Determination of Mineral Content in Methanolic Safflower (Carthamus tinctorius L.) Seed Extract and Its Effect on Osteoblast Markers


Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparison of mineral content between aqueous and methanolic safflower seed extracts
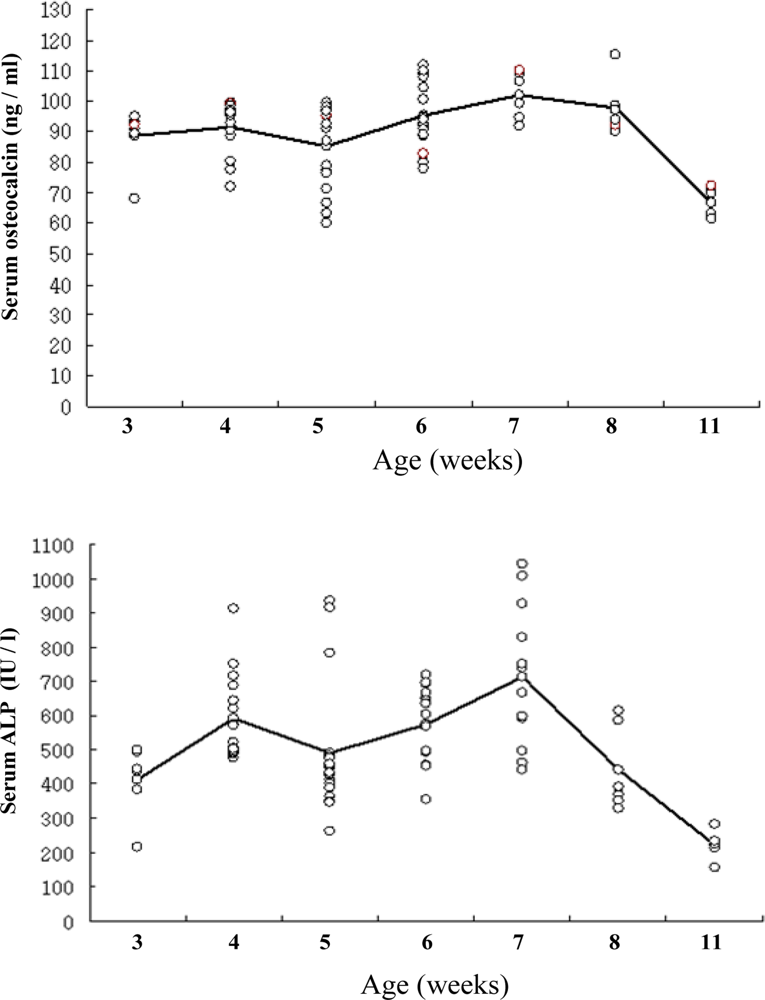
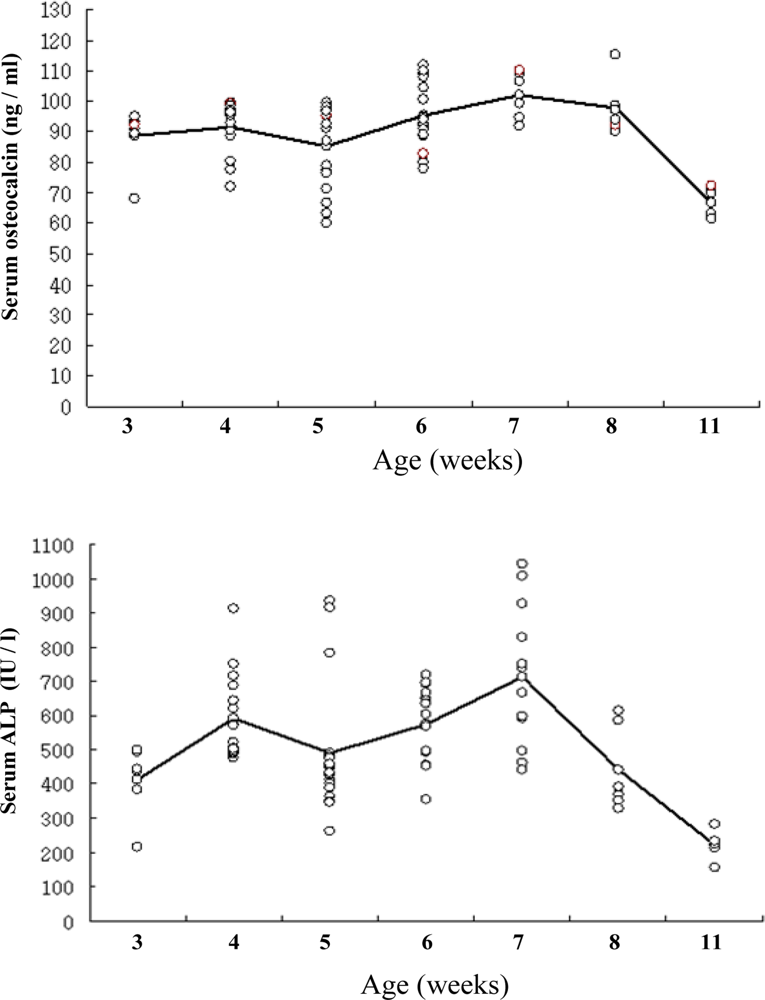
2.2. Dependence of OC content and B-ALP activity on aging of rats
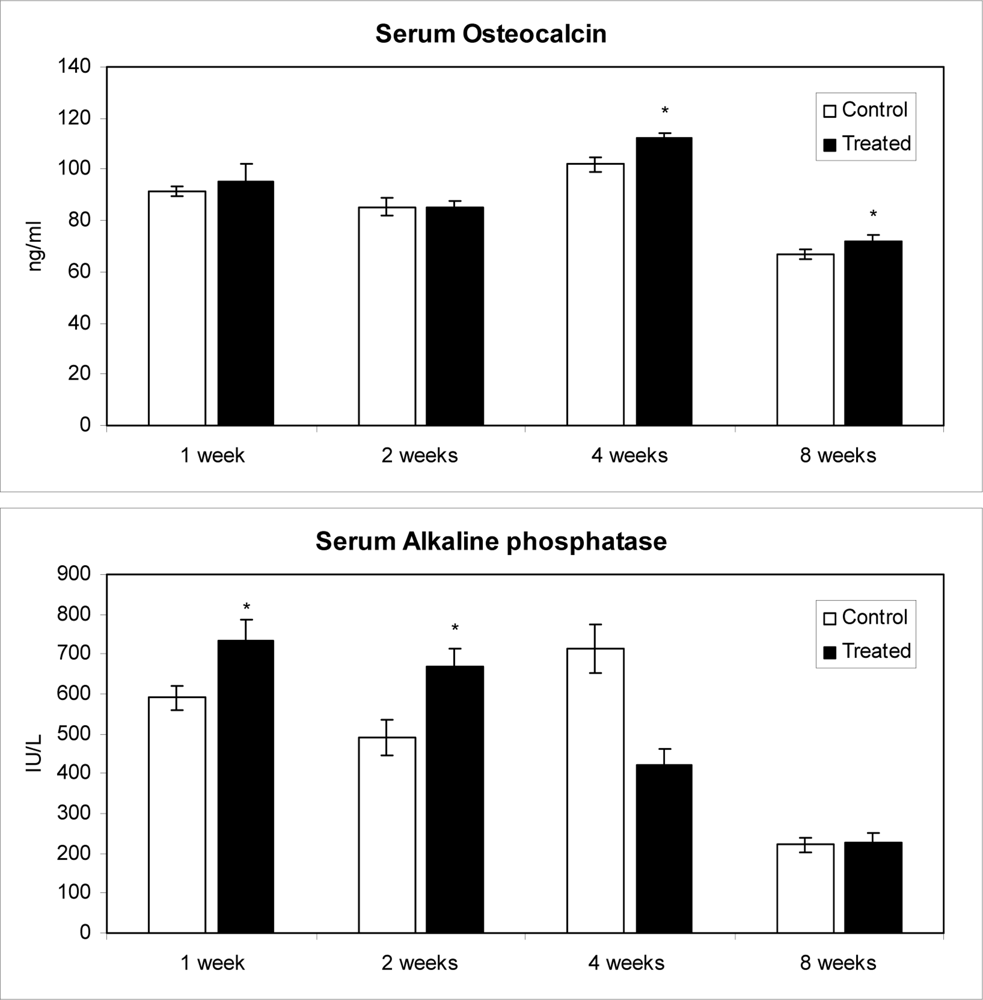
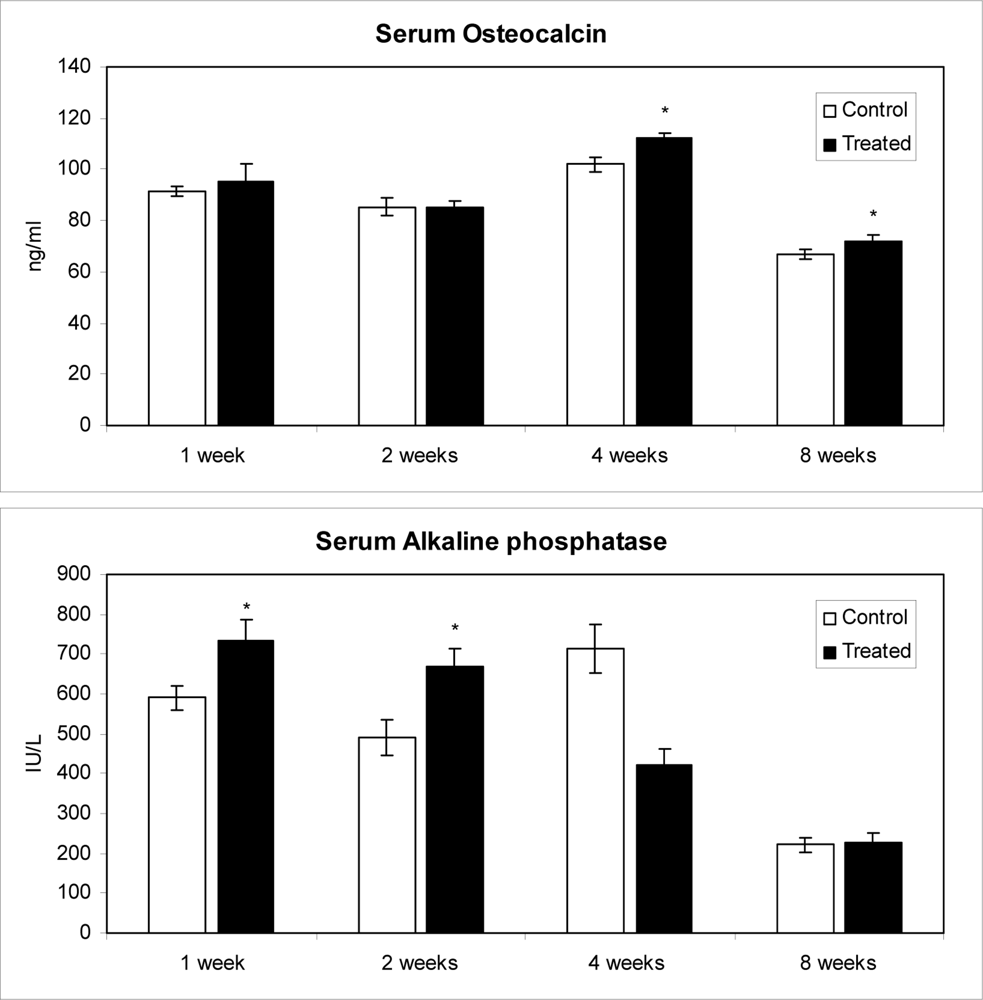
2.3. Effects of methanolic safflower seed extracts on OC contents and B-ALP activities
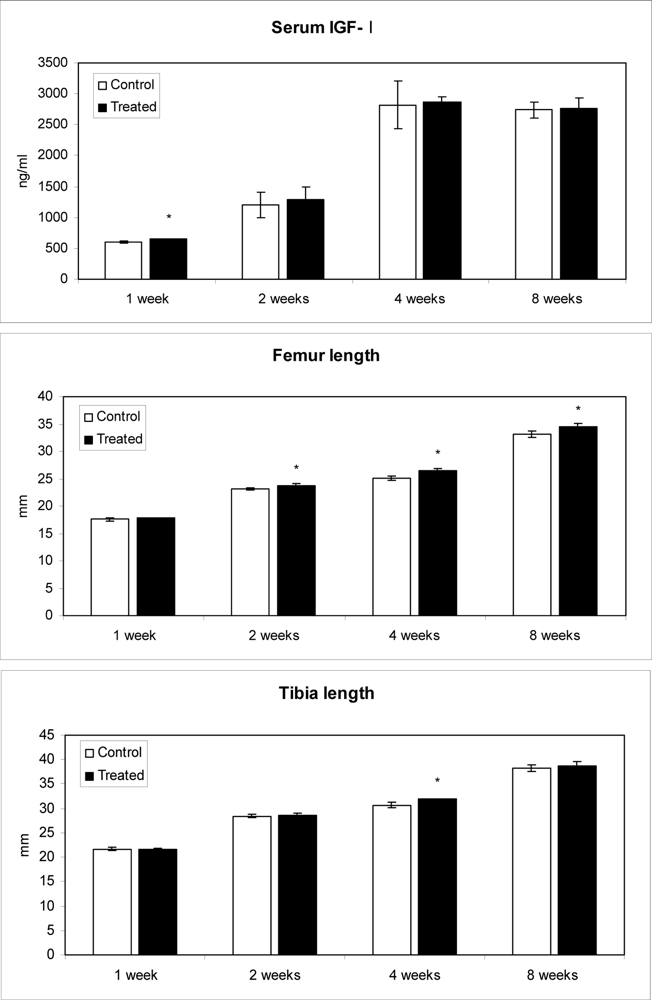
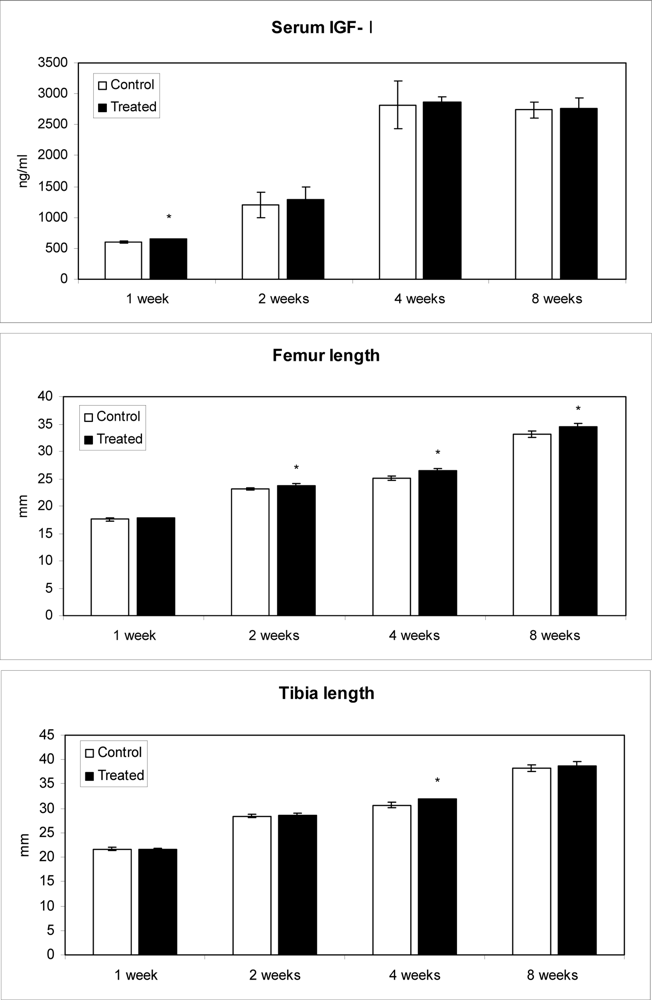
2.4. Effect of methanolic safflower seeds extract on serum IGF-I levels and long bone growth
3. Experimental Section
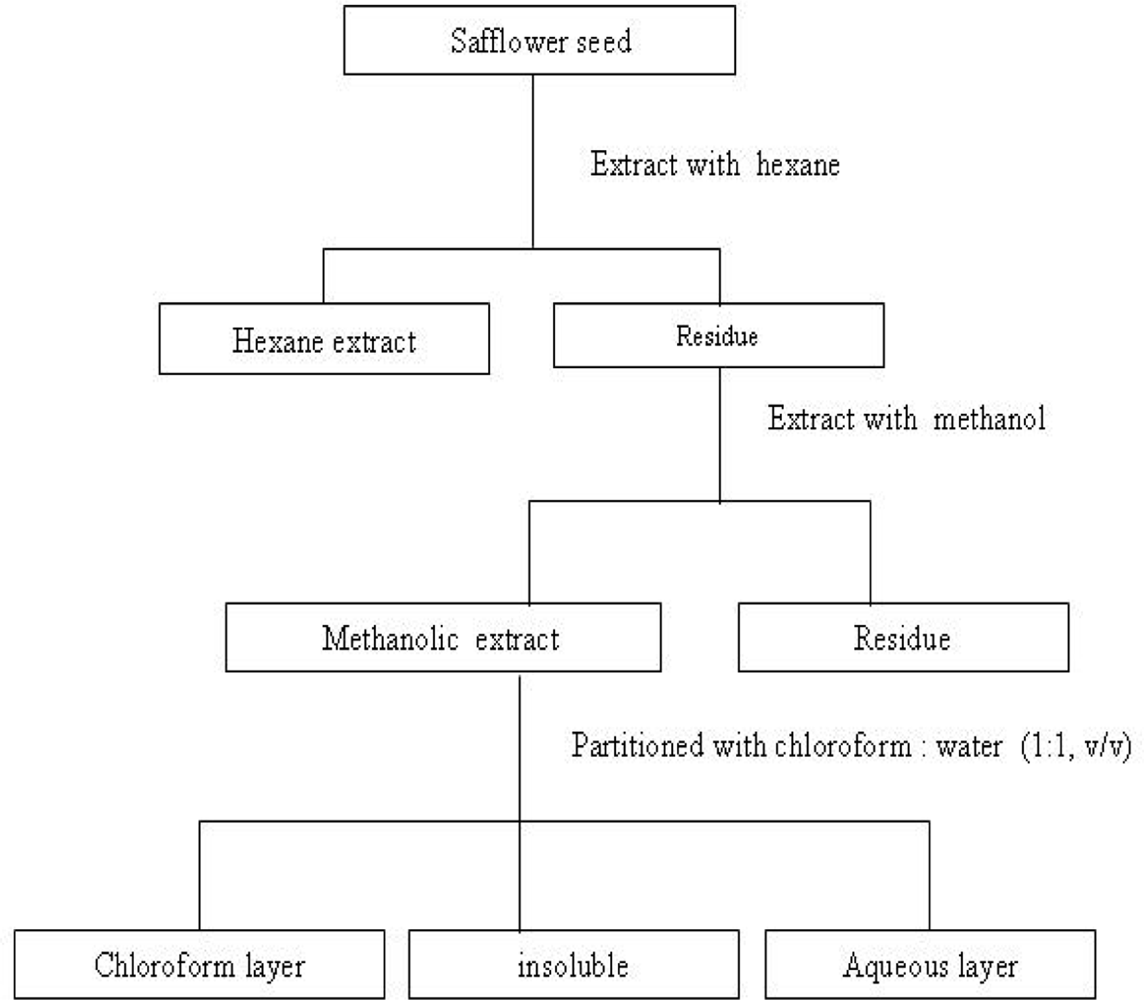
3.1. Preparation of safflower seeds extract and analysis of mineral content
3.2. Experimental animals and administration of extract
3.3. Biochemical analysis of serum
3.4. Measurement of long bone growth
3.5. Statistical analysis
4. Conclusions
Acknowledgments
References and Notes
- Rodan, GA; Martin, TJ. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar]
- Havill, LM; Hale, LG; Newman, DE; Witte, SM; Mahaney, MC. Bone ALP and OC reference standards in adult baboons (Papio hamadryas) by sex and age. J. Med. Primatol 2006, 35, 97–105. [Google Scholar]
- Szulc, P; Seeman, E; Delmas, PD. Biochemical measurements of bone turnover in children and adolescents. Osteoporosis Int 2000, 11, 281–294. [Google Scholar]
- Pedrazzoni, M; Alfano, FS; Girasole, G; Giuliani, N; Fantuzzi, M; Gatti, C; Campanini, C; Passeri, M. Clinical observations with a new specific assay for bone alkaline phosphatase: A cross-sectional study in osteoporotic and pagetic subjects and a longitudinal evaluation of the response to ovariectomy, estrogens, and biphosphonates. Calcified Tissue Int 1996, 59, 334–338. [Google Scholar]
- Ross, PD; Knowlton, W. Rapid bone loss is associated with increased levels of biochemical markers. J. Bone Miner. Res 1998, 13, 297–302. [Google Scholar]
- Ganero, P; Sornay-Rendu, E; Claustrat, B; Delmas, PD. Biochemical markers of bone turnover, endogenous hormones and the risk of fractures in postmenopausal women: the OFELY study. J. Bone Miner. Res 2000, 15, 1526–1536. [Google Scholar]
- Ross, PD; Kress, BC; Parson, RE; Wasnich, RD; Armour, KA; Mizrahi, IA. Serum bone alkaline phosphatase and calcaneus bone density predict fractures: A prospective study. Osteoporosis Int 2000, 11, 76–82. [Google Scholar]
- Hannon, R; Eastell, R. Preanalytical variability of biochemical markers of bone turnover. Osteoporosis Int 2000, 11(Supplment 6), S30–S44. [Google Scholar]
- He, J; Rosen, CJ; Adams, DJ; Kream, BE. Postnatal growth and bone mass in mice with IGF-I haploinsufficiency. Bone 2006, 38, 826–835. [Google Scholar]
- Le Roith, D. The insulin-like growth factor system. Exp. Diabet. Res 2003, 4, 205–212. [Google Scholar]
- Bikle, DD; Sakata, T; Leary, C; Elalieh, H; Ginzinger, D; Rosen, CJ; Beamer, W; Majumdar, S; Alloran, BP. Insulin-like growth factor I is required for the anabolic actions of parathyroid hormone on mouse bone. J. Bone Miner. Res 2002, 17, 1570–1578. [Google Scholar]
- Arjmandi, BH; Lucas, EA; Juma, S; Soliman, A; Stroecker, BJ; Khalil, DA; Smith, BJ; Wang, C. Dried plums prevent ovariectomy-induced bone loss in rats. J. Am. Nutr. Assoc 2001, 4, 50–56. [Google Scholar]
- Deyhim, F; Stroecker, BJ; Brusewitz, GH; Devareddy, L; Arjmandi, BH. Dried plum reverses bone loss in an osteopenic rat model of osteoporosis. Menopause 2005, 12, 755–762. [Google Scholar]
- Arjmandi, BH; Khalil, DA; Lucas, EA; Georgis, A; Stroecker, BJ; Hardin, C; Payton, ME; Wild, RA. Dried plums improve indices of bone formation in postmenopausal women. J. Womens Health Gend. Based Med 2002, 11, 61–68. [Google Scholar]
- Franklin, M; Bu, SY; Lerner, MR; Lancaster, EA; Bellmer, D; Marlow, D; Lightfoot, SA; Arjmandi, BH; Brackett, DJ; Lucas, EA; Smith, BJ. Dried plum prevents bone loss in a male osteoporosis model via IGF-I and the RANK pathway. Bone 2006, 39, 1331–1342. [Google Scholar]
- Cho, M-H; Paik, Y-S; Hahn, T-R. Enzymatic conversion of precarthamin to carthamin by a purified enzyme from the yellow petals of safflower. J. Agric. Food Chem 2000, 48, 3971–3921. [Google Scholar]
- Weiss, EA. Castor, sesame and safflower; Barnes and Noble, Inc: New York, 1971; pp. 529–744. [Google Scholar]
- Wang, G; Li, Y. Clinical application of safflower (Carthamus tinctorius) [in Chinese]. Zhejiang J. Trad. Chinese Med 1985, 20, 42–43. [Google Scholar]
- Wu, L. Yi Qi Huo Xue Prescription used for treatment of gastritis [in Chinese]. Jiangsu J. Trad. Chinese Med 1992, 13, 35–37. [Google Scholar]
- Dong, J. Huo Xue Hua Yu decoction was used for treatment on ankyloenteron of 50 cases [in Chinese]. J. Comb. Chinese Trad. West. Med 1988, 8, 237. [Google Scholar]
- Wang, Y; Wang, L. A clinical observation on the effect of rhematic prescription for treatment of rheumatoid arthritis of 50 cases [in Chinese]. Jiangsu J. Trad. Chinese Med 1990, 11, 1–3. [Google Scholar]
- Zhu, P. Hu Shen Hua Yu Decoction was used for treatment on renal function imperfection of 61 cases [in Chinese]. J. Comb. Trad. Chinese West. Med 1991, 11, 750–752. [Google Scholar]
- Huh, JS; Kang, JH; Yoo, YJ; Kim, CS; Cho, KS; Choi, SH. The effect of safflower seed fraction extract on human periodontal ligament fibroblast and MC3T3-E1 cell in vitro [in Korean]. J. Kor. Acad. Periodontol 2001, 31, 833–846. [Google Scholar]
- Seo, J-J; Kim, T; Pi, S-H; Yun, G-Y; You, H-K; Shin, H-S. Effects of safflower seed extracts and bovine bone on regeneration of bone defects in mongrel dogs [in Korean]. J. Kor. Acad. Periodontol 2000, 30, 553–569. [Google Scholar]
- Lee, S-J; Choi, H-C; Sun, K-J; Song, J-B; Pi, S-H; You, H-K; Shin, H-S. A study of safflower extracts on bone formation in vitro [in Korean]. J. Kor. Acad. Periodontol 2005, 30, 461–474. [Google Scholar]
- You, K-T; Choi, K-S; Yun, G-Y; Kim, E-C; You, H-K; Shin, H-S. Healing after implantation of bone substitutes and safflower seeds feeding in rat calvarial defects [in Korean]. J. Kor. Acad. Periodontol 2000, 30, 91–104. [Google Scholar]
- Kim, HJ; Bae, YC; Park, RW; Choi, SW; Cho, SH; Choi, YS; Lee, WJ. Bone-protecting effect of safflower seeds in ovariectomized rats. Calcified Tissue Int 2002, 71, 88–94. [Google Scholar]
- Hong, H-T; Kim, H-J; Lee, T-K; Kim, D-W; Kim, H-M; Choo, Y-K; Park, Y-G; Lee, Y-C; Kim, C-H. Inhibitory effect of a Korean traditional medicine, Honghwain-Jahage (water extracts of Carthamus tinctorius L. seed and Homonis placenta) on interleukin-1-mediated bone resorption. J. Ethnopharm 2002, 79, 143–148. [Google Scholar]
- Kim, K-W; Suh, S-J; Lee, T-K; Ha, K-T; Kim, J-K; Kim, K-H; Kim, D-I; Jeon, J-H; Moon, T-C; Kim, C-H. Effect of safflower seeds supplementation on stimulation of the proliferation, differentiation and mineralization of osteoblastic MC3T3-E1 cells. J. Ethnopharm 2008, 115, 42–49. [Google Scholar]
- Lee, JY; Chang, EJ; Kim, HJ; Park, JH; Choi, SW. Antioxidant flavonoids from leaves of Carthamus tinctorius. Arch. Pharm. Res 2002, 25, 313–319. [Google Scholar]
- Kim, EO; Oh, JH; Lee, SK; Lee, JY; Choi, SW. Antioxidant properties and quantification of phenolic compounds from safflower (Carthamus tinctorius L.) seeds. Food Sci. Biotechnol 2007, 16, 71–77. [Google Scholar]
- Dixon, RA. Phytoestrogens. Annu. Rev. Plant Biol 2004, 55, 225–261. [Google Scholar]
- Horvath, AL. Solubility of structurally complicated materials: II. Bone. J. Phys. Chem. Ref. Data 2006, 4, 1653–1668. [Google Scholar]
- Koo, WWK; Walters, JC; Esterlitz, J; Levine, RJ; Bush, AJ; Sibai, B. Maternal calcium supplementation and fetal bone mineralization. Obstet. Gynecol 1999, 94, 577–582. [Google Scholar]
- Lemann, J; Pleuss, JA. Potassium causes calcium retention in healthy adults. J. Nutr 1993, 123, 1623–1626. [Google Scholar]
- Macdonald, HM; New, SA; Fraser, WD; Campbell, MK; Reid, DM. Low dietary potassium intakes and high dietary estimates of net endogenous acid production are associated with low bone mineral density in premenopausal women and increased markers of bone resorption in postmenopausal women. Am. J. Clin. Nutr 2005, 81, 923–933. [Google Scholar]
- Koyama, Y; Rittling, SR; Tsuji, K; Hino, K; Salincarnboriboon, R; Yano, T; Taketani, Y; Nifuji, A; Denhardt, D; Noda, M. Osteopontin deficiency suppresses high phosphate load-induced bone loss via specific modulation of osteoclasts. Endocrinology 2006, 147, 3040–3049. [Google Scholar]
- Polak-Jonkisz, D; Zwolinska, D. Osteocalcin as a biochemical marker of bone turnover. Nephrology 1998, 4, 339–346. [Google Scholar]
- Glover, SJ; Garnero, P; Naylor, K; Rogers, A; Eastell, R. Establishing a reference range for bone turnover markers in young, healthy women. Bone 2008, 42, 623–630. [Google Scholar]
- Aizawa, T; Kokubun, S; Tanaka, Y. Apoptosis and proliferation of growth plate chondrocytes in rabbits. JBJS 1997, 79, 483–486. [Google Scholar]
- Roach, HI; Mehta, G; Oreffo, RO; Clarke, NM; Cooper, C. Temporal analysis of rat growth plates: cessation of growth with age despite presence of a physis. J. Histochem. Cytochem 2003, 51, 373–383. [Google Scholar]
- Kim, DH; Lee, JH; Ahn, EM; Lee, YH; Baek, NI; Lim, IH. Phenolic glycosides isolated from safflower (Carthamus tinctorius L.) seeds increase the alkaline phosphatase (ALP) activity of human osteoblast-like cells. Food Sci. Biotechnol 2006, 15, 781–785. [Google Scholar]
- Aubin, JE; Liu, F; Malaval, L; Gupta, AK. Osteoblast and chondroblast differentiation. Bone 1995, 17, 77S–83S. [Google Scholar]
- Lin, W-w; Oberbauer, AM. Spatiotemporal expression of alternatively spiced IGF-I mRNA in the rat costochondral growth plate. J. Endocrinol 1999, 160, 461–467. [Google Scholar]
- Schwander, SC; Hauri, C; Zapf, J; Froescj, ER. Synthesis and secretion of insulin-like growth factor and its binding protein by the perfused rat liver: Dependence on growth hormone status. Endocrinology 1983, 113, 297–305. [Google Scholar]
- Ezzat, S; Melmed, S; Endres, D; Eyre, DR; Singer, FR. Biochemical assessment of bone formation and resorption in acromegaly. J. Clin. Endocrinol. Metab 1993, 76, 1452–1457. [Google Scholar]
- Le Roith, D; Bondy, C; Yakar, S; Liu, JL; Butler, A. The somatomedin hypothesis. Endocrine Rev 2001, 22, 53–74. [Google Scholar]
- Yakar, S; Rosen, CJ. From mouse to man: Redefining the role of insulin-like growth factor-I in the acquisition of bone mass. Exp. Biol. Med 2003, 228, 245–252. [Google Scholar]
- Baxter, RC. Circulating levels and molecular distribution of the acid-labile (α) subunit of the high molecular weight insulin-like growth factor-binding protein complex. J. Clin. Endocrinol. Metab 1990, 70, 1347–1353. [Google Scholar]
- Daughaday, WH. Free insulin-like growth factor (IGF) in disorders of IGF binding protein 3 complex formation. J. Clin. Endocrinol 2004, 89, 3–5. [Google Scholar]
- Bouxsein, ML; Rosen, CJ; Turner, CH; Ackert, CL; Shultz, KL; Donahue, LR; Churchill, G; Adamo, ML; Powell, DR; Turner, RT; Müller, R; Beamer, WG. Generation of a new congenic mouse strain to test the relationships among serum insulin-like growth factor I, bone mineral density, and skeletal morphology in vivo. J. Bone Miner. Res 2002, 17, 570–579. [Google Scholar]
- Rosen, CJ. Insulin-like growth factor and bone mineral density: experience from animal models and human observational studies. Best Practice & Research. Clin. Endocrinol. Metab 2004, 18, 423–435. [Google Scholar]
- Song, H-R; Ra, D-K; Kim, J-S; Jung, T-S; Kim, Y-H; Kang, H-J; Kang, C-B; Yeon, S-C; Kim, EH; Lee, H-J; Shin, G-W; Park, M-R; Kim, G-S. Effects of safflower seed on new bone formation. J. Vet. Clinic 2002, 19, 66–72. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements | Contents (μg/g)
| |
|---|---|---|
| W1) | M2) | |
| Al | 0.175 | 0.019 |
| Ca | 0.075 | 3.752 |
| Cu | 0.055 | 0.015 |
| Fe | 0.100 | 0.170 |
| K | 2.306 | 1.313 |
| Mg | 0.474 | 0.023 |
| Na | 0.066 | 0.177 |
| P | 1.043 | 1.161 |
| Sr | 0.002 | 0.002 |
| Zn | 0.070 | 0.042 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, Y.S.; Choi, C.W.; Kim, J.J.; Ganapathi, A.; Udayakumar, R.; Kim, S.C. Determination of Mineral Content in Methanolic Safflower (Carthamus tinctorius L.) Seed Extract and Its Effect on Osteoblast Markers. Int. J. Mol. Sci. 2009, 10, 292-305. https://doi.org/10.3390/ijms10010292
Lee YS, Choi CW, Kim JJ, Ganapathi A, Udayakumar R, Kim SC. Determination of Mineral Content in Methanolic Safflower (Carthamus tinctorius L.) Seed Extract and Its Effect on Osteoblast Markers. International Journal of Molecular Sciences. 2009; 10(1):292-305. https://doi.org/10.3390/ijms10010292
Chicago/Turabian StyleLee, Young Seok, Chang Won Choi, Jae Jin Kim, Andy Ganapathi, Rajangam Udayakumar, and Sei Chang Kim. 2009. "Determination of Mineral Content in Methanolic Safflower (Carthamus tinctorius L.) Seed Extract and Its Effect on Osteoblast Markers" International Journal of Molecular Sciences 10, no. 1: 292-305. https://doi.org/10.3390/ijms10010292
APA StyleLee, Y. S., Choi, C. W., Kim, J. J., Ganapathi, A., Udayakumar, R., & Kim, S. C. (2009). Determination of Mineral Content in Methanolic Safflower (Carthamus tinctorius L.) Seed Extract and Its Effect on Osteoblast Markers. International Journal of Molecular Sciences, 10(1), 292-305. https://doi.org/10.3390/ijms10010292