
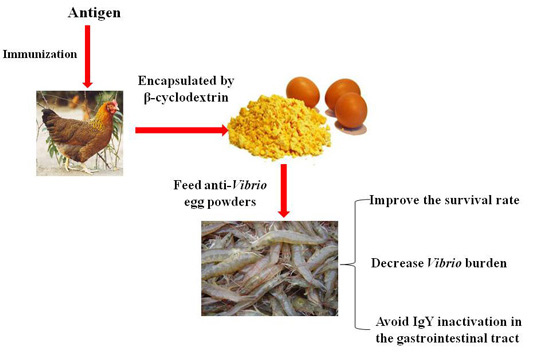
Passive Immune-Protection of Litopenaeus vannamei against Vibrio harveyi and Vibrio parahaemolyticus Infections with Anti-Vibrio Egg Yolk (IgY)-Encapsulated Feed


Abstract
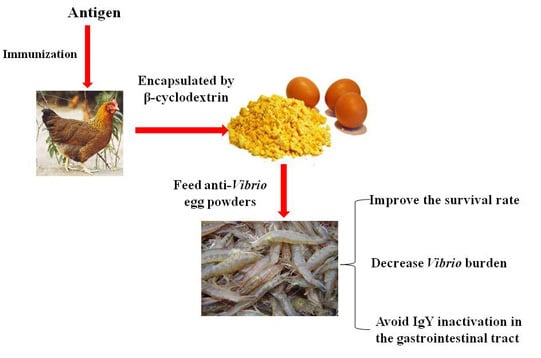
:
1. Introduction
2. Results
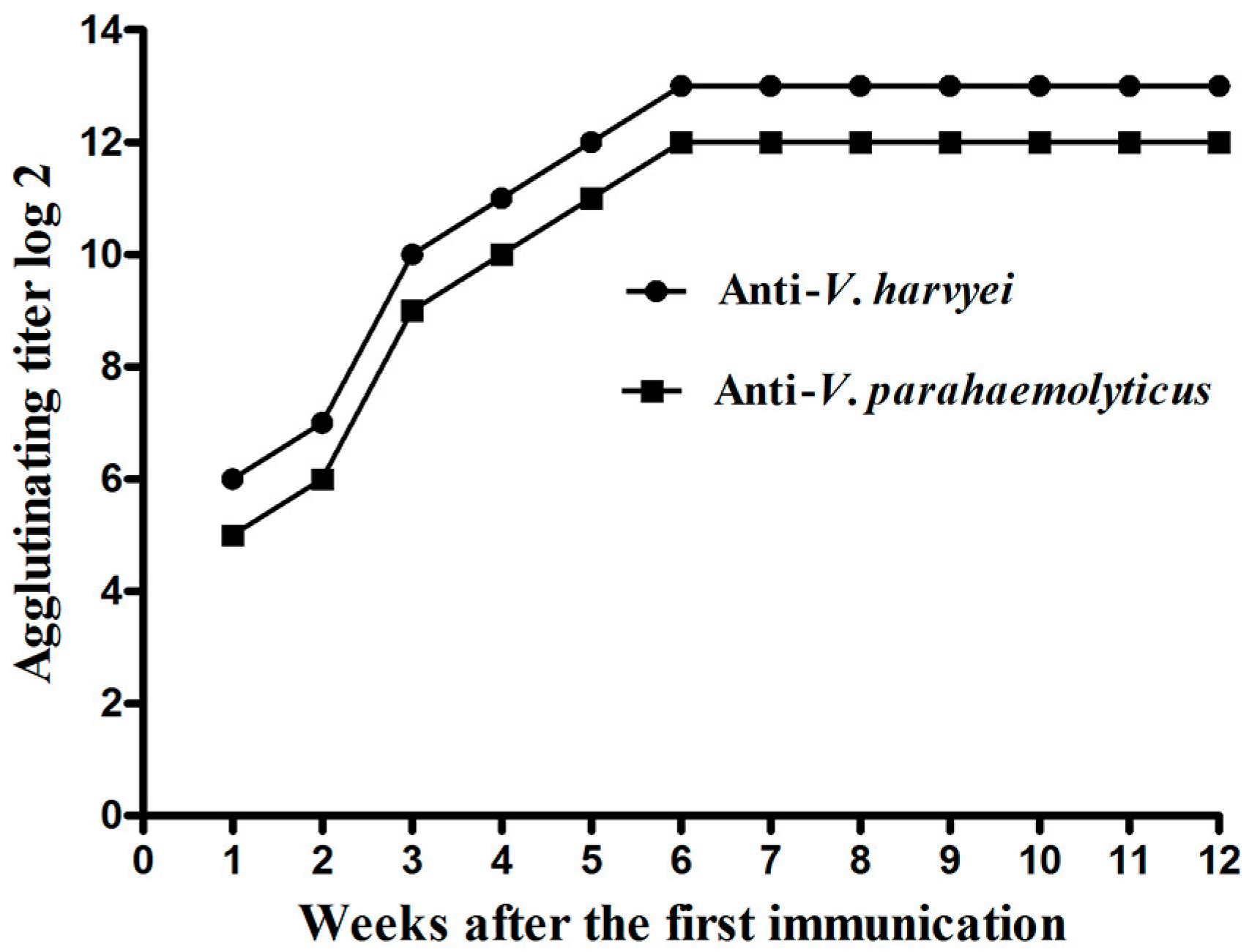
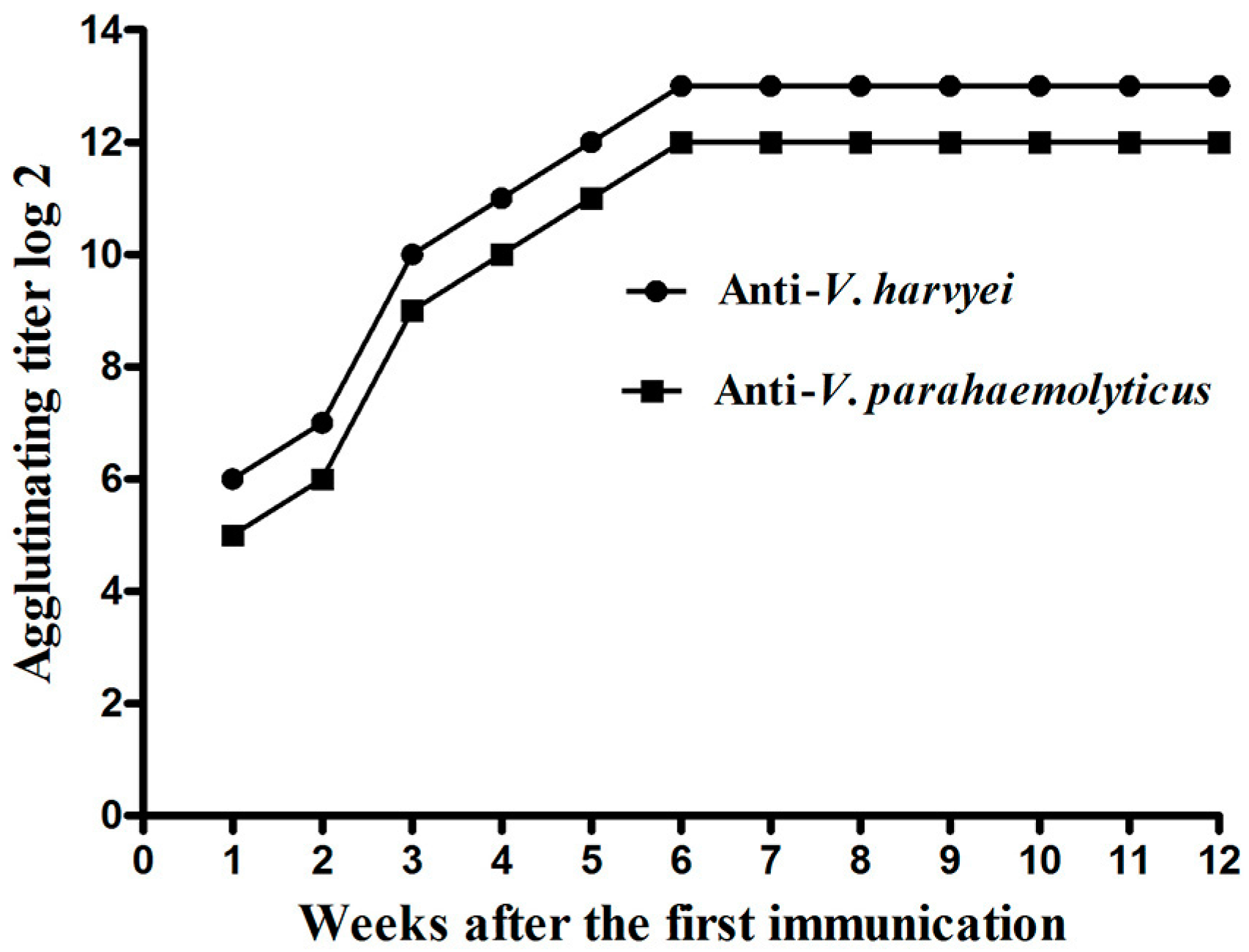
2.1. Titer of Anti-Vibrio IgY in Egg Yolk
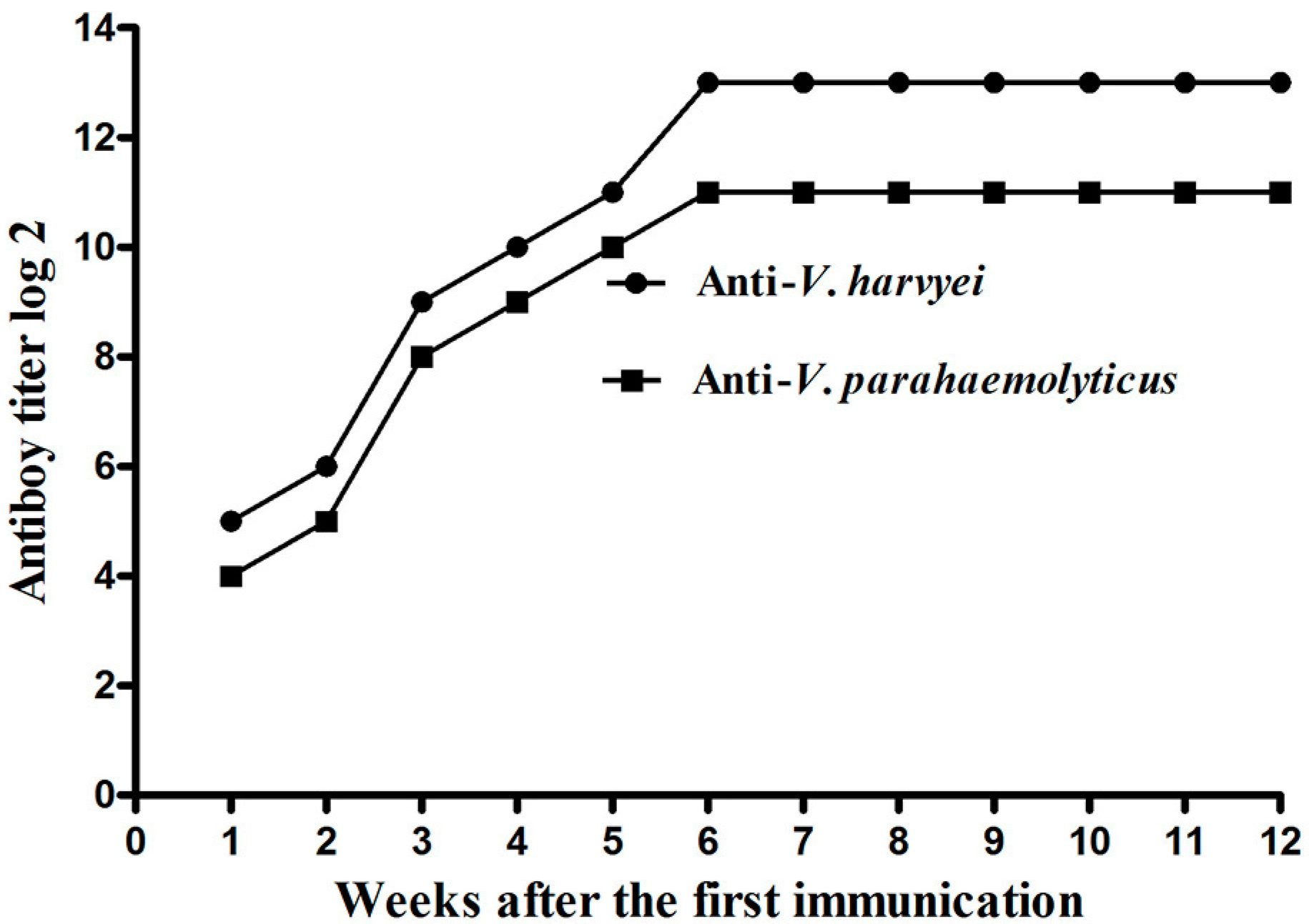
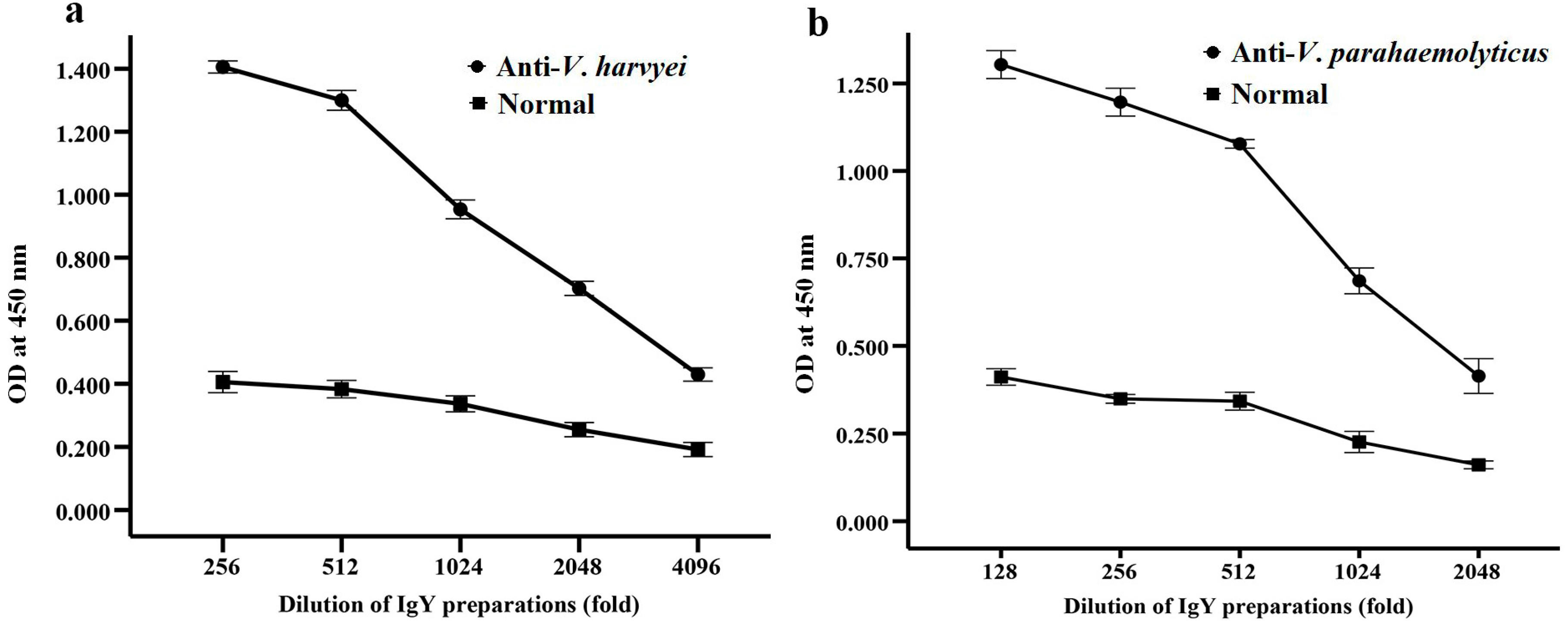
2.2. Titer of Anti-Vibrio IgY in Egg Yolk Powder
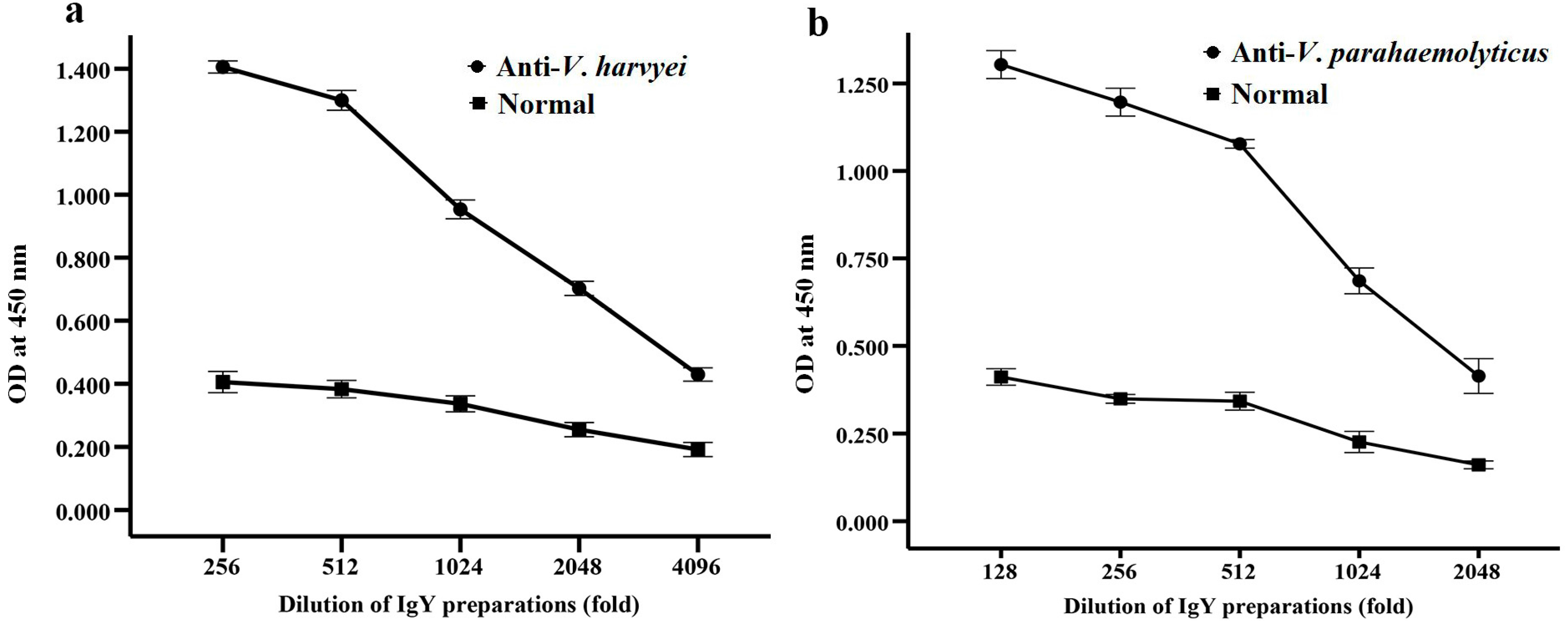
2.3. Inhibitory Effects of Anti-Vibrio IgY
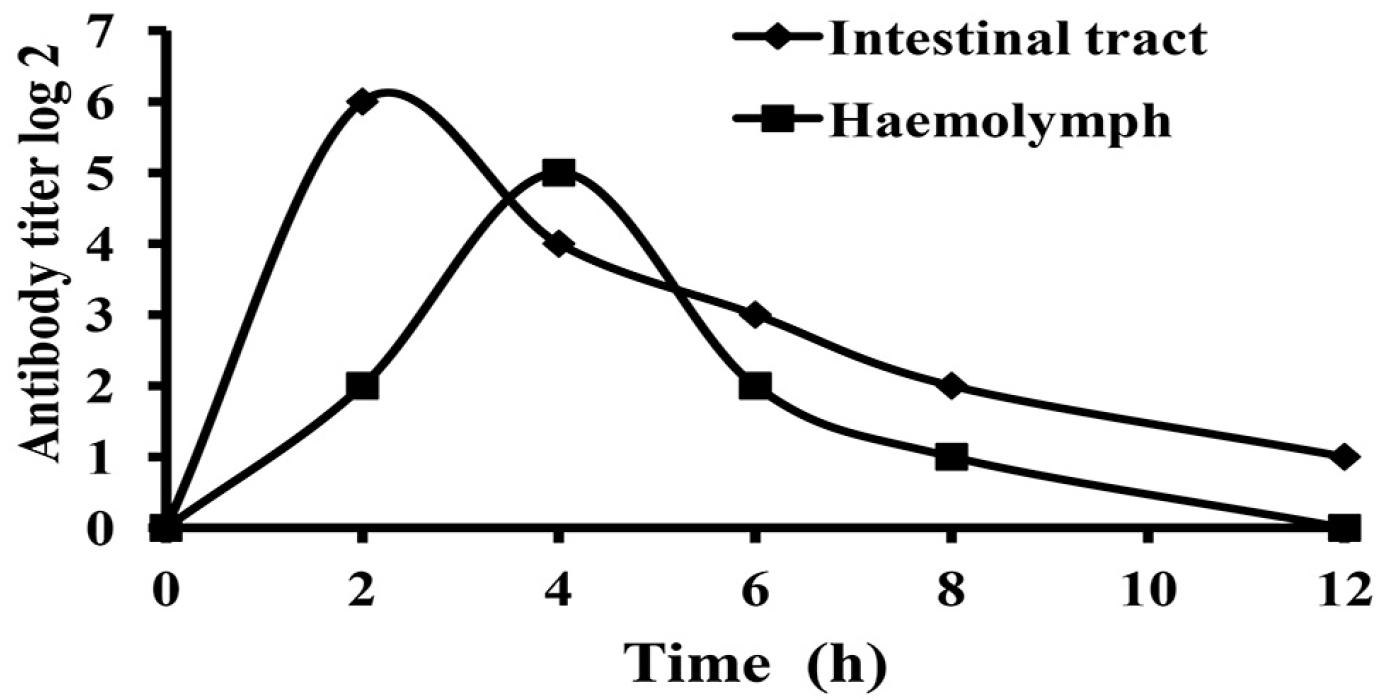
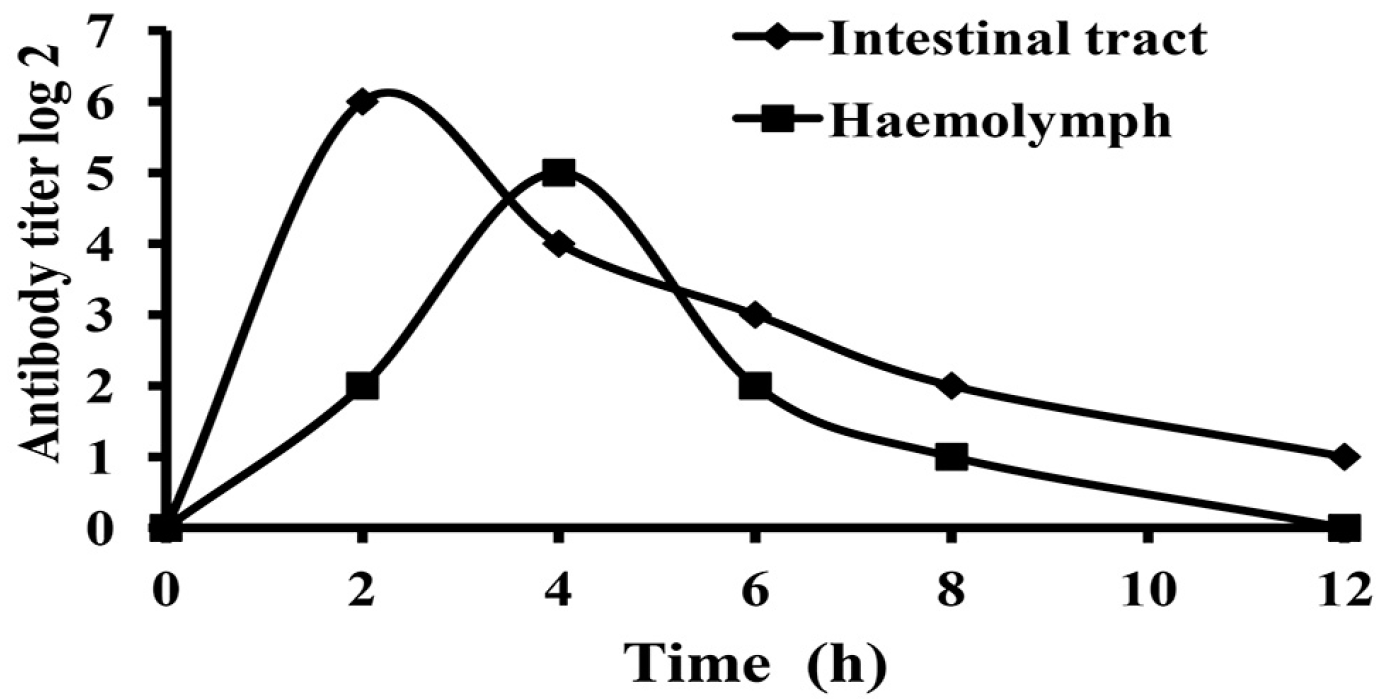
2.4. Absorption and Metabolism of IgY in White Shrimp
2.5. Effects of Dietary Anti-Vibrio Egg Yolk Powder on White Shrimp
2.6. Effects of Anti-Vibrio Yolk Powder on the Bacterial Burden in Postlarva Cultured in Prawn Seeding Ponds
3. Discussion
4. Materials and Methods
4.1. Preparation of Vaccine
4.2. Immunizations of Laying Hens
4.3. Detection of IgY in Yolk by Tube Agglutination Test
4.4. Detection of IgY in Yolk by ELISA
4.5. Preparation of Egg Yolk Powders Containing Anti-Vibiro IgY
4.6. Bacterial Inhibiting Assay
4.7. Kinetic of IgY in White Shrimp
4.8. Challenge Experiment
4.9. Effects of Anti-Vibrio Yolk Powder on the Bacterial Burden in Postlarva of White Shrimp
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lun, J.S.; Xia, C.Y.; Yuan, C.F.; Zhang, Y.L.; Zhong, M.Q.; Huang, T.W.; Hu, Z. The outer membrane protein, LamB (maltoporin), is a versatile vaccine candidate among the Vibrio species. Vaccine 2014, 32, 805–819. [Google Scholar] [CrossRef]
- Sun, R.; Qiu, L.M.; Yue, F.; Wang, L.L.; Liu, R.; Zhou, Z.; Zhang, H.; Song, L.S. Hemocytic immune responses triggered by CpG ODNs in shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2013, 34, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Rungrassamee, W.; Klanchui, A.; Maibunkaew, S.; Karoonuthaisiri, N. Bacterial dynamics in intestines of the black tiger shrimp and the Pacific white shrimp during Vibrio harveyi exposure. J. Invertebr. Pathol. 2016, 133, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rodriguez, S.A.; Gomez-Gil, B.; Lozano, R.; Rio-Rodríguez, R. Virulence of Vibrio harveyi responsible for the “Bright-red” Syndrome in the Pacific white shrimp Litopenaeus vannamei. J. Invertebr. Pathol. 2012, 109, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.J.; Lightner, D.V. Homologues of insecticidal toxin complex genes within a genomic island in the marine bacterium Vibrio parahaemolyticus. FEMS Microbiol. Lett. 2014, 361, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Lomelí-Ortega, C.O.; Martínez-Díaz, S.F. Phage therapy against Vibrio parahaemolyticus infection in thewhiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 2014, 434, 208–211. [Google Scholar] [CrossRef]
- Zhang, X.J.; Bi, K.R.; Yan, B.L.; Chen, L.; Bai, X.S.; Qin, L. Identification and virulence genes detection of pathogenic Vibrio harveyi isolated from mysis of Fenneropenaeus chinensis L. Prog. Fish. Sci. 2014, 35, 105–111. [Google Scholar]
- Zhang, X.J.; Chen, C.Z.; Yan, B.L.; Fang, H.; Qin, G.M.; Xu, J. Phenotypic and molecular characterization of pathogenic Vibrio parahaemolyticus isolated from Penaeus vannamei. Oceanol. Limnol. Sin. 2009, 40, 654–662. [Google Scholar]
- Dang, H.Y.; Zhao, J.Y.; Song, L.S.; Chen, M.N.; Chang, Y.Q. Molecular characterizations of chloramphenicol-and oxytetracycline-resistant bacteria and resistance genes in mariculture waters of China. Mar. Pollut. Bull. 2009, 58, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qiu, L.M.; Song, L.S.; Zhang, H.; Zhao, J.M.; Wang, L.L.; Yu, Y.D.; Li, C.H.; Li, F.M.; Xing, K.Z.; et al. Cloning and characterization of a novel C-type lectin gene from shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2009, 26, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.G.; Eto, S.F.; Ferraro, A.C.N.S.; Marioto, D.T.G.; Vieira, N.J.; Cheirubim, A.P.; Ramos, S.P.; Venâncio, E.J. The production and characterization of anti-bothropic and anti-crotalic IgY antibodies in laying hens: A long term experiment. Toxicon 2013, 66, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.A.; Franco, M.T.; Zapata-Quintanilla, L.; Carbonare, S.B. In vitro reactivity and growth inhibition of EPEC serotype O111 and STEC serotypes O111 and O157 by homologous and heterologous chicken egg yolk antibody. Vet. Res. Commun. 2008, 32, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.N.; Liu, J.J.; Jin, L.J.; Li, X.Y.; Zhen, Y.H.; Xue, H.Y.; You, J.S.; Xu, Y.P. Passive protection of shrimp against white spot syndrome virus (WSSV) using specific antibody from egg yolk of chickens immunized with inactivated virus or a WSSV-DNA vaccine. Fish Shellfish Immunol. 2008, 25, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Spillner, E.; Braren, I.; Greunke, K.; Seismann, H.; Blank, S.; Plessis, D. Avian IgY antibodies and their recombinant equivalents in research, diagnostics and therapy. Biologicals 2012, 40, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.J.; He, H.W.; Sato, A.; Hatta, H.; Nakao, M.; Somamoto, T. Ulcer disease prophylaxis in koi carp by bath immersion with chicken egg yolk containing anti-Aeromonas salmonicida IgY. Res. Vet. Sci. 2015, 99, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.D.; Tambourgi, D.V. IgY: A promising antibody for use in immunodiagnostic and in immunotherapy. Vet. Immunol. Immunopathol. 2010, 135, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.P.; Li, X.Y.; Jin, L.J.; Zhen, Y.H.; Lu, Y.N.; Li, S.Y.; You, J.S.; Wang, L.H. Application of chicken egg yolk immunoglobulins in the control of terrestrial and aquatic animal diseases: A review. Biotechnol. Adv. 2011, 29, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.J.; Li, X.Y.; Zou, D.L.; Li, S.Y.; Song, W.Q.; Xu, Y.P. Protection of crucian carp (Carassius auratus Gibelio) against septicaemia caused by Aeromonas hydrophila using specific egg yolk immunoglobulins. Aquac. Res. 2013, 44, 928–936. [Google Scholar] [CrossRef]
- Lu, Y.N.; Liu, J.J.; Jin, L.J.; Li, X.Y.; Zhen, Y.H.; Xue, H.Y.; Lin, Q.Y.; Xu, Y.P. Passive immunization of crayfish (Procambius clarkiaii) with chicken egg yolk immunoglobulin (IgY) against white spot syndrome virus (WSSV). Appl. Biochem. Biotechnol. 2009, 159, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Liu, H.; Xu, Y.P.; Xu, F.X.; Wang, L.H.; You, J.S.; Li, S.Y.; Jin, L.J. Chicken egg yolk antibody (IgY) controls Solobacterium moorei under in vitro and in vivo conditions. Appl. Biochem. Biotechnol. 2012, 168, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.M.; Rahman, A.K.M.S.; Isoda, R.; Umeda, K.; Sa, N.V.; Kodamaa, Y. In vitro and in vivo effectiveness of egg yolk antibody against Candida albicans (anti-CA IgY). Vaccine 2008, 26, 2073–2080. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Wang, H.; Chan, Y.L.; Li, T.L. Passive immune-protection of small abalone against Vibrio alginolyticus infection by anti-Vibrio IgY-encapsulated feed. Fish Shellfish Immunol. 2011, 30, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Mine, Y.; Stevenson, R.M. Effects of hen egg yolk immunoglobulin in passive protection of rainbow trout against Yersinia ruckeri. J. Agric. Food Chem. 1999, 48, 110–115. [Google Scholar] [CrossRef]
- Fan, J.H.; Zuo, Y.Z.; Li, T.Q.; Zhang, X.B. Preparation and physicochemical property of chicken yolk immunoglobulin (IgY) against porcine transmissible gastroenteritis virus (TGEV). Front. Agric. China 2009, 3, 466–470. [Google Scholar] [CrossRef]
- Li, X.Y.; Jin, L.J.; Uzonna, J.E.; Li, S.Y.; Liu, J.J.; Li, H.Q.; Lu, Y.N.; Zhen, Y.H.; Xu, Y.P. Chitosan-alginate microcapsules for oral delivery of egg yolk immunoglobulin (IgY): In vivo evaluation in a pig model of enteric colibacillosis. Vet. Immunol. Immunopathol. 2009, 129, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Bellingeri, R.V.; Picco, N.Y.; Alustiza, F.E.; Canova, J.V.; Molina, M.A.; Acevedo, D.F.; Barbero, C.; Vivas, A.B. pH-responsive hydrogels to protect IgY from gastric conditions: In vitro evaluation. J. Food Sci. Technol. 2015, 52, 3117–3122. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Jing, K.L.; Wang, X.T.; Li, Y.; Zhang, M.X.; Li, Z.; Xu, L.; Wang, L.L.; Xu, Y.P. Protective effects of chicken egg yolk antibody (IgY) against experimental Vibrio splendidus infection in the sea cucumber (Apostichopus japonicus). Fish Shellfish Immunol. 2016, 48, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.P.; Sharma, S.K.; Gautam, O.P.; Sarup, S. The use of spleen antigen in the tube agglutination test for diagnosis of anaplasmosis in cattle. Trop. Anim. Health Prod. 1978, 10, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Lu, X.J.; Li, D.F.; Chen, J. Passive protective effect of chicken egg yolk immunoglobulins against experimental Vibrio anguillarum infection in ayu (Plecoglossus altivelis). Fish Shellfish Immunol. 2014, 37, 108–114. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Dilution of Crude Extraction of IgY (Fold) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1:2 | 1:4 | 1:8 | 1:16 | 1:32 | 1:64 | 1:128 | 1:256 | 1:512 | |
| V. harveyi | − | − | − | − | + | + | + | + | + |
| V. parahaemolyticus | − | − | − | + | + | + | + | + | + |
| Bacteria | Groups | Feed | Dead Larva After Challenge (h) | Total Deaths | Mortality (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| 6 | 12 | 18 | 24 | 48 | |||||
| V. harveyi | Zoea | Specific IgY | 6 | 10 | 12 | 13 | 15 | 56 | 37.3 |
| Normal IgY | 24 | 28 | 21 | 29 | 24 | 126 | 84.0 | ||
| Mysis | Specific IgY | 5 | 10 | 13 | 15 | 16 | 59 | 39.3 | |
| Normal IgY | 19 | 28 | 30 | 23 | 27 | 127 | 84.7 | ||
| Postlarva | Specific IgY | 8 | 11 | 13 | 12 | 13 | 53 | 38.0 | |
| Normal IgY | 16 | 24 | 28 | 29 | 35 | 132 | 88.0 | ||
| V. parahaemolyticus | Zoea | Specific IgY | 4 | 12 | 15 | 17 | 12 | 60 | 40.0 |
| Normal IgY | 34 | 30 | 23 | 19 | 24 | 130 | 86.7 | ||
| Mysis | Specific IgY | 3 | 12 | 16 | 18 | 13 | 62 | 41.4 | |
| Normal IgY | 21 | 28 | 32 | 23 | 22 | 126 | 84.0 | ||
| Postlarva | Specific IgY | 8 | 18 | 13 | 11 | 15 | 65 | 43.3 | |
| Normal IgY | 15 | 23 | 29 | 26 | 38 | 131 | 87.3 | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Zhang, X.; Lin, L.; Yao, D.; Sun, J.; Du, X.; Li, X.; Zhang, Y. Passive Immune-Protection of Litopenaeus vannamei against Vibrio harveyi and Vibrio parahaemolyticus Infections with Anti-Vibrio Egg Yolk (IgY)-Encapsulated Feed. Int. J. Mol. Sci. 2016, 17, 723. https://doi.org/10.3390/ijms17050723
Gao X, Zhang X, Lin L, Yao D, Sun J, Du X, Li X, Zhang Y. Passive Immune-Protection of Litopenaeus vannamei against Vibrio harveyi and Vibrio parahaemolyticus Infections with Anti-Vibrio Egg Yolk (IgY)-Encapsulated Feed. International Journal of Molecular Sciences. 2016; 17(5):723. https://doi.org/10.3390/ijms17050723
Chicago/Turabian StyleGao, Xiaojian, Xiaojun Zhang, Li Lin, Dongrui Yao, Jingjing Sun, Xuedi Du, Xiumei Li, and Yue Zhang. 2016. "Passive Immune-Protection of Litopenaeus vannamei against Vibrio harveyi and Vibrio parahaemolyticus Infections with Anti-Vibrio Egg Yolk (IgY)-Encapsulated Feed" International Journal of Molecular Sciences 17, no. 5: 723. https://doi.org/10.3390/ijms17050723
APA StyleGao, X., Zhang, X., Lin, L., Yao, D., Sun, J., Du, X., Li, X., & Zhang, Y. (2016). Passive Immune-Protection of Litopenaeus vannamei against Vibrio harveyi and Vibrio parahaemolyticus Infections with Anti-Vibrio Egg Yolk (IgY)-Encapsulated Feed. International Journal of Molecular Sciences, 17(5), 723. https://doi.org/10.3390/ijms17050723