
Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes

 ,
,
Abstract
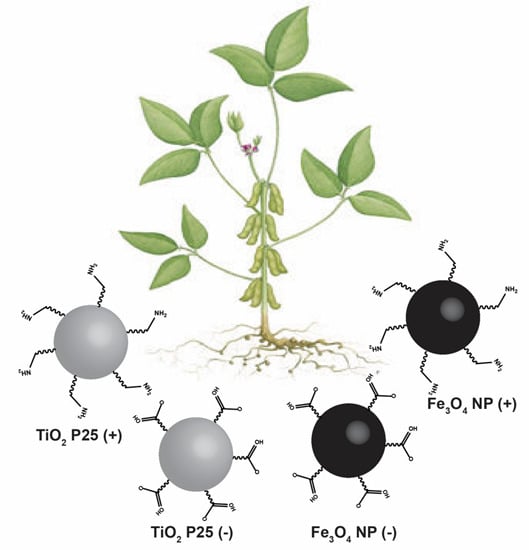
:
1. Introduction
2. Results and Discussion
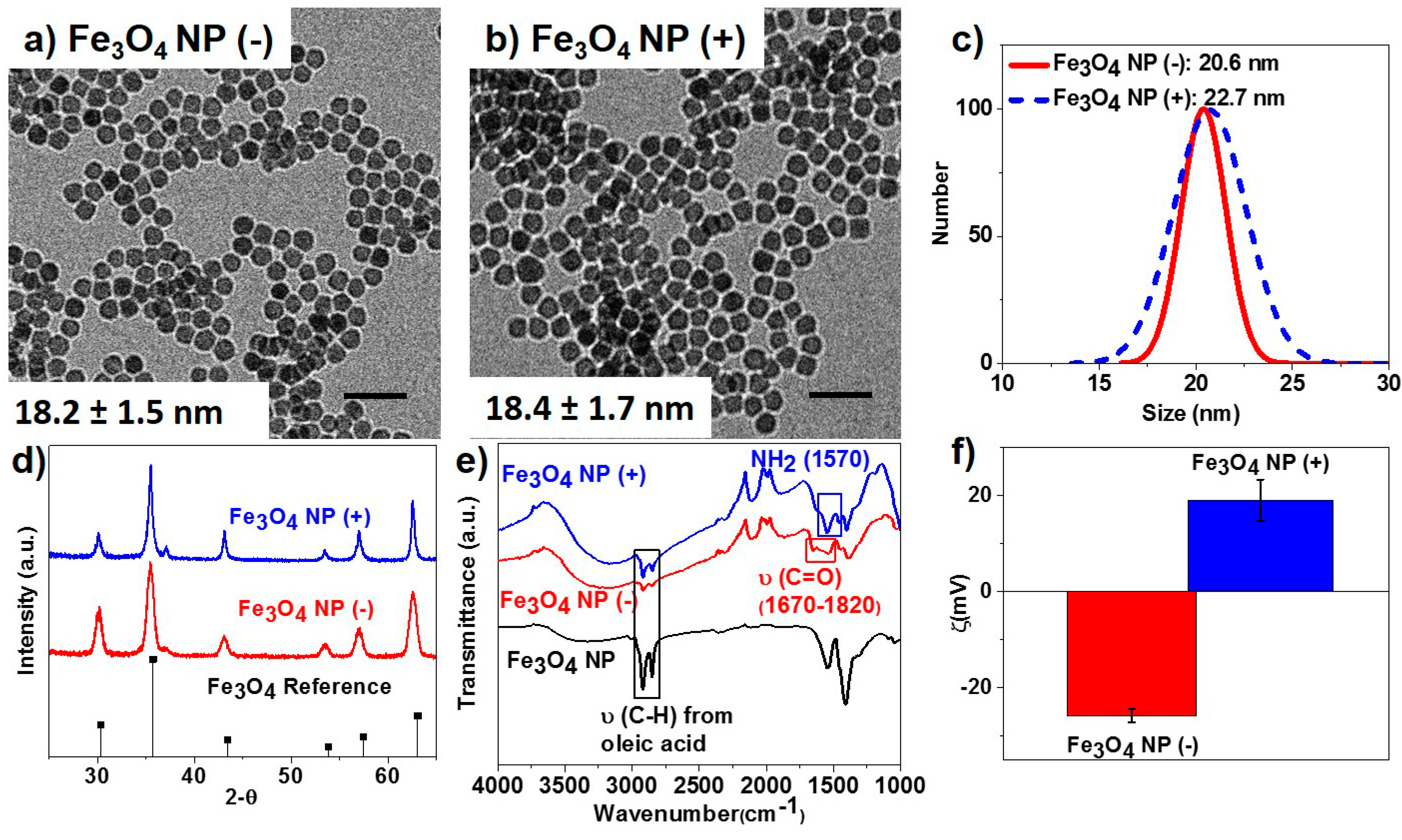
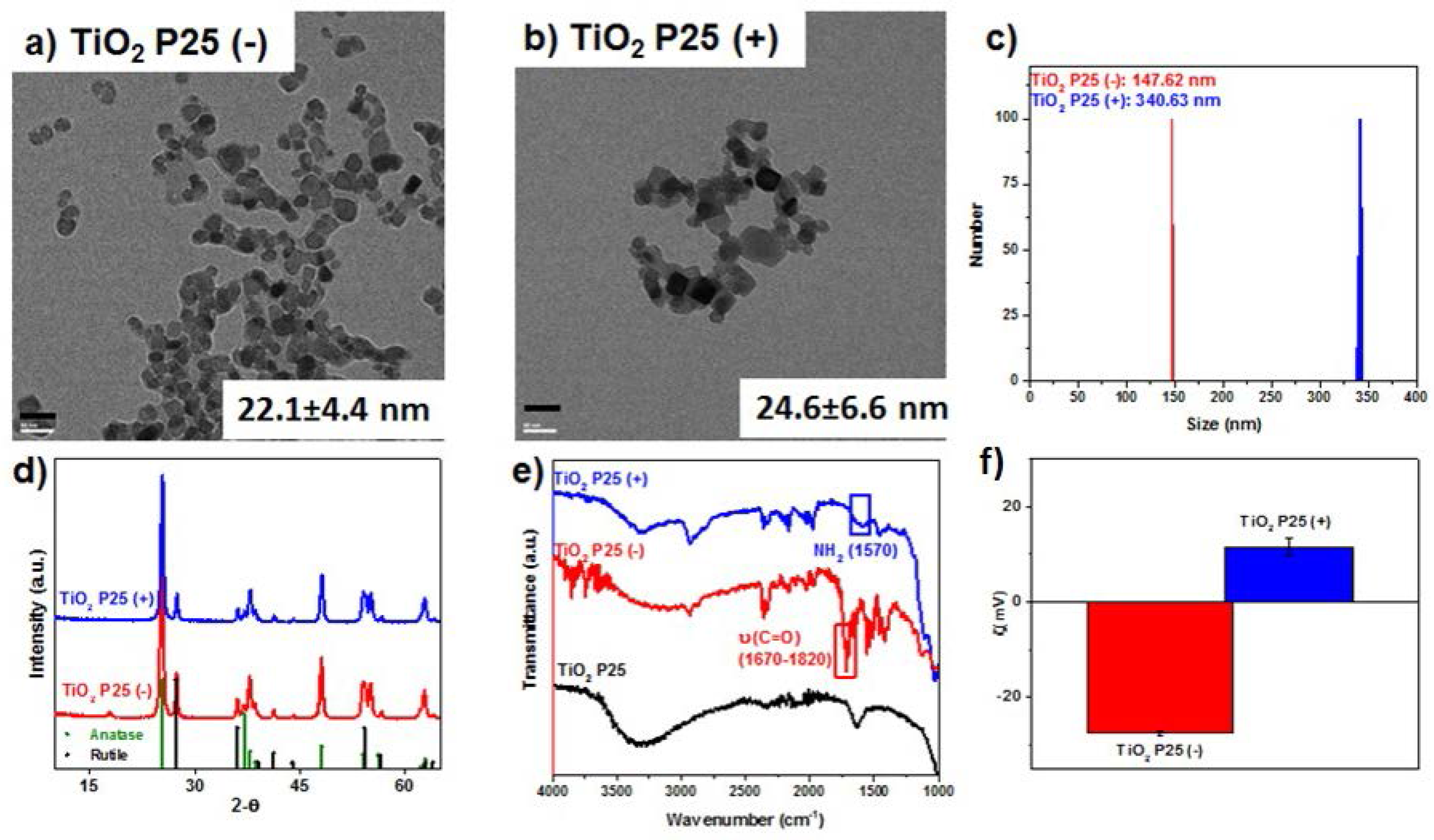
2.1. Nanoparticle Synthesis and Surface Functionalization
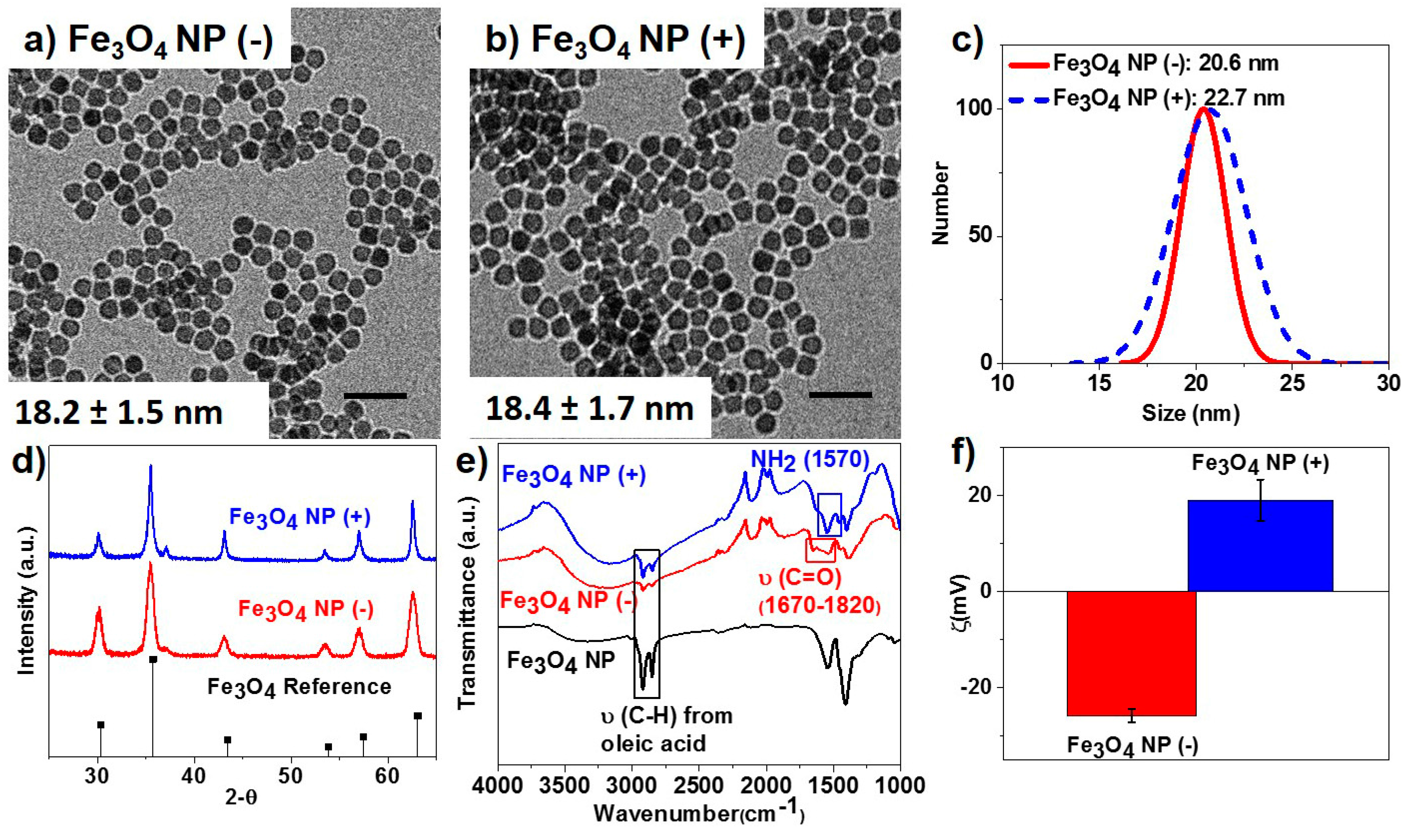
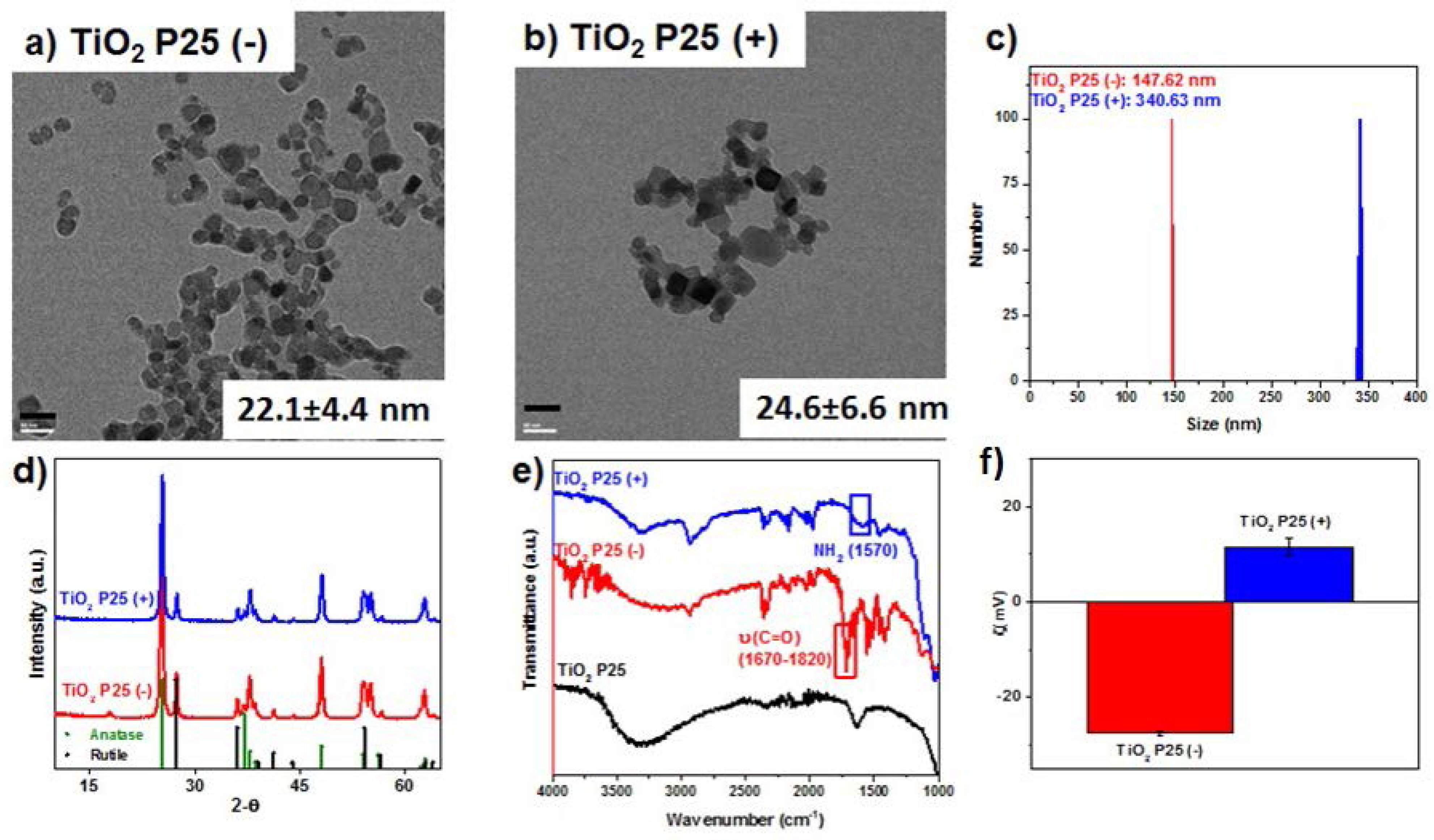
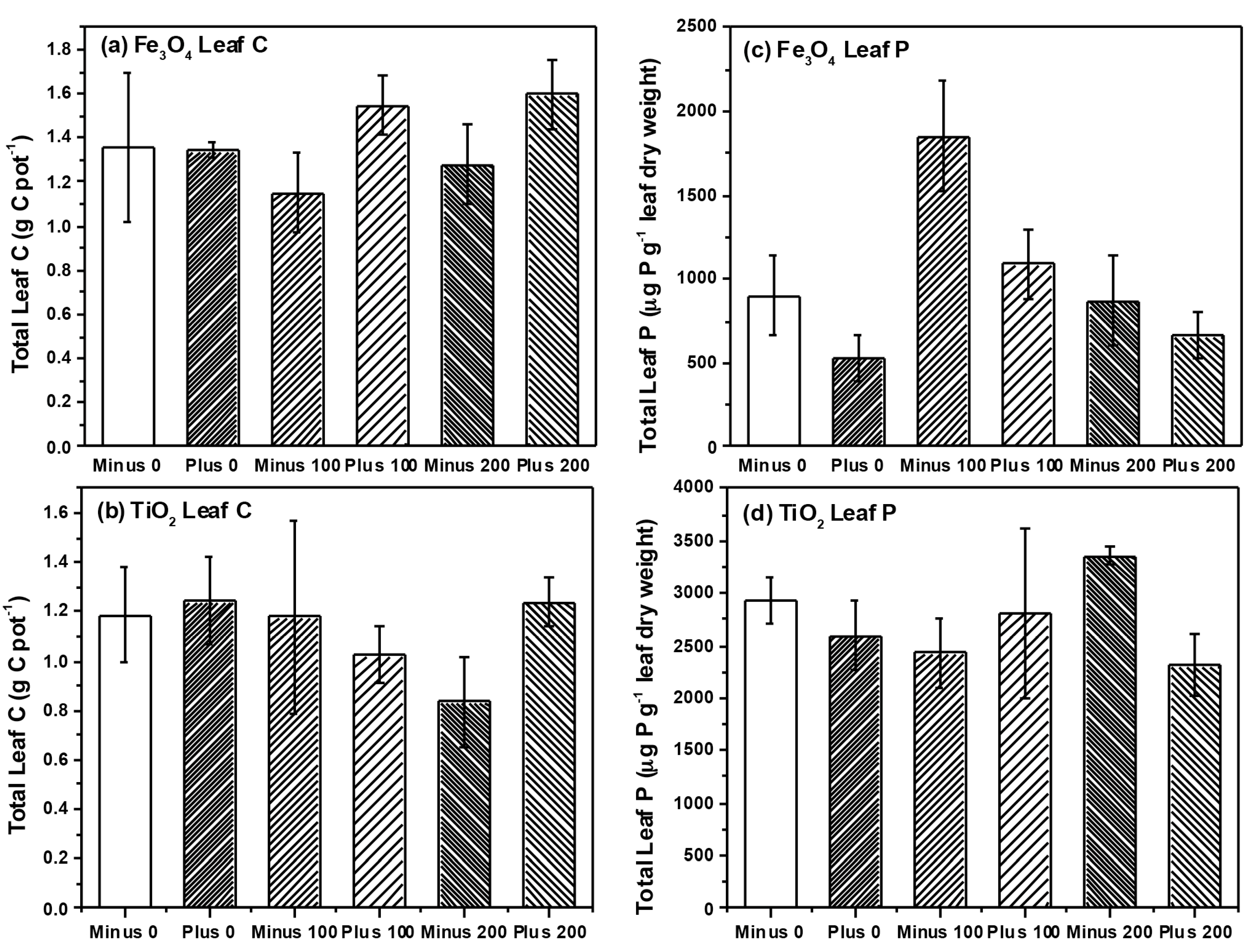
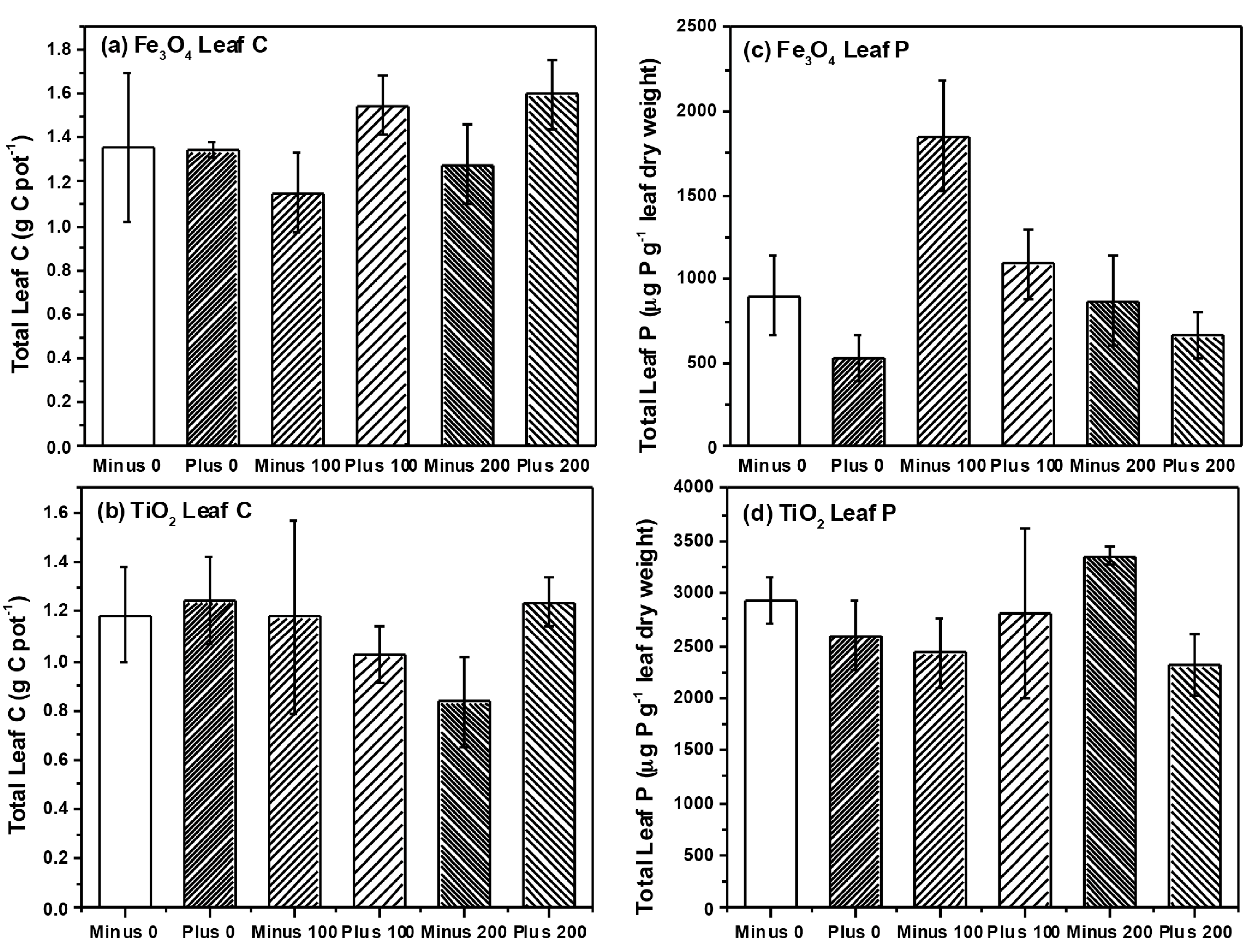
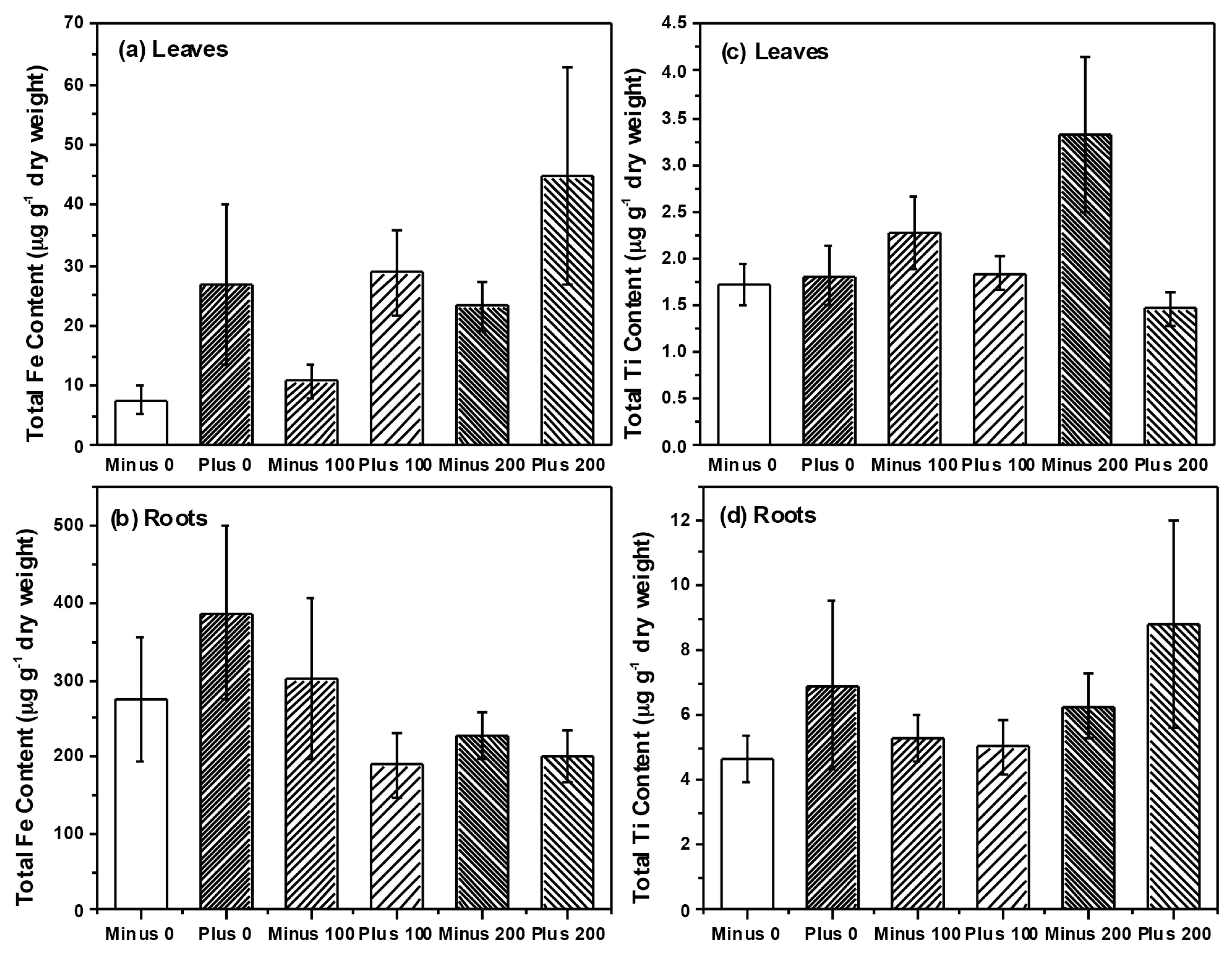
2.2. Plant Growth and Nutrient Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
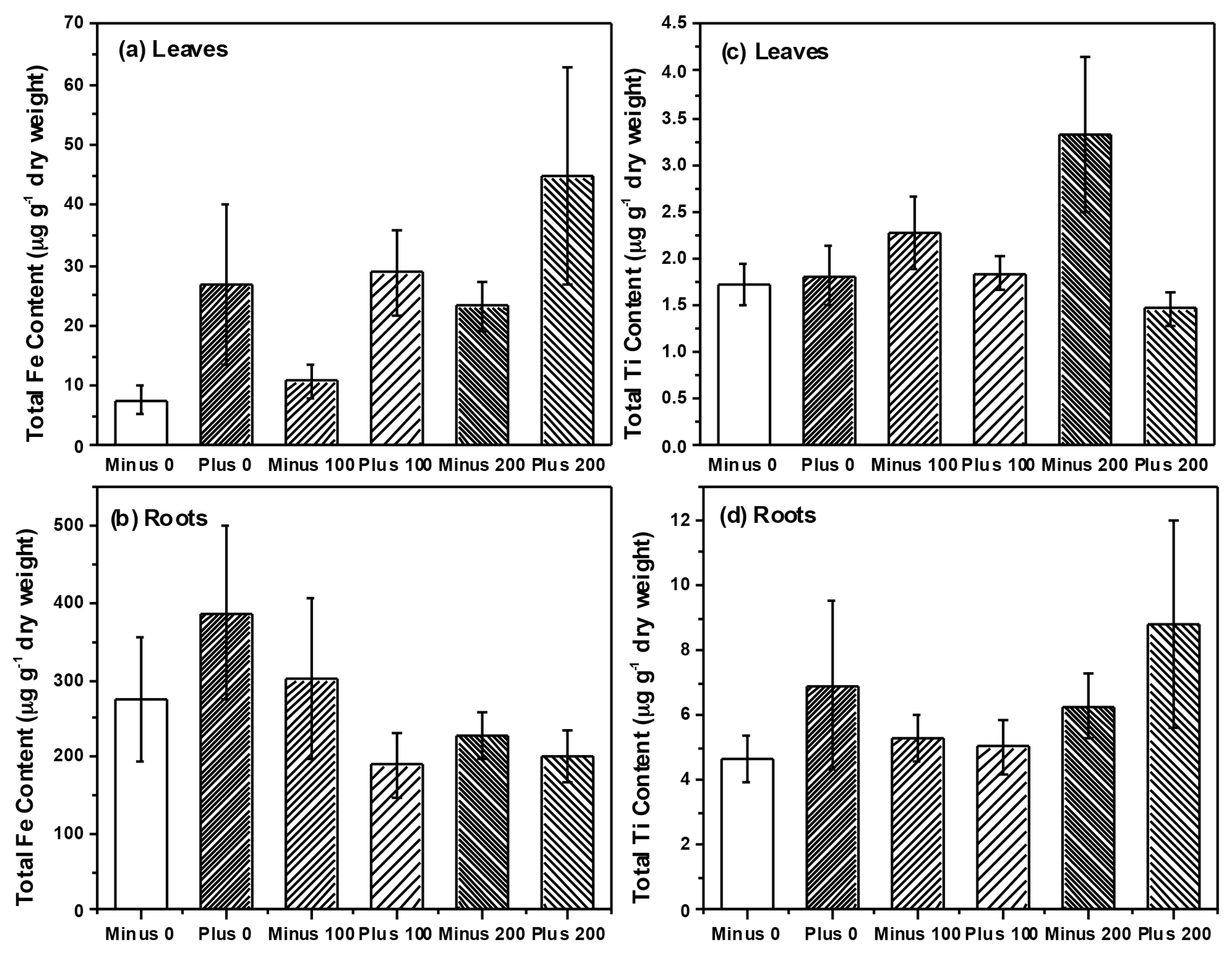{kind=link}
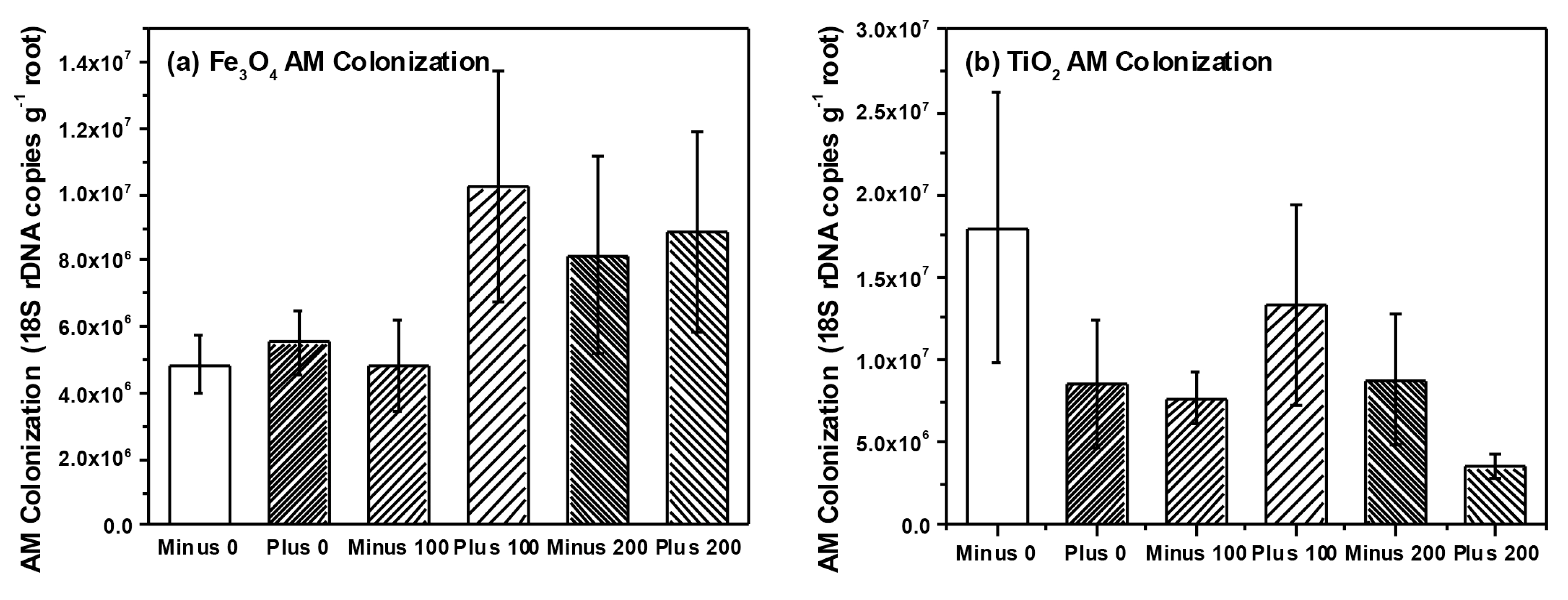{kind=link}
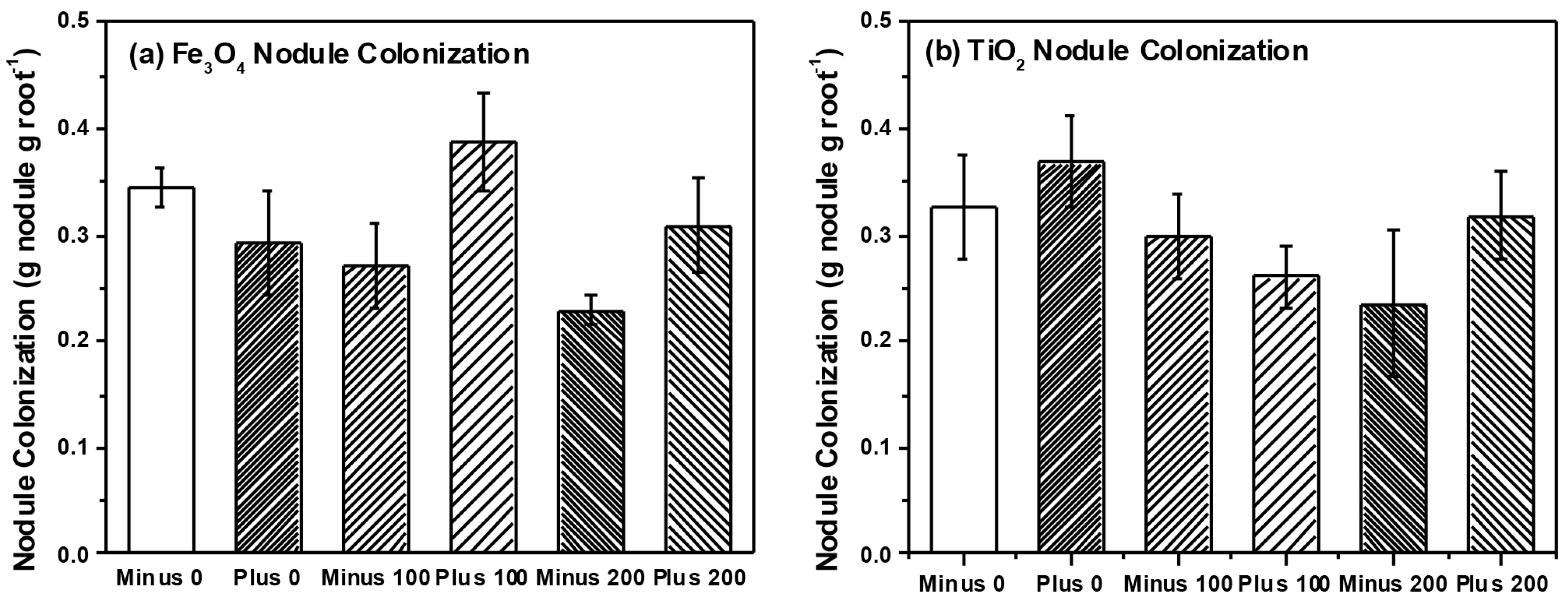{kind=link}
| Source of Variation | Above Wt 1 | Root Wt | Nodule Wt | Nodule/Root 2 | AM Col 3 | C 4 | N | P |
|---|---|---|---|---|---|---|---|---|
| Metal Type | 0.013 | 0.078 | 0.222 | 0.866 | 0.199 | 0.069 | 0.808 | <0.001 |
| Metal Concentration | 0.730 | 0.618 | 0.750 | 0.157 | 0.750 | 0.465 | 0.662 | 0.370 |
| Metal Charge | 0.373 | 0.228 | 0.082 | 0.073 | 0.878 | 0.110 | 0.938 | 0.047 |
| Metal × Concentration | 0.587 | 0.590 | 0.722 | 0.421 | 0.163 | 0.240 | 0.641 | 0.093 |
| Metal × Charge | 0.591 | 0.945 | 0.777 | 0.511 | 0.245 | 0.242 | 0.327 | 0.769 |
| Concentration × Charge | 0.182 | 0.317 | 0.396 | 0.559 | 0.171 | 0.194 | 0.570 | 0.665 |
| Metal × Conc. × charge | 0.100 | 0.972 | 0.322 | 0.225 | 0.630 | 0.234 | 0.977 | 0.134 |
| Source of Variation | Above Wt 1 | Root Wt | Nodule Wt | Nodule/Root 2 | AM Col 3 | C 4 | N | P |
|---|---|---|---|---|---|---|---|---|
| Fe3O4 Concentration | 0.772 | 0.647 | 0.749 | 0.257 | 0.842 | 0.528 | 0.778 | 0.003 |
| Fe3O4 Charge | 0.284 | 0.494 | 0.152 | 0.057 | 0.188 | 0.010 | 0.387 | 0.018 |
| Concentration × Charge | 0.138 | 0.590 | 0.307 | 0.258 | 0.604 | 0.887 | 0.507 | 0.442 |
| TiO2 Concentration | 0.578 | 0.521 | 0.719 | 0.267 | 0.156 | 0.297 | 0.631 | 0.899 |
| TiO2 Charge | 0.814 | 0.252 | 0.309 | 0.462 | 0.412 | 0.805 | 0.547 | 0.377 |
| Concentration × Charge | 0.159 | 0.475 | 0.441 | 0.460 | 0.479 | 0.157 | 0.863 | 0.350 |
2.3. Analysis of Microbial Communities
| Variable | Source of Variation | DF | SS | MS | F | p |
|---|---|---|---|---|---|---|
| Fe | Plant Tissue | 1 | 24.306 | 24.306 | 216.747 | <0.001 |
| Fe3O4 Concentration | 2 | 0.138 | 0.0691 | 0.616 | 0.543 | |
| Fe3O4 Charge | 1 | 0.379 | 0.379 | 3.381 | 0.071 | |
| Tissue × Concentration | 2 | 0.624 | 0.312 | 2.783 | 0.070 | |
| Tissue × Charge | 1 | 0.496 | 0.496 | 4.424 | 0.040 | |
| Concentration × Charge | 2 | 0.293 | 0.147 | 1.308 | 0.278 | |
| Tissue × Conc. × Charge | 2 | 0.182 | 0.0911 | 0.812 | 0.449 | |
| Residual | 60 | 6.728 | 0.112 | |||
| Total | 71 | 33.148 | 0.467 | |||
| Ti | Plant Tissue | 1 | 275.862 | 275.862 | 25.317 | <0.001 |
| TiO2 Concentration | 2 | 23.532 | 11.766 | 1.080 | 0.347 | |
| TiO2 Charge | 1 | 2.492 | 2.492 | 0.229 | 0.634 | |
| Tissue × Concentration | 2 | 10.868 | 5.434 | 0.499 | 0.610 | |
| Tissue × Charge | 1 | 20.869 | 20.869 | 1.915 | 0.172 | |
| Concentration × Charge | 2 | 6.466 | 3.233 | 0.297 | 0.744 | |
| Tissue × Conc. × Charge | 2 | 11.798 | 5.899 | 0.541 | 0.585 | |
| Residual | 55 | 599.305 | 10.896 | |||
| Total | 66 | 971.381 | 14.718 |
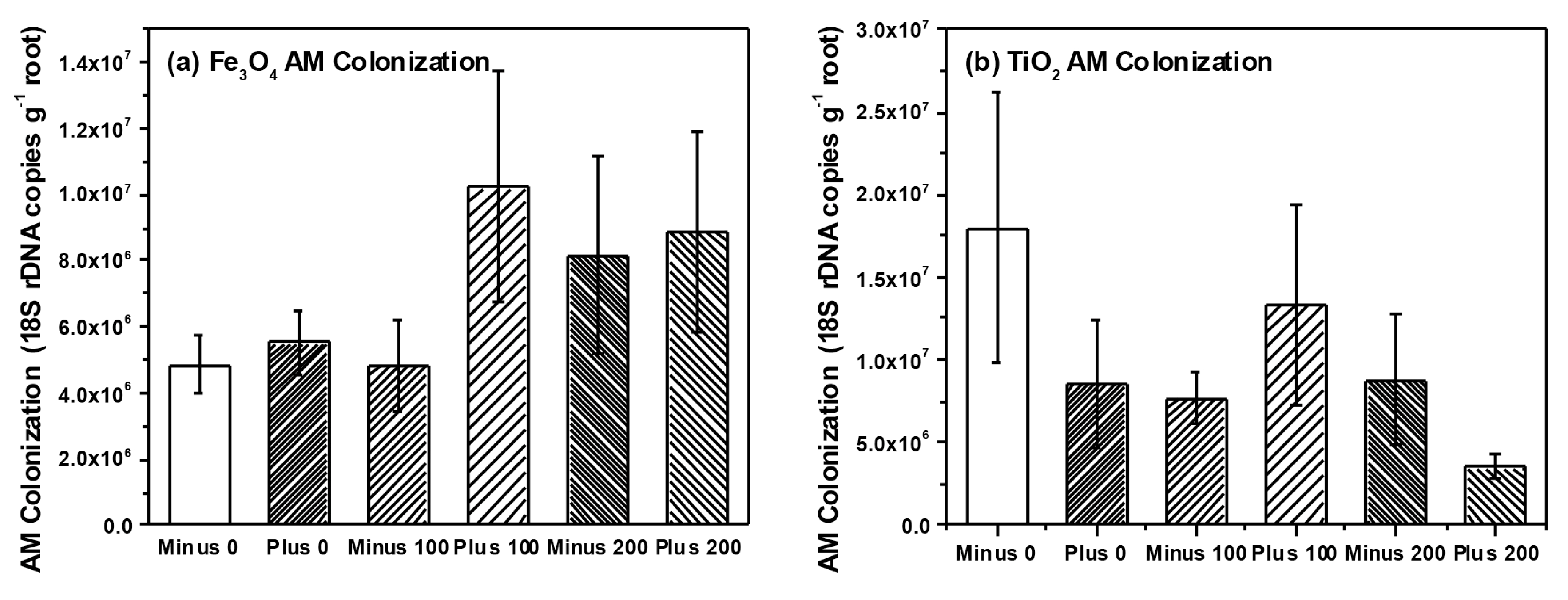
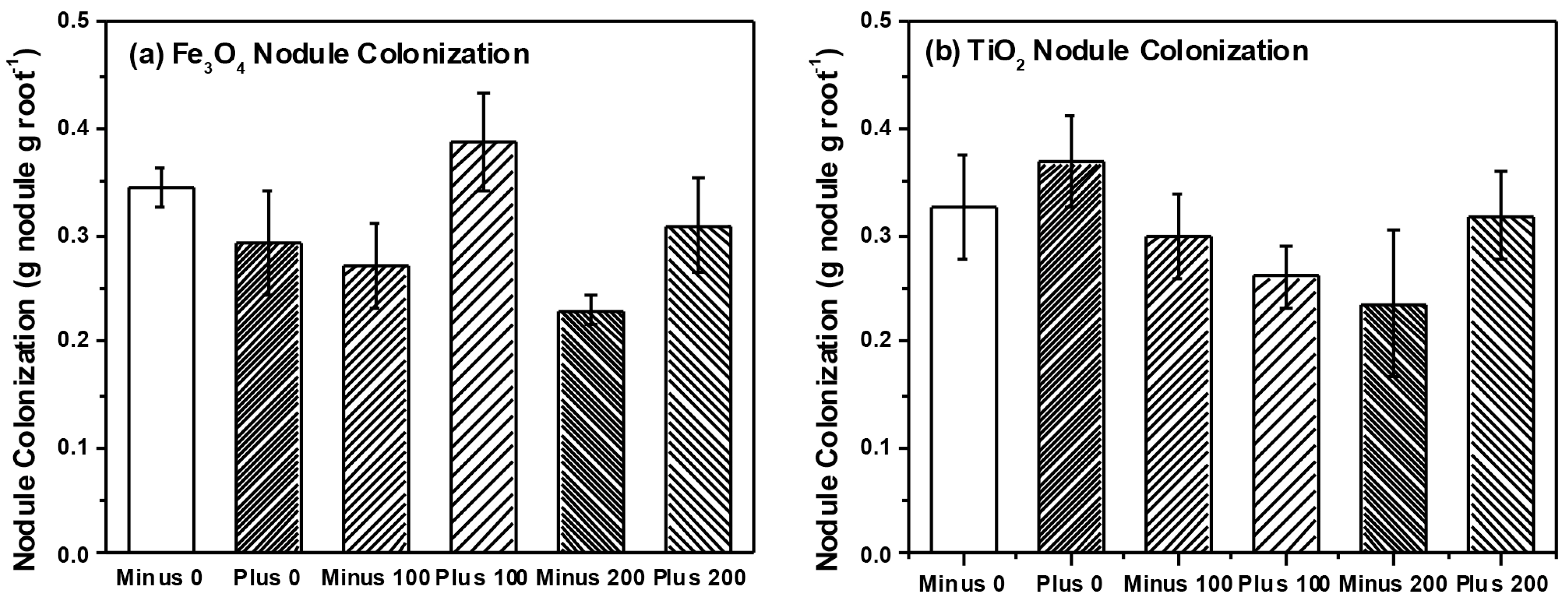
3. Experimental Section
3.1. Materials
3.2. Experimental Set-up and Plant/Soil Sampling
3.3. Nanoparticle Synthesis and Characterization
3.3.1. Synthesis of Magnetite Nanoparticles (Fe3O4 NPs)
3.3.2. Functionalization of Fe3O4 NPs to Introduce Surface Charges
3.3.3. Functionalization of Degussa TiO2 P25 NPs to Introduce Surface Charges
3.3.4. Characterization
3.4. Plant Growth and Elemental Analysis
3.5. Analysis of Microbial Communities
3.6. Data and Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ma, X.; Geisler-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Benn, T.M.; Westerhoff, P. Nanoparticle silver released into water from commercially available sock fabrics. Environ. Sci. Technol. 2008, 42, 4133–4139. [Google Scholar] [CrossRef] [PubMed]
- Klaine, S.J.; Alvarez, P.J.J.; Batley, G.E.; Fernandes, T.F.; Handy, R.D.; Lyon, D.Y.; Mahendra, S.; McLaughlin, M.J.; Lead, J.R. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ. Toxicol. Chem. 2008, 27, 1825. [Google Scholar] [CrossRef] [PubMed]
- Sobha, K.; Surendranath, K.; Meena, V.; Jwala, T.K.; Swetha, N.; Latha, K.S.M. Emerging trends in nanobiotechnology. In Biotechnology and Molecular Biology Reviews; Academic Journals: Nairobi, Kenya, 2010; Volume 4, pp. 1–12. [Google Scholar]
- Rai, M.; Ingle, A. Role of nanotechnology in agriculture with special reference to management of insect pests. Appl. Microbiol. Biotechnol. 2012, 94, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Throbäck, I.N.; Johansson, M.; Rosenquist, M.; Pell, M.; Hansson, M.; Hallin, S. Silver (Ag+) reduces denitrification and induces enrichment of novel nirk genotypes in soil. FEMS Microbiol. Lett. 2007, 270, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Priester, J.H.; Ge, Y.; Mielke, R.E.; Horst, A.M.; Moritz, S.C.; Espinosa, K.; Gelb, J.; Walker, S.L.; Nisbet, R.M.; An, Y.-J.; et al. Soybean susceptibility to manufactured nanomaterials with evidence for food quality and soil fertility interruption. Proc. Natl. Acad. Sci. USA 2012, 109, E2451–E2456. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Bandyopadhyay, M.; Mukherjee, A. Genotoxicity of titanium dioxide (TiO2) nanoparticles at two trophic levels: Plant and human lymphocytes. Chemosphere 2010, 81, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Asli, S.; Neumann, P.M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Schimel, J.P.; Holden, P.A. Identification of soil bacteria susceptible to TiO2 and ZnO nanoparticles. Appl. Environ. Microb. 2012, 78, 6749–6758. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.J.; Zhu, S.; Pablico-Lansigan, M.P.; Hewins, C.R.; Samia, A.C.S. Titanium oxide nanoparticle effects on composition of soil microbial communities and plant performance. Biol. Fertil. Soils 2014, 50, 1169–1173. [Google Scholar] [CrossRef]
- Boonyanitipong, P.K.B.; Kumar, P.; Baruah, S.; Dutta, J. Toxicity of ZnO and TiO2 nanoparticles on germinating rice seed oryza sativa L. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 282–285. [Google Scholar] [CrossRef]
- Kim, E.; Kim, S.-H.; Kim, H.-C.; Lee, S.G.; Lee, S.J.; Jeong, S.W. Growth inhibition of aquatic plant caused by silver and titanium oxide nanoparticles. ToxEHS 2011, 3, 1–6. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.J.; Kwak, J.I.; Lee, W.M.; Holden, P.A.; An, Y.J. Zinc oxide nanoparticles delay soybean development: A standard soil microcosm study. Ecotoxicol. Environ. Saf. 2014, 100, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Priester, J.H.; van de Werfhorst, L.C.; Walker, S.L.; Nisbet, R.M.; An, Y.J.; Schimel, J.P.; Gardea-Torresdey, J.L.; Holden, P.A. Soybean plants modify metal oxide nanoparticle effects on soil bacterial communities. Environ. Sci. Technol. 2014, 48, 13489–13496. [Google Scholar] [CrossRef] [PubMed]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef] [PubMed]
- Arruda, S.C.C.; Silva, A.L.D.; Galazzi, R.M.; Azevedo, R.A.; Arruda, M.A.Z. Nanoparticles applied to plant science: A review. Talanta 2015, 131, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Emami-Karvani, Z.; Chehrazi, P. Antibacterial activity of ZnO nanoparticle on gram-positive and gram-negative bacteria. Afr. J. Microbiol. Res. 2011, 5, 1368–1373. [Google Scholar]
- Murata, T.; Kanao-Koshikawa, M.; Takamatsu, T. Effects of Pb, Cu, Sb, In and Ag contamination on the proliferation of soil bacterial colonies, soil dehydrogenase activity, and phospholipid fatty acid profiles of soil microbial communities. Water Air Soil Pollut. 2005, 164, 103–118. [Google Scholar] [CrossRef]
- Yu, W.W.; Falkner, J.C.; Yavuz, C.T.; Colvin, V.L. Synthesis of monodisperse iron oxide nanocrystals by thermal decomposition of iron carboxylate salts. Chem. Commun. 2004, 2306, 2306–2307. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Viezcas, J.A.; Castillo-Michel, H.; Andrews, J.C.; Cotte, M.; Rico, C.; Peralta-Videa, J.R.; Ge, Y.; Priester, J.H.; Holden, P.A.; Gardea-Torresdey, J.L. In situ synchrotron X-ray fluorescence mapping and speciation of CeO2 and ZnO nanoparticles in soil cultivated soybean (Glycine max). ACS Nano 2013, 7, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gomez, C.; Babin, M.; Obrador, A.; Alvarez, J.M.; Fernandez, M.D. Integrating ecotoxicity and chemical approaches to compare the effects of ZnO nanoparticles, ZnO bulk, and ZnCl2 on plants and microorganisms in a natural soil. Environ. Sci. Pollut. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, C.; Wen, J.; Wu, G.; Tao, M. Research of the effect of nanometer materials on germination and growth enhancement of Glycine max and its mechanism. Soybean Sci. 2002, 21, 168–171. [Google Scholar]
- Gao, F.; Hong, F.; Liu, C.; Zheng, L.; Su, M.; Wu, X.; Yang, F.; Wu, C.; Yang, P. Mechanism of nano-anatase TiO2 on promoting photosynthetic carbon reaction of spinach: Inducing complex of rubisco-rubisco activase. Biol. Trace Elem. Res. 2006, 111, 239–253. [Google Scholar] [CrossRef]
- Yang, F.; Liu, C.; Gao, F.; Su, M.; Wu, X.; Zheng, L.; Hong, F.; Yang, P. The improvement of spinach growth by nano-anatase TiO2 treatment is related to nitrogen photoreduction. Biol. Trace Elem. Res. 2007, 119, 77–88. [Google Scholar] [CrossRef] [PubMed]
- English, J.E.; Maynard, D.N. A key to nutrient disorders of vegetable plants. HortScience 1977, 13, 28–29. [Google Scholar]
- Schlesinger, W.H. Biogeochemistry: An Analysis of Global Change, 2nd ed.; Academic Press: San Diego, CA, USA, 1997; p. 588. [Google Scholar]
- Farago, M.E. Plants and The Chemical Elements: Biochemistry, Uptake, Tolerance, and Toxicity; Wiley-VCH Verlag GmbH: Weinheim, Germany, 1994. [Google Scholar]
- Harris, A.T.; Bali, R. On the formation and extent of uptake of silver nanoparticles by live plants. J. Nanopart. Res. 2008, 10, 691–695. [Google Scholar] [CrossRef]
- Brear, E.M.; Day, D.A.; Smith, P.M.C. Iron: An essential micronutrient for the legume-rhizobiumsymbiosis. Front. Plant Sci. 2013, 4, 359. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhu, S.; Martins de Godoi, D.R.; Samia, A.C.S.; Scherson, D. Adsorption of Cd2+ on carboxyl-terminated superparamagnetic iron oxide nanoparticles. Anal. Chem. 2012, 84, 3764–3770. [Google Scholar] [CrossRef] [PubMed]
- Moore, K. Determination of phosphorus in plant tissue by colorimetry. In Plant Analysis Reference Procedures for the Southern Region of the United States; Plank, C., Ed.; Georgia Agricultural Experiment Station: Athens, GA, USA, 1992. [Google Scholar]
- Kuo, S. Phosphorus. In Methods of Soil Analysis. Part 3, Chemical Methods; Sparks, D., Ed.; American Society of Agronomy-Soil Science Society of America: Madison, WI, USA, 1996; pp. 869–919. [Google Scholar]
- Burke, D.J.; Kretzer, A.M.; Rygiewicz, P.T.; Topa, M.A. Soil bacterial diversity in a loblolly pine plantation: Influence of ectomycorrhizas and fertilization. FEMS Microbiol. Ecol. 2006, 57, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Helgason, T.; Daniell, T.J.; Husband, R.; Fitter, A.H.; Young, J.P. Ploughing up the wood-wide web? Nature 1998, 394, 431. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Lalonde, M.; Bruns, T.D. Specific amplification of 18s fungal ribosomal genes from vesicular-arbuscular endomycorrhizal fungi colonizing roots. Appl. Environ. Microb. 1992, 58, 291–295. [Google Scholar]
- Hewins, C.R.; Carrino-Kyker, S.R.; Burke, D.J. Seasonal variation in mycorrhizal fungi colonizing roots of Allium tricoccum (wild leek) in a mature mixed hardwood forest. Mycorrhiza 2015, 25, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.J. Effects of Alliaria petiolata (garlic mustard; Brassicaceae) on mycorrhizal colonization and community structure in three herbaceous plants in a mixed deciduous forest. Am. J. Bot. 2008, 95, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Widmer, F.; Shaffer, B.T.; Porteous, L.A.; Seidler, R.J. Analysis of nifh gene pool complexity in soil and litter at a douglas fir forest site in the oregon cascade mountain range. Appl. Environ. Microb. 1999, 65, 374–380. [Google Scholar]
- Burke, D.J.; Chan, C.R. Effects of the invasive plant garlic mustard (Alliaria petiolata) on bacterial communities in a northern hardwood forest soil. Can. J. Microbiol. 2010, 56, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.J.; Dunham, S.M.; Kretzer, A.M. Molecular analysis of bacterial communities associated with the roots of douglas fir (Pseudotsuga menziesii) colonized by different ectomycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, L.M.; Sul, W.J.; Blackwood, C.B. Assessment of bias associated with incomplete extraction of microbial DNA from soil. Appl. Environ. Microb. 2009, 75, 5428–5433. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities Book; Mjm Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burke, D.J.; Pietrasiak, N.; Situ, S.F.; Abenojar, E.C.; Porche, M.; Kraj, P.; Lakliang, Y.; Samia, A.C.S. Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes. Int. J. Mol. Sci. 2015, 16, 23630-23650. https://doi.org/10.3390/ijms161023630
Burke DJ, Pietrasiak N, Situ SF, Abenojar EC, Porche M, Kraj P, Lakliang Y, Samia ACS. Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes. International Journal of Molecular Sciences. 2015; 16(10):23630-23650. https://doi.org/10.3390/ijms161023630
Chicago/Turabian StyleBurke, David J., Nicole Pietrasiak, Shu F. Situ, Eric C. Abenojar, Mya Porche, Pawel Kraj, Yutthana Lakliang, and Anna Cristina S. Samia. 2015. "Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes" International Journal of Molecular Sciences 16, no. 10: 23630-23650. https://doi.org/10.3390/ijms161023630
APA StyleBurke, D. J., Pietrasiak, N., Situ, S. F., Abenojar, E. C., Porche, M., Kraj, P., Lakliang, Y., & Samia, A. C. S. (2015). Iron Oxide and Titanium Dioxide Nanoparticle Effects on Plant Performance and Root Associated Microbes. International Journal of Molecular Sciences, 16(10), 23630-23650. https://doi.org/10.3390/ijms161023630