Effectiveness of Chitosan against Mature Biofilms Formed by Food Related Bacteria

Abstract
:1. Introduction
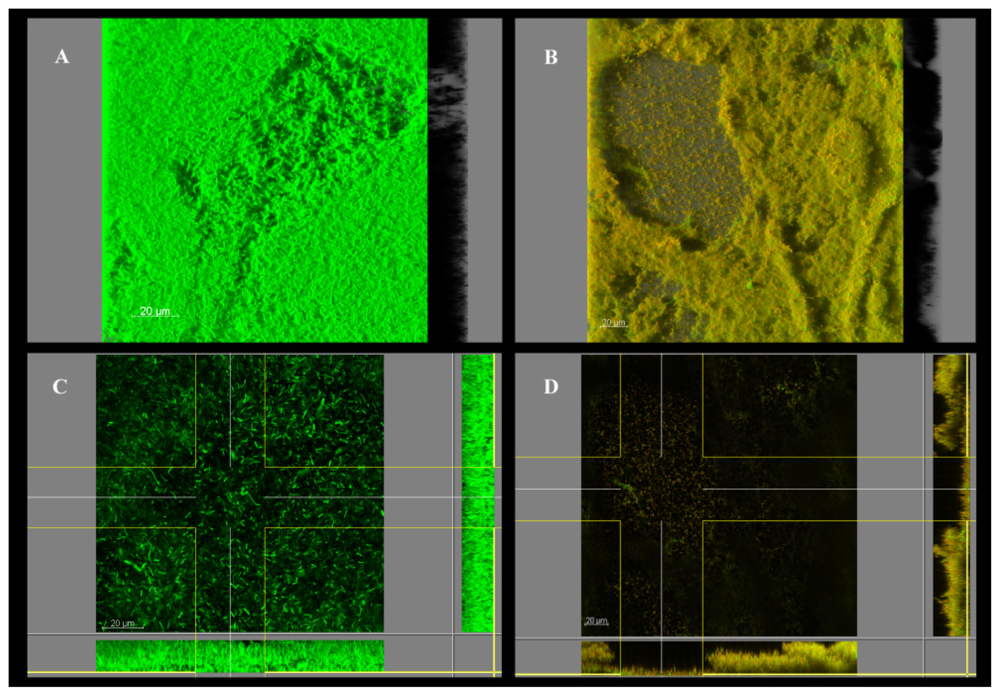
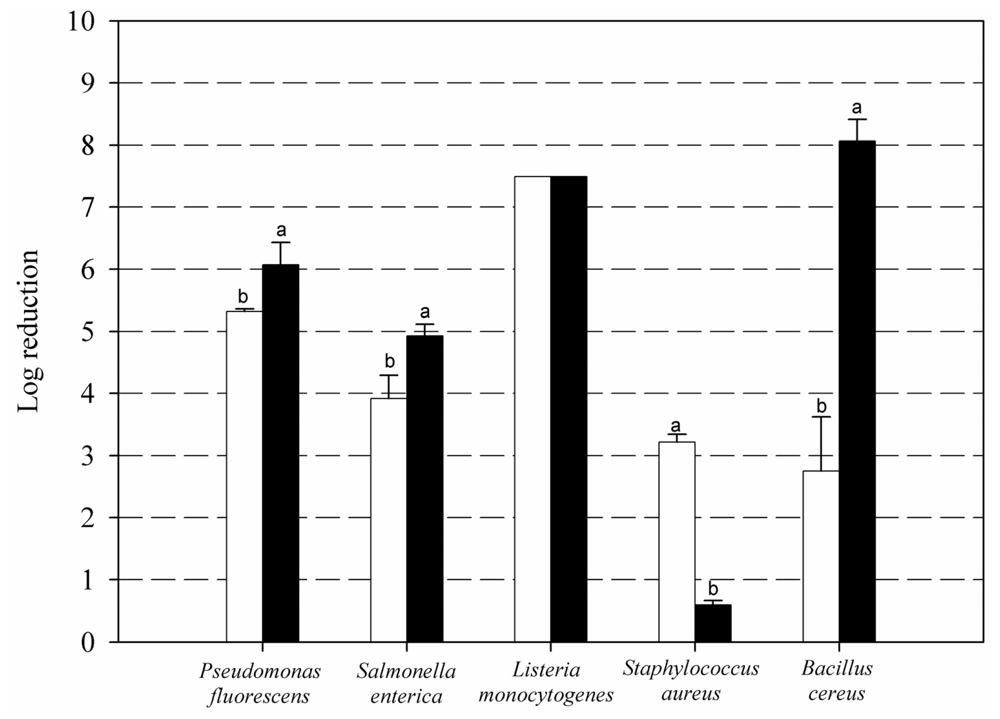
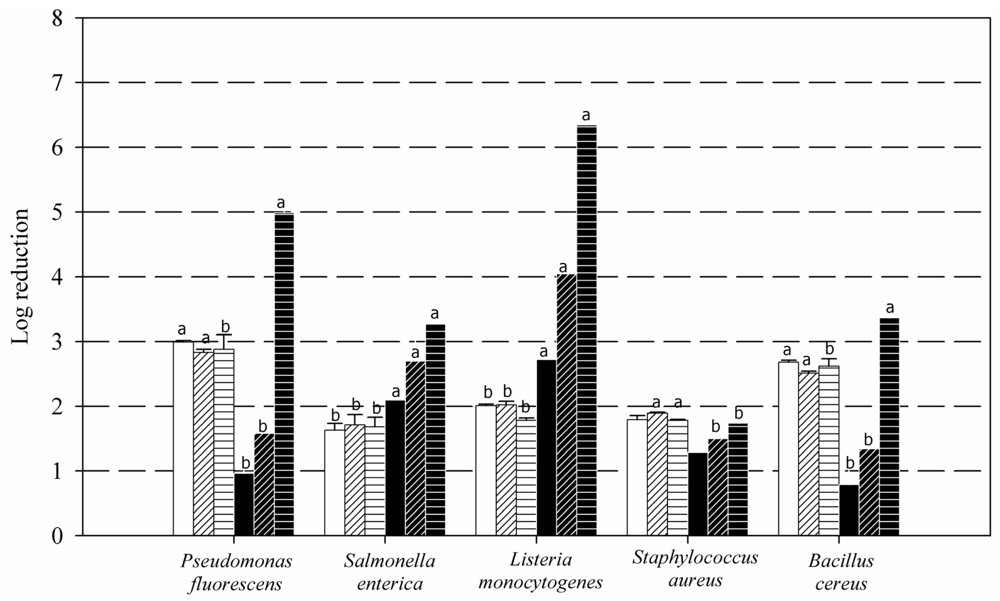
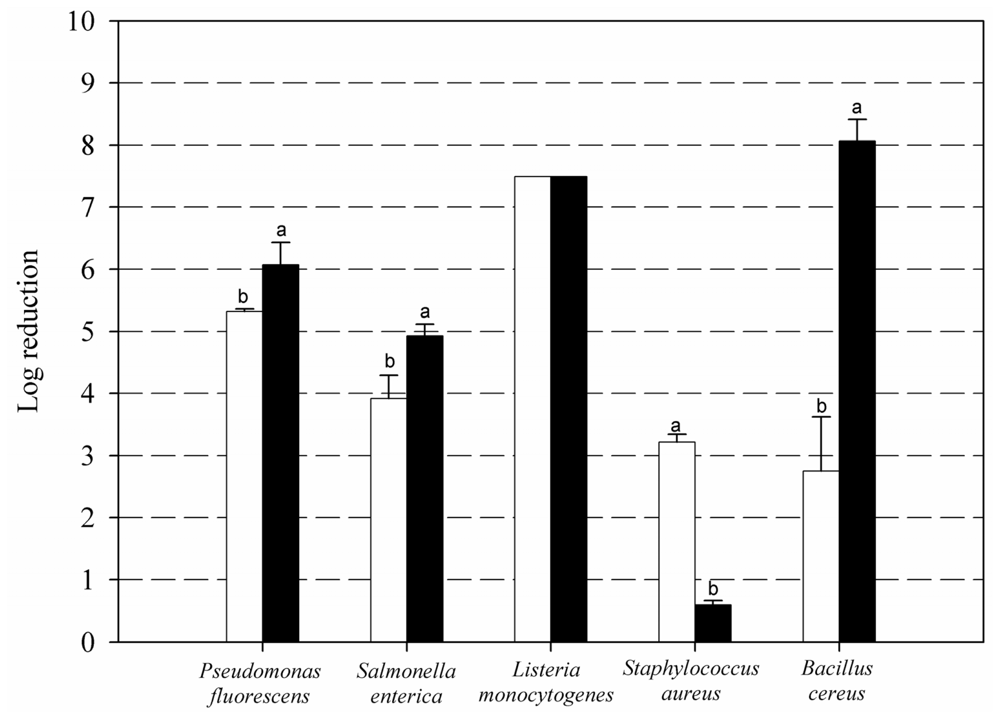
2. Results and Discussion
3. Experimental Section
3.1. Bacterial Strains
3.2. Experimental Systems
3.3. Chitosan Preparations and Treatments
3.4. Cell Recovery and Counting
3.5. Statistical Analysis
3.6. Confocal Scanning Laser Microscopy (CSLM)
4. Conclusions
Acknowledgements
References
- Costerton, JW. Toward a unified biofilm theory. In The Biofilm Primer, 1st ed; Heidelberg,, CE, Ed.; Springer: Berlin, Germany, 2007; Volume 1, pp. 169–180. [Google Scholar]
- Sauer, K. Minireview: The genomics and proteomics of biofilm formation. Genome Biol 2003, 4, 1–4. [Google Scholar]
- Flemming, HC; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol 2010, 8, 623–632. [Google Scholar]
- Verran, J; Airey, P; Packer, A; Whitehead, KA. Microbial retention on open food contact surfaces and implications for food contamination. Adv. Appl. Microbiol 2008, 64, 223–246. [Google Scholar]
- Shi, X; Zhu, X. Biofilm formation and food safety in food industries. Trends Food Sci. Technol 2009, 20, 407–413. [Google Scholar]
- Friedman, M; Juneja, VK. Review of antimicrobial and antioxidative activities of chitosans in food. J. Food Prot 2010, 73, 1737–1761. [Google Scholar]
- Kurita, K. Chitin and chitosan: Functional biopolymers from marine crustaceans. Mar. Biotechnol 2006, 8, 203–226. [Google Scholar]
- Prashanth, KVH; Tharanathan, RN. Chitin/chitosan: Modifications and their unlimited application potential an overview. Trends Food Sci. Technol 2007, 18, 117–131. [Google Scholar]
- Wang, SL; Lin, HT; Liang, TW; Chen, YJ; Yen, YH; Guo, SP. Reclamation of chitinous materials by bromelain for the preparation of antitumor and antifungal materials. Bioresource Technol 2008, 99, 4386–4393. [Google Scholar]
- Roller, S; Covill, N. The antifungal properties of chitosan in laboratory media and apple juice. Int. J. Food Microbiol 1999, 47, 67–77. [Google Scholar]
- Bae, K; Jun, EJ; Lee, SM; Paik, DI; Kim, JB. Effect of water-soluble reduced chitosan on Streptococcus mutans, plaque regrowth and biofilm vitality. Clin. Oral Invest 2006, 10, 102–107. [Google Scholar]
- Je, JY; Kim, SK. Chitosan derivates killed bacteria by disrupting the outer and inner membrane. J. Agric. Food Chem 2006, 54, 6629–6633. [Google Scholar]
- Kumar, VAB; Varadaraj, MC; Gowda, LR; Tharanathan, RN. Characterization of chitooligosaccharides prepared by chitosanolysis with the aid of papain and pronase, and their bactericidal action. Biochem. J 2005, 391, 167–175. [Google Scholar]
- Donzelli, BGG; Ostroff, G; Harman, GE. Enhanced enzymatic hydrolysis of langostino shell chitin with mixtures of enzymes from bacterial and fungal sources. Carbohydr. Res 2003, 338, 1823–1833. [Google Scholar]
- Kumar, VAB; Varadaraj, MC; Gowda, LR; Tharanathan, RN. Low molecular weight chitosans-Preparation with the aid of pronase, characterization and their bactericidal activity towards Bacillus cereus and Escherichia coli. Biochim Biophys Acta 2007, 495–505. [Google Scholar]
- Yalpani, M; Pantaleone, D. An examination of the unusual susceptibilities of aminoglycans to enzymatic hydrolysis. Carbohydr. Res 1994, 256, 159–175. [Google Scholar]
- Orgaz, B; Neufeld, RJ; Sanjose, C. Single-step biofilm removal with delayed release encapsulated pronase mixed with soluble enzymes. Enz. Microb. Technol 2007, 40, 1045–1051. [Google Scholar]
- Kim, IY; Seo, SJ; Moon, HS; Yoo, MK; Park, IY; Kim, BC; Cho, CS. Chitosan and its derivatives for tissue engineering applications. Biotechnol. Adv 2008, 26, 1–21. [Google Scholar]
- Chhabra, P; Huang, YW; Frank, JF; Chmielewski, R; Gates, K. Fate of Staphylococcus aureus, Salmonella enterica serovar Typhimurium, and Vibrio vulnificus in raw oysters treated with chitosan. J. Food Prot 2006, 69, 1600–1604. [Google Scholar]
- Fernandez-Saiz, P; Soler, C; Lagarón, JM; Ocio, J. Effects of chitosan films on the growth of Listeria monocytogenes, Staphylococcus aureus and Salmonella spp. in laboratory media and in fish soup. Int. J. Food Microbiol 2010, 137, 87–294. [Google Scholar]
- Tsai, GJ; Tsai, MT; Lee, JM; Zhong, MZ. Effects of chitosan and low-molecular-weight chitosan on Bacillus cereus and application in the preservation of cooked rice. J. Food Prot 2006, 69, 2168–2175. [Google Scholar]
- Fernandes, JC; Tavaria, FK; Soares, JC; Ramos, OS; Monteiro, MJ; Pintado, ME; Malcasa, FX. Antimicrobial effects of chitosan and chitooligosaccharides, upon Staphylococcus aureus and Escherichia coli, in food model systems. Food Microbiol 2008, 25, 922–928. [Google Scholar]
- Carlson, RP; William, RT; Davison, M; Stewart, PS. Anti-biofilm properties of chitosan-coated surfaces. J. Biomater. Sci. Polym 2008, 19, 1035–1046. [Google Scholar]
- van der Mei, HC; Engels, EV; Dijkstra, RJB; Busscher, HJ. Chitosan adsorption to salivary pellicles. Eur. J. Oral Sci 2007, 115, 303–307. [Google Scholar]
- Busscher, HJ; Engels, E; Rene, JB; Dijkstra; van der Mei, HC. Influence of a chitosan on oral bacterial adhesion and growth in vitro. Eur. J. Oral Sci 2008, 116, 493–495. [Google Scholar]
- Martinez, LR; Mihu, MR; Han, G; Frases, S; Cordero, RJB; Casadevall, A; Friedman, JM; Nosanchuk, JD. The use of chitosan to damage Cryptococcus neoformans biofilms. Biomaterials 2010, 31, 669–679. [Google Scholar]
- Flemming, HC. Biofouling in water systems-cases, causes and countermeasures. Appl. Microbiol. Biotechnol 2002, 59, 629–640. [Google Scholar]
- Donlan, RM; Costerton, JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev 2002, 15, 167–193. [Google Scholar]
- Narahashi, Y. Pronase. Methods Enzymol 1970, 9, 651–664. [Google Scholar]
- Rieu, A; Briandet, R; Habimana, O; Garmyn, D; Guzzo, J; Piveteau, P. Listeria monocytogenes EGD-e biofilms: no mushrooms but a network of knitted chains. Appl. Environ. Microbiol 2008, 74, 4491–4497. [Google Scholar]
- Bridier, A; Dubois-Brissonnet, F; Boubetra, A; Thomas, V; Briandet, R. The biofilm architecture of sixty opportunistic pathogens deciphered using a high throughput CLSM method. J. Microbiol. Methods 2010, 82, 64–70. [Google Scholar]
- Kives, J; Orgaz, B; Sanjose, C. Polysaccharide differences between planktonic and biofilm-associated EPS from Pseudomonas fluorescens. Colloids Surf. B: Biointerfaces 2006, 50, 104–108. [Google Scholar]
- Sutherland, IW. The biofilm matrix-an immobilized but dynamic microbial environment. Environ. Trends Microbiol 2001, 9, 222–227. [Google Scholar]
- Sutherland, IW. Kamerling, JP, Ed.; Comprehensive Glycoscience; Elsevier: Doordrecht, The Netherlands, 2007; Volume 2, pp. 521–558. [Google Scholar]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol 2002, 43, 1367–1378. [Google Scholar]
- Gilbert, P; McBain, AJ; Rickard, AH. Formation of microbial biofilm in hygienic situations: A problem of control. Int. Biodeterior. Biodegr 2003, 51, 245–248. [Google Scholar]
- Chavant, P; Gaillard-Martinie, B; Hebraud, M. Antimicrobial effects of sanitizers against planktonic and sessile Listeria monocytogenes cells according to the growth phase. FEMS Microbiol. Lett 2004, 236, 241–248. [Google Scholar]
- Cabo, ML; Herrera, JJ; Crespo, MD; Pastoriza, L. Comparison among the effectiveness of ozone, nisin and benzalkonium chloride for the elimination of planktonic cells and biofilms of Staphylococcus aureus CECT4459 on polypropylene. Food Control 2009, 20, 521–525. [Google Scholar]
- Raafat, D; Von Bargen, K; Haas, A; Sahl, HG. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol 2008, 74, 3764–3773. [Google Scholar]
- Kornacki, JL; Gurtler, JB. Incidence and control of Listeria in food processing facilities. In Listeria, listeriosis and food safety, 3rd ed; Ryser, ET, Marth, EH, Eds.; CRC Press: Boca Raton, FL USA, 2007; pp. 681–766. [Google Scholar]
- Eaton, P; Fernandes, JC; Pereira, E; Pintado, ME; Malcasa, FX. Atomic force microscopy study of the antibacterial effects of chitosans on Escherichia coli and Staphylococcus aureus. Ultramicroscopy 2008, 108, 1128–1134. [Google Scholar]
- Park, NHK; Lee, NY; Meyers, SP. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol 2002, 74, 65–72. [Google Scholar]
- Guillamon, E; Garcia-Lafuente, A; Lozano, M. Edible mushrooms: Role in the prevention of cardiovascular diseases. Fitoterapia 2010, 81, 715–723. [Google Scholar]
- Kives, J; Guadarrama, D; Orgaz, B; Rivera-Sen, A; Vazquez, J; SanJose, C. Interactions in biofilms of Lactococcus lactis ssp. cremoris and Pseudomonas fluorescens cultured in cold UHT milk. J. Dairy Sci 2005, 88, 4165–4171. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Serovar | Log reduction | SD |
|---|---|---|---|
| Scott A | 4b | 4.04a | 0.10 |
| INIA H66a | 1/2a | 3.02b | 0.62 |
| INIA H66b | 1/2c | 2.63b | 0.28 |
| INIA H63a | 1/2b | 1.47d | 0.18 |
| INIA CAL17a | 4b | 2.01c | 0.08 |
| F6861 | 4b | 1.84c,d | 0.08 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Orgaz, B.; Lobete, M.M.; Puga, C.H.; San Jose, C. Effectiveness of Chitosan against Mature Biofilms Formed by Food Related Bacteria. Int. J. Mol. Sci. 2011, 12, 817-828. https://doi.org/10.3390/ijms12010817
Orgaz B, Lobete MM, Puga CH, San Jose C. Effectiveness of Chitosan against Mature Biofilms Formed by Food Related Bacteria. International Journal of Molecular Sciences. 2011; 12(1):817-828. https://doi.org/10.3390/ijms12010817
Chicago/Turabian StyleOrgaz, Belen, Maria M. Lobete, Carmen H. Puga, and Carmen San Jose. 2011. "Effectiveness of Chitosan against Mature Biofilms Formed by Food Related Bacteria" International Journal of Molecular Sciences 12, no. 1: 817-828. https://doi.org/10.3390/ijms12010817
APA StyleOrgaz, B., Lobete, M. M., Puga, C. H., & San Jose, C. (2011). Effectiveness of Chitosan against Mature Biofilms Formed by Food Related Bacteria. International Journal of Molecular Sciences, 12(1), 817-828. https://doi.org/10.3390/ijms12010817