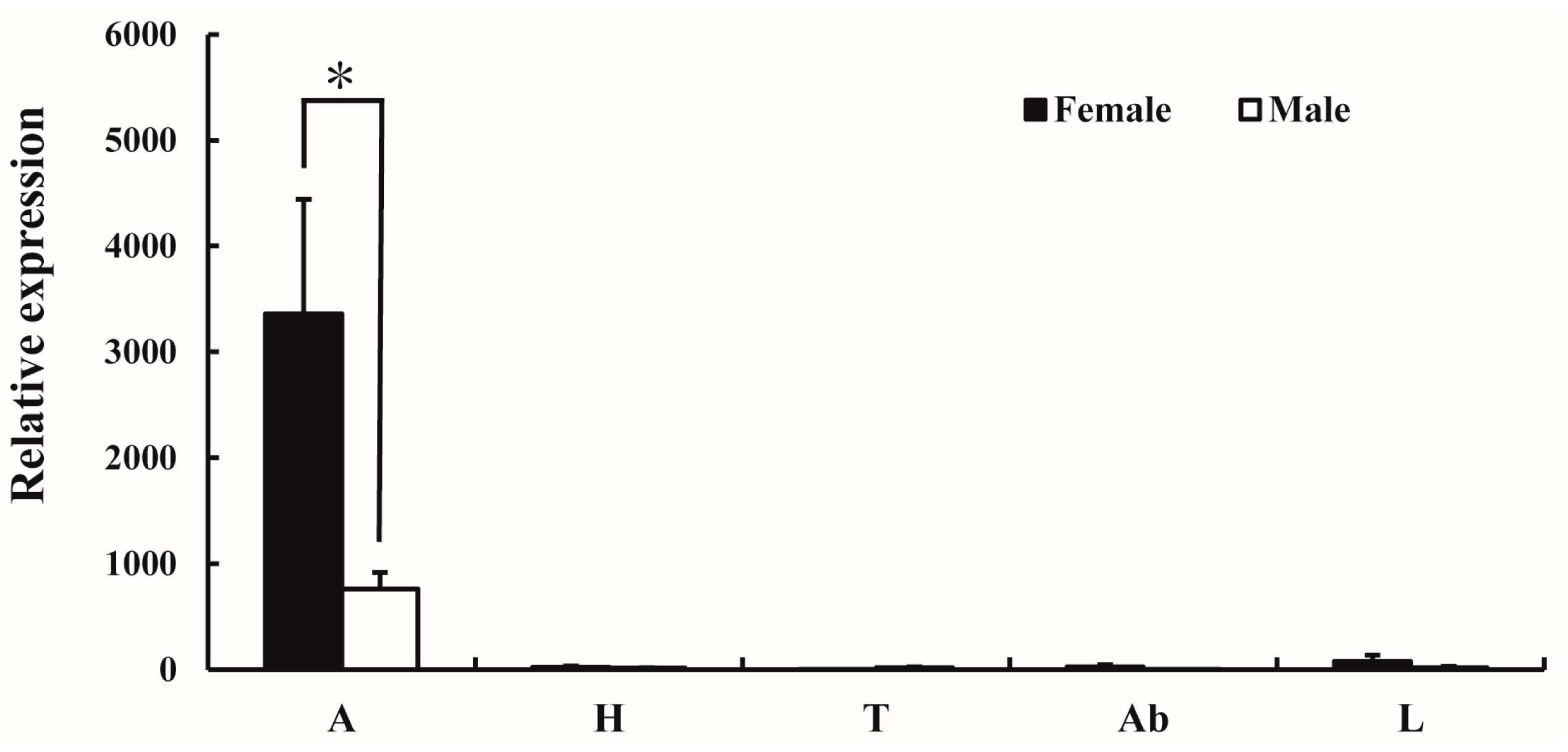
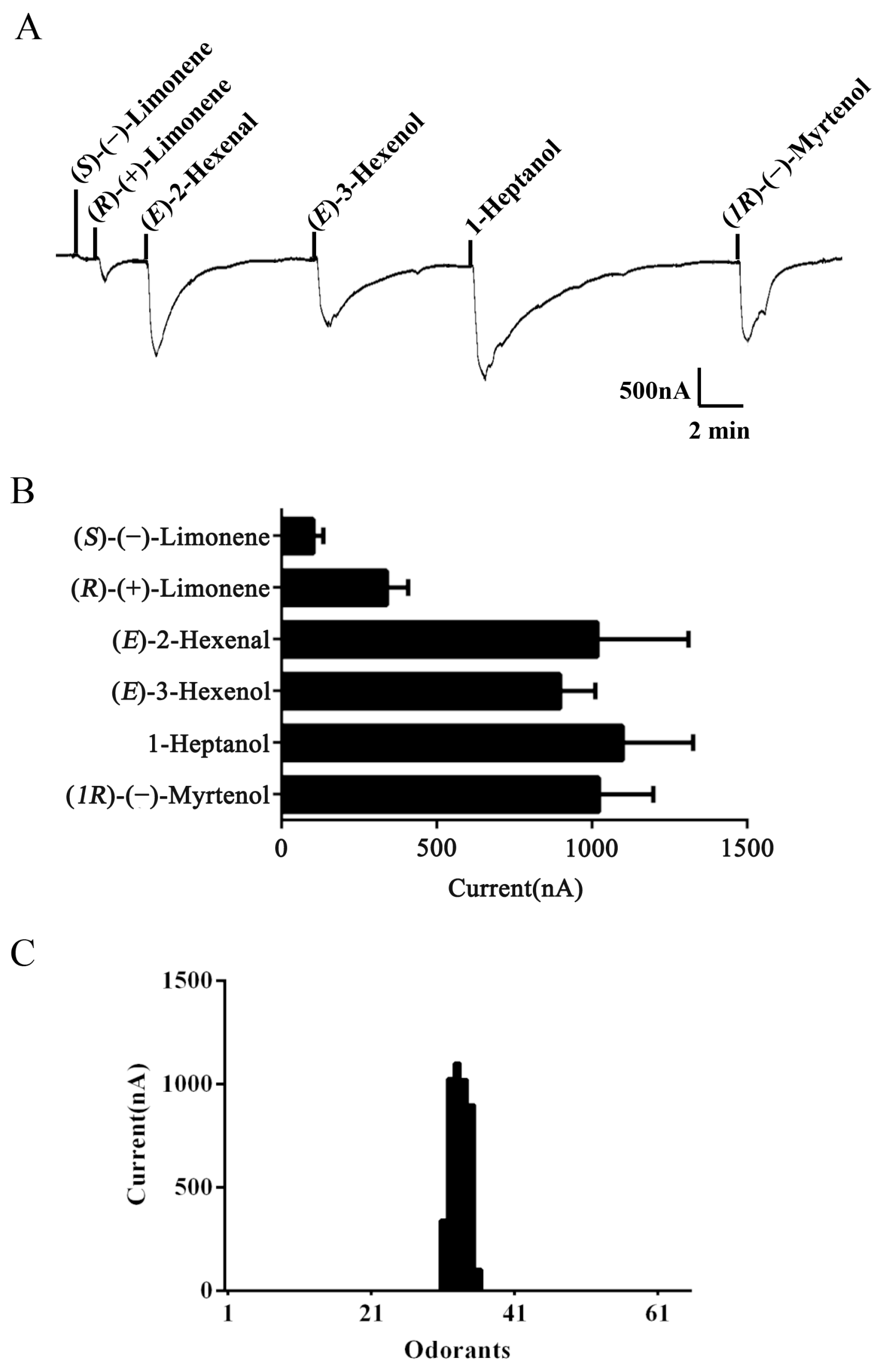
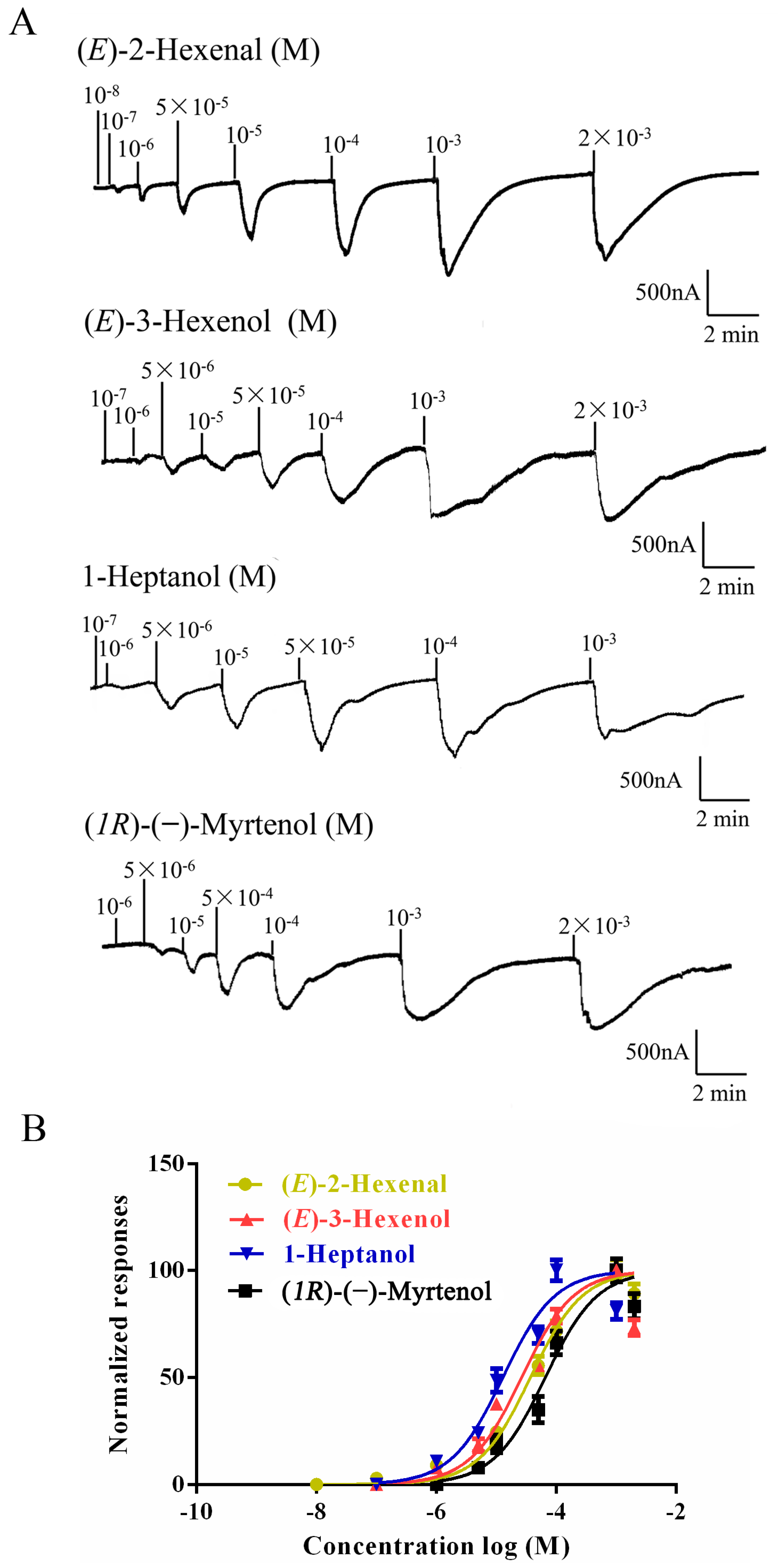
2.3. Functional Characterization of AlucOR46
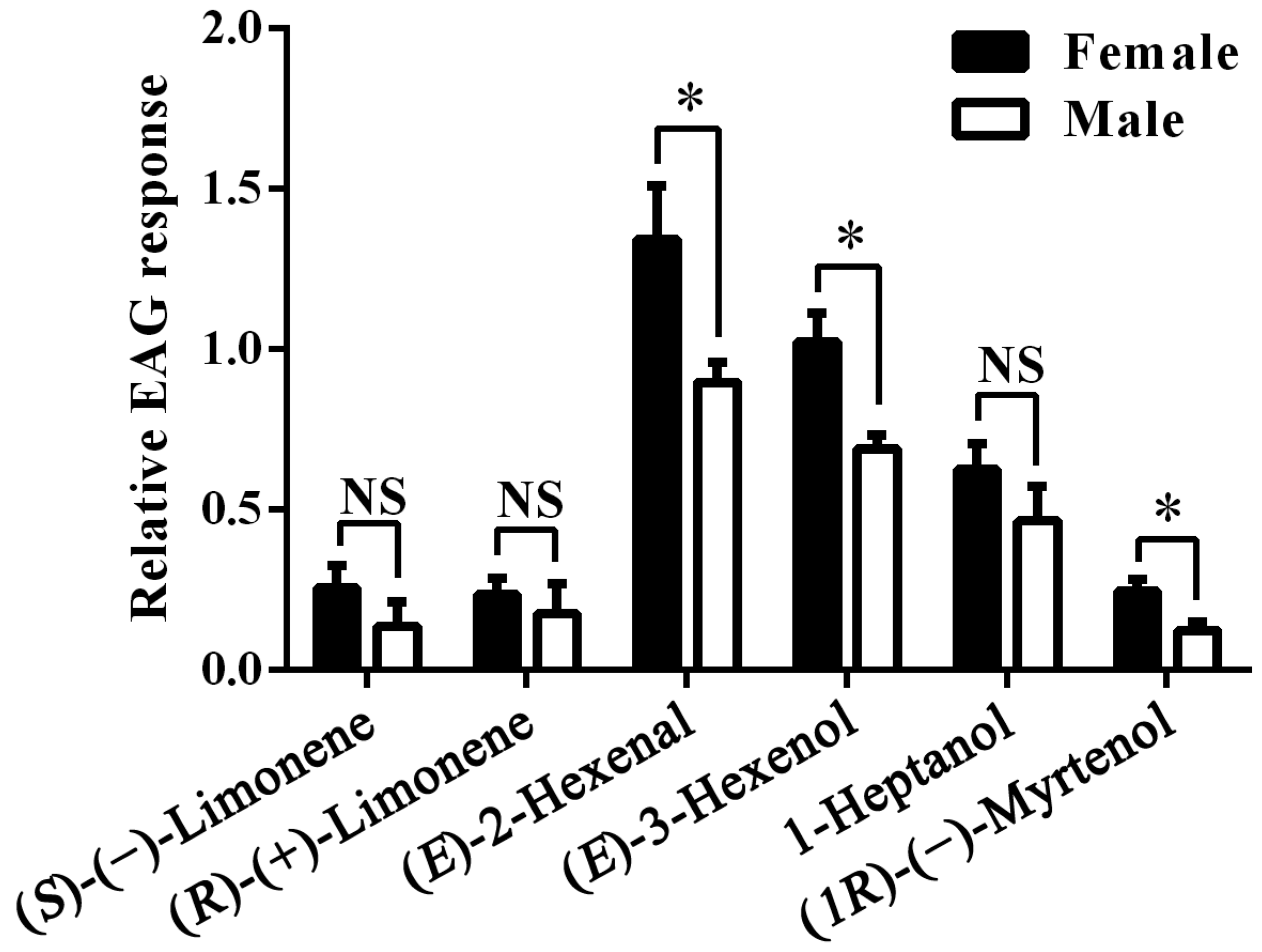
2.4. Electrophysiological Responses of A. lucorum Antennae
3. Discussion
4. Materials and Methods
4.1. Insect Rearing and Tissue Collection
4.2. Plant Volatile Compounds
4.3. RNA Isolation and cDNA Synthesis
4.4. Gene Cloning and Sequence Analysis
4.5. Quantitative Real-Time PCR
4.6. Vector Construction and cRNAs Synthesis
4.7. Two-Electrode Voltage Clamp Electrophysiological Recordings
4.8. Electroantennogram Experiment
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lu, Y.H.; Wu, K.M. Biology and Control of Cotton Mirids; Golden Shield Press: Beijing, China, 2008. [Google Scholar]
- Lu, Y.H.; Wu, K.M.; Wyckhuys, K.A.; Guo, Y.Y. Temperature-dependent life history of the green plant bug, Apolygus lucorum (Meyer-Dür)(Hemiptera: Miridae). Appl. Entomol. Zool. 2010, 45, 387–393. [Google Scholar] [CrossRef]
- Lu, Y.H.; Wu, K.M.; Wyckhuys, K.A.; Guo, Y.Y. Overwintering hosts of Apolygus lucorum (Hemiptera: Miridae) in northern China. Crop Prot. 2010, 29, 1026–1033. [Google Scholar] [CrossRef]
- Pan, H.S.; Liu, B.; Lu, Y.H.; Wyckhuys, K.A. Seasonal alterations in host range and fidelity in the polyphagous mirid bug, Apolygus lucorum (Heteroptera: Miridae). PLoS ONE 2015, 10, e0117153. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Jiao, Z.B.; Wu, K.M. Early season host plants of Apolygus lucorum (Heteroptera: Miridae) in northern China. J. Econ. Entomol. 2012, 105, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Schoonhoven, L.M.; van Loon, J.J.; Dicke, M. Insect-Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Bruce, T.J.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Q.; Xiao, L.B.; Tang, Y.A.; Sun, Y.; Bai, L.X. Relative preferences of Apolygus lucorum adults for six host species and their volatiles. Chin. J. Appl. Entomol. 2012, 49, 641–647. [Google Scholar]
- Chen, Z.C.; Su, L.; Ge, F.; Su, J.W. Electroantennogram responses of the green leaf bug, Lygus lucorum Meyer-Dür (Hemiptera: Miridae), to sex pheromone analogs and plant volatiles. Acta Entomol. Sin. 2010, 53, 47–54. [Google Scholar]
- Pan, H.S.; Lu, Y.H.; Wyckhuys, K.A.; Wu, K.M. Preference of a polyphagous mirid bug, Apolygus lucorum (Meyer-Dür) for flowering host plants. PLoS ONE 2013, 8, e68980. [Google Scholar] [CrossRef] [PubMed]
- Su, C.Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, E.M.; Gisselmann, G.; Zhang, W.; Dooley, R.; Stortkuhl, K.; Hatt, H. Odorant receptor heterodimerization in the olfactory system of Drosophila melanogaster. Nat. Neurosci. 2005, 8, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Hallem, E.A.; Ho, M.G.; Carlson, J.R. The molecular basis of odor coding in the Drosophila antenna. Cell 2004, 117, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Vosshall, L.B.; Hansson, B.S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 2011, 36, 497–498. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Zhu, X.Q.; Gu, S.H.; Cui, H.H.; Guo, Y.Y.; Zhou, J.J.; Zhang, Y.J. Silencing in Apolygus lucorum of the olfactory coreceptor Orco gene by RNA interference induces EAG response declining to two putative semiochemicals. J. Insect Physiol. 2014, 60, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.W.; Zhang, J.; Liu, Y.; Li, G.Q.; Wang, G.R. An olfactory receptor from Apolygus lucorum (Meyer-Dür) mainly tuned to volatiles from flowering host plants. J. Insect Physiol. 2015, 79, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.R.; Wanner, K.W.; Trowell, S.C.; Warr, C.G.; Jaquin-Joly, E.; Zagatti, P.; Robertson, H.; Newcomb, R.D. Molecular basis of female-specific odorant responses in Bombyx mori. Insect Biochem. Mol. Biol. 2009, 39, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Wanner, K.W.; Anderson, A.R.; Trowell, S.C.; Theilmann, D.A.; Robertson, H.M.; Newcomb, R.D. Female-biased expression of odorant receptor genes in the adult antennae of the silkworm, Bombyx mori. Insect Mol. Biol. 2007, 16, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, C.C.; Yan, S.W.; Liu, Y.; Guo, M.B.; Dong, S.L.; Wang, G.R. An odorant receptor from the common cutworm (Spodoptera litura) exclusively tuned to the important plant volatile cis-3-hexenyl acetate. Insect Mol. Biol. 2013, 22, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.R.; Carey, A.F.; Carlson, J.R.; Zwiebel, L.J. Molecular basis of odor coding in the malaria vector mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2010, 107, 4418–4423. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.J.; Liu, Y.; Walker, W.B.; Liu, C.C.; Lin, K.J.; Gu, S.H.; Zhang, Y.J.; Zhou, J.J.; Wang, G.R. Identification and characterization of pheromone receptors and interplay between receptors and pheromone binding proteins in the diamondback moth, Plutella xyllostella. PLoS ONE 2013, 8, e62098. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.F.; Wang, G.; Su, C.Y.; Zwiebel, L.J.; Carlson, J.R. Odorant reception in the malaria mosquito Anopheles gambiae. Nature 2010, 464, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Zhang, J.; Yan, S.W.; Chang, H.T.; Liu, Y.; Wang, G.R.; Dong, S.L. Functional characterization of sex pheromone receptors in the purple stem borer, Sesamia inferens (Walker). Insect Mol. Biol. 2014, 23, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Liu, Y.; Guo, M.B.; Cao, D.P.; Dong, S.L.; Wang, G.R. Narrow tuning of an odorant receptor to plant volatiles in Spodoptera exigua (Hübner). Insect Mol. Biol. 2014, 23, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Chinta, S.; Dickens, J.C.; Aldrich, J.R. Olfactory reception of potential pheromones and plant odors by tarnished plant bug, Lygus lineolaris (Hemiptera: Miridae). J. Chem. Ecol. 1994, 20, 3251–3267. [Google Scholar] [CrossRef] [PubMed]
- Groot, A.T.; Timmer, R.; Gort, G.; Lelyveld, G.P.; Drijfhout, F.P.; van Beek, T.A.; Visser, J.H. Sex-related perception of insect and plant volatiles in Lygocoris pabulinus. J. Chem. Ecol. 1999, 25, 2357–2371. [Google Scholar] [CrossRef]
- Loughrin, J.H.; Manukian, A.; Heath, R.R.; Turlings, T.C.; Tumlinson, J.H. Diurnal cycle of emission of induced volatile terpenoids by herbivore-injured cotton plant. Proc. Natl. Acad. Sci. USA 1994, 91, 11836–11840. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Scala, A.; Allmann, S.; Mirabella, R.; Haring, M.A.; Schuurink, R.C. Green leaf volatiles: A plant’s multifunctional weapon against herbivores and pathogens. Int. J. Mol. Sci. 2013, 14, 17781–17811. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Q. Identification of Host Palnt Semiochemicals and the Attraction for Apolygus lucorum (Meyer-Dür). Ph.D. Thesis, Agricultural University of Hebei Province, Baoding, China, 2011. [Google Scholar]
- Wardle, A.R.; Borden, J.H.; Pierce, H.D., Jr.; Gries, R. Volatile compounds released by disturbed and calm adults of the tarnished plant bug, Lygus lineolaris. J. Chem. Ecol. 2003, 29, 931–944. [Google Scholar] [CrossRef] [PubMed]
- ClustalW2. Available online: http://www.ebi.ac.uk/Tools/msa/clustalw2 (accessed on 14 October 2015).
- TMHMM Server v. 2.0. Available online: http://www.cbs.dtu.dk/services/TMHMM (accessed on 14 October 2015).
- Lu, T.; Qiu, Y.T.; Wang, G.R.; Kwon, J.Y.; Rutzler, M.; Kwon, H.W.; Pitts, R.J.; Van Loon, J.J.A.; Takken, W.; Carlson, J.R. Odor coding in the maxillary palp of the malaria vector mosquito Anopheles gambiae. Curr. Biol. 2007, 17, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Yu, H.; Zhou, J.J.; Pickett, J.A.; Wu, K.M. Plant volatile analogues strengthen attractiveness to insect. PLoS ONE 2014, 9, e99142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Odorants | Structure | EC50 (M) |
|---|---|---|
| (R)-(+)-Limonene | 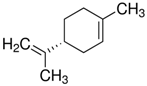 | - |
| (S)-(−)-Limonene | 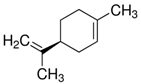 | - |
| (E)-2-Hexenal | 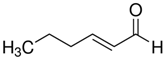 | 3.71 × 10−5 |
| (E)-3-Hexenol | 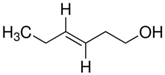 | 2.53 × 10−5 |
| 1-Heptanol | 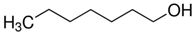 | 1.38 × 10−5 |
| (1R)-(−)-Myrtenol | 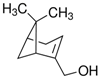 | 7.44 × 10−5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, M.; Yan, S.; Wang, G.; Liu, Y. A Female-Biased Odorant Receptor from Apolygus lucorum (Meyer-Dür) Tuned to Some Plant Odors. Int. J. Mol. Sci. 2016, 17, 1165. https://doi.org/10.3390/ijms17081165
Zhang Z, Zhang M, Yan S, Wang G, Liu Y. A Female-Biased Odorant Receptor from Apolygus lucorum (Meyer-Dür) Tuned to Some Plant Odors. International Journal of Molecular Sciences. 2016; 17(8):1165. https://doi.org/10.3390/ijms17081165
Chicago/Turabian StyleZhang, Zhixiang, Meiping Zhang, Shuwei Yan, Guirong Wang, and Yang Liu. 2016. "A Female-Biased Odorant Receptor from Apolygus lucorum (Meyer-Dür) Tuned to Some Plant Odors" International Journal of Molecular Sciences 17, no. 8: 1165. https://doi.org/10.3390/ijms17081165
APA StyleZhang, Z., Zhang, M., Yan, S., Wang, G., & Liu, Y. (2016). A Female-Biased Odorant Receptor from Apolygus lucorum (Meyer-Dür) Tuned to Some Plant Odors. International Journal of Molecular Sciences, 17(8), 1165. https://doi.org/10.3390/ijms17081165