
Aminomethylphosphonic Acid and Methoxyacetic Acid Induce Apoptosis in Prostate Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
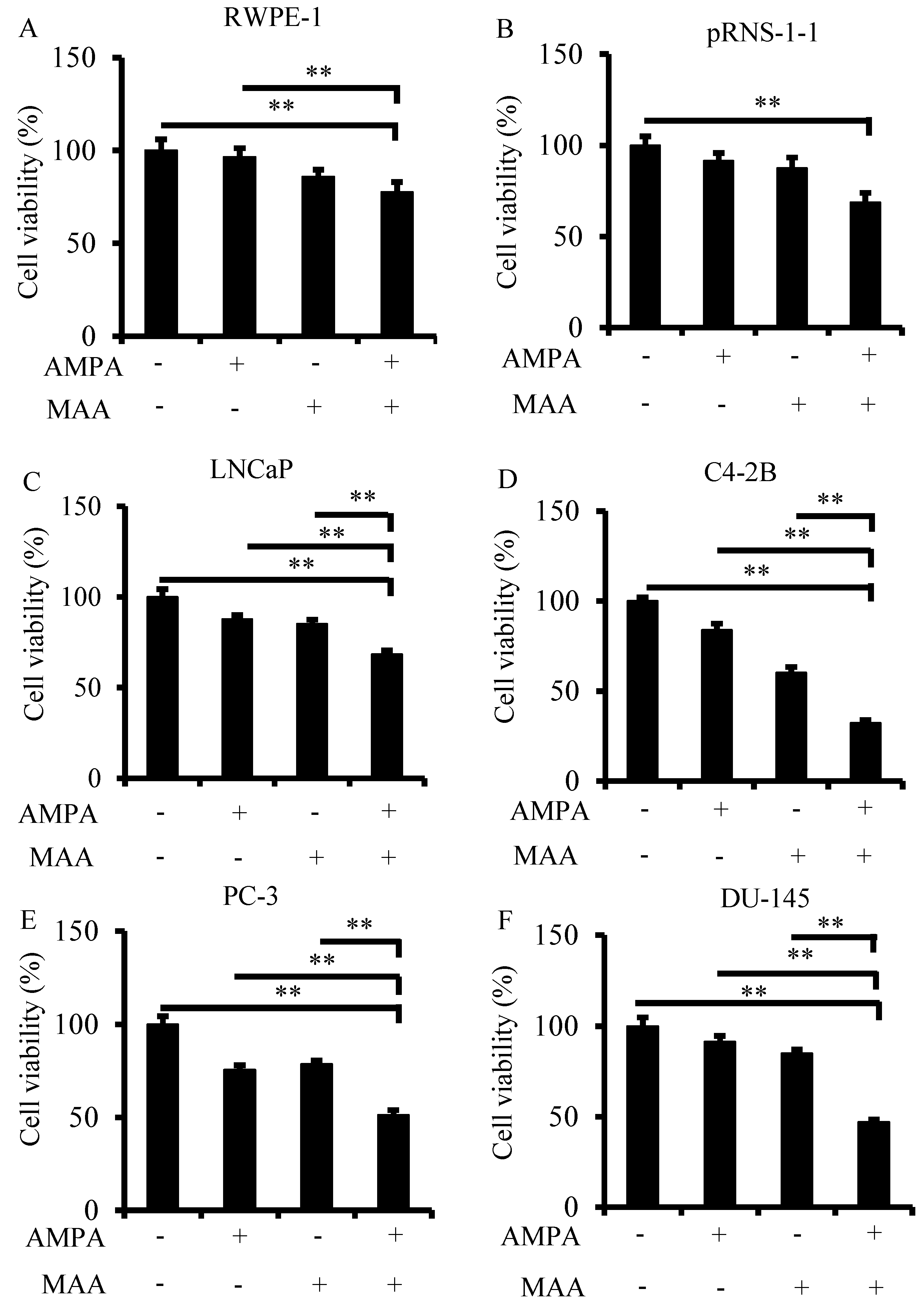
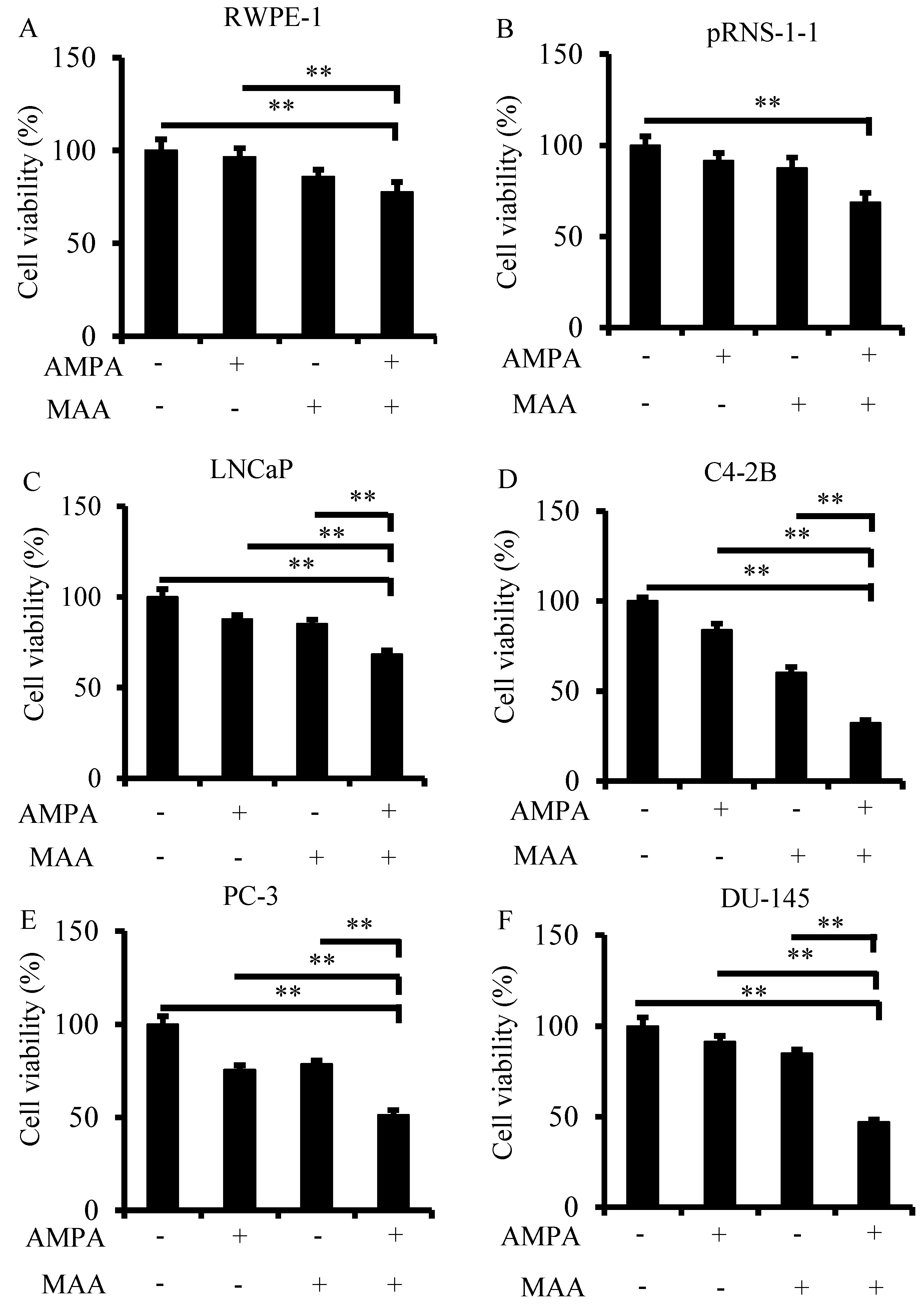
2.1. The Aminomethylphosphonic Acid (AMPA) and Methoxyacetic Acid (MAA) Combination Inhibits Prostate Cancer Cell Viability
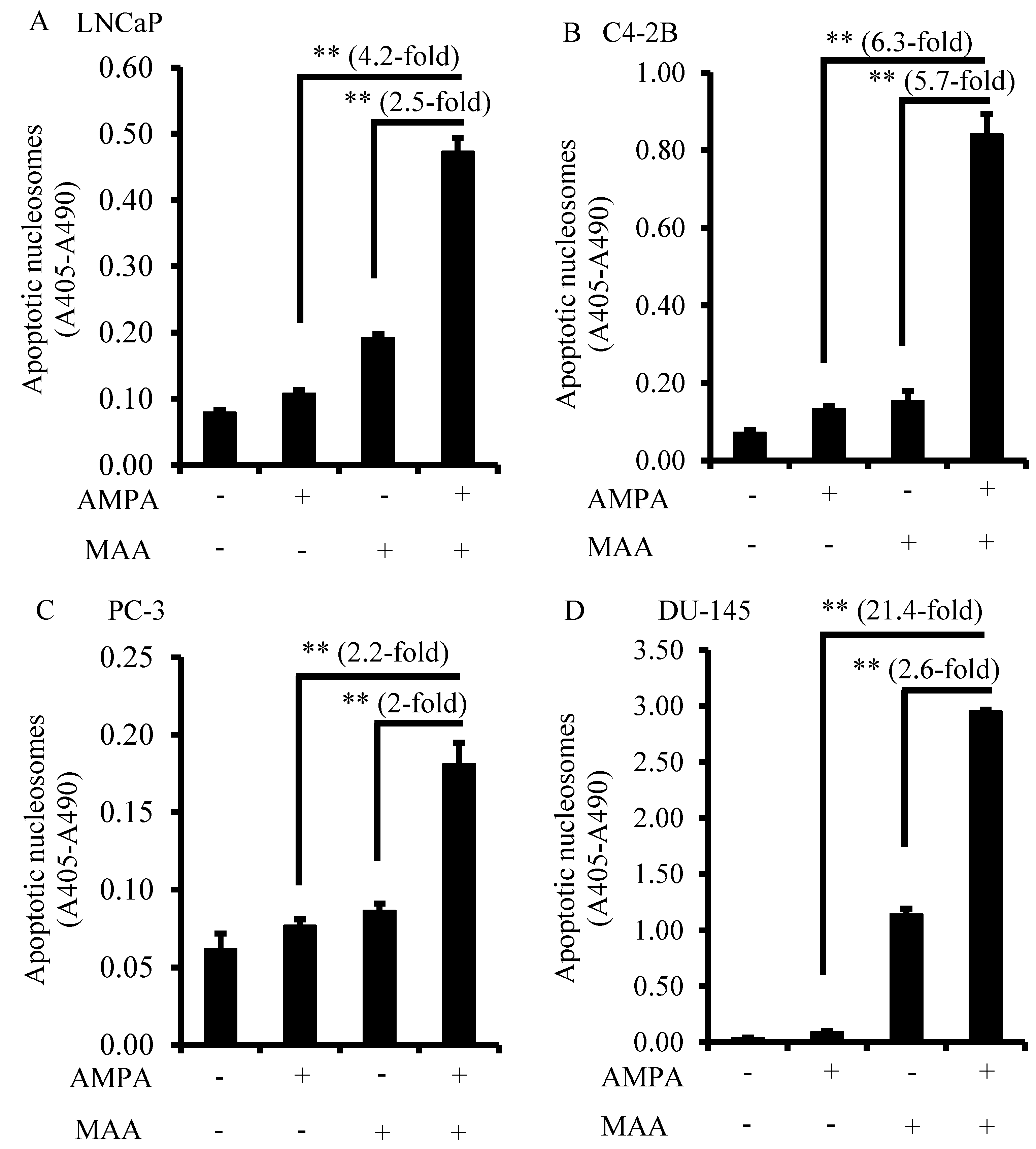
2.2. The Combination of AMPA and MAA Potentiates Apoptosis in Prostate Cancer Cells
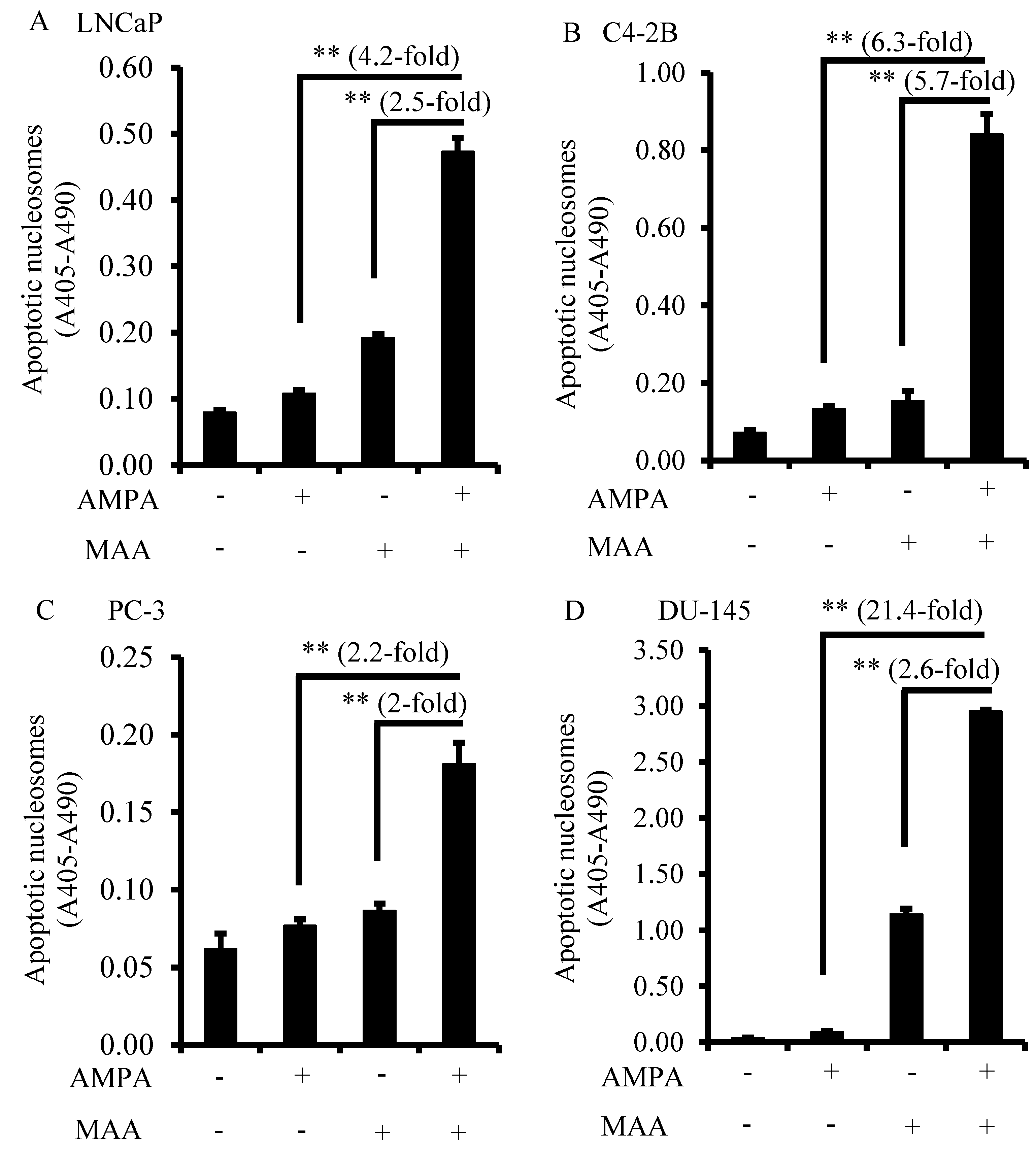
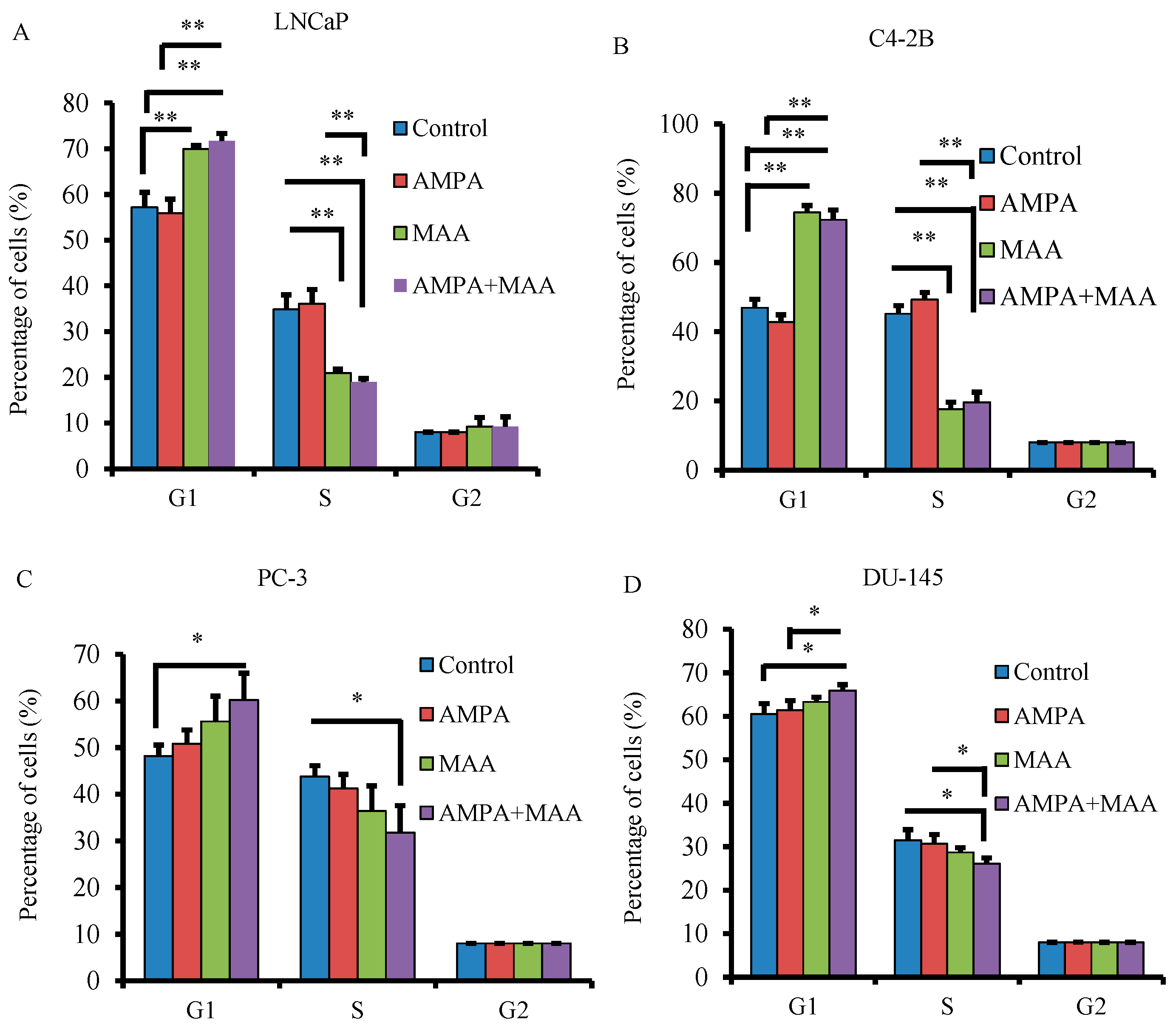
2.3. The Combination of AMPA and MAA Blocks the Entry of Cells from the G1 to S Phase of the Cell Cycle
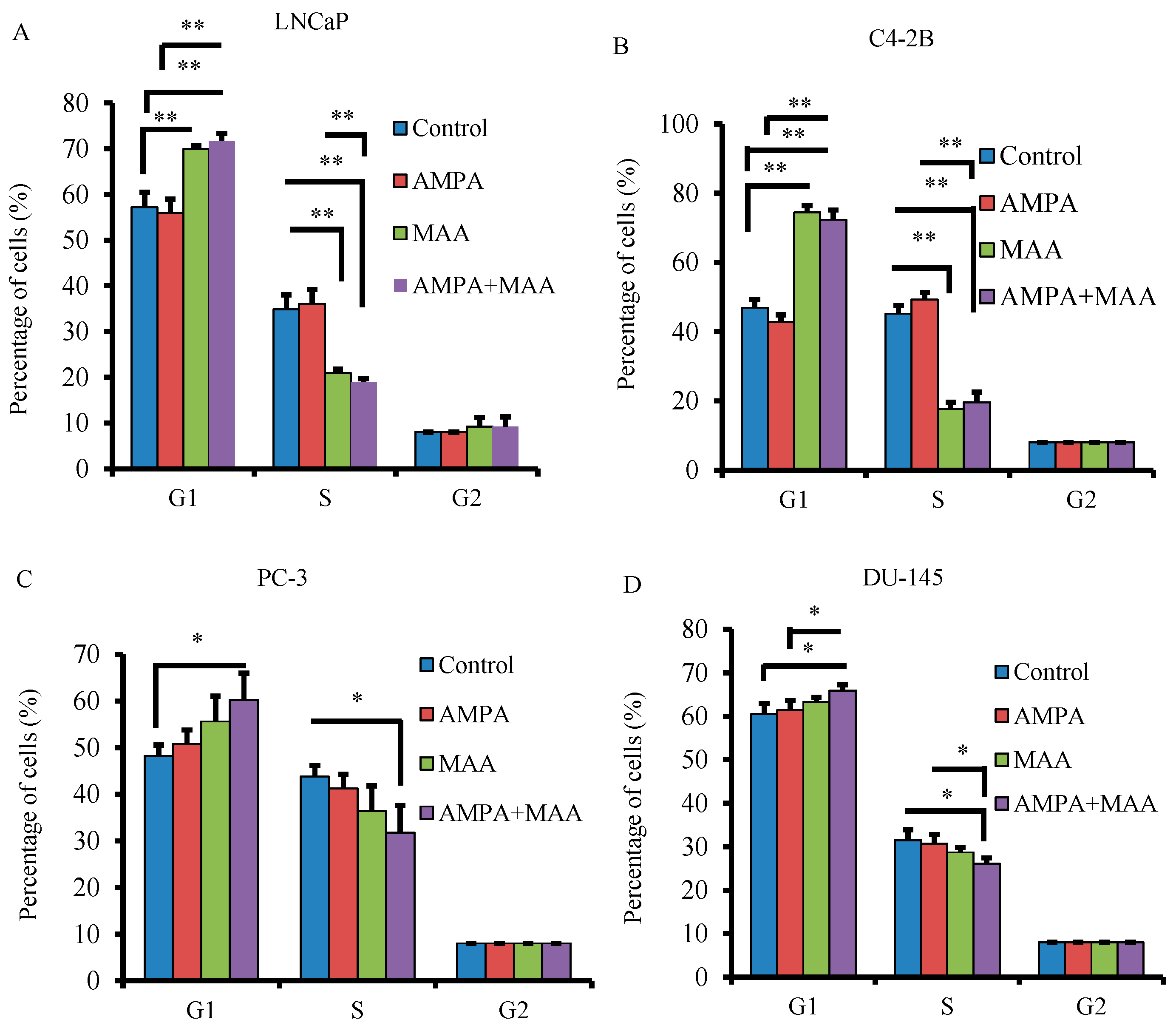
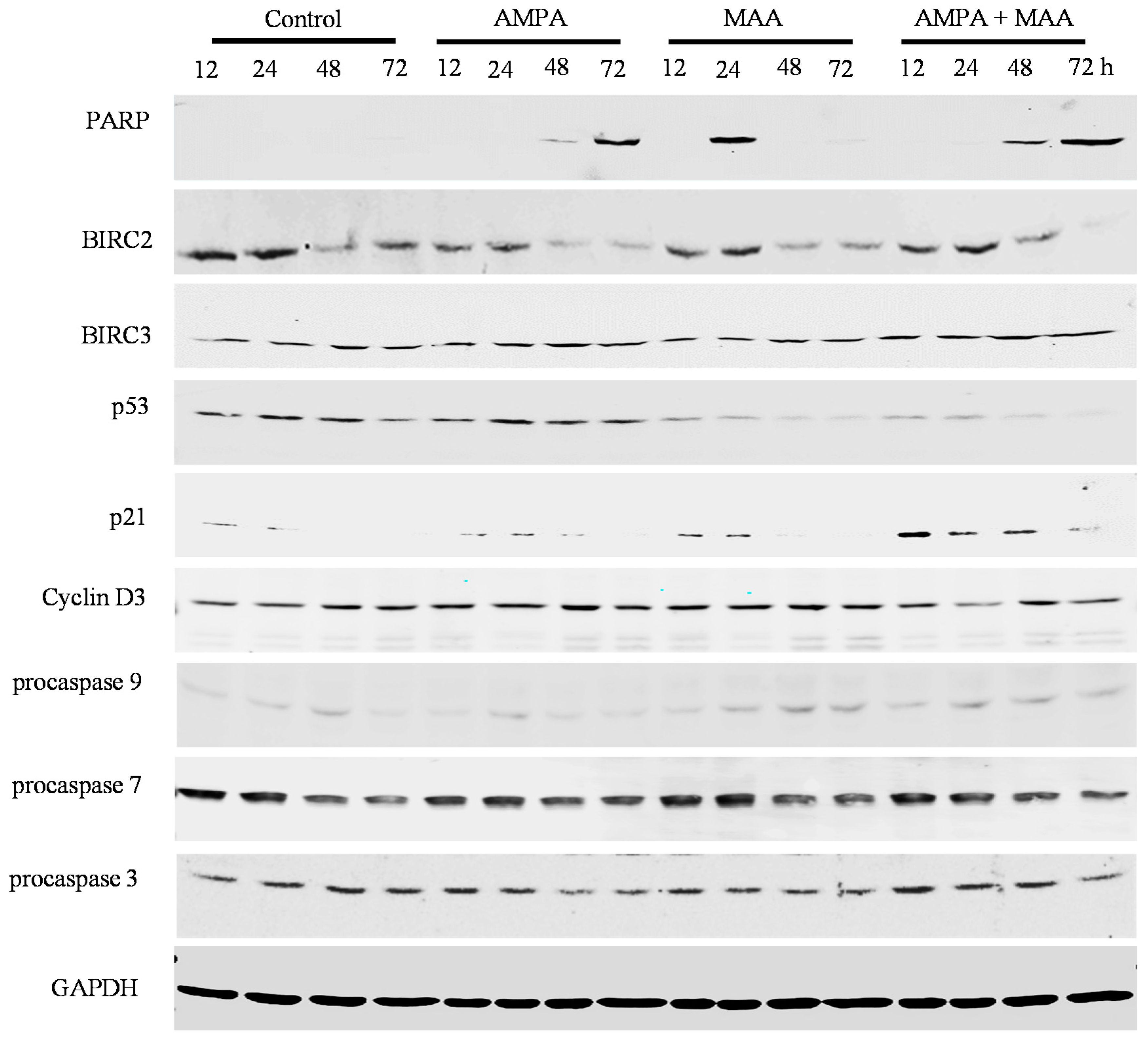
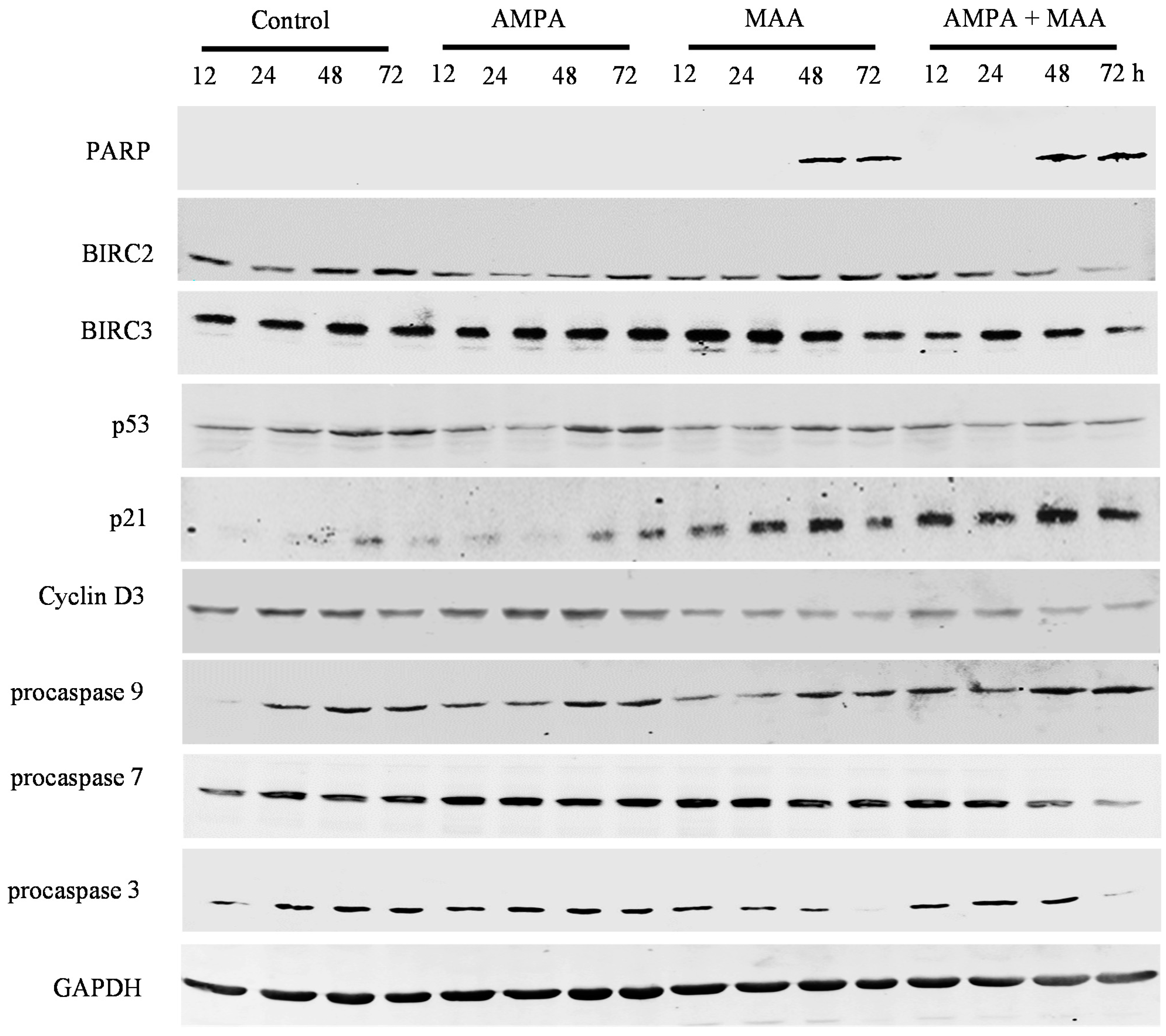
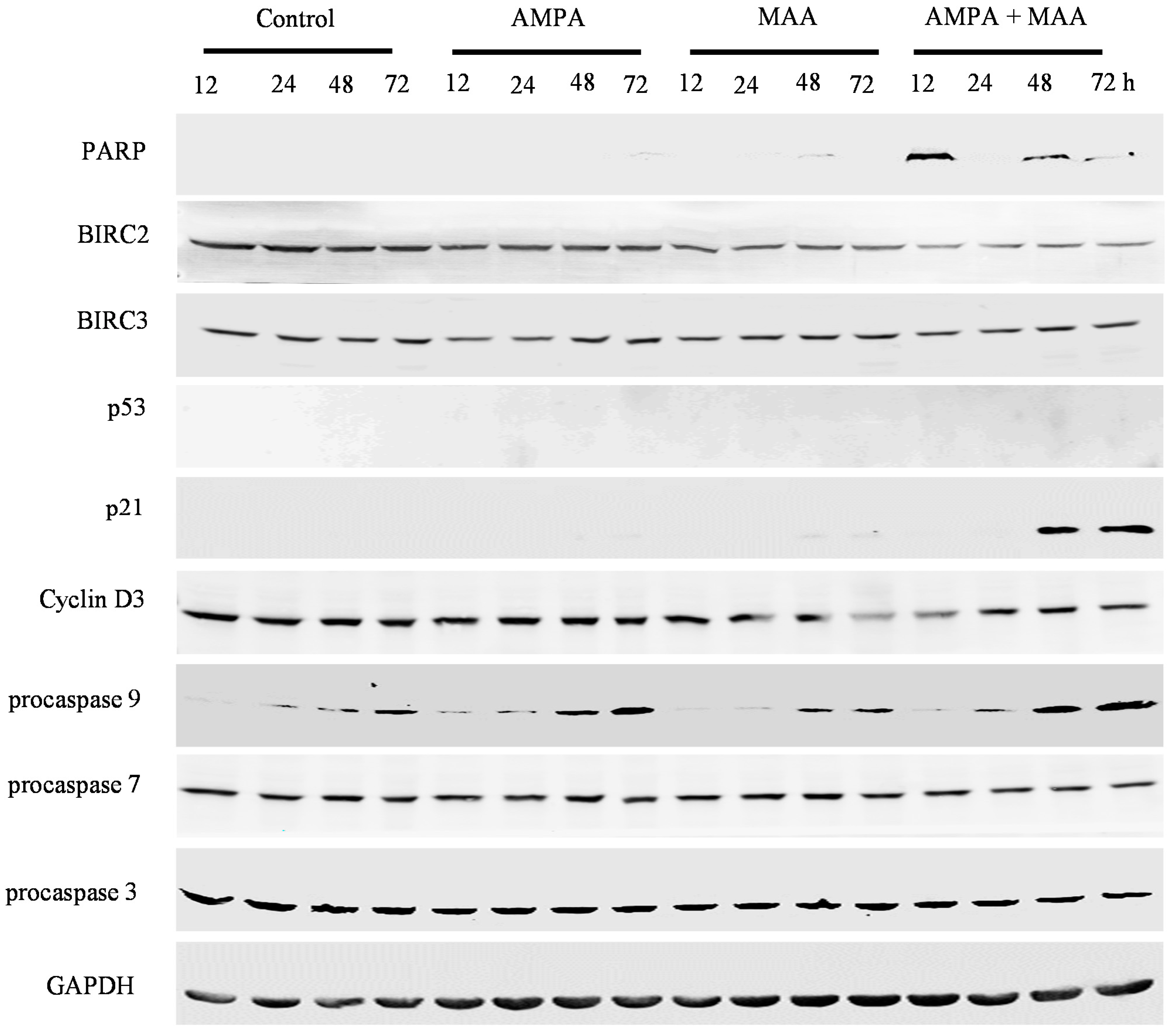
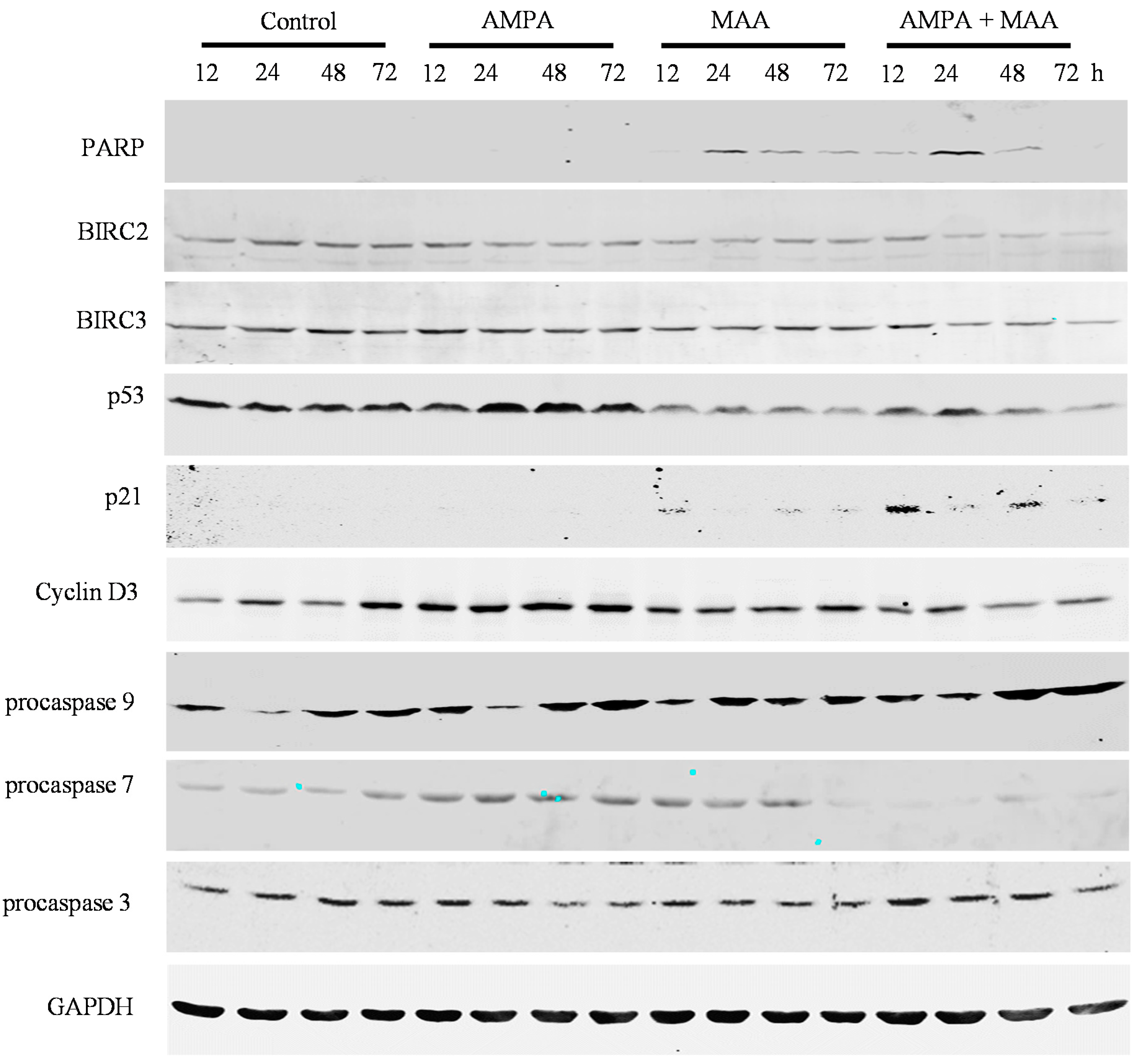
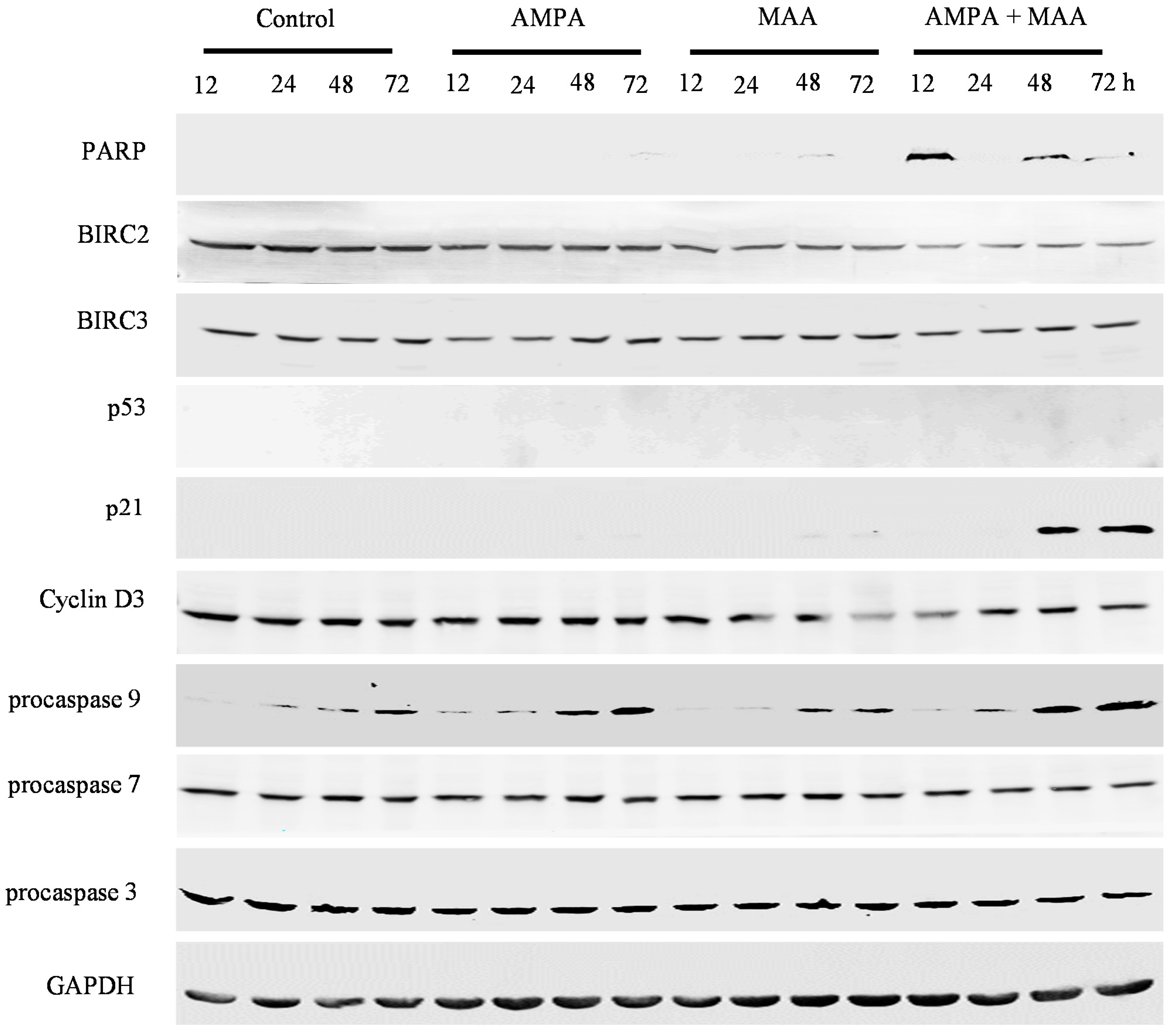
2.4. The AMPA and MAA Combination Induces Changes in the Expression Levels of Genes Involved in the Cell Cycle and Apoptosis
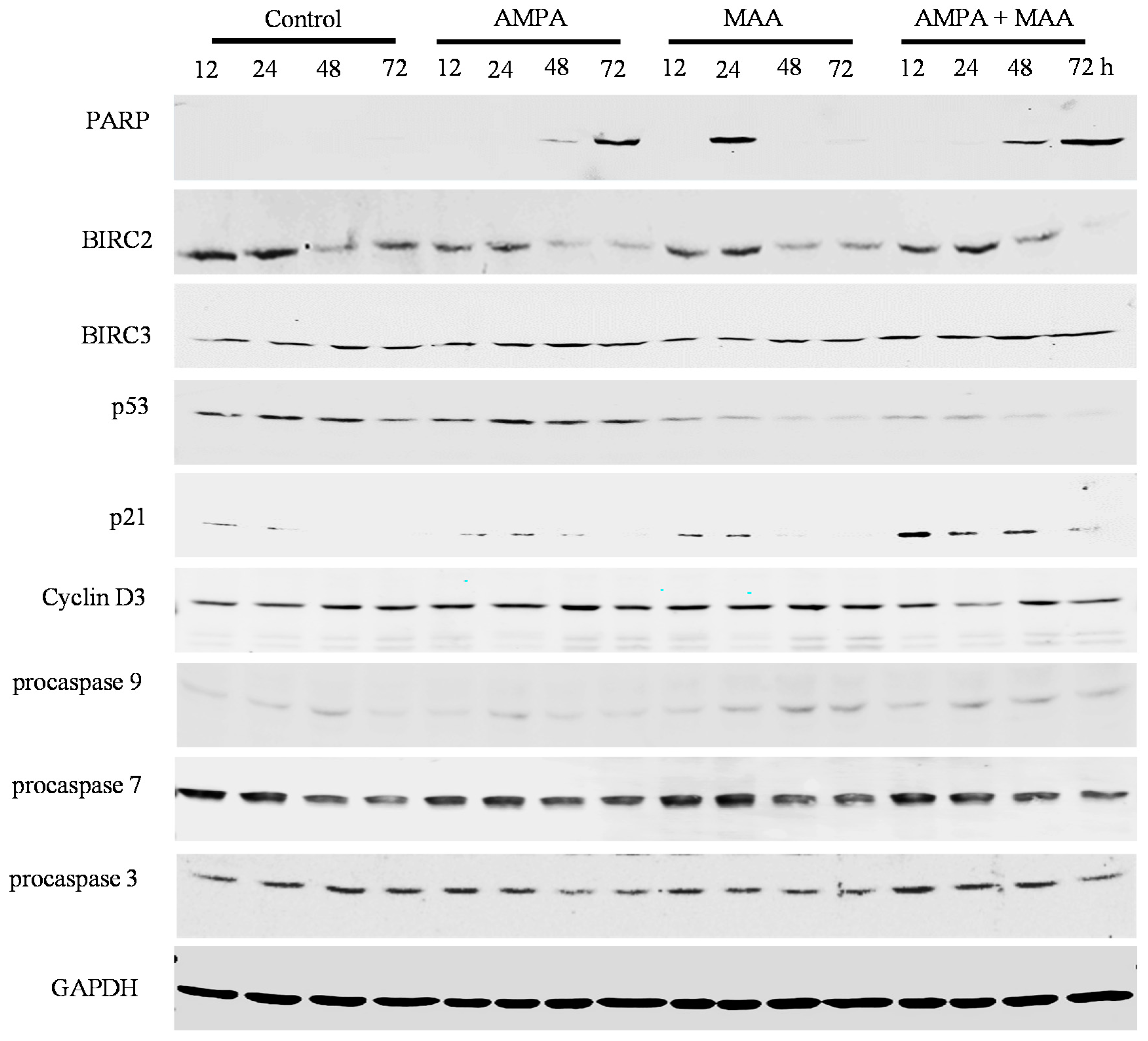
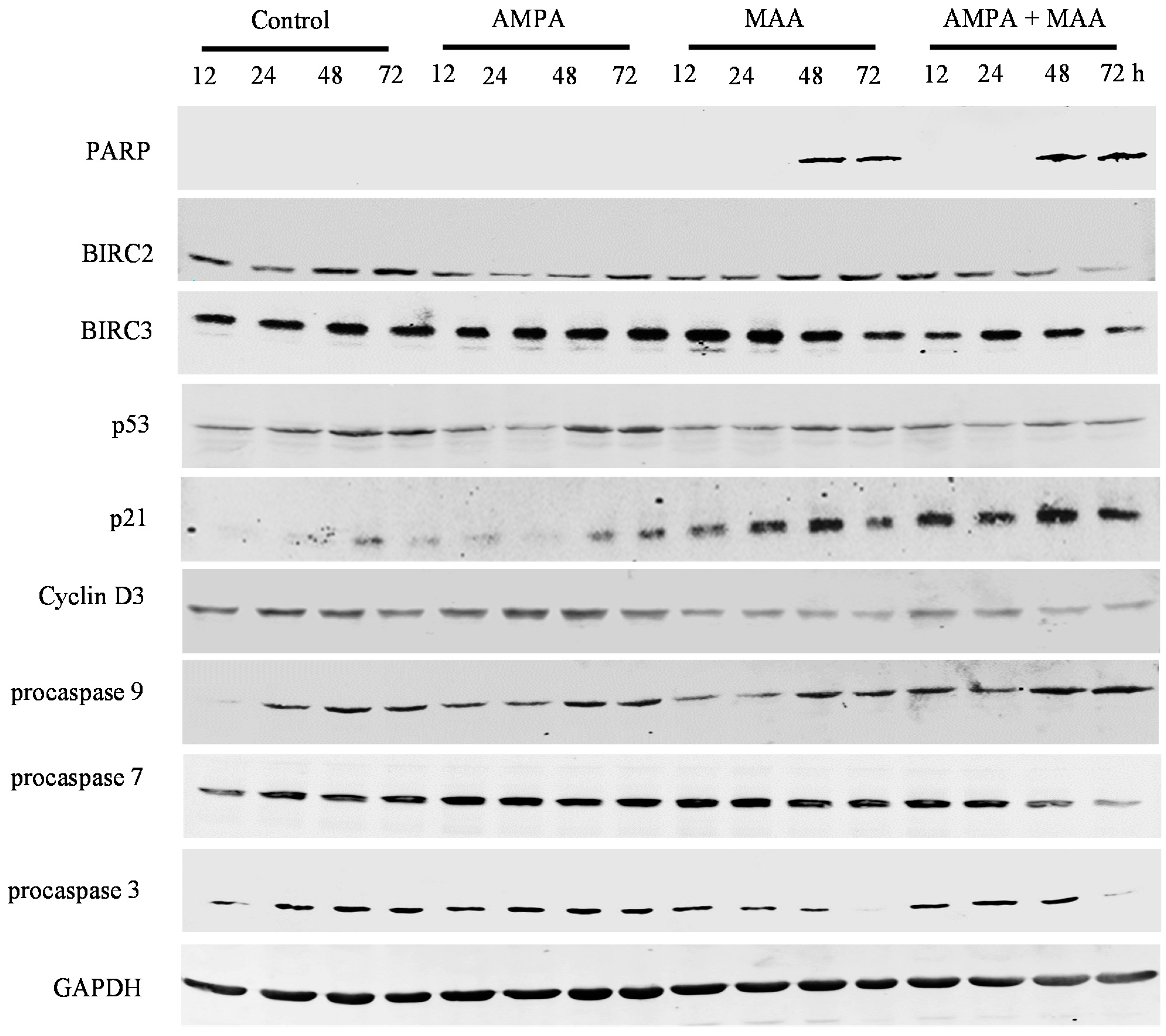

3. Experimental Section
3.1. Cell Culture
3.2. Cell Viability Assay
3.3. Detection of Apoptotic Nucleosomes
3.4. Cell Cycle Analysis
3.5. Western Blot Analysis
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kudzin, Z.H.; Gralak, D.K.; Drabowicz, J.; Luczak, J. Novel approach for the simultaneous analysis of glyphosate and its metabolites. J. Chromatogr. A 2002, 947, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Vereecken, H. Mobility and leaching of glyphosate: A review. Pest Manag. Sci. 2005, 61, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- World health organization. Glyphosate and AMPA in drinking-water. Background document for development of who guidelines for drinking-water quality. Available online: http://www.who.int/water_sanitation_health/ (accessed on 18 May 2015).
- Williams, G.M.; Kroes, R.; Munro, I.C. Safety evaluation and risk assessment of the herbicide roundup and its active ingredient, glyphosate, for humans. Regul. Toxicol. Pharmacol. 2000, 31, 117–165. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lambrechts, M.J.; Zhang, Q.; Liu, S.; Ge, D.; Yin, R.; Xi, M.; You, Z. Glyphosate and ampa inhibit cancer cell growth through inhibiting intracellular glycine synthesis. Drug Des. Dev. Ther. 2013, 7, 635–643. [Google Scholar]
- Priyandoko, D.; Ishii, T.; Kaul, S.C.; Wadhwa, R. Ashwagandha leaf derived withanone protects normal human cells against the toxicity of methoxyacetic acid, a major industrial metabolite. PLoS ONE 2011, 6, e19552. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Wine, R.N.; Chapin, R.E. 2-Methoxyacetic acid (MAA)-induced spermatocyte apoptosis in human and rat testes: An in vitro comparison. J. Androl. 1996, 17, 538–549. [Google Scholar] [PubMed]
- Bagchi, G.; Hurst, C.H.; Waxman, D.J. Interactions of methoxyacetic acid with androgen receptor. Toxicol. Appl. Pharmacol. 2009, 238, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, G.; Zhang, Y.; Waxman, D.J. Impact of methoxyacetic acid on mouse leydig cell gene expression. Reprod. Biol. Endocrinol. 2010, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Tirado, O.M.; Selva, D.M.; Toràn, N.; Suárez-Quian, C.A.; Jansen, M.; McDonnell, D.P.; Reventós, J.; Munell, F. Increased expression of estrogen receptor β in pachytene spermatocytes after short-term methoxyacetic acid administration. J. Androl. 2004, 25, 84–94. [Google Scholar] [PubMed]
- Henley, D.V.; Mueller, S.; Korach, K.S. The short-chain fatty acid methoxyacetic acid disrupts endogenous estrogen receptor-α-mediated signaling. Environ. Health Perspect. 2009, 117, 1702–1706. [Google Scholar] [PubMed]
- Bagchi, G.; Zhang, Y.; Stanley, K.A.; Waxman, D.J. Complex modulation of androgen responsive gene expression by methoxyacetic acid. Reprod. Biol. Endocrinol. 2011, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.S.; Nagel, S.C.; Miranda, P.J.; Lobenhofer, E.K.; Afshari, C.A.; McDonnell, D.P. Short-chain fatty acids enhance nuclear receptor activity through mitogen-activated protein kinase activation and histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2004, 101, 7199–7204. [Google Scholar] [CrossRef] [PubMed]
- Wade, M.G.; Kawata, A.; Williams, A.; Yauk, C. Methoxyacetic acid-induced spermatocyte death is associated with histone hyperacetylation in rats. Biol. Reprod. 2008, 78, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.; Xie, Y.; Jiang, Y.; Yingjie, Z.; Xu, W. Progress of hdac inhibitor panobinostat in the treatment of cancer. Curr. Drug Targets 2014, 15, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, K.R.; Zhang, Q.; Liu, S.; Patel, N.K.; Lu, H.; Zeng, S.X.; Wang, G.; Zhang, C.; You, Z. Methoxyacetic acid suppresses prostate cancer cell growth by inducing growth arrest and apoptosis. Am. J. Clin. Exp. Urol. 2014, 2, 300–312. [Google Scholar] [PubMed]
- Anderson, D.D.; Stover, P.J. SHMT1 and SHMT2 are functionally redundant in nuclear de novo thymidylate biosynthesis. PLoS ONE 2009, 4, e5839. [Google Scholar] [CrossRef] [PubMed]
- Porter, D.H.; Cook, R.J.; Wagner, C. Enzymatic properties of dimethylglycine dehydrogenase and sarcosine dehydrogenase from rat liver. Arch. Biochem. Biophys. 1985, 243, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Lorick, K.L.; Jensen, J.P.; Fang, S.; Ong, A.M.; Hatakeyama, S.; Weissman, A.M. Ring fingers mediate ubiquitin-conjugating enzyme (E2)-dependent ubiquitination. Proc. Natl. Acad. Sci. USA 1999, 96, 11364–11369. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; John, S.W.; Tenev, T.; Rautureau, G.J.; Hinds, M.G.; Francalanci, F.; Wilson, R.; Broemer, M.; Santoro, M.M.; Day, C.L.; et al. Card-mediated autoinhibition of cIAP1’s E3 ligase activity suppresses cell proliferation and migration. Mol. Cell 2011, 42, 569–583. [Google Scholar] [CrossRef]
- Huang, Y.; Park, Y.C.; Rich, R.L.; Segal, D.; Myszka, D.G.; Wu, H. Structural basis of caspase inhibition by XIAP: Differential roles of the linker versus the BIR domain. Cell 2001, 104, 781–790. [Google Scholar] [PubMed]
- Sartor, O.; Gillessen, S. Treatment sequencing in metastatic castrate-resistant prostate cancer. Asian J. Androl. 2014, 16, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Brinkworth, M.H.; Weinbauer, G.F.; Schlatt, S.; Nieschlag, E. Identification of male germ cells undergoing apoptosis in adult rats. J. Reprod. Fertil. 1995, 105, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Wine, R.N.; Miller, D.S.; Reece, J.M.; Smith, M.; Chapin, R.E. Protection against methoxyacetic-acid-induced spermatocyte apoptosis with calcium channel blockers in cultured rat seminiferous tubules: Possible mechanisms. Toxicol. Appl. Pharmacol. 1997, 144, 105–119. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Shi, X.B.; DuRaine, G.; Haudenschild, D.; Tepper, C.G.; Lo, S.H.; Gandour-Edwards, R.; de Vere White, R.W.; Reddi, A.H. Interleukin-17 receptor-like gene is a novel antiapoptotic gene highly expressed in androgen-independent prostate cancer. Cancer Res. 2006, 66, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Scheibye-Knudsen, M.; Brace, L.E.; Kassahun, H.; SenGupta, T.; Nilsen, H.; Mitchell, J.R.; Croteau, D.L.; Bohr, V.A. Defective mitophagy in XPA via PARP-1 hyperactivation and NAD(+)/SIRT1 reduction. Cell 2014, 157, 882–896. [Google Scholar] [CrossRef] [PubMed]
- De Almagro, M.C.; Vucic, D. The inhibitor of apoptosis (IAP) proteins are critical regulators of signaling pathways and targets for anti-cancer therapy. Exp. Oncol. 2012, 34, 200–211. [Google Scholar] [PubMed]
- Jin, S.; Kalkum, M.; Overholtzer, M.; Stoffel, A.; Chait, B.T.; Levine, A.J. cIAP1 and the serine protease HTRA2 are involved in a novel p53-dependent apoptosis pathway in mammals. Genes Dev. 2003, 17, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fang, S.; Jensen, J.P.; Weissman, A.M.; Ashwell, J.D. Ubiquitin protein ligase activity of IAPS and their degradation in proteasomes in response to apoptotic stimuli. Science 2000, 288, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Liu, L.; Teng, X.; Mukai-Sakai, R.; Shimada, H.; Kaji, R.; Mitani, T.; Matsumoto, M.; Toida, K.; Ishimura, K.; et al. Nucling recruits APAF-1/pro-caspase-9 complex for the induction of stress-induced apoptosis. J. Biol. Chem. 2004, 279, 41131–41140. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Shaha, C. N-Acetylcysteine prevents MAA induced male germ cell apoptosis: Role of glutathione and cytochrome c. FEBS Lett. 2002, 527, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Eckelman, B.P.; Salvesen, G.S. The human anti-apoptotic proteins cIAP1 and cIAP2 bind but do not inhibit caspases. J. Biol. Chem. 2006, 281, 3254–3260. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.P.; Smith, L.; Smith, J.B. cIAP1 cooperatively inhibits procaspase-3 activation by the caspase-9 apoptosome. J. Biol. Chem. 2010, 285, 30061–30068. [Google Scholar] [CrossRef] [PubMed]
- Alimirah, F.; Chen, J.; Basrawala, Z.; Xin, H.; Choubey, D. DU-145 and PC-3 human prostate cancer cell lines express androgen receptor: Implications for the androgen receptor functions and regulation. FEBS Lett. 2006, 580, 2294–2300. [Google Scholar] [CrossRef] [PubMed]
- Van Bokhoven, A.; Varella-Garcia, M.; Korch, C.; Johannes, W.U.; Smith, E.E.; Miller, H.L.; Nordeen, S.K.; Miller, G.J.; Lucia, M.S. Molecular characterization of human prostate carcinoma cell lines. Prostate 2003, 57, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Dong, J.; Berg, A.H. Identification and characterization of membrane androgen receptors in the ZIP9 zinc transporter subfamily: II. Role of human ZIP9 in testosterone-induced prostate and breast cancer cell apoptosis. Endocrinology 2014, 155, 4250–4265. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, N.; Charalampopoulos, I.; Anagnostopoulou, V.; Konstantinidis, G.; Föller, M.; Gravanis, A.; Alevizopoulos, K.; Lang, F.; Stournaras, C. Membrane androgen receptor activation triggers down-regulation of PI-3k/Akt/NF-κB activity and induces apoptotic responses via Bad, FasL and caspase-3 in DU145 prostate cancer cells. Mol. Cancer 2008, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Kampa, M.; Papakonstanti, E.A.; Hatzoglou, A.; Stathopoulos, E.N.; Stournaras, C.; Castanas, E. The human prostate cancer cell line lncap bears functional membrane testosterone receptors that increase PSA secretion and modify actin cytoskeleton. FASEB J. 2002, 16, 1429–1431. [Google Scholar] [PubMed]
- Kampa, M.; Kogia, C.; Theodoropoulos, P.A.; Anezinis, P.; Charalampopoulos, I.; Papakonstanti, E.A.; Stathopoulos, E.N.; Hatzoglou, A.; Stournaras, C.; Gravanis, A.; et al. Activation of membrane androgen receptors potentiates the antiproliferative effects of paclitaxel on human prostate cancer cells. Mol. Cancer Ther. 2006, 5, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, N.; Papakonstanti, E.A.; Kallergi, G.; Alevizopoulos, K.; Stournaras, C. Membrane androgen receptor activation in prostate and breast tumor cells: Molecular signaling and clinical impact. IUBMB Life 2009, 61, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Alevizopoulos, K.; Stournaras, C. Targeting membrane androgen receptors in tumors. Expert Opin. Ther. Targets 2013, 17, 951–963. [Google Scholar] [CrossRef] [PubMed]
- You, Z.; Saims, D.; Chen, S.; Zhang, Z.; Guttridge, D.C.; Guan, K.L.; MacDougald, O.A.; Brown, A.M.; Evan, G.; Kitajewski, J.; et al. Wnt signaling promotes oncogenic transformation by inhibiting c-Myc-induced apoptosis. J. Cell Biol. 2002, 157, 429–440. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parajuli, K.R.; Zhang, Q.; Liu, S.; You, Z. Aminomethylphosphonic Acid and Methoxyacetic Acid Induce Apoptosis in Prostate Cancer Cells. Int. J. Mol. Sci. 2015, 16, 11750-11765. https://doi.org/10.3390/ijms160511750
Parajuli KR, Zhang Q, Liu S, You Z. Aminomethylphosphonic Acid and Methoxyacetic Acid Induce Apoptosis in Prostate Cancer Cells. International Journal of Molecular Sciences. 2015; 16(5):11750-11765. https://doi.org/10.3390/ijms160511750
Chicago/Turabian StyleParajuli, Keshab R., Qiuyang Zhang, Sen Liu, and Zongbing You. 2015. "Aminomethylphosphonic Acid and Methoxyacetic Acid Induce Apoptosis in Prostate Cancer Cells" International Journal of Molecular Sciences 16, no. 5: 11750-11765. https://doi.org/10.3390/ijms160511750
APA StyleParajuli, K. R., Zhang, Q., Liu, S., & You, Z. (2015). Aminomethylphosphonic Acid and Methoxyacetic Acid Induce Apoptosis in Prostate Cancer Cells. International Journal of Molecular Sciences, 16(5), 11750-11765. https://doi.org/10.3390/ijms160511750