Claudins Overexpression in Ovarian Cancer: Potential Targets for Clostridium Perfringens Enterotoxin (CPE) Based Diagnosis and Therapy

Abstract
:1. Introduction
2. Structure and Function of Claudins
3. Claudins in Ovarian Cancer
3.1. Claudin-1 and Claudin-2
3.2. Claudin-3 and Claudin-4
3.3. Claudin-5
3.4. Claudin-6
3.5. Claudin-7
4. Claudin-3 and -4 are Potential Targets for CPE-Based Theranostics
5. Conclusions
Conflict of Interest
References
- Soini, Y.; Talvensaari-Mattila, A. Expression of claudins 1, 4, 5, and 7 in ovarian tumors of diverse types. Int. J. Gynecol. Pathol 2006, 25, 330–335. [Google Scholar]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol 2001, 2, 285–293. [Google Scholar]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J. Cell Biol 1998, 141, 1539–1550. [Google Scholar]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junctions. J. Cell Biol 1993, 123, 1777–1788. [Google Scholar]
- Swift, J.G.; Mukherjee, T.M.; Rowland, R. Intercellular junctions in hepatocellular carcinoma. J. Submicrosc. Cytol 1983, 15, 799–810. [Google Scholar]
- Furuse, M.; Sasaki, H.; Fujimoto, K.; Tsukita, S. A single gene product, claudin-1 or -2, reconstitutes tight junction strands and recruits occludin in fibroblasts. J. Cell Biol 1998, 143, 391–401. [Google Scholar]
- Tsukita, S.; Furuse, M. Pores in the wall: Claudins constitute tight junction strands containing aqueous pores. J. Cell Biol 2000, 149, 13–16. [Google Scholar]
- Martin, T.A.; Watkins, G.; Mansel, R.E.; Jiang, W.G. Loss of tight junction plaque molecules in breast cancer tissues is associated with a poor prognosis in patients with breast cancer. Eur. J. Cancer 2004, 40, 2717–2725. [Google Scholar]
- Mitic, L.L.; Anderson, J.M. Molecular architecture of tight junctions. Annu. Rev. Physiol 1998, 60, 121–142. [Google Scholar]
- Morin, P.J. Claudin proteins in human cancer: Promising new targets for diagnosis and therapy. Cancer Res 2005, 65, 9603–9606. [Google Scholar]
- Turksen, K.; Troy, T.C. Barriers built on claudins. J. Cell Sci 2004, 117, 2435–2447. [Google Scholar]
- Chen, Y.; Lu, Q.; Schneeberger, E.E.; Goodenough, D.A. Restoration of tight junction structure and barrier function by down-regulation of the mitogen-activated protein kinase pathway in ras-transformed madin-darby canine kidney cells. Mol. Biol. Cell 2000, 11, 849–862. [Google Scholar]
- Singh, A.B.; Sharma, A.; Dhawan, P. Claudin family of proteins and cancer: An overview. J. Oncol 2010, 2010, 541957. [Google Scholar]
- Rangel, L.B.; Agarwal, R.; D’Souza, T.; Pizer, E.S.; Alo, P.L.; Lancaster, W.D.; Gregoire, L.; Schwartz, D.R.; Cho, K.R.; Morin, P.J. Tight junction proteins claudin-3 and claudin-4 are frequently overexpressed in ovarian cancer but not in ovarian cystadenomas. Clin. Cancer Res 2003, 9, 2567–2575. [Google Scholar]
- Bignotti, E.; Tassi, R.A.; Calza, S.; Ravaggi, A.; Romani, C.; Rossi, E.; Falchetti, M.; Odicino, F.E.; Pecorelli, S.; Santin, A.D. Differential gene expression profiles between tumor biopsies and short-term primary cultures of ovarian serous carcinomas: Identification of novel molecular biomarkers for early diagnosis and therapy. Gynecol. Oncol 2006, 103, 405–416. [Google Scholar]
- Hough, C.D.; Sherman-Baust, C.A.; Pizer, E.S.; Montz, F.J.; Im, D.D.; Rosenshein, N.B.; Cho, K.R.; Riggins, G.J.; Morin, P.J. Large-scale serial analysis of gene expression reveals genes differentially expressed in ovarian cancer. Cancer Res 2000, 60, 6281–6287. [Google Scholar]
- Rangel, L.B.; Sherman-Baust, C.A.; Wernyj, R.P.; Schwartz, D.R.; Cho, K.R.; Morin, P.J. Characterization of novel human ovarian cancer-specific transcripts (hosts) identified by serial analysis of gene expression. Oncogene 2003, 22, 7225–7232. [Google Scholar]
- Santin, A.D.; Zhan, F.; Bellone, S.; Palmieri, M.; Cane, S.; Bignotti, E.; Anfossi, S.; Gokden, M.; Dunn, D.; Roman, J.J.; et al. Gene expression profiles in primary ovarian serous papillary tumors and normal ovarian epithelium: Identification of candidate molecular markers for ovarian cancer diagnosis and therapy. Int. J. Cancer 2004, 112, 14–25. [Google Scholar]
- Ismail, R.S.; Baldwin, R.L.; Fang, J.; Browning, D.; Karlan, B.Y.; Gasson, J.C.; Chang, D.D. Differential gene expression between normal and tumor-derived ovarian epithelial cells. Cancer Res 2000, 60, 6744–6749. [Google Scholar]
- Welsh, J.B.; Zarrinkar, P.P.; Sapinoso, L.M.; Kern, S.G.; Behling, C.A.; Monk, B.J.; Lockhart, D.J.; Burger, R.A.; Hampton, G.M. Analysis of gene expression profiles in normal and neoplastic ovarian tissue samples identifies candidate molecular markers of epithelial ovarian cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 1176–1181. [Google Scholar]
- Schwartz, D.R.; Kardia, S.L.; Shedden, K.A.; Kuick, R.; Michailidis, G.; Taylor, J.M.; Misek, D.E.; Wu, R.; Zhai, Y.; Darrah, D.M.; et al. Gene expression in ovarian cancer reflects both morphology and biological behavior, distinguishing clear cell from other poor-prognosis ovarian carcinomas. Cancer Res 2002, 62, 4722–4729. [Google Scholar]
- Ono, K.; Tanaka, T.; Tsunoda, T.; Kitahara, O.; Kihara, C.; Okamoto, A.; Ochiai, K.; Takagi, T.; Nakamura, Y. Identification by cdna microarray of genes involved in ovarian carcinogenesis. Cancer Res 2000, 60, 5007–5011. [Google Scholar]
- Shridhar, V.; Lee, J.; Pandita, A.; Iturria, S.; Avula, R.; Staub, J.; Morrissey, M.; Calhoun, E.; Sen, A.; Kalli, K.; et al. Genetic analysis of early- versus late-stage ovarian tumors. Cancer Res 2001, 61, 5895–5904. [Google Scholar]
- Hough, C.D.; Cho, K.R.; Zonderman, A.B.; Schwartz, D.R.; Morin, P.J. Coordinately up-regulated genes in ovarian cancer. Cancer Res 2001, 61, 3869–3876. [Google Scholar]
- Shridhar, V.; Sen, A.; Chien, J.; Staub, J.; Avula, R.; Kovats, S.; Lee, J.; Lillie, J.; Smith, D.I. Identification of underexpressed genes in early- and late-stage primary ovarian tumors by suppression subtraction hybridization. Cancer Res 2002, 62, 262–270. [Google Scholar]
- Jazaeri, A.A.; Lu, K.; Schmandt, R.; Harris, C.P.; Rao, P.H.; Sotiriou, C.; Chandramouli, G.V.; Gershenson, D.M.; Liu, E.T. Molecular determinants of tumor differentiation in papillary serous ovarian carcinoma. Mol. Carcinog 2003, 36, 53–59. [Google Scholar]
- Soini, Y. Expression of claudins 1, 2, 3, 4, 5 and 7 in various types of tumours. Histopathology 2005, 46, 551–560. [Google Scholar]
- Kleinberg, L.; Holth, A.; Trope, C.G.; Reich, R.; Davidson, B. Claudin upregulation in ovarian carcinoma effusions is associated with poor survival. Human Pathol 2008, 39, 747–757. [Google Scholar]
- Resnick, M.B.; Konkin, T.; Routhier, J.; Sabo, E.; Pricolo, V.E. Claudin-1 is a strong prognostic indicator in stage ii colonic cancer: A tissue microarray study. Mod. Pathol 2005, 18, 511–518. [Google Scholar]
- Sanada, Y.; Oue, N.; Mitani, Y.; Yoshida, K.; Nakayama, H.; Yasui, W. Down-regulation of the claudin-18 gene, identified through serial analysis of gene expression data analysis, in gastric cancer with an intestinal phenotype. J. Pathol 2006, 208, 633–642. [Google Scholar]
- Cheung, S.T.; Leung, K.L.; Ip, Y.C.; Chen, X.; Fong, D.Y.; Ng, I.O.; Fan, S.T.; So, S. Claudin-10 expression level is associated with recurrence of primary hepatocellular carcinoma. Clin. Cancer Res 2005, 11, 551–556. [Google Scholar]
- Gonzalez-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B.E. Tight junction proteins. Prog. Biophys. Mol. Biol 2003, 81, 1–44. [Google Scholar]
- Van Itallie, C.M.; Anderson, J.M. Claudins and epithelial paracellular transport. Annu. Rev. Physiol 2006, 68, 403–429. [Google Scholar]
- Lal-Nag, M.; Morin, P.J. The claudins. Genome Biol 2009, 10, 235. [Google Scholar]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar]
- Ikenouchi, J.; Furuse, M.; Furuse, K.; Sasaki, H.; Tsukita, S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J. Cell Biol 2005, 171, 939–945. [Google Scholar]
- Steed, E.; Rodrigues, N.T.; Balda, M.S.; Matter, K. Identification of marveld3 as a tight junction-associated transmembrane protein of the occludin family. BMC Cell Biol 2009, 10, 95. [Google Scholar]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and function of claudins. Biochim. Biophys. Acta 2008, 1778, 631–645. [Google Scholar]
- Umeda, K.; Ikenouchi, J.; Katahira-Tayama, S.; Furuse, K.; Sasaki, H.; Nakayama, M.; Matsui, T.; Tsukita, S.; Furuse, M. Zo-1 and zo-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell 2006, 126, 741–754. [Google Scholar]
- D’Souza, T.; Agarwal, R.; Morin, P.J. Phosphorylation of claudin-3 at threonine 192 by camp-dependent protein kinase regulates tight junction barrier function in ovarian cancer cells. J. Biol. Chem 2005, 280, 26233–26240. [Google Scholar]
- Hewitt, K.J.; Agarwal, R.; Morin, P.J. The claudin gene family: Expression in normal and neoplastic tissues. BMC Cancer 2006, 6, 186. [Google Scholar]
- Shang, X.; Lin, X.; Alvarez, E.; Manorek, G.; Howell, S.B. Tight junction proteins claudin-3 and claudin-4 control tumor growth and metastases. Neoplasia 2012, 14, 974–985. [Google Scholar]
- Hibbs, K.; Skubitz, K.M.; Pambuccian, S.E.; Casey, R.C.; Burleson, K.M.; Oegema, T.R., Jr; Thiele, J.J.; Grindle, S.M.; Bliss, R.L.; Skubitz, A.P. Differential gene expression in ovarian carcinoma: Identification of potential biomarkers. Am. J. Pathol 2004, 165, 397–414. [Google Scholar]
- Daugherty, B.L.; Ward, C.; Smith, T.; Ritzenthaler, J.D.; Koval, M. Regulation of heterotypic claudin compatibility. J. Biol. Chem 2007, 282, 30005–30013. [Google Scholar]
- Piontek, J.; Winkler, L.; Wolburg, H.; Muller, S.L.; Zuleger, N.; Piehl, C.; Wiesner, B.; Krause, G.; Blasig, I.E. Formation of tight junction: Determinants of homophilic interaction between classic claudins. FASEB J 2008, 22, 146–158. [Google Scholar]
- Piehl, C.; Piontek, J.; Cording, J.; Wolburg, H.; Blasig, I.E. Participation of the second extracellular loop of claudin-5 in paracellular tightening against ions, small and large molecules. Cell Mol. Life Sci 2010, 67, 2131–2140. [Google Scholar]
- Huber, J.D.; Witt, K.A.; Hom, S.; Egleton, R.D.; Mark, K.S.; Davis, T.P. Inflammatory pain alters blood-brain barrier permeability and tight junctional protein expression. Am. J. Physiol. Heart Circ. Physiol 2001, 280, H1241–H1248. [Google Scholar]
- Sawada, N.; Murata, M.; Kikuchi, K.; Osanai, M.; Tobioka, H.; Kojima, T.; Chiba, H. Tight junctions and human diseases. Med. Electron. Microsc 2003, 36, 147–156. [Google Scholar]
- Wolburg, H.; Wolburg-Buchholz, K.; Liebner, S.; Engelhardt, B. Claudin-1, claudin-2 and claudin-11 are present in tight junctions of choroid plexus epithelium of the mouse. Neurosci. Lett 2001, 307, 77–80. [Google Scholar]
- Bronstein, J.M.; Tiwari-Woodruff, S.; Buznikov, A.G.; Stevens, D.B. Involvement of osp/claudin-11 in oligodendrocyte membrane interactions: Role in biology and disease. J. Neurosci. Res 2000, 59, 706–711. [Google Scholar]
- Wilcox, E.R.; Burton, Q.L.; Naz, S.; Riazuddin, S.; Smith, T.N.; Ploplis, B.; Belyantseva, I.; Ben-Yosef, T.; Liburd, N.A.; Morell, R.J.; et al. Mutations in the gene encoding tight junction claudin-14 cause autosomal recessive deafness dfnb29. Cell 2001, 104, 165–172. [Google Scholar]
- Gye, M.C. Changes in the expression of claudins and transepithelial electrical resistance of mouse sertoli cells by leydig cell coculture. Int. J. Androl 2003, 26, 271–278. [Google Scholar]
- Swisshelm, K.; Macek, R.; Kubbies, M. Role of claudins in tumorigenesis. Adv. Drug Deliv. Rev 2005, 57, 919–928. [Google Scholar]
- Lu, K.H.; Patterson, A.P.; Wang, L.; Marquez, R.T.; Atkinson, E.N.; Baggerly, K.A.; Ramoth, L.R.; Rosen, D.G.; Liu, J.; Hellstrom, I.; et al. Selection of potential markers for epithelial ovarian cancer with gene expression arrays and recursive descent partition analysis. Clin. Cancer Res 2004, 10, 3291–3300. [Google Scholar]
- Zhu, Y.; Brannstrom, M.; Janson, P.O.; Sundfeldt, K. Differences in expression patterns of the tight junction proteins,claudin 1, 3, 4 and 5, in human ovarian surface epithelium as compared to epithelia in inclusion cysts and epithelial ovarian tumours. Int. J. Cancer 2006, 118, 1884–1891. [Google Scholar]
- Heinzelmann-Schwarz, V.A.; Gardiner-Garden, M.; Henshall, S.M.; Scurry, J.; Scolyer, R.A.; Davies, M.J.; Heinzelmann, M.; Kalish, L.H.; Bali, A.; Kench, J.G.; et al. Overexpression of the cell adhesion molecules ddr1, claudin 3, and ep-cam in metaplastic ovarian epithelium and ovarian cancer. Clin. Cancer Res 2004, 10, 4427–4436. [Google Scholar]
- Kominsky, S.L.; Argani, P.; Korz, D.; Evron, E.; Raman, V.; Garrett, E.; Rein, A.; Sauter, G.; Kallioniemi, O.P.; Sukumar, S. Loss of the tight junction protein claudin-7 correlates with histological grade in both ductal carcinoma in situ and invasive ductal carcinoma of the breast. Oncogene 2003, 22, 2021–2033. [Google Scholar]
- Johnson, A.H.; Frierson, H.F.; Zaika, A.; Powell, S.M.; Roche, J.; Crowe, S.; Moskaluk, C.A.; El-Rifai, W. Expression of tight-junction protein claudin-7 is an early event in gastric tumorigenesis. Am. J. Pathol 2005, 167, 577–584. [Google Scholar]
- Agarwal, R.; D’Souza, T.; Morin, P.J. Claudin-3 and claudin-4 expression in ovarian epithelial cells enhances invasion and is associated with increased matrix metalloproteinase-2 activity. Cancer Res 2005, 65, 7378–7385. [Google Scholar]
- Higashi, Y.; Suzuki, S.; Sakaguchi, T.; Nakamura, T.; Baba, S.; Reinecker, H.C.; Nakamura, S.; Konno, H. Loss of claudin-1 expression correlates with malignancy of hepatocellular carcinoma. J. Surg. Res 2007, 139, 68–76. [Google Scholar]
- Hadj-Rabia, S.; Baala, L.; Vabres, P.; Hamel-Teillac, D.; Jacquemin, E.; Fabre, M.; Lyonnet, S.; De Prost, Y.; Munnich, A.; Hadchouel, M.; et al. Claudin-1 gene mutations in neonatal sclerosing cholangitis associated with ichthyosis: A tight junction disease. Gastroenterology 2004, 127, 1386–1390. [Google Scholar]
- Lameris, A.L.; Huybers, S.; Burke, J.R.; Monnens, L.A.; Bindels, R.J.; Hoenderop, J.G. Involvement of claudin 3 and claudin 4 in idiopathic infantile hypercalcaemia: A novel hypothesis? Nephrol. Dial. Transplant 2010, 25, 3504–3509. [Google Scholar]
- Morita, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Endothelial claudin: Claudin-5/tmvcf constitutes tight junction strands in endothelial cells. J. Cell Biol 1999, 147, 185–194. [Google Scholar]
- Gonzalez-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of tight junction components with signaling pathways. Biochim. Biophys. Acta 2008, 1778, 729–756. [Google Scholar]
- Matsuda, M.; Kubo, A.; Furuse, M.; Tsukita, S. A peculiar internalization of claudins, tight junction-specific adhesion molecules, during the intercellular movement of epithelial cells. J. Cell Sci 2004, 117, 1247–1257. [Google Scholar]
- Ikenouchi, J.; Matsuda, M.; Furuse, M.; Tsukita, S. Regulation of tight junctions during the epithelium-mesenchyme transition: Direct repression of the gene expression of claudins/occludin by snail. J. Cell Sci 2003, 116, 1959–1967. [Google Scholar]
- Escaffit, F.; Boudreau, F.; Beaulieu, J.F. Differential expression of claudin-2 along the human intestine: Implication of gata-4 in the maintenance of claudin-2 in differentiating cells. J. Cell. Physiol 2005, 203, 15–26. [Google Scholar]
- Singh, A.B.; Harris, R.C. Epidermal growth factor receptor activation differentially regulates claudin expression and enhances transepithelial resistance in madin-darby canine kidney cells. J. Biol. Chem 2004, 279, 3543–3552. [Google Scholar]
- Grande, M.; Franzen, A.; Karlsson, J.O.; Ericson, L.E.; Heldin, N.E.; Nilsson, M. Transforming growth factor-beta and epidermal growth factor synergistically stimulate epithelial to mesenchymal transition (emt) through a mek-dependent mechanism in primary cultured pig thyrocytes. J. Cell Sci 2002, 115, 4227–4236. [Google Scholar]
- Flores-Benitez, D.; Ruiz-Cabrera, A.; Flores-Maldonado, C.; Shoshani, L.; Cereijido, M.; Contreras, R.G. Control of tight junctional sealing: Role of epidermal growth factor. Am. J. Physiol. Renal. Physiol 2007, 292, F828–F836. [Google Scholar]
- Ogawa, M.; Kojima, T.; Someya, M.; Nomura, K.; Takasawa, A.; Murata, M.; Tanaka, S.; Saito, T.; Sawada, N. Epidermal growth factor modulates claudins and tight junctional functions in ovarian cancer cell lines. Histochem. Cell Biol 2012, 138, 323–338. [Google Scholar]
- Zhu, Y.; Sundfeldt, K. Tight junction formation in epithelial ovarian adenocarcinoma. Acta Obstet. Gynecol. Scand 2007, 86, 1011–1019. [Google Scholar]
- Qin, W.; Ren, Q.; Liu, T.; Huang, Y.; Wang, J. Microrna-155 is a novel suppressor of ovarian cancer-initiating cells that targets cldn1. FEBS Lett 2013, 587, 1434–1439. [Google Scholar]
- laSuh, Y.; Yoon, C.H.; Kim, R.K.; Lim, E.J.; Oh, Y.S.; Hwang, S.G.; An, S.; Yoon, G.; Gye, M.C.; Yi, J.M.; et al. Claudin-1 induces epithelial-mesenchymal transition through activation of the c-abl-erk signaling pathway in human liver cells. Oncogene 2012. [Google Scholar] [CrossRef]
- Liu, T.; Cheng, W.; Lai, D.; Huang, Y.; Guo, L. Characterization of primary ovarian cancer cells in different culture systems. Oncol. Rep 2010, 23, 1277–1284. [Google Scholar]
- Soini, Y. Claudins 2, 3, 4, and 5 in paget’s disease and breast carcinoma. Human Pathol 2004, 35, 1531–1536. [Google Scholar]
- Patonai, A.; Erdelyi-Belle, B.; Korompay, A.; Somoracz, A.; Straub, B.K.; Schirmacher, P.; Kovalszky, I.; Lotz, G.; Kiss, A.; Schaff, Z. Claudins and tricellulin in fibrolamellar hepatocellular carcinoma. Virchows Archiv 2011, 458, 679–688. [Google Scholar]
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M.J. Cancer statistics, 2009. CA Cancer J. Clin 2009, 59, 225–249. [Google Scholar]
- Bristow, R.E.; Tomacruz, R.S.; Armstrong, D.K.; Trimble, E.L.; Montz, F.J. Survival effect of maximal cytoreductive surgery for advanced ovarian carcinoma during the platinum era: A meta-analysis. J. Clin. Oncol 2002, 20, 1248–1259. [Google Scholar]
- Monk, B.J.; Coleman, R.L. Changing the paradigm in the treatment of platinum-sensitive recurrent ovarian cancer: From platinum doublets to nonplatinum doublets and adding antiangiogenesis compounds. Int. J. Gynecol. Cancer 2009, 19, S63–S67. [Google Scholar]
- Boylan, K.L.; Misemer, B.; Derycke, M.S.; Andersen, J.D.; Harrington, K.M.; Kalloger, S.E.; Gilks, C.B.; Pambuccian, S.E.; Skubitz, A.P. Claudin 4 is differentially expressed between ovarian cancer subtypes and plays a role in spheroid formation. Int. J. Mol. Sci 2011, 12, 1334–1358. [Google Scholar]
- Li, J.; Chigurupati, S.; Agarwal, R.; Mughal, M.R.; Mattson, M.P.; Becker, K.G.; Wood, W.H., 3rd; Zhang, Y.; Morin, P.J. Possible angiogenic roles for claudin-4 in ovarian cancer. Cancer Biol. Ther 2009, 8, 1806–1814. [Google Scholar]
- Bellone, S.; Betti, M.; English, D.; Cocco, E.; Gasparrini, S.; Bortolomai, I.; Todeschini, P.; Romani, C.; Ravaggi, A.; Bignotti, E.; et al. Mammaglobin b (scgb2a1) is a novel tumor rejection antigen and highly differentially expressed gene in all major histological types of ovarian cancer. Br. J. Cancer 2013. submitted for publication. [Google Scholar]
- Banz, C.; Ungethuem, U.; Kuban, R.J.; Diedrich, K.; Lengyel, E.; Hornung, D. The molecular signature of endometriosis-associated endometrioid ovarian cancer differs significantly from endometriosis-independent endometrioid ovarian cancer. Fertil. Steril 2010, 94, 1212–1217. [Google Scholar]
- Tassi, R.A.; Bignotti, E.; Falchetti, M.; Ravanini, M.; Calza, S.; Ravaggi, A.; Bandiera, E.; Facchetti, F.; Pecorelli, S.; Santin, A.D. Claudin-7 expression in human epithelial ovarian cancer. Int. J. Gynecol. Cancer 2008, 18, 1262–1271. [Google Scholar]
- Dahiya, N.; Becker, K.G.; Wood, W.H., 3rd; Zhang, Y.; Morin, P.J. Claudin-7 is frequently overexpressed in ovarian cancer and promotes invasion. PLoS One 2011, 6, e22119. [Google Scholar]
- Szabo, I.; Kiss, A.; Schaff, Z.; Sobel, G. Claudins as diagnostic and prognostic markers in gynecological cancer. Histol. Histopathol 2009, 24, 1607–1615. [Google Scholar]
- Pan, X.Y.; Wang, B.; Che, Y.C.; Weng, Z.P.; Dai, H.Y.; Peng, W. Expression of claudin-3 and claudin-4 in normal, hyperplastic, and malignant endometrial tissue. Int. J. Gynecol. Cancer 2007, 17, 233–241. [Google Scholar]
- Santin, A.D.; Bellone, S.; Siegel, E.R.; McKenney, J.K.; Thomas, M.; Roman, J.J.; Burnett, A.; Tognon, G.; Bandiera, E.; Pecorelli, S. Overexpression of clostridium perfringens enterotoxin receptors claudin-3 and claudin-4 in uterine carcinosarcomas. Clin. Cancer Res 2007, 13, 3339–3346. [Google Scholar]
- Lee, J.W.; Lee, S.J.; Seo, J.; Song, S.Y.; Ahn, G.; Park, C.S.; Lee, J.H.; Kim, B.G.; Bae, D.S. Increased expressions of claudin-1 and claudin-7 during the progression of cervical neoplasia. Gynecol. Oncol 2005, 97, 53–59. [Google Scholar]
- Sobel, G.; Paska, C.; Szabo, I.; Kiss, A.; Kadar, A.; Schaff, Z. Increased expression of claudins in cervical squamous intraepithelial neoplasia and invasive carcinoma. Human Pathol 2005, 36, 162–169. [Google Scholar]
- Sobel, G.; Szabo, I.; Paska, C.; Kiss, A.; Kovalszky, I.; Kadar, A.; Paulin, F.; Schaff, Z. Changes of cell adhesion and extracellular matrix (ecm) components in cervical intraepithelial neoplasia. Pathol. Oncol. Res 2005, 11, 26–31. [Google Scholar]
- Honda, H.; Pazin, M.J.; Ji, H.; Wernyj, R.P.; Morin, P.J. Crucial roles of sp1 and epigenetic modifications in the regulation of the cldn4 promoter in ovarian cancer cells. J. Biol. Chem 2006, 281, 21433–21444. [Google Scholar]
- Kwon, M.J.; Kim, S.S.; Choi, Y.L.; Jung, H.S.; Balch, C.; Kim, S.H.; Song, Y.S.; Marquez, V.E.; Nephew, K.P.; Shin, Y.K. Derepression of cldn3 and cldn4 during ovarian tumorigenesis is associated with loss of repressive histone modifications. Carcinogenesis 2010, 31, 974–983. [Google Scholar]
- Casagrande, F.; Cocco, E.; Bellone, S.; Richter, C.E.; Bellone, M.; Todeschini, P.; Siegel, E.; Varughese, J.; Arin-Silasi, D.; Azodi, M.; et al. Eradication of chemotherapy-resistant cd44+ human ovarian cancer stem cells in mice by intraperitoneal administration of clostridium perfringens enterotoxin. Cancer 2011, 117, 5519–5528. [Google Scholar]
- Stewart, J.J.; White, J.T.; Yan, X.; Collins, S.; Drescher, C.W.; Urban, N.D.; Hood, L.; Lin, B. Proteins associated with cisplatin resistance in ovarian cancer cells identified by quantitative proteomic technology and integrated with mrna expression levels. Mol. Cell Proteomics 2006, 5, 433–443. [Google Scholar]
- Santin, A.D.; Cane, S.; Bellone, S.; Palmieri, M.; Siegel, E.R.; Thomas, M.; Roman, J.J.; Burnett, A.; Cannon, M.J.; Pecorelli, S. Treatment of chemotherapy-resistant human ovarian cancer xenografts in c.B-17/scid mice by intraperitoneal administration of clostridium perfringens enterotoxin. Cancer Res 2005, 65, 4334–4342. [Google Scholar]
- Yoshida, H.; Sumi, T.; Zhi, X.; Yasui, T.; Honda, K.; Ishiko, O. Claudin-4: A potential therapeutic target in chemotherapy-resistant ovarian cancer. Anticancer Res 2011, 31, 1271–1277. [Google Scholar]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar]
- Yilmaz, M.; Christofori, G. Emt, the cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev 2009, 28, 15–33. [Google Scholar]
- Foty, R.A.; Steinberg, M.S. Cadherin-mediated cell-cell adhesion and tissue segregation in relation to malignancy. Int. J. Dev. Biol 2004, 48, 397–409. [Google Scholar]
- Szasz, A.M.; Nemeth, Z.; Gyorffy, B.; Micsinai, M.; Krenacs, T.; Baranyai, Z.; Harsanyi, L.; Kiss, A.; Schaff, Z.; Tokes, A.M.; et al. Identification of a claudin-4 and e-cadherin score to predict prognosis in breast cancer. Cancer Sci 2011, 102, 2248–2254. [Google Scholar]
- Lee, K.W.; Lee, N.K.; Kim, J.H.; Kang, M.S.; Yoo, H.Y.; Kim, H.H.; Um, S.H.; Kim, S.H. Twist1 causes the transcriptional repression of claudin-4 with prognostic significance in esophageal cancer. Biochem. Biophys. Res. Commun 2012, 423, 454–460. [Google Scholar]
- Ersoz, S.; Mungan, S.; Cobanoglu, U.; Turgutalp, H.; Ozoran, Y. Prognostic importance of claudin-1 and claudin-4 expression in colon carcinomas. Pathol. Res. Pract 2011, 207, 285–289. [Google Scholar]
- Tsutsumi, K.; Sato, N.; Tanabe, R.; Mizumoto, K.; Morimatsu, K.; Kayashima, T.; Fujita, H.; Ohuchida, K.; Ohtsuka, T.; Takahata, S.; et al. Claudin-4 expression predicts survival in pancreatic ductal adenocarcinoma. Ann. Surg. Oncol 2012, 19, S491–S499. [Google Scholar]
- Finch, A.; Shaw, P.; Rosen, B.; Murphy, J.; Narod, S.A.; Colgan, T.J. Clinical and pathologic findings of prophylactic salpingo-oophorectomies in 159 brca1 and brca2 carriers. Gynecol. Oncol 2006, 100, 58–64. [Google Scholar]
- Lee, Y.; Miron, A.; Drapkin, R.; Nucci, M.R.; Medeiros, F.; Saleemuddin, A.; Garber, J.; Birch, C.; Mou, H.; Gordon, R.W.; et al. A candidate precursor to serous carcinoma that originates in the distal fallopian tube. J. Pathol 2007, 211, 26–35. [Google Scholar]
- Karst, A.M.; Levanon, K.; Drapkin, R. Modeling high-grade serous ovarian carcinogenesis from the fallopian tube. Proc. Natl. Acad. Sci. USA 2011, 108, 7547–7552. [Google Scholar]
- Shang, X.; Lin, X.; Manorek, G.; Howell, S.B. Claudin-3 and claudin-4 regulate sensitivity to cisplatin by controlling expression of the copper and cisplatin influx transporter ctr1. Mol. Pharmacol 2013, 83, 85–94. [Google Scholar]
- Litkouhi, B.; Kwong, J.; Lo, C.M.; Smedley, J.G., 3rd; McClane, B.A.; Aponte, M.; Gao, Z.; Sarno, J.L.; Hinners, J.; Welch, W.R.; et al. Claudin-4 overexpression in epithelial ovarian cancer is associated with hypomethylation and is a potential target for modulation of tight junction barrier function using a C-terminal fragment of clostridium perfringens enterotoxin. Neoplasia 2007, 9, 304–314. [Google Scholar]
- Choi, Y.L.; Kim, J.; Kwon, M.J.; Choi, J.S.; Kim, T.J.; Bae, D.S.; Koh, S.S.; In, Y.H.; Park, Y.W.; Kim, S.H.; et al. Expression profile of tight junction protein claudin 3 and claudin 4 in ovarian serous adenocarcinoma with prognostic correlation. Histol. Histopathol 2007, 22, 1185–1195. [Google Scholar]
- Huang, Y.H.; Bao, Y.; Peng, W.; Goldberg, M.; Love, K.; Bumcrot, D.A.; Cole, G.; Langer, R.; Anderson, D.G.; Sawicki, J.A. Claudin-3 gene silencing with sirna suppresses ovarian tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2009, 106, 3426–3430. [Google Scholar]
- Facchetti, F.; Lonardi, S.; Gentili, F.; Bercich, L.; Falchetti, M.; Tardanico, R.; Baronchelli, C.; Lucini, L.; Santin, A.; Murer, B. Claudin 4 identifies a wide spectrum of epithelial neoplasms and represents a very useful marker for carcinoma versus mesothelioma diagnosis in pleural and peritoneal biopsies and effusions. Virchows Archiv 2007, 451, 669–680. [Google Scholar]
- Rodewald, M.; Herr, D.; Duncan, W.C.; Fraser, H.M.; Hack, G.; Konrad, R.; Gagsteiger, F.; Kreienberg, R.; Wulff, C. Molecular mechanisms of ovarian hyperstimulation syndrome: Paracrine reduction of endothelial claudin 5 by hcg in vitro is associated with increased endothelial permeability. Hum. Reprod 2009, 24, 1191–1199. [Google Scholar]
- Dvorak, H.F.; Nagy, J.A.; Feng, D.; Brown, L.F.; Dvorak, A.M. Vascular permeability factor/vascular endothelial growth factor and the significance of microvascular hyperpermeability in angiogenesis. Curr. Top Microbiol. Immunol 1999, 237, 97–132. [Google Scholar]
- Dvorak, H.F. Vascular permeability factor/vascular endothelial growth factor: A critical cytokine in tumor angiogenesis and a potential target for diagnosis and therapy. J. Clin. Oncol 2002, 20, 4368–4380. [Google Scholar]
- Olson, T.A.; Mohanraj, D.; Carson, L.F.; Ramakrishnan, S. Vascular permeability factor gene expression in normal and neoplastic human ovaries. Cancer Res 1994, 54, 276–280. [Google Scholar]
- Hazelton, D.; Nicosia, R.F.; Nicosia, S.V. Vascular endothelial growth factor levels in ovarian cyst fluid correlate with malignancy. Clin. Cancer Res 1999, 5, 823–829. [Google Scholar]
- Herr, D.; Sallmann, A.; Bekes, I.; Konrad, R.; Holzheu, I.; Kreienberg, R.; Wulff, C. Vegf induces ascites in ovarian cancer patients via increasing peritoneal permeability by downregulation of claudin 5. Gynecol. Oncol 2012, 127, 210–216. [Google Scholar]
- Turunen, M.; Talvensaari-Mattila, A.; Soini, Y.; Santala, M. Claudin-5 overexpression correlates with aggressive behavior in serous ovarian adenocarcinoma. Anticancer Res 2009, 29, 5185–5189. [Google Scholar]
- Wang, L.; Xue, Y.; Shen, Y.; Li, W.; Cheng, Y.; Yan, X.; Shi, W.; Wang, J.; Gong, Z.; Yang, G.; et al. Claudin 6: A novel surface marker for characterizing mouse pluripotent stem cells. Cell Res 2012, 22, 1082–1085. [Google Scholar]
- Turksen, K.; Troy, T.C. Claudin-6: A novel tight junction molecule is developmentally regulated in mouse embryonic epithelium. Dev. Dyn 2001, 222, 292–300. [Google Scholar]
- Anderson, W.J.; Zhou, Q.; Alcalde, V.; Kaneko, O.F.; Blank, L.J.; Sherwood, R.I.; Guseh, J.S.; Rajagopal, J.; Melton, D.A. Genetic targeting of the endoderm with claudin-6creer. Dev. Dyn 2008, 237, 504–512. [Google Scholar]
- Birks, D.K.; Kleinschmidt-DeMasters, B.K.; Donson, A.M.; Barton, V.N.; McNatt, S.A.; Foreman, N.K.; Handler, M.H. Claudin 6 is a positive marker for atypical teratoid/rhabdoid tumors. Brain Pathol 2010, 20, 140–150. [Google Scholar]
- Yafang, L.; Qiong, W.; Yue, R.; Xiaoming, X.; Lina, Y.; Mingzi, Z.; Ting, Z.; Yulin, L.; Chengshi, Q. Role of estrogen receptor-alpha in the regulation of claudin-6 expression in breast cancer cells. J. Breast Cancer 2011, 14, 20–27. [Google Scholar]
- Rendon-Huerta, E.; Teresa, F.; Teresa, G.M.; Xochitl, G.S.; Georgina, A.F.; Veronica, Z.Z.; Montano, L.F. Distribution and expression pattern of claudins 6, 7, and 9 in diffuse- and intestinal-type gastric adenocarcinomas. J. Gastrointest Cancer 2010, 41, 52–59. [Google Scholar]
- Lal-Nag, M.; Battis, M.; Santin, A.D.; Morin, P.J. Claudin-6: A novel receptor for cpe-mediated cytotoxicity in ovarian cancer. Oncogenesis 2012, 1, e33. [Google Scholar]
- Smedley, J.G., 3rd; McClane, B.A. Fine mapping of the N-terminal cytotoxicity region of clostridium perfringens enterotoxin by site-directed mutagenesis. Infect Immun 2004, 72, 6914–6923. [Google Scholar]
- Kim, C.J.; Lee, J.W.; Choi, J.J.; Choi, H.Y.; Park, Y.A.; Jeon, H.K.; Sung, C.O.; Song, S.Y.; Lee, Y.Y.; Choi, C.H.; et al. High claudin-7 expression is associated with a poor response to platinum-based chemotherapy in epithelial ovarian carcinoma. Eur. J. Cancer 2011, 47, 918–925. [Google Scholar]
- Usami, Y.; Chiba, H.; Nakayama, F.; Ueda, J.; Matsuda, Y.; Sawada, N.; Komori, T.; Ito, A.; Yokozaki, H. Reduced expression of claudin-7 correlates with invasion and metastasis in squamous cell carcinoma of the esophagus. Human Pathol 2006, 37, 569–577. [Google Scholar]
- Al Moustafa, A.E.; Alaoui-Jamali, M.A.; Batist, G.; Hernandez-Perez, M.; Serruya, C.; Alpert, L.; Black, M.J.; Sladek, R.; Foulkes, W.D. Identification of genes associated with head and neck carcinogenesis by cdna microarray comparison between matched primary normal epithelial and squamous carcinoma cells. Oncogene 2002, 21, 2634–2640. [Google Scholar]
- Sheehan, G.M.; Kallakury, B.V.; Sheehan, C.E.; Fisher, H.A.; Kaufman, R.P., Jr; Ross, J.S. Loss of claudins-1 and -7 and expression of claudins-3 and -4 correlate with prognostic variables in prostatic adenocarcinomas. Human pathology 2007, 38, 564–569. [Google Scholar]
- Sauer, T.; Pedersen, M.K.; Ebeltoft, K.; Naess, O. Reduced expression of claudin-7 in fine needle aspirates from breast carcinomas correlate with grading and metastatic disease. Cytopathology 2005, 16, 193–198. [Google Scholar]
- Katahira, J.; Sugiyama, H.; Inoue, N.; Horiguchi, Y.; Matsuda, M.; Sugimoto, N. Clostridium perfringens enterotoxin utilizes two structurally related membrane proteins as functional receptors in vivo. J. Biol. Chem 1997, 272, 26652–26658. [Google Scholar]
- Cocco, E.; Casagrande, F.; Bellone, S.; Richter, C.E.; Bellone, M.; Todeschini, P.; Holmberg, J.C.; Fu, H.H.; Montagna, M.K.; Mor, G.; et al. Clostridium perfringens enterotoxin carboxy-terminal fragment is a novel tumor-homing peptide for human ovarian cancer. BMC Cancer 2010, 10, 349. [Google Scholar]
- Kakutani, H.; Kondoh, M.; Saeki, R.; Fujii, M.; Watanabe, Y.; Mizuguchi, H.; Yagi, K. Claudin-4-targeting of diphtheria toxin fragment a using a C-terminal fragment of clostridium perfringens enterotoxin. Eur. J. Pharm. Biopharm 2010, 75, 213–217. [Google Scholar]
- Ebihara, C.; Kondoh, M.; Hasuike, N.; Harada, M.; Mizuguchi, H.; Horiguchi, Y.; Fujii, M.; Watanabe, Y. Preparation of a claudin-targeting molecule using a C-terminal fragment of clostridium perfringens enterotoxin. J. Pharmacol. Exp. Ther 2006, 316, 255–260. [Google Scholar]
- Yuan, X.; Lin, X.; Manorek, G.; Kanatani, I.; Cheung, L.H.; Rosenblum, M.G.; Howell, S.B. Recombinant cpe fused to tumor necrosis factor targets human ovarian cancer cells expressing the claudin-3 and claudin-4 receptors. Mol. Cancer Ther 2009, 8, 1906–1915. [Google Scholar]
- Michl, P.; Buchholz, M.; Rolke, M.; Kunsch, S.; Lohr, M.; McClane, B.; Tsukita, S.; Leder, G.; Adler, G.; Gress, T.M. Claudin-4: A new target for pancreatic cancer treatment using clostridium perfringens enterotoxin. Gastroenterology 2001, 121, 678–684. [Google Scholar]
- Long, H.; Crean, C.D.; Lee, W.H.; Cummings, O.W.; Gabig, T.G. Expression of clostridium perfringens enterotoxin receptors claudin-3 and claudin-4 in prostate cancer epithelium. Cancer Res 2001, 61, 7878–7881. [Google Scholar]
- Kominsky, S.L.; Vali, M.; Korz, D.; Gabig, T.G.; Weitzman, S.A.; Argani, P.; Sukumar, S. Clostridium perfringens enterotoxin elicits rapid and specific cytolysis of breast carcinoma cells mediated through tight junction proteins claudin 3 and 4. Am. J. Pathol . 2004, 164, 1627–1633. [Google Scholar]
- McClane, B.A. An overview of clostridium perfringens enterotoxin. Toxicon 1996, 34, 1335–1343. [Google Scholar]
- Kokai-Kun, J.F.; McClane, B.A. Deletion analysis of the clostridium perfringens enterotoxin. Infect Immun 1997, 65, 1014–1022. [Google Scholar]
- Kokai-Kun, J.F.; Benton, K.; Wieckowski, E.U.; McClane, B.A. Identification of a clostridium perfringens enterotoxin region required for large complex formation and cytotoxicity by random mutagenesis. Infect Immun 1999, 67, 5634–5641. [Google Scholar]
- Kokai-Kun, J.F.; McClane, B.A. Evidence that a region(s) of the clostridium perfringens enterotoxin molecule remains exposed on the external surface of the mammalian plasma membrane when the toxin is sequestered in small or large complexes. Infect Immun 1996, 64, 1020–1025. [Google Scholar]
- Yamamoto, A.; Uchiyama, T.; Nishikawa, R.; Fujita, T.; Muranishi, S. Effectiveness and toxicity screening of various absorption enhancers in the rat small intestine: Effects of absorption enhancers on the intestinal absorption of phenol red and the release of protein and phospholipids from the intestinal membrane. J. Pharm. Pharmacol 1996, 48, 1285–1289. [Google Scholar]
- Kondoh, M.; Takahashi, A.; Fujii, M.; Yagi, K.; Watanabe, Y. A novel strategy for a drug delivery system using a claudin modulator. Biol. Pharm. Bull 2006, 29, 1783–1789. [Google Scholar]
- Gao, Z.; Xu, X.; McClane, B.; Zeng, Q.; Litkouhi, B.; Welch, W.R.; Berkowitz, R.S.; Mok, S.C.; Garner, E.I. C-terminus of clostridium perfringens enterotoxin downregulates cldn4 and sensitizes ovarian cancer cells to taxol and carboplatin. Clin. Cancer Res 2011, 17, 1065–1074. [Google Scholar]
- Alberts, D.S.; Markman, M.; Armstrong, D.; Rothenberg, M.L.; Muggia, F.; Howell, S.B. Intraperitoneal therapy for stage iii ovarian cancer: A therapy whose time has come! J. Clin. Oncol 2002, 20, 3944–3946. [Google Scholar]
- Kominsky, S.L.; Tyler, B.; Sosnowski, J.; Brady, K.; Doucet, M.; Nell, D.; Smedley, J.G., 3rd; McClane, B.; Brem, H.; Sukumar, S. Clostridium perfringens enterotoxin as a novel-targeted therapeutic for brain metastasis. Cancer Res 2007, 67, 7977–7982. [Google Scholar]
- Santin, A.D.; Bellone, S.; Marizzoni, M.; Palmieri, M.; Siegel, E.R.; McKenney, J.K.; Hennings, L.; Comper, F.; Bandiera, E.; Pecorelli, S. Overexpression of claudin-3 and claudin-4 receptors in uterine serous papillary carcinoma: Novel targets for a type-specific therapy using clostridium perfringens enterotoxin (cpe). Cancer 2007, 109, 1312–1322. [Google Scholar]
- Walther, W.; Petkov, S.; Kuvardina, O.N.; Aumann, J.; Kobelt, D.; Fichtner, I.; Lemm, M.; Piontek, J.; Blasig, I.E.; Stein, U.; et al. Novel clostridium perfringens enterotoxin suicide gene therapy for selective treatment of claudin-3- and -4-overexpressing tumors. Gene Ther 2012, 19, 494–503. [Google Scholar]
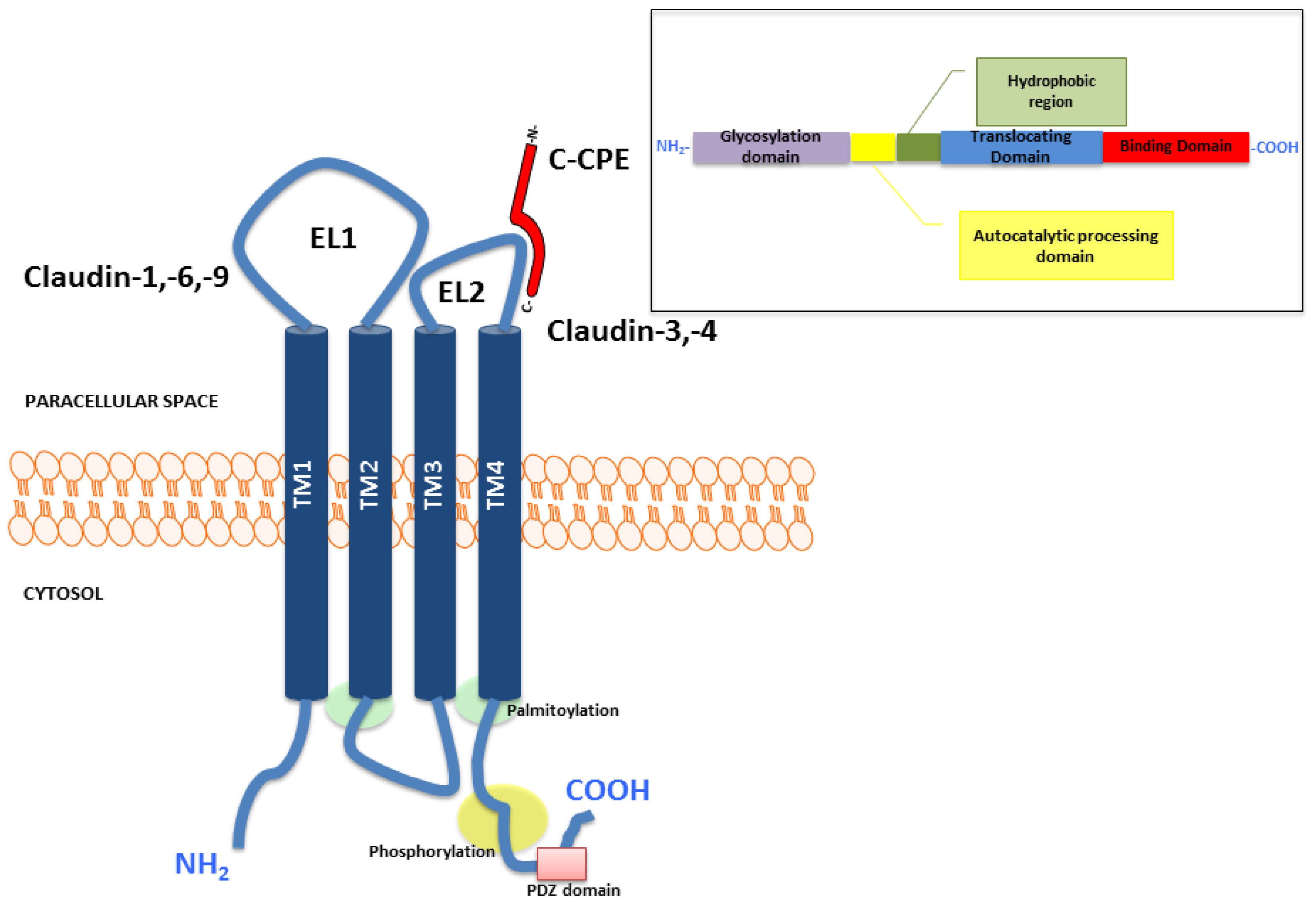


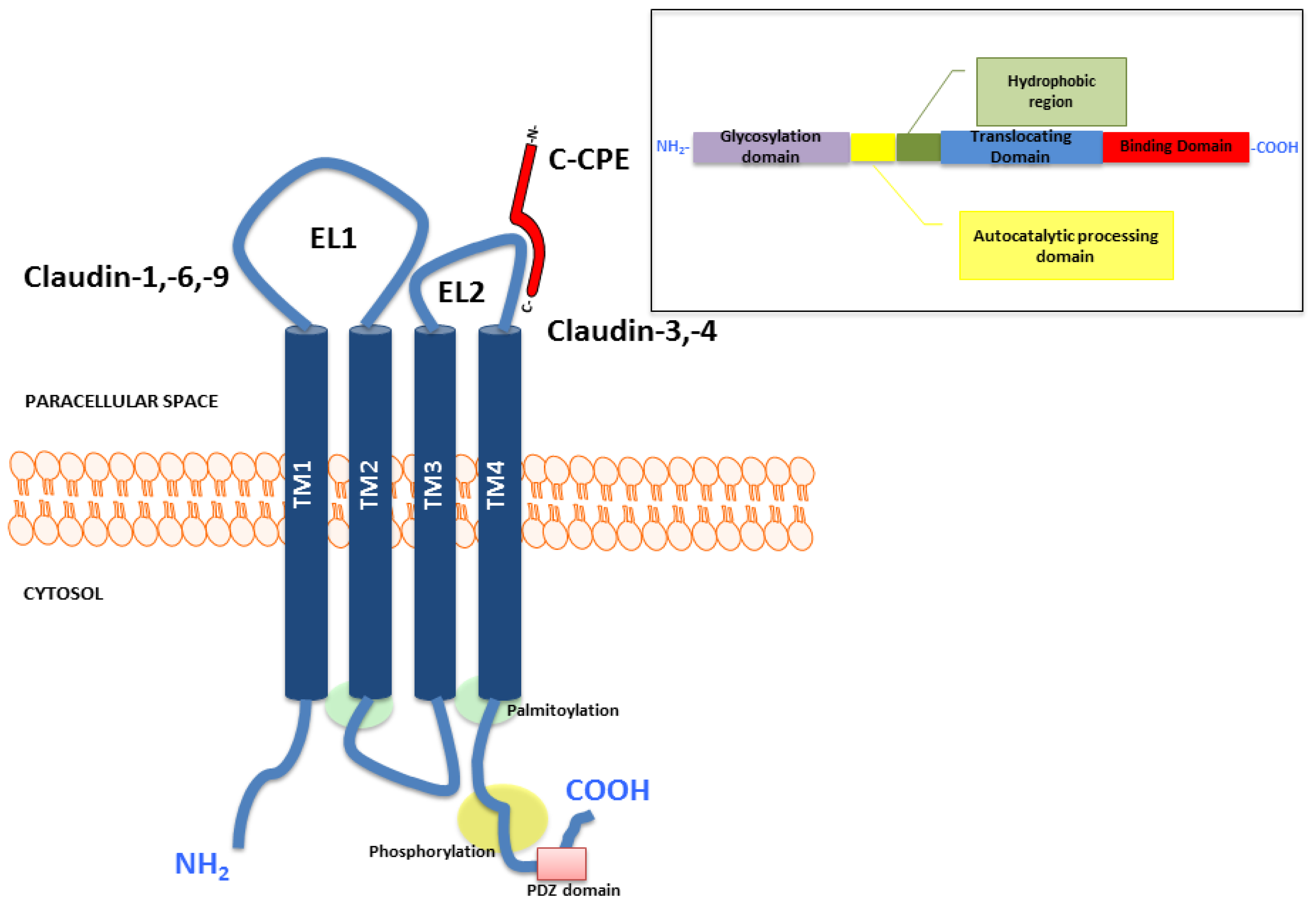{kind=link}
{kind=link}
| Claudin | Function | Tissue specificity | Involvement in disease |
|---|---|---|---|
| Claudin-1 | TJ-specific obliteration of the intercellular space through Ca2+-independent cell-adhesion activity. Acts as a co-receptor for HCV entry into hepatic cells | Strongly expressed in liver and kidney. Also expressed in heart, brain, spleen, lung and testis | Ichthyosis [61] |
| Claudin-3 | TJ-specific obliteration of the intercellular space through Ca2+-independent cell-adhesion activity (CPE is the natural ligand) | Strongly expressed in ovary, lung, pancreas, salivary gland, kidney, adrenal, small intestine, colon and thyroid | Williams-Beuren syndrome [62] |
| Claudin-4 | TJ-specific obliteration of the intercellular space (CPE is the natural ligand) | Strongly expressed in ovary, lung, pancreas, salivary gland, kidney, adrenal, small intestine, colon and thyroid | Williams-Beuren syndrome [62] |
| Claudin-5 | Target molecule of hypoxia | Strongly expressed in vascular endothelial cells. Transiently expressed during development of RPE. Expressed in lung | Velocardiofacial syndrome [63] |
| Claudin-7 | TJ-specific obliteration of the intercellular space. Co-localizes with EPCAM at the lateral cell membrane and TJ | Strongly expressed in kidney, GI tract, thyroid, adrenal gland and lung. Also expressed in prostate tissue | Related to ability of breast cancer cells to disseminate. Downregulation correlates with histological grade [57] |
| Tumor type | Claudin gene | Expression compared to normal tissues | References |
|---|---|---|---|
| Ovarian | CLDN3 | High | [16,18,54,56] |
| CLDN4 | High | [16,18,43] | |
| CLDN7 | High | [18,84–86] | |
| CLDN16 | High | [17] | |
| Endometrial | CLDN2 | High | [87] |
| CLDN3 | High | [87–89] | |
| CLDN4 | High | [53,88] | |
| Cervical | CLDN1 | High | [90,91] |
| CLDN2 | High | [91] | |
| CLDN4 | High | [91,92] | |
| CLDN7 | High | [90,92] | |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
English, D.P.; Santin, A.D. Claudins Overexpression in Ovarian Cancer: Potential Targets for Clostridium Perfringens Enterotoxin (CPE) Based Diagnosis and Therapy. Int. J. Mol. Sci. 2013, 14, 10412-10437. https://doi.org/10.3390/ijms140510412
English DP, Santin AD. Claudins Overexpression in Ovarian Cancer: Potential Targets for Clostridium Perfringens Enterotoxin (CPE) Based Diagnosis and Therapy. International Journal of Molecular Sciences. 2013; 14(5):10412-10437. https://doi.org/10.3390/ijms140510412
Chicago/Turabian StyleEnglish, Diana P., and Alessandro D. Santin. 2013. "Claudins Overexpression in Ovarian Cancer: Potential Targets for Clostridium Perfringens Enterotoxin (CPE) Based Diagnosis and Therapy" International Journal of Molecular Sciences 14, no. 5: 10412-10437. https://doi.org/10.3390/ijms140510412
APA StyleEnglish, D. P., & Santin, A. D. (2013). Claudins Overexpression in Ovarian Cancer: Potential Targets for Clostridium Perfringens Enterotoxin (CPE) Based Diagnosis and Therapy. International Journal of Molecular Sciences, 14(5), 10412-10437. https://doi.org/10.3390/ijms140510412