
Neonatal Heart-Enriched miR-708 Promotes Differentiation of Cardiac Progenitor Cells in Rats

 ,
,
Abstract
:
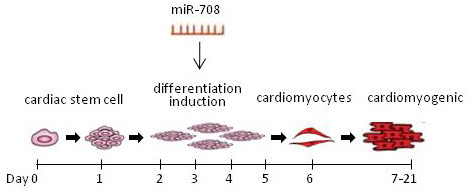{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
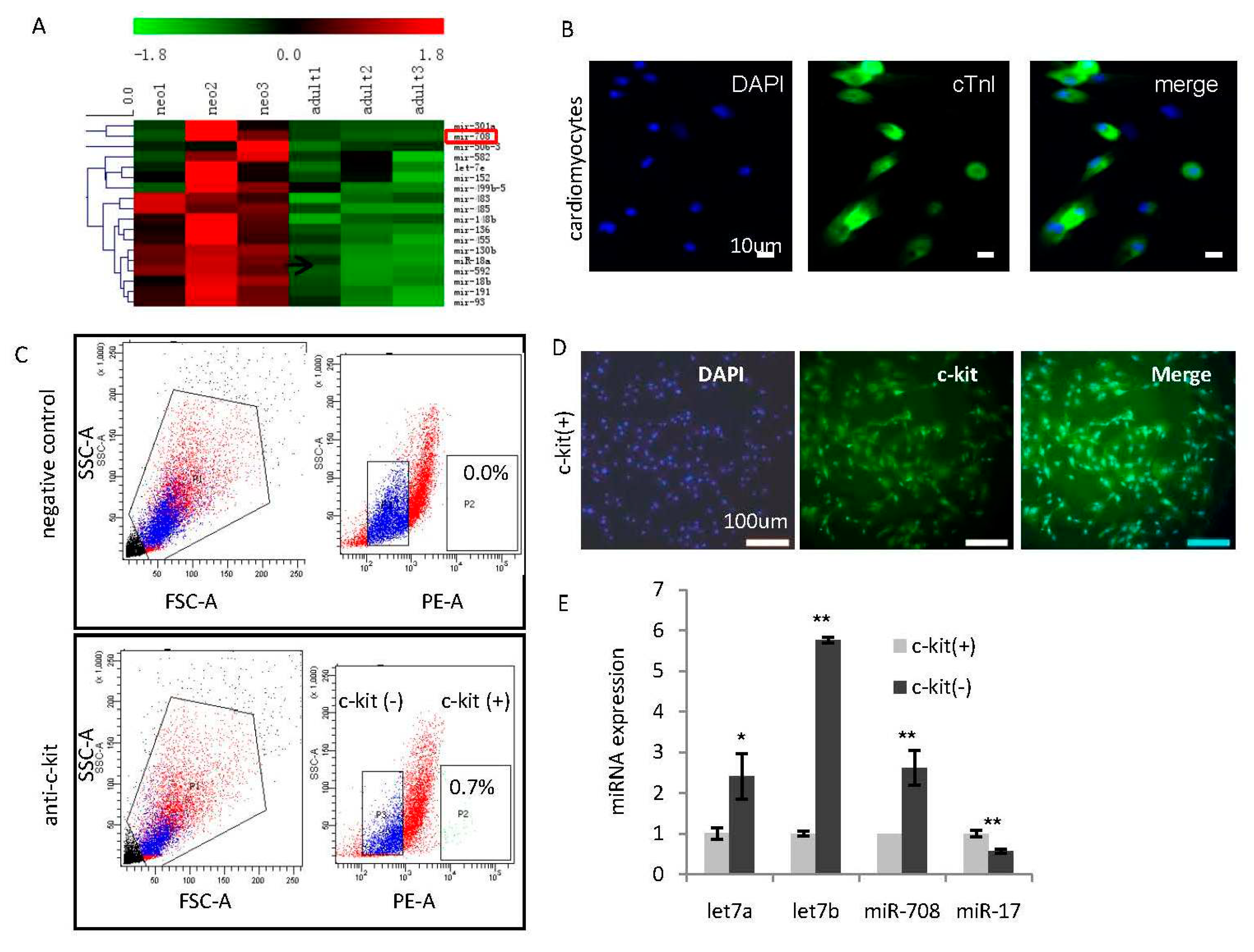
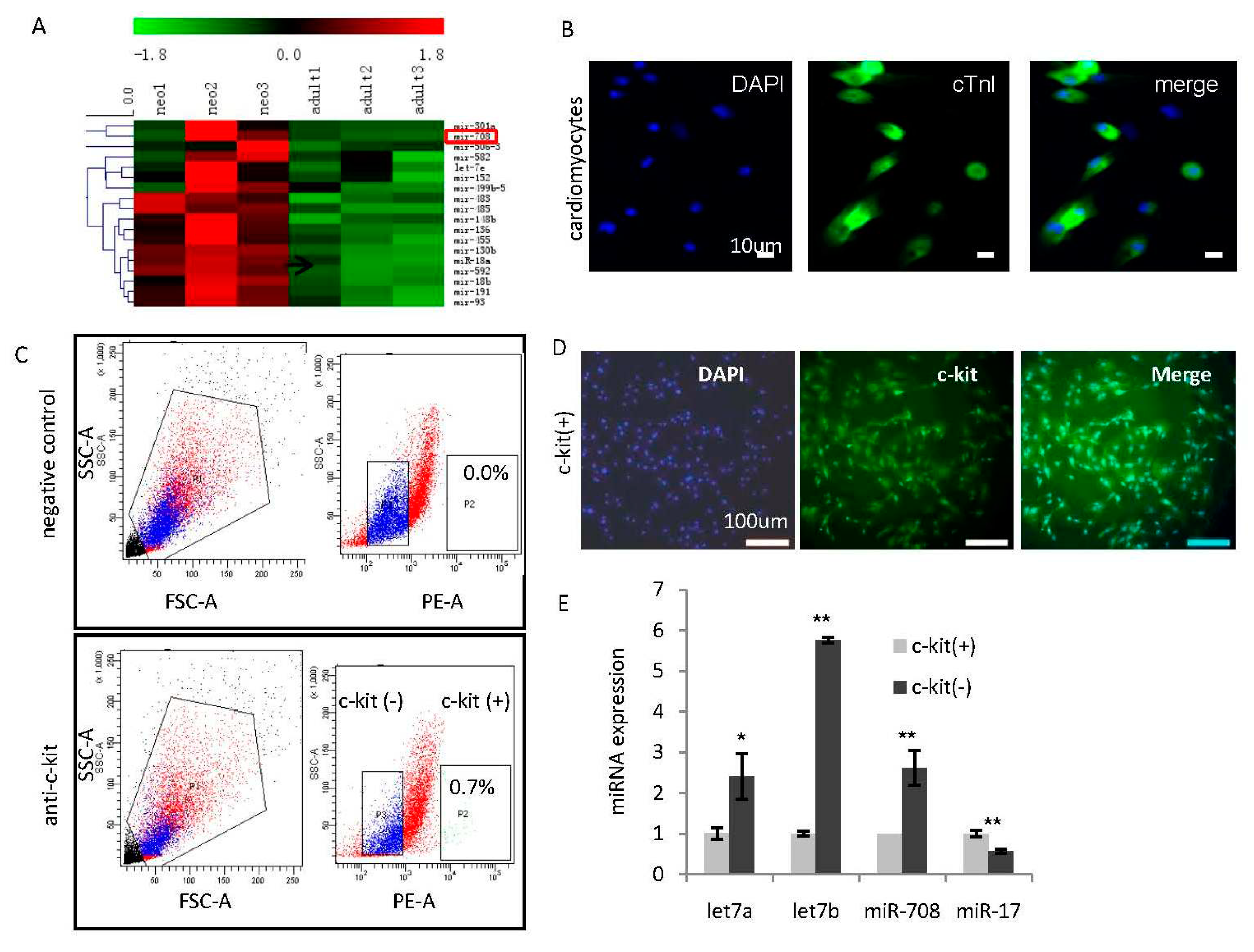
2.1. Identification of miR-708 as a Cardiomyocytes-Enriched miRNA in the Heart of Neonatal Rats
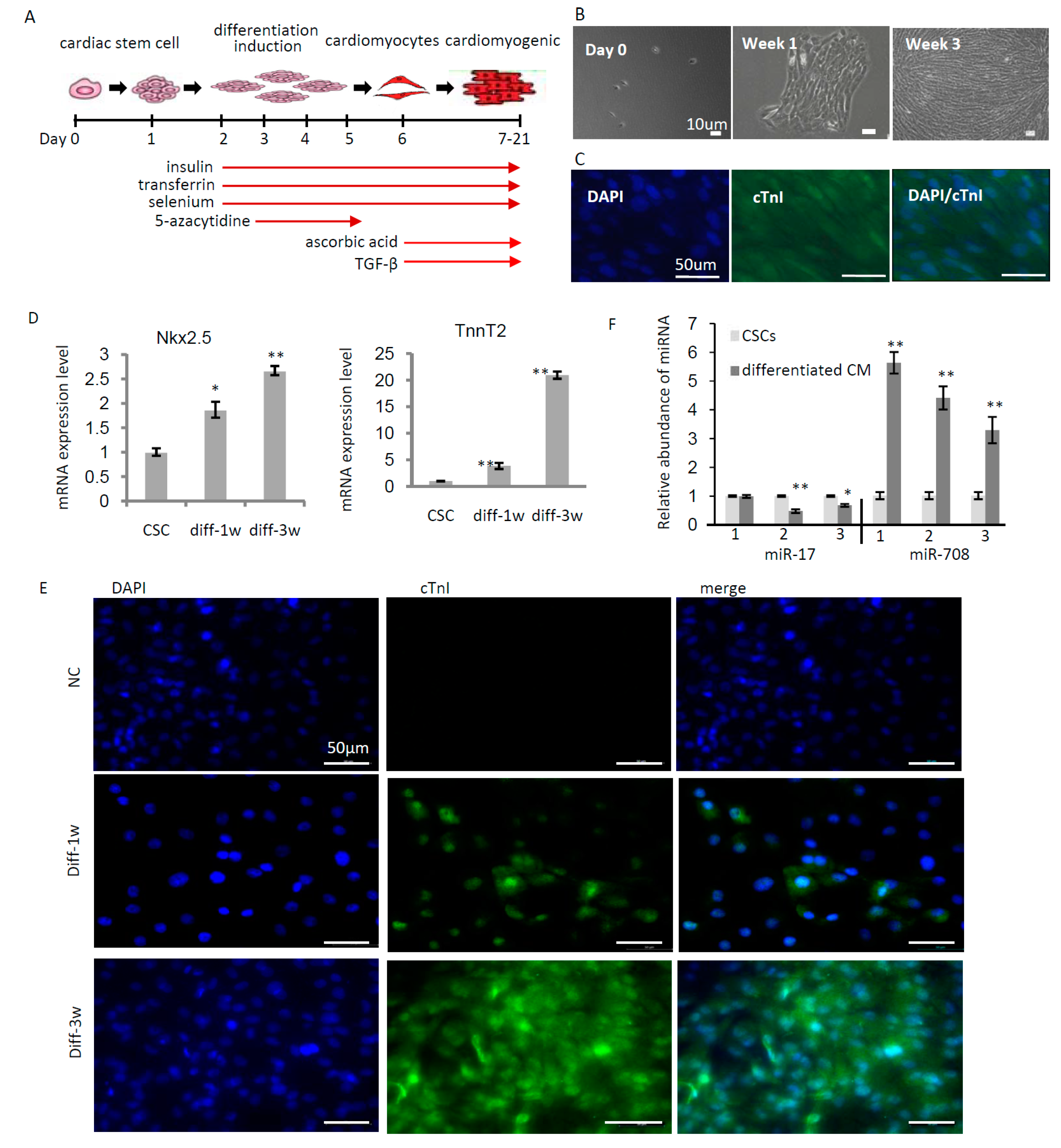
2.2. miR-708 Was Upregulated upon Differentiation of Cardiac Stem/Progenitor Cells
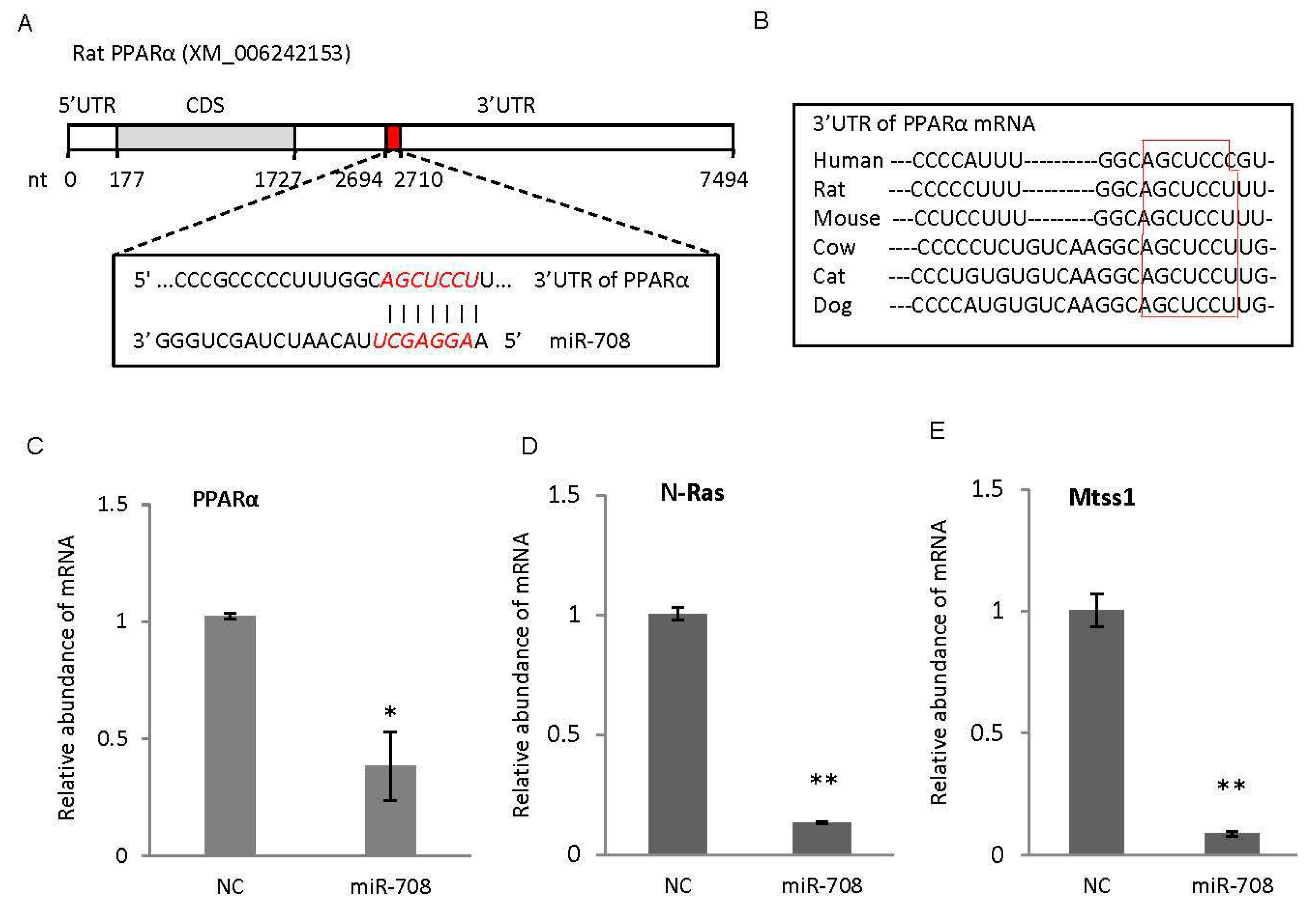
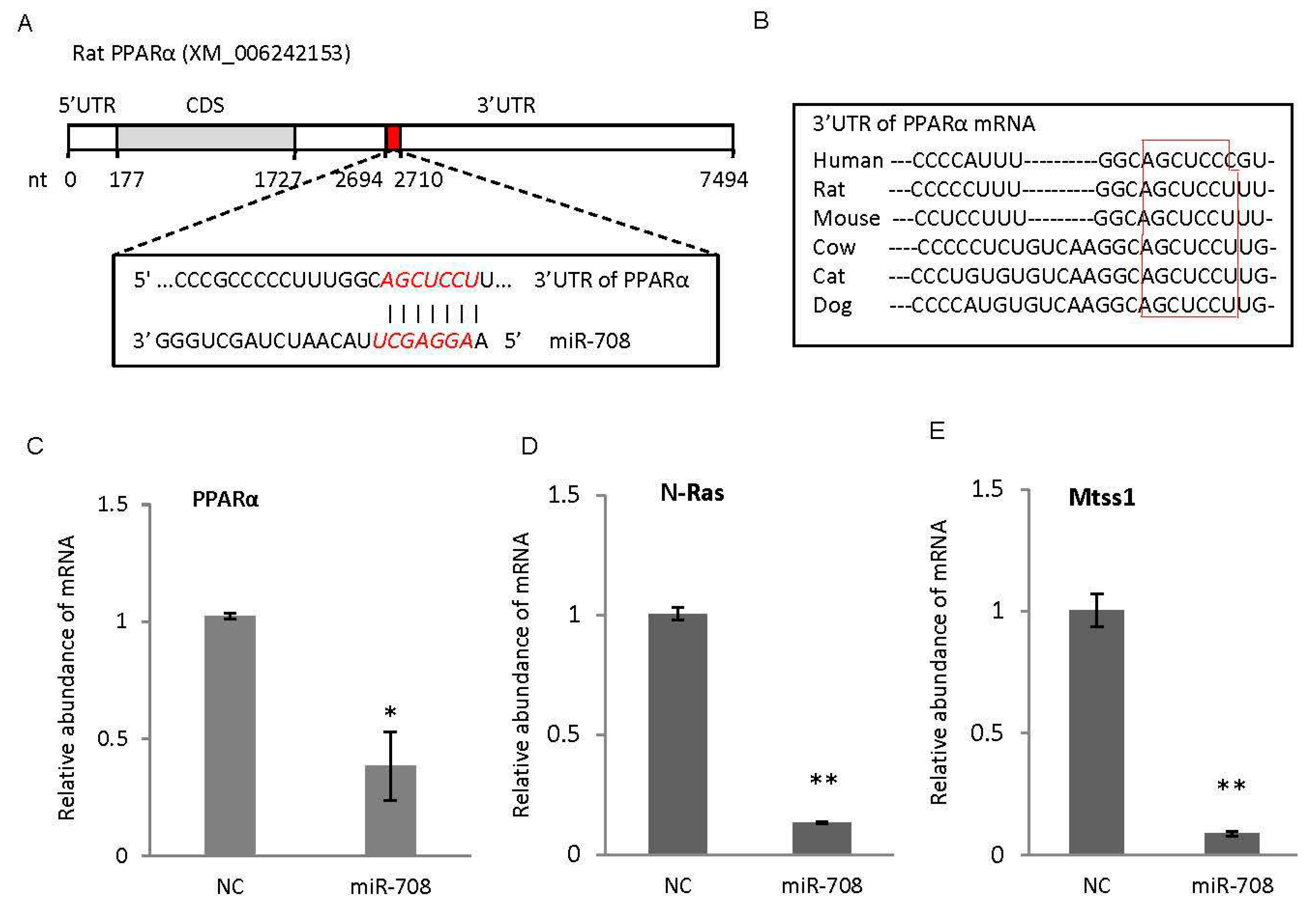
2.3. miR-708 Promoted Differentiation of Cardiac Stem/Progenitor Cells into Cardiomyocytes
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Culture
4.2. Oligos and Transfection
4.3. miRNA QRT-PCR Analysis
4.4. miRNA Profiling Analysis in Neonatal and Adult Rat Heart
4.5. Immunofluorescence
4.6. Induced Differentiation of CSCs
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef]
- Li, F.; Wang, X.; Capasso, J.M.; Gerdes, A.M. Rapid transition of cardiac myocytes from hyperplasia to hypertrophy during postnatal development. J. Mol. Cell. Cardiol. 1996, 28, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Jiang, X.; Martin-Puig, S.; Caron, L.; Zhu, S.; Shao, Y.; Roberts, D.J.; Huang, P.L.; Domian, I.J.; Chien, K.R. Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature 2009, 460, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Mouquet, F.; Pfister, O.; Jain, M.; Oikonomopoulos, A.; Ngoy, S.; Summer, R.; Fine, A.; Liao, R. Restoration of cardiac progenitor cells after myocardial infarction by self-proliferation and selective homing of bone marrow-derived stem cells. Circ. Res. 2005, 97, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Haider, H.K.; Jiang, S.; Ashraf, M. Sca-1+ stem cell survival and engraftment in the infarcted heart: Dual role for preconditioning-induced connexin-43. Circulation 2009, 119, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Linke, A.; Müller, P.; Nurzynska, D.; Casarsa, C.; Torella, D.; Nascimbene, A.; Castaldo, C.; Cascapera, S.; Böhm, M.; Quaini, F.; et al. Stem cells in the dog heart are self-renewing, clonogenic, and multipotent and regenerate infarcted myocardium, improving cardiac function. Proc. Natl. Acad. Sci. USA 2005, 102, 8966–8971. [Google Scholar] [CrossRef] [PubMed]
- Dawn, B.; Stein, A.B.; Urbanek, K.; Rota, M.; Whang, B.; Rastaldo, R.; Torella, D.; Tang, X.L.; Rezazadeh, A.; Kajstura, J.; et al. Cardiac stem cells delivered intravascularly traverse the vessel barrier, regenerate infarcted myocardium, and improve cardiac function. Proc. Natl. Acad. Sci. USA 2005, 102, 3766–3771. [Google Scholar] [CrossRef] [PubMed]
- Chamuleau, S.A.; Vrijsen, K.R.; Rokosh, D.G.; Tang, X.L.; Piek, J.J.; Bolli, R. Cell therapy for ischaemic heart disease: Focus on the role of resident cardiac stem cells. Neth. Heart J. 2009, 17, 199–207. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Urbanek, K.; Torella, D.; Sheikh, F.; de Angelis, A.; Nurzynska, D.; Silvestri, F.; Beltrami, C.A.; Bussani, R.; Beltrami, A.P.; Quaini, F. Myocardial regeneration by activation of multipotent cardiac stem cells in ischemic heart failure. Proc. Natl. Acad. Sci. USA 2005, 102, 8692–8697. [Google Scholar] [CrossRef] [PubMed]
- Stamm, C.; Nasseri, B.; Hetzer, R. Cardiac stem cells in patients with ischaemic cardiomyopathy. Lancet 2012, 379, 891. [Google Scholar] [CrossRef]
- Makkar, R.R.; Smith, R.R.; Cheng, K.; Malliaras, K.; Thomson, L.E.; Berman, D.; Czer, L.S.; Marbán, L.; Mendizabal, A.; Johnston, P.V. Intracoronary cardiosphere-derived cells for heart regeneration after myocardial infarction (CADUCEUS): A prospective, randomised phase 1 trial. Lancet 2012, 379, 895–904. [Google Scholar] [CrossRef]
- Siu, C.W.; Tse, H.F. Cardiac regeneration: Messages from CADUCEUS. Lancet 2012, 379, 870–871. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Li, Y.; Liang, C.L.; Ma, H.Z.; Zhao, Q.; Lu, Y.; Xiang, Z.D.; Li, L.; Qin, J.; Chen, Y.J.; Cho, W.C.; et al. miR-221/222 promotes S-phase entry and cellular migration in control of basal-like breast cancer. Molecules 2014, 19, 7122–7137. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Yu, Z.; Purisima, E.O.; Wang, E. Principles of microRNA regulation of a human cellular signaling network. Mol. Syst. Biol. 2006, 2, 46. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.J.; Gandhi, V.; Plunkett, W.; Sampath, D.; Calin, G.A. MiRNAs and their potential for use against cancer and other diseases. Future Oncol. 2007, 3, 521–537. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Small, E.M.; Olson, E.N. Pervasive roles of microRNAs in cardiovascular biology. Nature 2011, 469, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.M.; van Vliet, P.; Metz, C.H.; Korfage, T.; Sluijter, J.P.; Doevendans, P.A.; Goumans, M.J. Human cardiomyocyte progenitor cells differentiate into functional mature cardiomyocytes: An in vitro model for studying human cardiac physiology and pathophysiology. Nat. Protoc. 2009, 4, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Furtado, K.S.; de Oliveira Andrade, F.; Campos, A.; Rosim, M.P.; Vargas-Mendez, E.; Henriques, A.; de Conti, A.; Scolastici, C.; Barbisan, L.F.; Carvalho, R.F.; et al. β-ionone modulates the expression of miRNAs and genes involved in the metastatic phenotype of microdissected persistent preneoplastic lesions in rats submitted to hepatocarcinogenesis. Mol. Carcinog. 2016. [Google Scholar] [CrossRef] [PubMed]
- Forstner, A.J.; Forstner, A.J.; Hofmann, A.; Maaser, A.; Sumer, S.; Khudayberdiev, S.; Mühleisen, T.W.; Leber, M.; Schulze, T.G.; Strohmaier, J.; et al. Genome-wide analysis implicates microRNAs and their target genes in the development of bipolar disorder. Transl. Psychiatry 2015, 5, e678. [Google Scholar] [CrossRef] [PubMed]
- Van Berlo, J.H.; van Berlo, J.H.; Kanisicak, O.; Maillet, M.; Vagnozzi, R.J.; Karch, J.; Lin, S.C.; Middleton, R.C.; Marbán, E.; Molkentin, J.D. c-kit+ cells minimally contribute cardiomyocytes to the heart. Nature 2014, 509, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Sultana, N.; Sultana, N.; Zhang, L.; Yan, J.; Chen, J.; Cai, W.; Razzaque, S.; Jeong, D.; Sheng, W.; Bu, L.; et al. Resident c-kit(+) cells in the heart are not cardiac stem cells. Nat. Commun. 2015, 6, 8701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Deng, S.Q.; Wang, G.X.; Liu, C.C.; Meng, L.Y.; Qiao, S.S.; Shen, L.; Zhang, Y.; Lü, J.H.; Li, W.S.; et al. A direct quantification method for measuring plasma MicroRNAs identified potential biomarkers for detecting metastatic breast cancer. Oncotarget 2016. [Google Scholar] [CrossRef]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.; Zhao, Q.; Zhou, X.; Zhang, L.; Bao, L.; Zhen, L.; Zhang, Y.; Fan, H.; Liu, Z.; Yu, Z. Neonatal Heart-Enriched miR-708 Promotes Differentiation of Cardiac Progenitor Cells in Rats. Int. J. Mol. Sci. 2016, 17, 875. https://doi.org/10.3390/ijms17060875
Deng S, Zhao Q, Zhou X, Zhang L, Bao L, Zhen L, Zhang Y, Fan H, Liu Z, Yu Z. Neonatal Heart-Enriched miR-708 Promotes Differentiation of Cardiac Progenitor Cells in Rats. International Journal of Molecular Sciences. 2016; 17(6):875. https://doi.org/10.3390/ijms17060875
Chicago/Turabian StyleDeng, Shengqiong, Qian Zhao, Xianjin Zhou, Lin Zhang, Luer Bao, Lixiao Zhen, Yuzhen Zhang, Huimin Fan, Zhongmin Liu, and Zuoren Yu. 2016. "Neonatal Heart-Enriched miR-708 Promotes Differentiation of Cardiac Progenitor Cells in Rats" International Journal of Molecular Sciences 17, no. 6: 875. https://doi.org/10.3390/ijms17060875
APA StyleDeng, S., Zhao, Q., Zhou, X., Zhang, L., Bao, L., Zhen, L., Zhang, Y., Fan, H., Liu, Z., & Yu, Z. (2016). Neonatal Heart-Enriched miR-708 Promotes Differentiation of Cardiac Progenitor Cells in Rats. International Journal of Molecular Sciences, 17(6), 875. https://doi.org/10.3390/ijms17060875