
Comparison of Polyphenolic Content and Bioactivities Between Extracts from the Living Plants and Beach Deposits of the Submerged Brackish Water Angiosperm Ruppia maritima


 , and
, and 
Abstract
1. Introduction
2. Results and Discussion
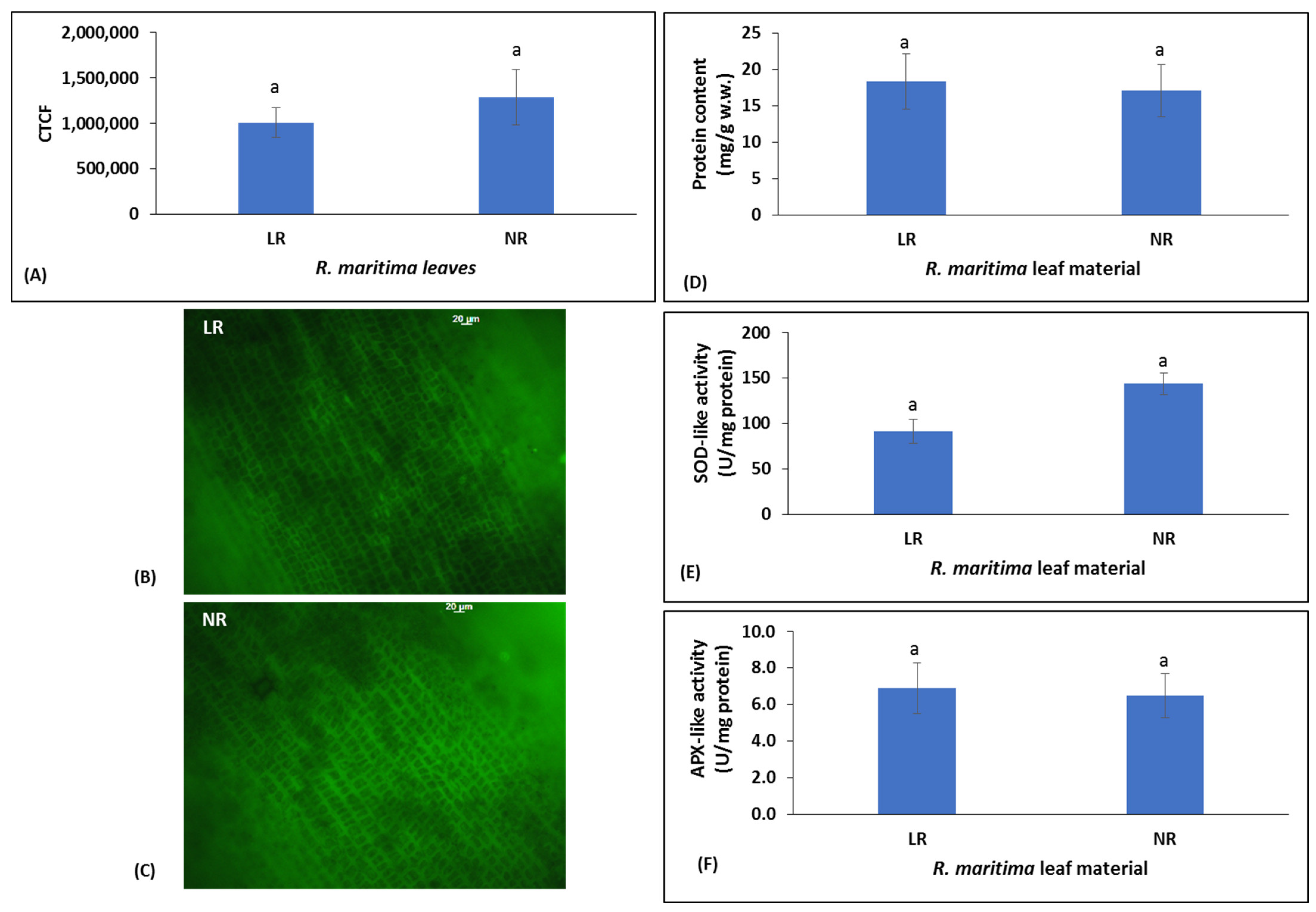
2.1. Oxidative Stress, Antioxidant Enzyme Activity, and Protein Content of Ruppia maritima Living Plant (LR) and Beach Deposit (NR) Samples
2.2. Determination of (Poly)phenolic Composition of LR and NR Extracts from Ruppia maritima
2.3. Antioxidant Activity of LR and NR Extracts from Ruppia maritima
2.4. Inhibitory Activity of LR and NR Extracts from Ruppia maritima Against Cancer Cell Growth
2.5. Case Study for the Quantification of the Bioactive Extract Produced by Ruppia maritima Beach Deposits in the Collection Area
3. Materials and Methods
3.1. Sampling of Living Plants and Beach Deposits of Ruppia maritima
3.2. Imaging of Hydrogen Peroxide Production in Leaves from Living Plants (LR) and Beach Deposits (NR) of Ruppia maritima
3.3. Assessment of the Activity of the Antioxidant Enzymes SOD-like and APX-like and Protein Content in Leaves from Living Plants (LR) and Beach Deposits (NR) of Ruppia maritima
3.4. Preparation of Ruppia maritima Extracts
3.5. Assessment of Phenolic Coompounds in Ruppia maritima Extracts
3.6. Free Radical Scavenging Activity of Ruppia maritima Extracts
3.7. Reducing Power (RP) Activity of Ruppia maritima Extracts
3.8. Cell Culture Conditions
3.9. XTT Assay for Assessing the Inhibitory Activity of Ruppia maritima Extracts Against Cancer Cell Proliferation
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2,2-Azino-bis(3-ethylbenzthiazoline-6-sulfonic acid | ABTS•+ |
| 2,2-Diphenyl-picrylhydrazyl | DPPH |
| Ascorbic peroxidase | APX |
| Corrected total cell fluorescence | CTCF |
| Ethylenediaminetetraacetic acid | EDTA |
| Hydrogen peroxide | H2O2 |
| Hydroxyl radical | OH• |
| Polyvinylpolypyrrolidone | PVPP |
| Reactive oxygen species | ROS |
| Reducing power | RP |
| Ruppia maritima beach deposits (or necromass) | NR |
| Ruppia maritima living plants | LR |
| Selectivity index | SI |
| Superoxide dismutase | SOD |
| Superoxide radical | O2•− |
References
- Davis, W.P.; Davis, M.R.; Flemer, D.A. Observations on the Regrowth of Subaquatic Vegetation Following Transplantation: A Potential Method to Assess Environmental Health of Coastal Habitats. In Seagrasses: Monitoring, Ecology, Physiology and Management; Bortone, S.A., Ed.; CRC Press: Boca Raton, FL, USA, 2000; p. 318. [Google Scholar]
- den Hartog, C.; Kuo, J. Taxonomy and Biogeography of Seagrasses. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–23. [Google Scholar]
- Agulles, M.; Marbà, N.; Duarte, C.M.; Jordà, G. Mediterranean Seagrasses Provide Essential Coastal Protection under Climate Change. Sci. Rep. 2024, 14, 30269. [Google Scholar] [CrossRef]
- Malea, P.; Kevrekidis, T.; Mogias, A. Annual versus Perennial Growth Cycle in Ruppia maritima L.: Temporal Variation in Population Characteristics in Mediterranean Lagoons (Monolimni and Drana Lagoons, Northern Aegean Sea). Bot. Mar. 2004, 47, 357–366. [Google Scholar] [CrossRef]
- Ito, Y.; Ohi-Toma, T.; Murata, J.; Tanaka, N. Hybridization and Polyploidy of an Aquatic Plant, Ruppia (Ruppiaceae), Inferred from Plastid and Nuclear DNA Phylogenies. Am. J. Bot. 2010, 97, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Agostini, S.; Capiomont, A.; Marchand, B.; Pergent, G. Distribution and Estimation of Basal Area Coverage of Subtidal Seagrass Meadows in a Mediterranean Coastal Lagoon. Estuar. Coast. Shelf Sci. 2003, 56, 1021–1028. [Google Scholar] [CrossRef]
- Nonnis Marzano, C.; Scalera Liaci, L.; Fianchini, A.; Gravina, F.; Mercurio, M.; Corriero, G. Distribution, Persistence and Change in the Macrobenthos of the Lagoon of Lesina (Apulia, Southern Adriatic Sea). Oceanol. Acta 2003, 26, 57–66. [Google Scholar] [CrossRef]
- Malea, P.; Kevrekidis, T.; Mogias, A.; Adamakis, I.-D.S. Kinetics of Cadmium Accumulation and Occurrence of Dead Cells in Leaves of the Submerged Angiosperm Ruppia maritima. Bot. Mar. 2014, 57, 111–122. [Google Scholar] [CrossRef]
- Christia, C.; Tziortzis, I.; Fyttis, G.; Kashta, L.; Papastergiadou, E.A. Survey of the Benthic Aquatic Flora in Transitional Water Systems of Greece and Cyprus (Mediterranean Sea). Bot. Mar. 2011, 54, 169–178. [Google Scholar] [CrossRef]
- Ragupathi Raja Kannan, R.; Arumugam, R.; Thangaradjou, T.; Anantharaman, P. Phytochemical Constituents, Antioxidant Properties and p-Coumaric Acid Analysis in Some Seagrasses. Food Res. Int. 2013, 54, 1229–1236. [Google Scholar] [CrossRef]
- Kim, D.H.; Mahomoodally, M.F.; Sadeer, N.B.; Seok, P.G.; Zengin, G.; Palaniveloo, K.; Khalil, A.A.; Rauf, A.; Rengasamy, K.R. Nutritional and Bioactive Potential of Seagrasses: A Review. S. Afr. J. Bot. 2021, 137, 216–227. [Google Scholar] [CrossRef]
- Zamani, N.P.; Rahman, L.; Rosada, R.L.; Tirtama, W. Overview of Bioactivity Studies on Marine Natural Products. IOP Conf. Ser. Earth Environ. Sci. 2021, 944, 012029. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Fakhfakh, J.; Krichen, F.; Jribi, I.; Chiarore, A.; Patti, F.P.; Blecker, C.; Allouche, N.; Belghith, H.; Belghith, K. Structural Characterization and Functional Properties of Antihypertensive Cymodocea nodosa Sulfated Polysaccharide. Carbohydr. Polym. 2016, 151, 511–522. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Salah, H.B.; Saidi, S.A.; Allouche, N.; Belghith, H.; Belghith, K. Evaluation of Nutritional Value, Characteristics, Functional Properties of Cymodocea nodosa and Its Benefits on Health Diseases. Lipids Health Dis. 2017, 16, 238. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Taberner, M.D.M.; Mir-Rossello, P.M.; Gil, L.; Sureda, A.; Capó, X. Potential Use of Marine Plants as a Source of Bioactive Compounds. Molecules 2025, 30, 485. [Google Scholar] [CrossRef] [PubMed]
- Kevrekidou, A.; Assimopoulou, A.N.; Trachana, V.; Stagos, D.; Malea, P. Antioxidant Activity, Inhibition of Intestinal Cancer Cell Growth and Polyphenolic Compounds of the Seagrass Posidonia oceanica’s Extracts from Living Plants and Beach Casts. Mar. Drugs 2024, 22, 130. [Google Scholar] [CrossRef]
- Hasle Enerstvedt, K.; Lundberg, A.; Jordheim, M. Characterization of Polyphenolic Content in the Aquatic Plants Ruppia cirrhosa and Ruppia maritima—A Source of Nutritional Natural Products. Molecules 2017, 23, 16. [Google Scholar] [CrossRef]
- Malea, P.; Boubonari, T.; Kevrekidis, T. Iron, Zinc, Copper, Lead and Cadmium Contents in Ruppia maritima from a Mediterranean Coastal Lagoon: Monthly Variation and Distribution in Different Plant Fractions. Bot. Mar. 2008, 51, 320–330. [Google Scholar] [CrossRef]
- Malea, P.; Mylona, Z.; Kevrekidis, T. Trace Elements in the Seagrass Posidonia oceanica: Compartmentation and Relationships with Seawater and Sediment Concentrations. Sci. Total Environ. 2019, 686, 63–74. [Google Scholar] [CrossRef]
- Papathanasiou, V.; Kariofillidou, G.; Malea, P.; Orfanidis, S. Effects of Air Exposure on Desiccation and Photosynthetic Performance of Cymodocea nodosa with and without Epiphytes and Ulva rigida in Comparison, under Laboratory Conditions. Mar. Environ. Res. 2020, 158, 104948. [Google Scholar] [CrossRef]
- Malea, P.; Charitonidou, K.; Sperdouli, I.; Mylona, Z.; Moustakas, M. Zinc Uptake, Photosynthetic Efficiency and Oxidative Stress in the Seagrass Cymodocea nodosa Exposed to ZnO Nanoparticles. Materials 2019, 12, 2101. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Malea, P.; Sperdouli, I.; Panteris, E.; Kokkinidi, D.; Moustakas, M. Evaluation of the Spatiotemporal Effects of Bisphenol A on the Leaves of the Seagrass Cymodocea nodosa. J. Hazard. Mater. 2021, 404, 124001. [Google Scholar] [CrossRef]
- Liu, J.; Tang, X.; Wang, Y.; Zang, Y.; Zhou, B. A Zostera Marina Manganese Superoxide Dismutase Gene Involved in the Responses to Temperature Stress. Gene 2016, 575, 718–724. [Google Scholar] [CrossRef]
- Malea, P.; Emmanouilidis, A.; Kevrekidis, D.P.; Moustakas, M. Copper Uptake Kinetics and Toxicological Effects of Ionic Cu and CuO Nanoparticles on the Seaweed Ulva rigida. Environ Sci. Pollut. Res. 2022, 29, 57523–57542. [Google Scholar] [CrossRef] [PubMed]
- Malea, P.; Kokkinidi, D.; Kevrekidou, A.; Adamakis, I.-D.S. The Enzymatic and Non-Enzymatic Antioxidant System Response of the Seagrass Cymodocea nodosa to Bisphenol-A Toxicity. Int. J. Mol. Sci. 2022, 23, 1348. [Google Scholar] [CrossRef]
- Malea, P.; Dermentzis, M.; Patronia, M.-M.; Kevrekidis, D.P.; Kevrekidou, A.; Siopi, V. Mechanism of Up-Regulated H2O2 BPA-Derived Production and Production of (Poly)Phenols by Two Seaweeds of the Genus Ulva. Environ Sci. Pollut. Res. 2024, 31, 52017–52031. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, P.; Umamaheswari, A.; Banupriya, R.; Elumalai, S. Phytochemical Screening and in Vitro Anticancer Activity of Ethyl Acetate Fraction of Seagrass Halodule Uninervis from Mandapam Coastal Region, Rameswaram, Gulf of Mannar, India. Int. J. Pharm. Sci. Drug Res. 2021, 13, 677–684. [Google Scholar] [CrossRef]
- Mohamed, S.I.A.; Elsayed, G.H.; El Shaffai, A.; Yahya, S.M.M.; Mettwally, W.S.A. In-Vitro Study of Cytotoxic and Apoptotic Potential of Thalassia hemprichii (Ehren.) Asch. And Enhalus acoroides (L.f.) Royle against Human Breast Cancer Cell Line (MCF-7) with Correlation to Their Chemical Profile. BMC Complement. Med. Ther. 2024, 24, 244. [Google Scholar] [CrossRef]
- Rubio-Portillo, E.; Martin-Cuadrado, A.-B.; Ramos-Esplá, A.Á.; Antón, J. Metagenomics Unveils Posidonia oceanica “Banquettes” as a Potential Source of Novel Bioactive Compounds and Carbohydrate Active Enzymes (CAZymes). mSystems 2021, 6, e0086621. [Google Scholar] [CrossRef]
- Vasarri, M.; De Biasi, A.M.; Barletta, E.; Pretti, C.; Degl’Innocenti, D. An Overview of New Insights into the Benefits of the Seagrass Posidonia oceanica for Human Health. Mar. Drugs 2021, 19, 476. [Google Scholar] [CrossRef]
- Harrison, P.G. Detrital Processing in Seagrass Systems: A Review of Factors Affecting Decay Rates, Remineralization and Detritivory. Aquat. Bot. 1989, 35, 263–288. [Google Scholar] [CrossRef]
- Achamlale, S.; Rezzonico, B.; Grignon-Dubois, M. Evaluation of Zostera detritus as a Potential New Source of Zosteric Acid. J. Appl. Phycol. 2009, 21, 347–352. [Google Scholar] [CrossRef]
- United Nations Environment Programme-World Conservation Monitoring Centre, UNEP-WCMC. Available online: https://unepwcmc.org/en/news/new-research-shows-which-mediterranean-coastal-communities-must-plan-now-for-seagrass-loss (accessed on 10 March 2025).
- Apostoloumi, C.; Malea, P.; Kevrekidis, T. Principles and Concepts about Seagrasses: Towards a Sustainable Future for Seagrass Ecosystems. Mar. Pollut. Bull. 2021, 173, 112936. [Google Scholar] [CrossRef]
- de los Santos, C.B.; Scott, A.; Arias-Ortiz, A.; Jones, B.; Kennedy, H.; Mazarrasa, I.; McKenzie, L.; Nordlund, L.M.; de la Torre-Castro, M.; Unsworth, R.K.F.; et al. Seagrass Ecosystem Services: Assessment and Scale of Benefits. In Out of the Blue: The Value of Seagrasses to the Environment and to People; UNEP, Ed.; UNEP: Nairobi, Kenya, 2020; pp. 48–55. [Google Scholar]
- Tutar, O.; Marín-Guirao, L.; Ruiz, J.M.; Procaccini, G. Antioxidant Response to Heat Stress in Seagrasses. A Gene Expression Study. Mar. Environ. Res. 2017, 132, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Bharathi, N.P.; Jayalakshmi, M.; Amudha, P.; Vanitha, V. Phytochemical screening and in vitro antioxidant activity of the seagrass Cymodocea serrulata. Indian J. Geo-Mar. Sci. 2019, 48, 1216–1221. [Google Scholar]
- Messina, C.M.; Arena, R.; Manuguerra, S.; Pericot, Y.; Curcuraci, E.; Kerninon, F.; Renda, G.; Hellio, C.; Santulli, A. Antioxidant Bioactivity of Extracts from Beach Cast Leaves of Posidonia oceanica (L.) Delile. Mar. Drugs 2021, 19, 560. [Google Scholar] [CrossRef] [PubMed]
- Krastanov, A.; Alexiera, Z.; Yernendzhiev, H. Microbial degradation of phenol and phenolic derivatives. Eng. Life Sci. 2013, 13, 76–87. [Google Scholar] [CrossRef]
- Agarry, S.S.A.; Durojaiye, A.O.; Solomon, B. Microbial degradation of phenols: A review. Int. J. Environ. Pollut. 2008, 32, 12–28. [Google Scholar] [CrossRef]
- Mejiaa, S.Y.; Rotini, A.; Lacasell, F.; Bookman, R.; Thaller, M.C.; Shem-Tov, R.; Winters, G.; Migliore, L. Assessing the ecological status of seagrasses using morphology, biochemical descriptors and microbial community analyses.A study in Halophila stipulacea (Forsk.) Aschers meadows in thenorthern Red Sea. Ecol. Indic. 2016, 60, 1150–1163. [Google Scholar] [CrossRef]
- Soto-Hernandez, M.; Palma-Tenango, M.; Garcia-Mateos, R. (Eds.) Phenolic Compounds—Natural Sources, Importance and Applications; InTech: Houston, TX, USA, 2017; p. 456. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S. Microbes and Enzymes in Soil Health and Bioremediation; Kumar, A.N., Ed.; Microorganisms for Sustainability; Springer: Cham, Switzerland, 2019; Volume 16. [Google Scholar]
- Ameen, H.M.; Jayadev, A.; Prasad, G.; Nair, D.I. Seagrass Meadows: Prospective Candidates for Bioactive Molecules. Molecules 2024, 29, 4596. [Google Scholar] [CrossRef]
- Adams, J.; Bate, G. The tolerance to desiccation of the submerged macrophytes Ruppia cirrhosa (Petagna) Grande and Zostera capensis Setchell. J. Experim. Mar. Biol. Ecol. 1994, 183, 53–62. [Google Scholar] [CrossRef]
- Shafer, D.J.; Sherman, T.D.; Wyllie-Echeverria, S. Do desiccation tolerances control the vertical distribution of intertidal seagrasses? Aquat. Bot. 2007, 87, 161–166. [Google Scholar] [CrossRef]
- Lee, J.; Scagel, C.F. Chicoric Acid: Chemistry, Distribution, and Production. Front. Chem. 2013, 1, 40. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, M.; Kalavagunta, P.K.; Li, J.; He, Q.; Zhang, Y.; Ahmad, O.; Yin, H.; Wang, T.; Shang, J. Protective Effects of Chicoric Acid on H2O2-Induced Oxidative Injury in Hepatocytes and Larval Zebrafish Models. Biomed. Pharmacother. 2018, 104, 679–685. [Google Scholar] [CrossRef]
- Li, Z.; Feng, H.; Han, L.; Ding, L.; Shen, B.; Tian, Y.; Zhao, L.; Jin, M.; Wang, Q.; Qin, H.; et al. Chicoric Acid Ameliorate Inflammation and Oxidative Stress in Lipopolysaccharide and D -galactosamine Induced Acute Liver Injury. J. Cell Mol. Med. 2020, 24, 3022–3033. [Google Scholar] [CrossRef]
- Nobela, O.; Renslow, R.S.; Thomas, D.G.; Colby, S.M.; Sitha, S.; Njobeh, P.B.; du Preez, L.; Tugizimana, F.; Madala, N.E. Efficient Discrimination of Natural Stereoisomers of Chicoric Acid, an HIV-1 Integrase Inhibitor. J. Photochem. Photobiol. B Biol. 2018, 189, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Sun, Q.; Gao, R.; Park, Y. AAK-2 and SKN-1 Are Involved in Chicoric-Acid-Induced Lifespan Extension in caenorhabditis Elegans. J. Agric. Food Chem. 2019, 67, 9178–9186. [Google Scholar] [CrossRef]
- Kim, M.; Yoo, G.; Randy, A.; Kim, H.S.; Nho, C.W. Chicoric Acid Attenuate a Nonalcoholic Steatohepatitis by Inhibiting Key Regulators of Lipid Metabolism, Fibrosis, Oxidation, and Inflammation in Mice with Methionine and Choline Deficiency. Mol. Nutr. Food Res. 2017, 61, 1600632. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wu, C.; Zhang, T.; Shi, L.; Li, J.; Liang, H.; Lv, X.; Jing, F.; Qin, L.; Zhao, T.; et al. Chicoric Acid: Natural Occurrence, Chemical Synthesis, Biosynthesis, and Their Bioactive Effects. Front Chem. 2022, 10, 888673. [Google Scholar] [CrossRef]
- DellaGreca, M.; Fiorentino, A.; Isidori, M.; Monaco, P.; Zarrelli, A. Antialgal Ent-Labdane Diterpenes from Ruppia maritima. Phytochemistry 2000, 55, 909–913. [Google Scholar] [CrossRef]
- Haznedaroglu, M.Z.; Zeybek, U. HPLC Determination of Chicoric Acid in Leaves of Posidonia oceanica. Pharm. Biol. 2007, 45, 745–748. [Google Scholar] [CrossRef]
- Grignon-Dubois, M.; Rezzonico, B. Phenolic Fingerprint of the Seagrass Posidonia oceanica from Four Locations in the Mediterranean Sea: First Evidence for the Large Predominance of Chicoric Acid. Bot. Mar. 2015, 58, 379–391. [Google Scholar] [CrossRef]
- Grignon-Dubois, M.; Rezzonico, B. The Economic Potential of Beach-Cast Seagrass—Cymodocea nodosa: A Promising Renewable Source of Chicoric Acid. Bot. Mar. 2013, 56, 303–311. [Google Scholar] [CrossRef]
- Papenbrock, J. Highlights in Seagrasses’ Phylogeny, Physiology, and Metabolism: What Makes Them Special? ISRN Bot. 2012, 2012, 103892. [Google Scholar] [CrossRef]
- Zeng, W.; Long, X.; Liu, P.; Xie, X. The Interplay of Oncogenic Signaling, Oxidative Stress and Ferroptosis in Cancer. Int. J. Cancer 2023, 153, 918–931. [Google Scholar] [CrossRef]
- Tsukatani, T.; Ide, S.; Ono, M.; Matsumoto, K. New tetrazolium method for phosphatase assay using ascorbic acid 2-phosphate as a substrate. Talanta 2007, 73, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 2. Hydrogen Atom Transfer (HAT)-Based, Mixed-Mode (Electron Transfer (ET)/HAT), and Lipid Peroxidation Assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef]
- Peng, Y.; Sun, Q.; Park, Y. The Bioactive Effects of Chicoric Acid as a Functional Food Ingredient. J. Med. Food 2019, 22, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Diao, Z.; Li, J.; Ren, B.; Zhu, D.; Liu, Q.; Liu, Z.; Liu, X. Chicoric Acid Supplementation Ameliorates Cognitive Impairment Induced by Oxidative Stress via Promotion of Antioxidant Defense System. RSC Adv. 2017, 7, 36149–36162. [Google Scholar] [CrossRef]
- Thygesen, L.; Thulin, J.; Mortensen, A.; Skibsted, L.H.; Molgaard, P. Antioxidant Activity of Cichoric Acid and Alkamides from Echinacea Purpurea, Alone and in Combination. Food Chem. 2007, 101, 74–81. [Google Scholar] [CrossRef]
- Nantitanon, W. Comparison of Antioxidant Activity of Compounds Isolated from Guava Leaves and a Stability Study of the Most Active Compound. Drug. Discov. Ther. 2012, 6, 38–43. [Google Scholar] [CrossRef]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic Studies on the Antioxidant Activity of p-Coumaric Acid. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Sheng, K.; Li, Y.; Wang, Z.; Hang, K.; Ye, Z. p-Coumaric Acid Suppresses Reactive Oxygen Species-induced Senescence in Nucleus Pulposus Cells. Exp. Ther. Med. 2021, 23, 183. [Google Scholar] [CrossRef] [PubMed]
- Nivetha, S.; Asha, K.R.T.; Srinivasan, S.; Murali, R.; Kanagalakshmi, A. p-Coumaric Acid Pronounced Protective Effect against Potassium Bromate-induced Hepatic Damage in Swiss Albino Mice. Cell Biochem. Funct. 2024, 42, e4076. [Google Scholar] [CrossRef] [PubMed]
- Khelifi, I.; Hayouni, E.A.; Cazaux, S.; Ksouri, R.; Bouajila, J. Evaluation of in Vitro Biological Activities: Antioxidant; Anti-Inflammatory; Anti-Cholinesterase; Anti- Xanthine Oxidase, Anti-Superoxyde Dismutase, Anti-Glucosidase and Cytotoxic of 19 Bioflavonoids. Cell Mol. Biol. 2020, 66, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Nićiforović, N.; Polak, T.; Makuc, D.; Poklar Ulrih, N.; Abramovič, H. A Kinetic Approach in the Evaluation of Radical-Scavenging Efficiency of Sinapic Acid and Its Derivatives. Molecules 2017, 22, 375. [Google Scholar] [CrossRef]
- Akdemir, F.N.E.; Güler, M.C.; Eraslan, E.; Tanyeli, A.; Yildirim, S. Assessment of Sinapic Acid’s Protective Effects against Ethanol-Induced Gastric Ulcers in Rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 398, 7133–7145. [Google Scholar] [CrossRef]
- Ortasoz, A.M.; Ozdemir, E.; Taskıran, A.S.; Ozturk, A. Sinapic Acid Alleviates Glutamate-Induced Excitotoxicity by Inhibiting Neuroinflammation and Endoplasmic Reticulum Stress Pathway in C6 Glioma Cells. Toxicol. Vitr. 2025, 103, 105977. [Google Scholar] [CrossRef]
- Akarsu, S.A.; İleritürk, M.; Küçükler, S.; Akaras, N.; Gür, C.; Kandemir, F.M. Ameliorative Effects of Sinapic Acid against Vancomycin-Induced Testicular Oxidative Damage, Apoptosis, Inflammation, Testicular Histopathologic Disorders and Decreased Epididymal Sperm Quality. Reprod. Toxicol. 2024, 129, 108666. [Google Scholar] [CrossRef]
- Mehmood, A.; Soliman, M.M.; Almalki, D.A.; Alotaibi, K.S.; Youssef, G.B.A.; Althobaiti, S. Ameliorative Impacts of Sinapic Acid against Mercuric Chloride-Induced Renal Toxicity: Role of Antioxidants and Inflammatory Cytokines. Toxicol. Res. 2024, 13, tfae066. [Google Scholar] [CrossRef]
- Campoccia, D.; Ravaioli, S.; Santi, S.; Mariani, V.; Santarcangelo, C.; De Filippis, A.; Montanaro, L.; Arciola, C.R.; Daglia, M. Exploring the anticancer effects of standardized extracts of poplar-type propolis: In vitro cytotoxicity toward cancer and normal cell lines. Biomed. Pharmacother. 2021, 141, 111895. [Google Scholar] [CrossRef]
- Mouna, R.; Broisat, A.; Ahmed, A.; Debiossat, M.; Boumendjel, A.; Ghezzi, C.; Kabouche, Z. Antiproliferative Activity, Cell-Cycle Arrest, Apoptotic Induction and LC-HRMS/MS Analyses of Extracts from Two Linum Species. Pharm. Biol. 2022, 60, 1491–1501. [Google Scholar] [CrossRef]
- Yoon, J.-Y.; Cho, H.-S.; Lee, J.-J.; Lee, H.-J.; Jun, S.Y.; Lee, J.-H.; Song, H.-H.; Choi, S.; Saloura, V.; Park, C.G.; et al. Novel TRAIL Sensitizer Taraxacum officinale F.H. Wigg Enhances TRAIL-Induced Apoptosis in Huh7 Cells. Mol. Carcinog. 2016, 55, 387–396. [Google Scholar] [CrossRef]
- Velusamy, P.; Muthusami, S.; Arumugam, R. In Vitro Evaluation of P-Coumaric Acid and Naringin Combination in Human Epidermoid Carcinoma Cell Line (A431). Med. Oncol. 2023, 41, 4. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Martins, J.I.; Gonzatti, M.B.; Varela, M.T.; Sousa, M.E.P.; Costa, L.V.S.; Rodrigues, E.G.; Fernandes, J.P.S.; Keller, A.C. Esterification of P-Coumaric Acid Improves the Control over Melanoma Cell Growth. Biomedicines 2023, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Radziejewska, I.; Supruniuk, K.; Tomczyk, M.; Izdebska, W.; Borzym-Kluczyk, M.; Bielawska, A.; Bielawski, K.; Galicka, A. P-Coumaric Acid, Kaempferol, Astragalin and Tiliroside Influence the Expression of Glycoforms in AGS Gastric Cancer Cells. Int. J. Mol. Sci. 2022, 23, 8602. [Google Scholar] [CrossRef]
- Cui, K.; Wu, H.; Fan, J.; Zhang, L.; Li, H.; Guo, H.; Yang, R.; Li, Z. The Mixture of Ferulic Acid and P-Coumaric Acid Suppresses Colorectal Cancer through lncRNA 495810/PKM2 Mediated Aerobic Glycolysis. Int. J. Mol. Sci. 2022, 23, 12106. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.A.; Castaldo, S.; Rotondo, R.; Staffieri, S.; Sanchez, M.; Arcella, A. Inhibiting Effect of p-Coumaric Acid on U87MG Human Glioblastoma Cell Growth. J. Chemother. 2022, 34, 173–183. [Google Scholar] [CrossRef]
- Tehami, W.; Nani, A.; Khan, N.A.; Hichami, A. New Insights Into the Anticancer Effects of p-Coumaric Acid: Focus on Colorectal Cancer. Dose-Response 2023, 21, 15593258221150704. [Google Scholar] [CrossRef]
- Eroğlu, C.; Avcı, E.; Vural, H.; Kurar, E. Anticancer Mechanism of Sinapic Acid in PC-3 and LNCaP Human Prostate Cancer Cell Lines. Gene 2018, 671, 127–134. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, H.; Tan, P.; Huang, M.; Shi, H.; Sun, B.; Cheng, Y.; Li, T.; Mou, Z.; Li, Q.; et al. Sinapic Acid Inhibits Pancreatic Cancer Proliferation, Migration, and Invasion via Downregulation of the AKT/Gsk-3β Signal Pathway. Drug Dev. Res. 2021, 83, 721–734. [Google Scholar]
- Taştemur, Ş.; Hacısüleyman, L.; Karataş, Ö.; Yulak, F.; Ataseven, H. Anticancer Activity of Sinapic Acid by Inducing Apoptosis in HT-29 Human Colon Cancer Cell Line. Can. J. Physiol. Pharmacol. 2023, 101, 361–368. [Google Scholar] [CrossRef]
- Vasarri, M.; De Marchi, L.; Pretti, C.; Barletta, E.; Degl’Innocenti, D. Antioxidant and Anti-Inflammatory Properties of Four Native Mediterranean Seagrasses: A Review of Bioactive Potential and Ecological Context. Mar. Drugs 2025, 23, 206. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G.; Martínez-Zamora, L.; Peñalver, R.; Marín-Iniesta, F.; Taboada-Rodríguez, A.; López-Gómez, A.; Martínez-Hernández, G.B. Applications of Plant Bioactive Compounds as Replacers of Synthetic Additives in the Food Industry. Foods 2023, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Pastorino, G.; Borghesi, B.; Salis, A.; Clericuzio, M.; Marchetti, C.; Damonte, G.; Burlando, B. Posidonia oceanica (L.) Delile Ethanolic Extract Modulates Cell Activities with Skin Health Applications. Mar. Drugs 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Tahir, M.; Hong, Z.; Zia, M.A.; Rafeeq, H.; Ahmad, M.S.; Rehman, S.U.; Sun, J. Plant and marine-derived natural products: Sustainable pathways for future drug discovery and therapeutic development. Front. Pharmacol. 2025, 15, 1497668. [Google Scholar] [CrossRef]
- Jeong, J.S.; Kim, J.W.; Kim, J.H.; Chung, E.H.; Lee, D.R.; Choi, B.K.; Ko, J.W.; Kim, T.W. Oral toxicity and genotoxicity assessment of standardized Echinacea purpurea (L.) extract and the pharmacokinetic profile of its active ingredient chicoric acid. Toxicol. Res. 2024, 40, 457–472. [Google Scholar] [CrossRef]
- Ren, J.; Ren, X.; Ma, L.; Liu, J.; Yuan, S.; Wang, G. Pharmacokinetics and antioxidant activity of dihydrocaffeic acid grafted chitosan nanomicelles loaded with chicoric acid in broilers. Poult. Sci. 2024, 103, 03776. [Google Scholar] [CrossRef]
- Petersen, B.; Egert, S.; Bosy-Westphal, A.; Müller, M.J.; Wolffram, S.; Hubbermann, E.M.; Rimbach, G.; Schwarz, K. Bioavailability of quercetin in humans and the influence of food matrix comparing quercetin capsules and different apple sources. Food Res. Int. 2016, 88 Pt A, 159–165. [Google Scholar] [CrossRef]
- Konishi, Y.; Hitomi, Y.; Yoshioka, E. Intestinal absorption of p-coumaric and gallic acids in rats after oral administration. J. Agric. Food Chem. 2004, 52, 2527–2532. [Google Scholar] [CrossRef]
- Malik, N.; Dhiman, P. New Approaches and Advancements in Drug Development from Phenolic P-coumaric Acid. Curr. Top Med. Chem. 2022, 22, 1515–1529. [Google Scholar] [CrossRef]
- Posadino, A.M.; Cossu, A.; Giordo, R.; Zinellu, A.; Sotgia, S.; Vardeu, A.; Hoa, P.T.; Deiana, L.; Carru, C.; Pintus, G. Coumaric acid induces mitochondrial damage and oxidative- mediated cell death of human endothelial cells. Cardiovasc. Toxicol. 2013, 13, 301–306. [Google Scholar] [CrossRef]
- Seo, Y.K.; Kim, S.J.; Boo, Y.C.; Baek, J.H.; Lee, S.H.; Koh, J.S. Effects of p-coumaric acid on erythema and pigmentation of human skin exposed to ultraviolet radiation. Clin. Exp. Dermatol. 2011, 36, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, N.P.; Bondonno, C.P.; Rich, L.; Mas, E.; Shinde, S.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. Acute effects of quercetin-3-O-glucoside on endothelial function and blood pressure: A randomized dose-response study. Am. J. Clin. Nutr. 2016, 104, 97–103. [Google Scholar] [CrossRef] [PubMed]
- ElGamal, R.; Song, C.; Rayan, A.M.; Liu, C.; Al-Rejaie, S.; ElMasry, G. Thermal Degradation of Bioactive Compounds during Drying Process of Horticultural and Agronomic Products: A Comprehensive Overview. Agronomy 2023, 13, 1580. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Goutzourelas, N.; Kevrekidis, D.P.; Barda, S.; Malea, P.; Trachana, V.; Savvidi, S.; Kevrekidou, A.; Assimopoulou, A.N.; Goutas, A.; Liu, M.; et al. Antioxidant Activity and Inhibition of Liver Cancer Cells’ Growth of Extracts from 14 Marine Macroalgae Species of the Mediterranean Sea. Foods 2023, 12, 1310. [Google Scholar] [CrossRef]
- “Quest Graph™ IC50 Calculator.” AAT Bioquest, Inc., 28 April 2025. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 1 February 2025).
- Tsagias, N.; Koliakos, I.; Karagiannis, V.; Eleftheriadou, M.; Koliakos, G.G. Isolation of mesenchymal stem cells using the total length of umbilical cord for transplantation purposes. Transfus. Med. 2011, 21, 253–261. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Rt (min) | LR Extract (mg/g d.w.) | NR Extract (mg/g d.w.) | LR Material b (mg/g d.w.) | NR Material b (mg/g d.w.) |
|---|---|---|---|---|---|
| Caftaric acid | 4.56 | T | T | T | T |
| Chicoric acid | 12.42 | 41.09 | 45.40 | 11.50 | 4.68 |
| Quercetin-3-O-glucopyranoside | 22.53 | 10.22 | 10.74 | 2.86 | 1.10 |
| Caffeic acid | ND | ND | ND | ND | ND |
| (-)-Epigallocatechin gallate | ND | ND | ND | ND | ND |
| p-Coumaric acid | 11.87 | 9.14 | 10.76 | 2.55 | 1.10 |
| trans-Ferulic acid | 12.98 | 0.72 a | 0.82 | 0.20 | 0.08 |
| Sinapic acid (3,5,Dimethy-4-hydroxycinnamic acid) | 13.51 | 1.35 | 1.58 | 0.37 | 0.16 |
| Rutin hydrate | 16.82 | 5.26 | 6.00 | 1.47 | 0.61 |
| trans-Cinnamic acid | 20.66 | 0.17 a | 0.21 a | 0.04 | 0.02 |
| Hesperidin | 17.14 | NQ | NQ | NQ | NQ |
| 4′,5,7-Trihydroxyflavone | 27.45 | NQ | NQ | NQ | NQ |
| Total identified phenolics | 67.95 | 75.51 | 19.02 | 7.76 | |
| Extraction yield (%) | 28.00 | 10.30 |
| Phenolic Compounds | Amount (g) |
|---|---|
| Chicoric acid | 263.88 |
| Quercetin-3-O-glucopyranoside | 62.36 |
| p-Coumaric acid | 62.48 |
| trans-Ferulic acid | 4.76 |
| 3,5-Dimethy-4-hydroxycinnamic acid (Sinapic acid) | 9.17 |
| Rutin hydrate | 34.84 |
| trans-Cinnamic acid | 1.21 |
| Total identified phenolics | 438.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kevrekidou, A.; Goutzourelas, N.; Savvidi, S.; Trachana, V.; Assimopoulou, A.N.; Liu, M.; Malea, P.; Stagos, D. Comparison of Polyphenolic Content and Bioactivities Between Extracts from the Living Plants and Beach Deposits of the Submerged Brackish Water Angiosperm Ruppia maritima. Molecules 2025, 30, 2800. https://doi.org/10.3390/molecules30132800
Kevrekidou A, Goutzourelas N, Savvidi S, Trachana V, Assimopoulou AN, Liu M, Malea P, Stagos D. Comparison of Polyphenolic Content and Bioactivities Between Extracts from the Living Plants and Beach Deposits of the Submerged Brackish Water Angiosperm Ruppia maritima. Molecules. 2025; 30(13):2800. https://doi.org/10.3390/molecules30132800
Chicago/Turabian StyleKevrekidou, Alkistis, Nikolaos Goutzourelas, Stavroula Savvidi, Varvara Trachana, Andreana N. Assimopoulou, Ming Liu, Paraskevi Malea, and Dimitrios Stagos. 2025. "Comparison of Polyphenolic Content and Bioactivities Between Extracts from the Living Plants and Beach Deposits of the Submerged Brackish Water Angiosperm Ruppia maritima" Molecules 30, no. 13: 2800. https://doi.org/10.3390/molecules30132800
APA StyleKevrekidou, A., Goutzourelas, N., Savvidi, S., Trachana, V., Assimopoulou, A. N., Liu, M., Malea, P., & Stagos, D. (2025). Comparison of Polyphenolic Content and Bioactivities Between Extracts from the Living Plants and Beach Deposits of the Submerged Brackish Water Angiosperm Ruppia maritima. Molecules, 30(13), 2800. https://doi.org/10.3390/molecules30132800