Serum Oxidant and Antioxidant Status Following an All-Out 21-km Run in Adolescent Runners Undergoing Professional Training—A One-Year Prospective Trial



Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.2. Discussion
3. Experimental Section
3.1. Subjects
3.2. Professional Long-Distance Run Training
3.3. Study Design
3.4. Procedures
3.5. Serum Measurements
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Finaud, J.; Lac, G.; Filaire, E. Oxidative stress: Relationship with exercise and training. Sports Med 2006, 36, 327–358. [Google Scholar]
- Jackson, M.J. Reactive oxygen species and redox-regulation of skeletal muscle adaptations to exercise. Philos. Trans. R. Soc. Lond. B Biol. Sci 2005, 360, 2285–2291. [Google Scholar]
- Jenkins, R.R. Exercise and oxidative stress methodology: A critique. Am. J. Clin. Nutr 2000, 72, 670S–674S. [Google Scholar]
- Scheele, C.; Nielsen, S.; Pedersen, B.K. ROS and myokines promote muscle adaptation to exercise. Trends Endocrinol. Metab 2009, 20, 95–99. [Google Scholar]
- Carlsohn, A.; Rohn, S.; Mayer, F.; Schweigert, F.J. Physical activity, antioxidant status, and protein modification in adolescent athletes. Med. Sci. Sports Exerc 2010, 42, 1131–1139. [Google Scholar]
- Goldfarb, A.H. Nutritional antioxidants as therapeutic and preventive modalities in exercise-induced muscle damage. Can. J. Appl. Physiol 1999, 24, 249–266. [Google Scholar]
- Reid, M.B.; Haack, K.E.; Franchek, K.M.; Valberg, P.A.; Kobzik, L.; West, M.S. Reactive oxygen in skeletal muscle. I. Intracellular oxidant kinetics and fatigue in vitro. J. Appl. Physiol 1992, 73, 1797–1804. [Google Scholar]
- Sen, C.K.; Kolosova, I.; Hänninen, O.; Orlov, S.N. Inward potassium transport systems in skeletal muscle derived cells are highly sensitive to oxidant exposure. Free Radic. Biol. Med 1995, 18, 795–800. [Google Scholar]
- Miles, M.V.; Horn, P.S.; Tang, P.H.; Morrison, J.A.; Miles, L.; De Grauw, T.; Pesce, A.J. Age-related changes in plasma coenzyme Q10 concentrations and redox state in apparently healthy children and adults. Clin. Chim. Acta 2004, 347, 139–144. [Google Scholar]
- Riddell, M.C. The endocrine response and substrate utilization during exercise in children and adolescents. J. Appl. Physiol 2008, 105, 725–733. [Google Scholar]
- Zanconato, S.; Buchthal, S.; Barstow, T.J.; Cooper, D.M. 31P-magnetic resonance spectroscopy of leg muscle metabolism during exercise in children and adults. J. Appl. Physiol 1993, 74, 2214–2218. [Google Scholar]
- Tong, T.K.; Lin, H.; Lippi, G.; Nie, J.; Tian, Y. Serum oxidant and antioxidant status in adolescents undergoing professional endurance sports training. Oxid. Med. Cell Longev. 2012, 2012. [Google Scholar] [CrossRef]
- Tian, Y.; Nie, J.; Tong, T.K.; Baker, J.S.; Thomas, N.E.; Shi, Q. Serum oxidant and antioxidant status during early and late recovery periods following an all-out 21-km run in trained adolescent runners. Eur. J. Appl. Physiol 2010, 110, 971–976. [Google Scholar]
- Kim, S.Y.; Noguera, J.C.; Morales, J.; Velando, A. Heritability of resistance to oxidative stress in early life. J. Evol. Biol 2010, 23, 769–775. [Google Scholar]
- Miyagi, S.J.; Brown, I.W.; Chock, J.M.; Collier, A.C. Developmental changes in hepatic antioxidant capacity are age-and sex-dependent. J. Pharmacol. Sci 2009, 111, 440–445. [Google Scholar]
- Metcalfe, N.B.; Monaghan, P. Compensation for a bad start: Grow now, pay later? Trends Ecol. Evol 2001, 16, 254–260. [Google Scholar]
- Goldfarb, A.H.; McKenzie, M.J.; Bloomer, R.J. Gender comparisons of exercise-induced oxidative stress: influence of antioxidant supplementation. Appl. Physiol. Nutr. Metab 2007, 32, 1124–1131. [Google Scholar]
- Gougoura, S.; Nikolaidis, M.G.; Kostaropoulos, I.A.; Jamurtas, A.Z.; Koukoulis, G.; Kouretas, D. Increased oxidative stress indices in the blood of child swimmers. Eur. J. Appl. Physiol 2007, 100, 235–239. [Google Scholar]
- Tessier, F.; Margaritis, I.; Richard, M.J.; Moynot, C.; Marconnet, P. Selenium and training effects on the glutathione system and aerobic performance. Med. Sci. Sports Exerc 1995, 27, 390–396. [Google Scholar]
- Chang, C.K.; Tseng, H.F.; Hsuuw, Y.D.; Chan, W.H.; Shieh, L.C. Higher LDL oxidation at rest and after a rugby game in weekend warriors. Ann. Nutr. Metab 2002, 46, 103–107. [Google Scholar]
- Schippinger, G.; Wonisch, W.; Abuja, P.M.; Fankhauser, F.; Winklhofer-Roob, B.M.; Halwachs, G. Lipid peroxidation and antioxidant status in professional American football players during competition. Eur. J. Clin. Invest 2002, 32, 686–692. [Google Scholar]
- Kohen, R.; Vellaichamy, E.; Hrbac, J.; Gati, I.; Tirosh, O. Quantification of the overall reactive oxygen species scavenging capacity of biological fluids and tissues. Free Radic. Biol. Med 2000, 28, 871–879. [Google Scholar]
- Sen, C.K. Oxidants and antioxidants in exercise. J. Appl. Physiol 1995, 79, 675–686. [Google Scholar]
- Vertuani, S.; Angusti, A.; Manfredini, S. The antioxidants and pro-antioxidants network: An overview. Curr. Pharm. Des 2004, 10, 1677–1694. [Google Scholar]
- Hellsten, Y. Xanthine dehydrogenase and purine metabolism in man. With special reference to exercise. Acta Physiol. Scand. Suppl 1994, 621, 1–73. [Google Scholar]
- Groussard, C.; Rannou-Bekono, F.; Machefer, G.; Chevanne, M.; Vincent, S.; Sergent, O.; Cillard, J.; Gratas-Delamarche, A. Changes in blood lipid peroxidation markers and antioxidants after a single sprint anaerobic exercise. Eur. J. Appl. Physiol 2003, 89, 14–20. [Google Scholar]
- Blum, J.; Fridovich, I. Inactivation of glutathione peroxidase by superoxide radical. Arch. Biochem. Biophys 1985, 240, 500–508. [Google Scholar]
- Concepcion-Huertas, M.; Chirosa, L.J.; De Haro, T.; Chirosa, I.J.; Romero, V.; Aguilar-Martinez, D.; Leonardo-Mendonça, R.C.; Doerrier, C.; Escames, G.; Acuña-Castroviejo, D. Changes in the redox status and inflammatory response in handball players during one-year of competition and training. J. Sports Sci 2013, 31, 1197–1207. [Google Scholar]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kouretas, D. Blood reflects tissue oxidative stress depending on biomarker and tissue studied. Free Radic. Biol. Med 2009, 47, 1371–1374. [Google Scholar]
- Nikolaidis, M.G.; Jamurtas, A.Z.; Paschalis, V.; Fatouros, I.G.; Koutedakis, Y.; Kouretas, D. The effect of muscle-damaging exercise on blood and skeletal muscle oxidative stress: Magnitude and time-course considerations. Sports Med 2008, 38, 579–606. [Google Scholar]
- Fischer, G.; Schwartz, D.D.; Quindry, J.; Barberio, M.D.; Foster, E.B.; Jones, K.W.; Pascoe, D.D. Lymphocyte enzymatic antioxidant responses to oxidative stress following high-intensity interval exercise. J. Appl. Physiol 2011, 110, 730–737. [Google Scholar]
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolau, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K.; et al. Oxidative stress biomarkers responses to physical overtraining: implications for diagnosis. Free Radic. Biol. Med 2007, 43, 901–910. [Google Scholar]
- Chung, S.C.; Goldfarb, A.H.; Jamurtas, A.Z.; Hegde, S.S.; Lee, J. Effect of exercise during the follicular and luteal phases on indices of oxidative stress in healthy women. Med. Sci. Sports Exerc 1999, 31, 409–413. [Google Scholar]
- Benzie, I.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzymol 1999, 299, 15–27. [Google Scholar]
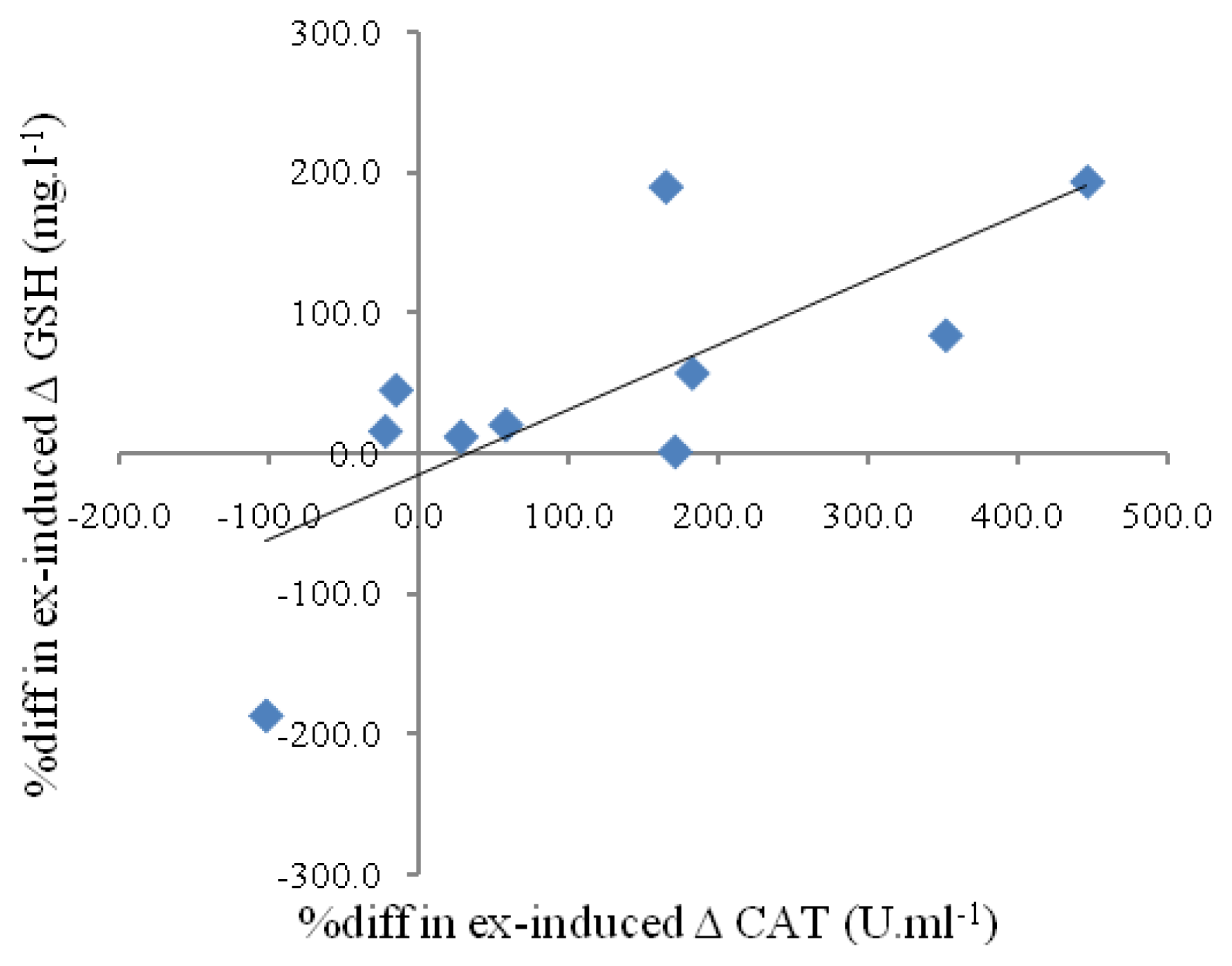

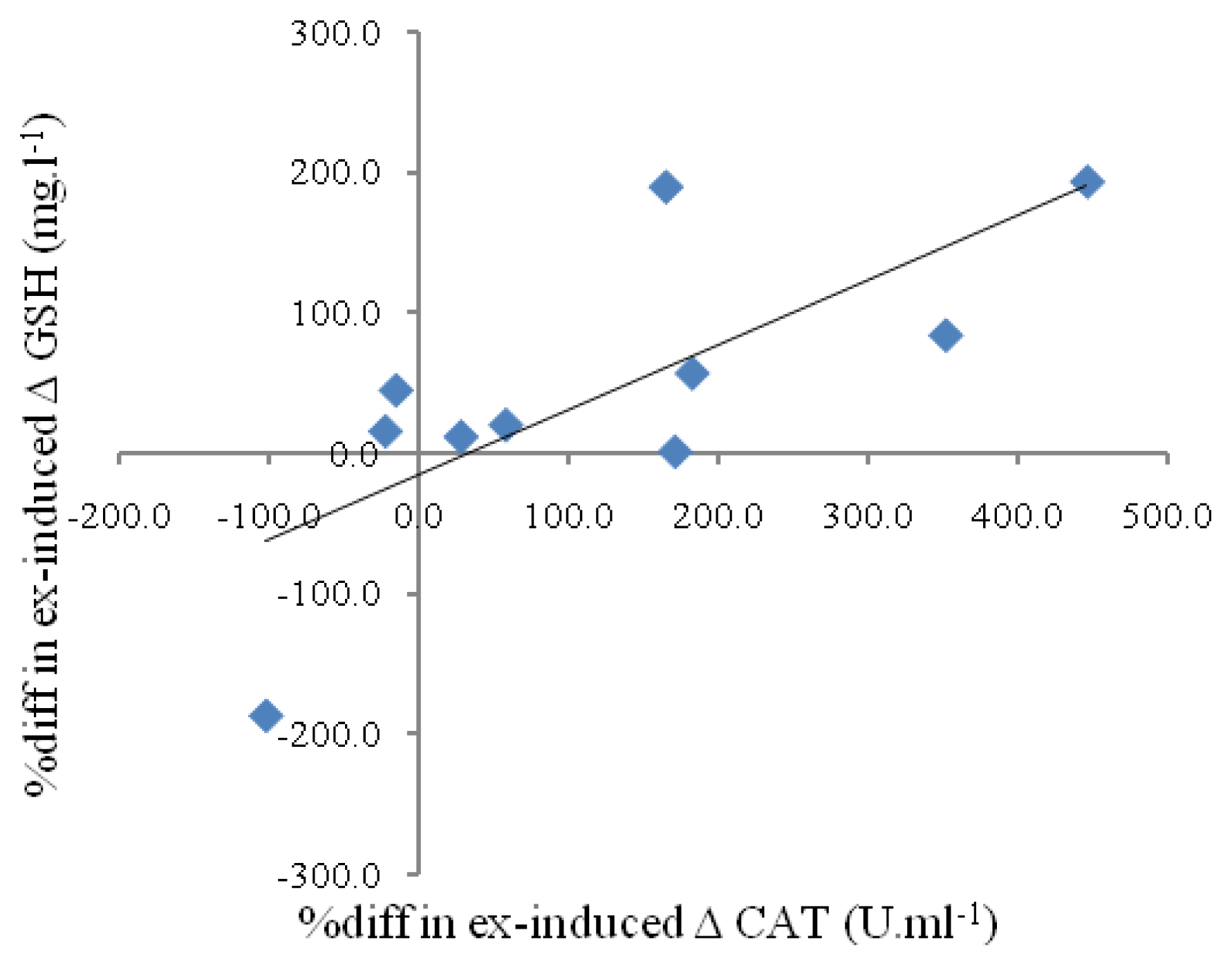{kind=link}
| Pre-evaluation | Post-evaluation | ||
|---|---|---|---|
| TBARS (nmol·mL−1) | Pre-ex | 5.64 ± 0.92 | 5.61 ± 0.91 |
| Post-ex | 4.24 ± 0.85 * | 5.40 ± 1.27 * | |
| XO (U·L−1) | Pre-ex | 14.7 ± 2.9 | 16.4 ± 4.5 |
| Post-ex | 13.4 ± 3.9 | 24.4 ± 7.8 *,a | |
| GSH (mg·L−1) | Pre-ex | 20.9 ± 10.3 | 21.1 ± 8.9 |
| Post-ex | 10.1 ± 4.7 | 21.1 ± 12.7 | |
| CAT (U·mL−1) | Pre-ex | 4.48 ± 1.51 | 3.84 ± 1.63 |
| Post-ex | 3.12 ± 4.19 | 7.79 ± 2.79 *,a | |
| T-AOC (U·mL−1) | Pre-ex | 11.2 ± 1.92 | 13.3 ± 1.0 a |
| Post-ex | 12.3 ± 1.33 | 13.7 ± 1.59 a | |
| SOD (U·mL−1) | Pre-ex | 69.2 ± 12.3 | 67.0 ± 13.5 |
| Post-ex | 63.4 ± 15.6 * | 60.9 ± 13.9 * | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tong, T.K.; Kong, Z.; Lin, H.; Lippi, G.; Zhang, H.; Nie, J. Serum Oxidant and Antioxidant Status Following an All-Out 21-km Run in Adolescent Runners Undergoing Professional Training—A One-Year Prospective Trial. Int. J. Mol. Sci. 2013, 14, 15167-15178. https://doi.org/10.3390/ijms140715167
Tong TK, Kong Z, Lin H, Lippi G, Zhang H, Nie J. Serum Oxidant and Antioxidant Status Following an All-Out 21-km Run in Adolescent Runners Undergoing Professional Training—A One-Year Prospective Trial. International Journal of Molecular Sciences. 2013; 14(7):15167-15178. https://doi.org/10.3390/ijms140715167
Chicago/Turabian StyleTong, Tom K., Zhaowei Kong, Hua Lin, Giuseppe Lippi, Haifeng Zhang, and Jinlei Nie. 2013. "Serum Oxidant and Antioxidant Status Following an All-Out 21-km Run in Adolescent Runners Undergoing Professional Training—A One-Year Prospective Trial" International Journal of Molecular Sciences 14, no. 7: 15167-15178. https://doi.org/10.3390/ijms140715167
APA StyleTong, T. K., Kong, Z., Lin, H., Lippi, G., Zhang, H., & Nie, J. (2013). Serum Oxidant and Antioxidant Status Following an All-Out 21-km Run in Adolescent Runners Undergoing Professional Training—A One-Year Prospective Trial. International Journal of Molecular Sciences, 14(7), 15167-15178. https://doi.org/10.3390/ijms140715167