
Anti-Inflammatory Activity of Thymol and Thymol-Rich Essential Oils: Mechanisms, Applications, and Recent Findings



Abstract
1. Introduction
2. Inflammation Mechanism
2.1. MAPK Signaling Pathway
2.2. NF-κB Signaling Pathway
2.3. JAK/STAT Signaling Pathway
2.4. Arachidonic Acid Signaling Pathway
3. Results and Discussion
3.1. Thymol-Rich Essential Oils/Volatiles
3.2. Anti-Inflammatory Activity of Essential Oils or Volatiles with Thymol
4. Thymol
4.1. In Vitro Anti-Inflammatory Activity of Thymol
4.2. In Vivo Anti-Inflammatory Properties of Thymol in Some Disease Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Disease Induction | Treatment | Results | Reference |
|---|---|---|---|---|
| Acute kidney injury | Glycerol injection in rats | 20 mg/kg and 40 mg/kg 24 h before glycerol administrations continued for 72 h, by gavage | IL-6↓, TNF-α↓, monocyte chemoattractant protein-1 (MCP-1)↓, NF-kB p65↓ | [79] |
| Chronic obstructive pulmonary disease | Porcine pancreatic elastase nasal instillation to mice | 20 mg/kg later the administration of elastase and then at days 7, 14 and 28, by instillation | IL-1β↓, IL-6↓, Il-8, IL-17, matrix metalloproteinase-9 (MMP-9)↓, p-65-NF-kB↓, 8-iso-PGF2α↓, NO exhalation↓ | [80] |
| Allergic inflammation | Intraperitoneal (i.p.) injection of ovalbumin at day 0, and on days 21, 22 and 23 the mice were submitted to intranasal inhalations | 16 mg/kg per os (p.o.) had similar response to 2 mg/kg of dexamethasone (positive control) | Ovalbumin-specific IgE↓, recruitment inhibition of eosinophils, neutrophils, macropages and lymphocytes into airway, IL-4↓, IL-5↓, IL-13, NF-kB↓ | [74] |
| Allergic rhinitis | Injection i.p. of ovalbumin in rats every other day for 14 days, then administration into both nostrils for 7 days | 20 mg/kg through intragastric lavage | IgE↓, IL-5↓, IL-13↓, total antioxidant capacity↓ | [81] |
| Ulcerative colitis | Dextrane sulfate sodium in drinking water to mice | 60 mg/kg p.o. | mRNA expression of TNF-α↓, IL-1β↓, IL-6↓. Malondalehyde (MDA) ↓, superoxide dismutase (SOD) activity↓, glutathione↑ | [82] |
| Colitis | Acetic acid intrarectally to rats | 100 mg/kg p.o. 5 days after the administration of acetic acid | MPO↓, TNF-α↓, upregulation inhibition of NF-kB p65 protein | [83] |
| Colitis | Dextrane sulfate sodium in the drink water to mice | 40 µL/kg thymol:carvacrol (1:1) p.o. for 14 day following 12 days of dextrane sulfate sodium exposure | Bifidobacterium pesudolongum↑, hyodeoxycholic acid (HDCA)↑, 12-ketodeoxycholic acid (12-KCAC)↑, protein kinase G (PKG) activated, mammalian target of rapamycin (mTOR) signaling pathway suppressed | [85] |
| Myocardial infarction | Isoproterenol injection subcutaneous at an interval of 24 h for 2 days to rats | Thymol (7.5 mg/kg) during pre- and cotreatment | Expression of TNF-α↓, IL-6↓, IL-1β↓. Thiobarbituric acid reactive substances (TBARS) ↓ | [86] |
| Atherosclerosis | Rabbits with high-fat and high cholesterol diet | Rabbits with a diet supplemented with 3 or 6 mg/kg/day | IL-1β↓, IL-6↓, TNF-α↓, TNF-β↓, vascular cell adhesion molecule-1 (VCAM-1) expression level↓, monocyte chemoattractant protein 1 (MCP-1)↓, matrix metalloproteinase 9 (MMP-9)↓ | [87] |
| Endometriosis | Female mice submitted with 17β-estradiol subcutaneously for 3 consecutive days. At the last injection day, the mice underwent surgical induction of endometriosis | 30 mg/kg/day or 60 mg/kg/day p.o. for 3 weeks | IL-1β↓, IL-6↓, TNF-α↓, TNF-β↓, neutrophil and macrophage counts↓, estrogen signaling inhibited by competing with estrogen for binding to estrogen receptor 1 (ESR1) | [89] |
| Hepatic encephalopathy | Thioacetamide i.p. 3 times per week, for 2 weeks to rats | 30 or 60 mg/kg p.o., daily for 1 month | Alanine aminotransferase (ALT)↓, aspartate aminotransferase (AST)↓, ammonia↓, brain oxidative stress↓, brain ATP↑, astrocyte swelling↓, brain edema↓, NF-kB protein expression↓, glial fibrillary acidic protein (GFAP) protein expression↑ | [90] |
| Parkinson’s disease | MnCl2 i.p. for 5 weeks to rats | 30 mg/kg/day p.o., 1 h before Mn administration for 5 weeks | TNF-α↓, TLR4↓, NF-kB↓, NLRP3↓, IL-1↓, COX-2↓ | [91] |
| Autism | Valproic acid to male rats | Thymol 30 mg/kg | Phosphorylated p38 MAPK↓, IL-1β↓, TNF-α↓, Pin1↓, postsynaptic density protein 95 (PSD95)↑, synaptophysin (SYP)↑ | [93] |
| Aspergillus fumigatus corneal keratitis | Right eyes of female mice infected with conidia (2.5 μL, 5 × 107 CFU/mL) | 50 µg/mL subconjunctival injection 1 day and 2 h before infection and treated with subconjunctival injection twice a day | TLR4↓, MyD88↓, NF-kB↓, IL-1β↓ | [94] |
5. Anti-Inflammatory Activity of Thymol in Drug-Induced Inflammation
| Drug/Side Effect | Treatment | Results | Reference |
|---|---|---|---|
| Cisplatin 6 mg/kg (single dose) intraperitoneally (i.p.) to male rats/nefrotoxicity | Thymol 20 mg/kg/day for 14 days prior to cisplatin and 7 days following cisplatin | Urea↓, creatinine↓, TNF-α↓, caspase-3↓ (43%) and MDA↓, (GSH)↑, SOD↑, catalase↑ | [99] |
| Doxorubicin 10 mg/kg (single dose) intravenously to rats/cardiotoxicity | Thymol 20 mg/kg/day for 14 days before doxorubicin and for 2 days after doxorubicin | Lactate dehydrogenase (LDH)↓, aspartate aminotransferase (AST)↓, creatine phosphokinase (CPK)↓, creatine kinase isoenzyme-MB (CK-MB)↓, cardiac troponin-I (cTn-I)↓, TNF-α↓ | [100] |
| Gentamicin 80 mg/kg/day i.p. starting from the 8th day to rats/nefrotoxicity | Thymol 20 mg/kg/day, p.o., for 15 days | Creatinine↓, MDA↓, NO↓, TNF-α↓, IL-18↓, Bax↓, caspases-3↓ and 9↓, renal NF-κB p65↓, kidney injury molecule-1 (KIM-1) expressions↓ | [101] |
| Bleomycin 15 mg/kg i.p., twice a week for 4 weeks to mice/lung fibrosis | Thymol 50 and 100 mg/kg p.o., for 4 weeks | MDA↓, GSH↑, SOD↑, TNF-α↓, IL-1β↓, IL-6↓, NF-kB↓, TGF-β↓, phospho-Akt↓ and PI3K↓ expression | [102] |
| 5-Fluorouracil (FU) 150 mg/kg on 6th and 7th days, i.p. to rats,/severe intestinal damages | Thymol 60 but particularly 120 mg/kg, p.o., for 11 days | GSH↑, SOD↑, glutathione peroxidase (GPx)↑, NF-κB expression↓, TNF-α protein level expression↓, NF-κB↓, TGF-β1↓, PGE2↓, COX-2↓, IL-6↓, IL-10↑, expression of p38↓, phosphorylated c-Jun N-terminal kinases (p-JNK)↓ | [104] |
| 5-Fluorouracil (FU) 150 mg/kg on 6th and 7th days, i.p. to rats/intestinal mucositis | Thymol 60 but particularly 120 mg/kg, p.o., for 11 days | IL-17↓, intestinal Notch1 expression↓, intestinal phospho-STAT3/total-STAT3 ratio↑ | [105] |
| Absolute ethanol 4 mL/kg p.o. 45 min after treatment/gastric ulcer | Thymol 30 mg/kg, p.o. | Total lesion area↓, amount of mucus↑, | [107] |
| Indomethacin 30 mg/kg p.o., to rats/hepatotoxicity | Thymol 250 mg/kg p.o. 10 min after the administration of indomethacin | AST↓, ALT↓, LDH↓, TNF-α↓, endothelial nitric oxide synthase (eNOS)↓, and caspase-3 activation↓, decreased PGE2↓ | [108] |
| Imidacloprid (pesticide) 22.5 mg/kg to rat/hepatotoxicity and brain disorder | Thymol 30 mg/kg | Hepatic enzyme leakage↓, hepatic oxidative stress↓, lipid peroxidation↓, DNA damage↓, NO↓ inflammatory cell infiltrations (portal and intralobular)↓ caspase-3↓, myeloperoxidase (MPO)↓ in brain | [110,111] |
6. Anti-Inflammatory Activity of Thymol in Livestock
7. Encapsulated Thymol in Micro- and Nanoparticles
8. Synergism Effect of Thymol and Other Natural Products
9. Material and Methods
10. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marinelli, L.; di Stefano, A.; Cacciatore, I. Carvacrol and its derivatives as antibacterial agents. Phytochem. Rev. 2018, 17, 903–921. [Google Scholar] [CrossRef]
- Mączka, W.; Twardawska, M.; Grabarczyk, M.; Wińska, K. Carvacrol—A natural phenolic compound with antimicrobial properties. Antibiotics 2023, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Haynes, W.M.; Lide, D.R.; Bruno, T.J. CRC Handbook of Chemistry and Physics, 95th ed.; CRC Press: Boca Raton, FL, USA; Taylor and Francis Group: London, UK; New York, NY, USA, 2014; eBook-pdf; ISBN 978-1-4822-0868-9. [Google Scholar]
- Escobar, A.; Pérz, M.; Romanelli, G.; Blustein, G. Thymol bioactivity: A review focusing on practical applications. Arab. J. Chem. 2020, 13, 9243–9269. [Google Scholar] [CrossRef]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; di Lorenzo, A.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef]
- Kazemi, M. Chemical composition and antimicrobial, antioxidant activities and anti-inflammatory potential of Achillea millefolium L., Anethum graveolens L., and Carum copticum L. essential oils. J. Herb. Med. 2015, 5, 217–222. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Kopaei, S.R.; Sajjadi, S.E. Anti-nociceptive and anti-inflammatory effects of Ferulago angulata. Immunopathol. Persa 2020, 6, e28. [Google Scholar] [CrossRef]
- Bourgou, S.; Rebey, I.B.; Kaab, S.B.; Hammami, M.; Dakhlaoui, S.; Sawsen, S.; Msaada, K.; Isoda, H.; Ksouri, R.; Fauconnier, M.-L. Green solvent to substitute hexane for bioactive lipids extraction from black cumin and basil seeds. Foods 2021, 10, 1493. [Google Scholar] [CrossRef]
- Mirahmad, A.; Ghoran, S.H.; Alipour, P.; Taktaz, F.; Hassan, S.; Naderian, M.; Moradalipour, A.; Faizi, M.; Kobarfard, F.; Ayatollahi, S.A. Oliveria decumbens Vent. (Apiaceae): Biological screening and chemical compositions. J. Ethnopharmacol. 2024, 318, 117053. [Google Scholar] [CrossRef]
- Kerbouche, L.; Hazzit, M.; Ferhat, M.-A.; Baaliouamer, A.; Miguel, M.G. Biological activities of essential oils and ethanol extracts of Teucrium polium subsp. capitatum (L.) Briq. and Origanum floribundum Munby. J. Essent. Oil Bear. Plants 2015, 18, 1197–1208. [Google Scholar] [CrossRef]
- Marrelli, M.M.; Araniti, F.; Abenavoli, M.R.; Statti, G.; Conforti, F. Potential health benefits of Origanum heracleoticum essential oil: Phytochemical and biological variability among different Calabrian populations. Nat. Prod. Commun. 2018, 13, 1183–1187. [Google Scholar] [CrossRef]
- Zinno, P.; Guantario, B.; Lombardi, G.; Ranaldi, G.; Finamore, A.; Allegra, S.; Mammano, M.M.; Fascella, G.; Raffo, A.; Roselli, M. Chemical composition and biological activities of essential oils from Origanum vulgare genotypes belonging to the carvacrol and thymol chemotypes. Plants 2023, 12, 1344. [Google Scholar] [CrossRef] [PubMed]
- Abiodun, O.O.; Oke, T.A.; Adeyemi, F.O.; Oshinloye, A.O.; Akande, A.O. Salacia pallescens Oliv. (Celastraceae) scavenges free radicals and inhibits pro-inflammatory mediators in lipopolysaccharide-activated RAW cells 264.7 macrophages. Turk. J. Pharm. Sci. 2021, 18, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Elbouny, H.; Ouahzizi, B.; El-Guourrami, O.; Drioua, S.; Mbarek, A.N.; Sellam, K.; Alem, C. Chemical profile and biological properties of the essential oil of Thymus atlanticus (Ball) Roussine. S. Afr. J. Bot. 2022, 151, 475–480. [Google Scholar] [CrossRef]
- Aazza, S.; El-Guendouz, S.; Miguel, M.G.; Antunes, M.D.; Faleiro, M.L.; Correia, A.I.; Figueiredo, A.C. Antioxidant, anti-inflammatory and anti-hyperglycaemic activities of essential oils from Thymbra capitata, Thymus albicans, Thymus caespititius, Thymus carnosus, Thymus lotocephalus and Thymus mastichina from Portugal. Nat. Prod. Commun. 2016, 11, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Chandra, M.; Prakash, O.; Bachheti, R.K.; Kumar, M.; Pant, A.K. Essential oil composition, phenolic constituents, antioxidant and pharmacological activities of Thymus linearis Benth. Collected from Uttarakhand region of India. J. Essent. Oil Bear. Plants 2016, 19, 277–289. [Google Scholar] [CrossRef]
- Demirci, F.; Karaca, N.; Tekin, M.; Demirci, B. Anti-inflammatory and antibacterial evaluation of Thymus sipyleus Boiss. subsp. sipyleus var. sipyleus essential oil against rhinosinusitis pathogens. Microb. Pathog. 2018, 122, 117–121. [Google Scholar]
- Pandur, E.; Micalizzi, G.; Mondello, L.; Horváth, A.; Sipos, K.; Horváth, G. Antioxidant and anti-inflammatory effects of thyme (Thymus vulgaris L.) essential oils prepared at different plant phenophases on Pseudomonas aeruginosa LPS-activated THP-1 macrophages. Antioxidants 2022, 11, 1330. [Google Scholar] [CrossRef] [PubMed]
- Warman, D.J.; Jia, H.; Kato, H. Effects of thyme (Thymus vulgaris L.) essential oil on aging-induced brain inflammation and blood telomere attrition in chronologically aged C57BL/6J mice. Antioxidants 2023, 12, 1178. [Google Scholar] [CrossRef]
- Spréa, R.M.; Caleja, C.; Finimundy, T.C.; Calhelha, R.C.; Pires, T.C.S.P.; Amaral, J.S.; Prieto, M.A.; Ferreira, I.C.F.R.; Pereira, E.; Marros, L. Chemical and bioactive evaluation of essential oils from edible and aromatic Mediterranean Lamiaceae plants. Molecules 2024, 29, 2827. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Bhattacharya, S.; Dutta, A.; Tauchen, J.; Landa, P.; Urbanová, K.; Houdková, M.; Fernández-Cusimamani, E.; Leuner, O. Synthetic polyploidization induces enhanced phytochemical profile and biological activities in Thymus vulgaris L. essential oil. Sci. Rep. 2024, 14, 5608. [Google Scholar] [CrossRef]
- Abdelli, W.; Bahri, F.; Romane, A.; Höferl, M.; Wanner, J.; Schmidt, E.; Jirovetz, L. Chemical composition and anti-inflammatory activity of Algerian Thymus vulgaris essential oil. Nat. Prod. Commun. 2017, 12, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Cutillas, A.-B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Thyme essential oils from Spain: Aromatic profile ascertained by GC-MS, and their antioxidant, anti-lipoxygenase and antimicrobial activities. J. Food Drug. Anal. 2018, 26, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.; Cabral, C.; Évora, L.; Ferreira, I.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Chemical composition, anti-inflammatory activity and cytotoxicity of Thymus zygis L. subsp. sylvestris (Hoffmanns. & Link) Cout. Essential oil and its main compounds. Arab. J. Chem. 2019, 12, 3236–3243. [Google Scholar]
- Kazemi, M. Chemical composition, antimicrobial, antioxidant and anti-inflammatory activity of Carum copticum L. essential oil. J. Essent. Oil Bear. Plants 2014, 17, 1040–1045. [Google Scholar] [CrossRef]
- Vanitha, K.G.; Natarajan, A.; Sudhkar, N.; Hirad, A.H.; Alarfaj, A.A.; Arulselvan, P.; Raja, R. Enhancing therapeutic potential: Investigating traditional detoxification methods and assessing their influence on anti-microbial efficacy, phytochemical composition, heavy metal content and anti-inflammatory properties in Trachyspermum ammi. Ind. J. Pharm. Edu. Res. 2025, 59, 230–241. [Google Scholar] [CrossRef]
- Dutta, P.; Sarma, N.; Saikia, S.; Gogoi, R.; Begum, T.; Lal, M. Pharmacological activity of Trachyspermum ammi L. seeds essential oil grown from Northeast India. J. Essent. Oil Bear. Plants 2021, 24, 1373–1388. [Google Scholar] [CrossRef]
- Bahuguna, A.; Ramalingam, S.; Arumugam, A.; Natarajan, D.; Kim, M. Molecular and in silico evidences explain the anti-inflammatory effect of Trachyspermum ammi essential oil in lipopolysaccharide induced macrophages. Process Biochem. 2020, 96, 138–145. [Google Scholar] [CrossRef]
- Jayawantha, D.; Hettigoda, L.; Mudalige, T.D.; Paranagama, P.A. Exploring the bioactivity of siddhalepa asamodagam spirit from seeds of Trachyspermum roxburghianum (DC.) H. Wolff. Nat. Prod. Commun. 2024, 19, 1934578X241271629. [Google Scholar] [CrossRef]
- Gabbai-Armelin, P.R.; Sales, L.S.; Ferrisse, T.M.; de Oliveira, A.B.; de Oliveira, J.R.; Giro, E.M.A.; Brighenti, F.L. A systematic review and meta-analysis of the effect of thymol as an anti-inflammatory and wound healing agent. Phytother. Res. 2022, 36, 3415–3443. [Google Scholar] [CrossRef]
- Najafloo, R.; Behyari, M.; Imani, R.; Nour, S. A mini-review of thymol incorporated materials: Applications in antibacterial wound dressing. J. Drug Deliv. Sci. Technol. 2020, 60, 101904. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef] [PubMed]
- El Boukhari, R.; Fatimi, A. Thymol’s Innovative health and medicine applications: An overview based on patent analysis and patent review. Curr. Trad. Med. 2025, 11, 1–12. [Google Scholar] [CrossRef]
- Peter, S.; Sotondoshe, N.; Aderibigbe, B.A. Carvacrol and thymol hybrids: Potential anticancer and antibacterial therapeutics. Molecules 2024, 29, 2277. [Google Scholar] [CrossRef] [PubMed]
- Favier, A.; Nikovics, K. Molecular and cellular mechanisms of inflammation and tissue regeneration. Biomedicines 2023, 11, 1416. [Google Scholar] [CrossRef]
- Deng, Z.; Liu, S. Inflammation-responsive delivery systems for the treatment of chronic inflammatory diseases. Drug Deliv. Transl. Res. 2021, 11, 1475–1497. [Google Scholar] [CrossRef] [PubMed]
- Chagas, M.S.S.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Gonçalves-de-Albuquerque, C.F. Flavonols and flavones as potential anti-inflammatory, antioxidant, and antibacterial compounds. Oxid. Med. Cell Longev. 2022, 2022, 9966750. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Cote, B.; Elbarbry, F.; Bui, F.; Su, J.W.; Seo, K.; Nguyen, A.; Lee, M.; Rao, D.A. Mechanistic basis for the role of phytochemicals in inflammation-associated chronic diseases. Molecules. 2022, 27, 781. [Google Scholar] [CrossRef] [PubMed]
- Ivanenkov, Y.A.; Balakin, K.V.; Lavrovsky, Y. Small molecule inhibitors of NF-kB and JAK/STAT signal transduction pathways as promising anti-inflammatory therapeutics. Mini Rev. Med. Chem. 2011, 11, 55–78. [Google Scholar] [CrossRef]
- Sharma, V.; Tiwari, R.K.; Shukla, S.S.; Pandey, R.K. Current and future molecular mechanism in inflammation and arthritis. J. Pharmacopunct. 2020, 23, 54–61. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr Rev. 2007, 65 Pt 2, S140–S146. [Google Scholar] [CrossRef] [PubMed]
- Tabbas, I.; Glass, C.K. Anti-inflammatory therapy in chronic disease: Challenges and opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.L.R.; Wilairatana, P.; Silva, L.R.; Moreira, P.S.; Barbosa, N.M.M.V.; da Silva, P.R.; Coutinho, H.D.M.; de Menezes, I.R.A.; Felipe, C.F.B. Biochemical aspects of the inflammatory process: A narrative review. Biomed. Pharmacother. 2023, 168, 115764. [Google Scholar] [CrossRef]
- Jantan, I.; Haque, M.A.; Arshad, L.; Harikrishnan, H.; Septama, A.W.; Mohamed-Hussein, Z. Dietary polyphenols suppress chronic inflammation by modulation of multiple inflammation-associated cell signaling pathways. J. Nutr. Biochem. 2021, 93, 108634. [Google Scholar] [CrossRef] [PubMed]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Anuar, N.N.M.; Jalil, J.; Nor, N.A.M.; Budin, S.B. The role of PKC-MAPK signalling pathways in the development of hyperglycemia-induced cardiovascular complications. Int. J. Mol. Sci. 2022, 23, 8582. [Google Scholar] [CrossRef]
- Gur-Arie, L.; Eitan-Wexler, M.; Weinberger, N.; Rosenshine, I.; Livnah, O. The bacterial metalloprotease NleD selectively cleaves mitogen-activated protein kinases that have high flexibility in their activation loop. J. Biol. Chem. 2020, 295, 9409–9420. [Google Scholar] [CrossRef]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Nadipelly, J. Molecular mechanisms involved in inflammatory cascade: A review. Texila Int. J. Basic Med. Sci. 2017, 2, 1–12. [Google Scholar] [CrossRef]
- Wu, Y.-X.; Lu, W.-W.; Geng, Y.-C.; Yu, C.-H.; Sun, H.-J.; Kim, Y.-J.; Zhang, G.; Kim, T. Antioxidant, antimicrobial and anti-inflammatory activities of essential oil derived from the wild rhizome of Atractylodes macrocephala. Chem. Biodivers. 2020, 17, e2000268. [Google Scholar] [CrossRef]
- Foudah, A.I.; Alqarni, M.H.; Alam, A.; Salkini, M.A.; Ahmed, E.O.I.; Yusufoglu, H.S. Evaluation of the composition and in vitro antimicrobial, antioxidant, and anti-inflammatory activities of Cilantro (Coriandrum sativum L. leaves) cultivated in Saudi Arabia (Al-Kharj). Saudi J. Biol. Sci. 2021, 28, 3461–3468. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.; Kumar, A.; Bawankule, D.U.; Tandon, S.; Singh, A.K.; Verma, R.S.; Saikia, D. Chemical composition, bactericidal kinetics, mechanism of action, and anti-inflammatory activity of Isodon melissoides (Benth.) H. Hara essential oil. Nat. Prod. Res. 2021, 35, 690–695. [Google Scholar] [CrossRef]
- Nea, F.; Kambiré, D.A.; Genva, M.; Tanoh, E.A.; Wognin, E.L.; Martin, H.; Brostaux, Y.; Tomi, F.; Lognay, G.C.; Tonzibo, Z.F.; et al. Composition, seasonal variation, and biological activities of Lantana camara essential oils from Côte d’Ivoire. Molecules 2020, 25, 2400. [Google Scholar] [CrossRef] [PubMed]
- Nea, F.; Tanoh, E.A.; Wognin, E.L.; Kemene, T.K.; Genva, M.; Saive, M.; Tonzibo, Z.F.; Fauconnier, M.-L. A new chemotype of Lantana rhodesiensis Moldenke essential oil from Côte d’Ivoire: Chemical composition and biological activities. Ind. Crops Prod. 2019, 141, 111766. [Google Scholar] [CrossRef]
- Bayala, B.; Bassole, I.H.N.; Gnoula, C.; Nebie, R.; Yonli, A.; Morel, L.; Figueredo, G.; Nikiema, J.-B.; Lobaccaro, J.-M.A.; Simpore, J. Chemical composition, antioxidant, anti-inflammatory and anti-proliferative activities of essential oils of plants from Burkina Faso. PLoS ONE 2014, 9, e92122. [Google Scholar] [CrossRef]
- Su, Y.-C.; Hsu, K.P.; Ho, C.-L. Composition, in vitro anti-inflammatory, antioxidant and antimicrobial activities of rhe leaf essential oil of Machilus konishii from Taiwan. Nat. Prod. Commun. 2016, 11, 1363–1366. [Google Scholar]
- Fraternale, D.; Dufat, H.; Albertini, M.C.; Bouzidi, C.; d’Adderio, R.; Coppari, S.; Giacomo, B.; Melandri, D.; Ramakrishbna, S.; Colomba, M. Chemical composition, antioxidant and anti-inflammatory properties of Monarda didyma L. essential oil. PeerJ 2022, 10, e14433. [Google Scholar] [CrossRef] [PubMed]
- Kanyal, J.; Prakash, O.; Kumar, R.; Rawat, D.S. Essential oil composition and biological activities determination of Mosla dianthera (Buch.-Ham. ex Roxb.) Maxim. and its major isolated component, carvone. Braz. J. Pharm. Sci. 2022, 58, e201031. [Google Scholar] [CrossRef]
- Kazemi, M. Phytochemical composition, antioxidant, anti-inflammatory and antimicrobial activity of Nigella sativa L. essential oil. J. Essent. Oil Bear. Plants 2014, 17, 1002–1011. [Google Scholar] [CrossRef]
- Yuan, Z.; Shafiq, M.; Zheng, H.; Zhang, L.; Wang, Z.; Yu, X.; Song, J.; Sun, B.; El-Newehy, M.; El-Hamshary, H.; et al. Multi-functional fibrous dressings for infectious injury treatment with anti-adhesion wound healing. Mater. Des. 2023, 235, 112459. [Google Scholar] [CrossRef]
- Al-Mijalli, S.H.; Mrabti, N.N.; Ouassou, H.; Sheikh, R.A.; Assaggaf, H.; Bakrim, S.; Abdallah, E.M.; Alshahrani, M.M.; Al Awadh, A.A.; Lee, L.-H.; et al. Chemical composition and antioxidant, antimicrobial, and anti-inflammatory properties of Origanum compactum Benth essential oils from two regions: In vitro and in vivo evidence and in silico molecular investigations. Molecules. 2022, 27, 7329. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, N.M.; Mitić, K.V.; Nešić, M.; Stanković, M.; Petrović, V.; Baralić, M.; Randjelović, P.J.; Sokolović, D.; Radulović, N. Oregano (Origanum vulgare) essential oil and its constituents prevent rat kidney tissue injury and inflammation induced by a high dose of L-arginine. Int. J. Mol. Sci. 2024, 25, 941. [Google Scholar] [CrossRef]
- Naccari, C.; Ginestra, G.; Micale, N.; Palma, E.; Galletta, B.; Costa, R.; Vadalà, R.; Nostro, A.; Cristani, M. Binary combinations of essential oils: Antibacterial activity against Staphylococcus aureus, and antioxidant and anti-inflammatory properties. Molecules 2025, 30, 438. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, A.; Perez, E.; Cutillas, A.-B.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Origanum vulgare and Thymbra capitata essential oils from Spain: Determination of aromatic profile and bioactivities. Nat. Prod. Commun. 2016, 11, 113–120. [Google Scholar] [CrossRef]
- Poulouse, A.J.; Croteau, R. Biosynthesis of aromatic monoterpenes: Conversion of γ-terpinene to p-cymene and thymol in Thymus vulgaris L. Arch. Biochem. Biophys. 1978, 187, 307–314. [Google Scholar] [CrossRef]
- Rudolph, K.; Parthier, C.; Egerer-Sieber, C.; Geiger, D.; Muller, Y.A.; Kreiss, W.; Müller-Uri, F. Expression, crystallization and structure elucidation of γ-terpinene synthase from Thymus vulgaris. Acta Cryst. 2016, F72, 16–23. [Google Scholar]
- Maryam, S.; Khan, M.R.; Shah, S.A.; Zahra, Z.; Batool, R.; Zai, J.A. Evaluation of anti-inflammatory potential of the leaves of Wendlandia heynei (Schult.) Santapau & Merchant in Sprague Dawley rat. J. Ethnopharmacol. 2019, 238, 111849. [Google Scholar] [CrossRef] [PubMed]
- Mangal, C.S.K.; Anitha, R.; Lakshmi, T. Inhibition of nitric oxide production and nitric oxide gene expression in LPS activated RAW264.7 macrophages thyme oleoresin from Thymus vulgaris. J. Young Pharm. 2018, 10, 481–483. [Google Scholar] [CrossRef]
- Erzurumlu, Y.; Dogan, H.K.; Catakli, D. Thymol reduces the lipopolysaccharide-induced acute kidney inflammation by modulating lysosomal stress. J. Res. Pharm. 2023, 27, 375–385. [Google Scholar]
- Wan, L.; Meng, D.; Wang, H.; Wan, S.; Jiang, S.; Huang, S.; Wei, L.; Yu, P. Preventive and therapeutic effects of thymol in a lipopolysaccharide-induced acute lung injury mice model. Inflammation 2018, 41, 183–192. [Google Scholar] [CrossRef]
- Yao, L.; Hou, G.; Wang, L.; Zuo, X.-s.; Liu, Z. Protective effects of thymol on LPS-induced acute lung injury in mice. Microb. Pathog. 2018, 116, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, M.; Romilde, I.; Beatrice, M.M.; Matteo, V.; Giovanna, N.; Consuelo, A.; Claudio, C.; Giorgio, S.; Filippo, M.; Massimo, N. Evaluations of thyme extract effects in human normal bronchial and tracheal epithelial cell lines and in human lung cancer cell line. Chem. Biol. Interact. 2016, 256, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Zhou, E.; Fu, Y.; Wei, Z.; Yu, Y.; Zhang, X.; Yang, Z. Thymol attenuates allergic airway inflammation in ovalbumin (OVA)-induced mouse asthma. Fitoterapia 2014, 96, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Jiang, K.; Yin, N.; Ma, X.; Zhao, G.; Qiu, C.; Deng, G. Thymol mitigates lipopolysaccharide-induced endometritis by regulating the TLR4- and ROS-mediated NF-κB signaling pathways. Oncotarget 2017, 8, 20042–20055. [Google Scholar] [CrossRef]
- Javadian, S.; Sabouni, F.; Haghbeen, K. Origanum vulgare L. extracts versus thymol: An anti-inflammatory study on activated microglial and mixed glial cells. J. Food Biochem. 2016, 40, 100–108. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, L.; Zhang, Y.; Shu, X.; Hu, Y.; Chen, D.; Zhang, Z.; Xia, S.; Yang, H.; Bao, X.; et al. Thymol improves ischemic brain injury by inhibiting microglia-mediated neuroinflammation. Brain Res. Bull. 2024, 215, 111029. [Google Scholar] [CrossRef]
- Lanzarin, G.A.B.; Félix, L.M.; Monteiro, S.M.; Ferreira, J.M.; Oliveira, P.A.; Venâncio, C. Anti-inflammatory, anti-oxidative and anti-apoptotic effects of thymol and 24-epibrassinolide in zebrafish larvae. Antioxidants 2023, 12, 1297. [Google Scholar] [CrossRef]
- Wang, Q.; Qi, G.; Zhou, H.; Cheng, F.; Yang, X.; Liu, X.; Wang, R. Protective effect of thymol on glycerol-induced acute kidney injury. Ren. Fail. 2023, 45, 12227728. [Google Scholar] [CrossRef]
- Games, E.; Guerreiro, M.; Santana, F.R.; Pinheiro, N.M.; de Oliveira, E.A.; Lopes, F.D.Q.S.; Olivo, C.R.; Tibério, I.F.L.C.; Martins, M.A.; Lago, J.H.G.; et al. Structurally related monoterpenes p-cymene, carvacrol and thymol isolated from essential oil from leaves of Lippia sidoides Cham. (Verbenaceae) protect mice against elastase-induced emphysema. Molecules 2016, 21, 1390. [Google Scholar] [CrossRef]
- Kilic, K.; Sakat, M.S.; Yildirim, S.; Kandemir, F.M.; Gozeler, M.S.; Dortbudak, M.B.; Kucukler, S. The amendatory effect of hesperidin and thymol in allergic rhinitis: An ovalbumin-induced rat mode. Eur. Arch. Otorhinolaryngol. 2019, 276, 407–415. [Google Scholar] [CrossRef]
- Liu, D.-M.; Zhou, C.-Y.; Meng, X.-L.; Wang, P.; Li, W. Thymol exerts anti-inflammatory effect in dextran sulfate sodium-induced experimental murine colitis. Trop. J. Pharm. Res. 2018, 17, 1803–1810. [Google Scholar] [CrossRef]
- Chamanara, C.; Abdollahi, A.; Rezayat, S.M.; Ghazi-Khanasari, M.; Dehpour, A.; Nassireslami, E.; Rashidian, A. Thymol reduces acetic acid-induced inflammatory response through inhibition of NF-κB signaling pathway in rat colon tissue. Inflammopharmacology 2019, 27, 1275–1283. [Google Scholar] [CrossRef]
- Laurindo, L.F.; dos Santos, A.R.O.; de Carvalho, A.C.A.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.A.; Sinatora, R.V.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and regulation of NF-κB in inflammatory bowel diseases: An overview of in vitro and in vivo effects. Metabolites 2023, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, Y.; Zheng, Y.; Zhang, T.; Wu, Y.; Yan, Y.; Lei, Y.; Cao, X.; Wang, X.; Yan, F.; et al. Bifidobacterium pseudolongum-derived bile acid from dietary carvacrol and thymol supplementation attenuates colitis via cGMP-PKG-mTORC1 pathway. Adv. Sci. 2024, 11, 2406917. [Google Scholar] [CrossRef]
- Meeran, M.F.N.; Jagadeesh, G.S.; Selvaraj, P. Thymol attenuates inflammation is isoproterenol induced myocardial infacted rats by inhibiting the release of lysosomal enzymes and downregulating the expressions of proinflammatory cytokines. Eur. J. Pharmacol. 2015, 754, 153–161. [Google Scholar] [CrossRef]
- Yu, Y.-M.; Chao, T.-Y.; Chang, W.-C.; Chang, M.J.; Lee, M.-F. Thymol reduces oxidative stress, aortic intimal thickening, and inflammation-related gene expression in hyperlipidemic rabbits. J. Food Drug Anal. 2016, 24, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Luo, C.; Li, X.; Zhou, Y.; He, H. The anti-inflammatory effect of kaempferol on early atherosclerosis in high cholesterol fed rabbits. Lipids Health Dis. 2013, 12, 115. Available online: http://www.lipidworld.com/content/12/1/115 (accessed on 2 May 2025). [CrossRef]
- Zhang, Y.; Shaukat, A.; Zhang, H.; Yang, Y.-F.; Li, H.-X.; Li, G.-Y.; Liu, Y.-N.; Liang, C.; Kang, J.-W.; Li, S.-C.; et al. Thymol impacts the progression of endometriosis by disrupting estrogen signaling pathways and inflammatory responses. Int. J. Mol. Sci. 2024, 25, 13150. [Google Scholar] [CrossRef]
- Ogaly, H.A.; Abdel-Rahman, R.F.; Mohamed, M.A.E.; Ahmed-Farid, O.A.; Khattab, M.S.; Abd-Elsalam, R.M. Thymol ameliorated neurotoxicity and cognitive deterioration in a thioacetamide-induced hepatic encephalopathy rat model; involvement of the BDNF/CREB signaling pathway. Food Funct. 2022, 13, 6180. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elfotuh, K.; Hamdan, A.M.E.; Mohammed, A.A.; Atwa, A.M.; Kozman, M.R.; Ibrahim, A.M.; Motawea, S.M.; Selim, H.M.R.M.; Tohamy, S.T.K.; El-Din, M.N.; et al. Neuroprotective effects of some nutraceutical against manganese-induced Parlinson’s disease in rats: Possible modulatory effects on TLR4/NLRP3/NF-κB, GSK-3β, Nrf2/HO-1, and apoptotic pathways. Pharmaceuticals 2022, 15, 1554. [Google Scholar] [CrossRef] [PubMed]
- Herrera, T.E.S.; Tello, I.P.S.; Mustafa, M.A.; Jamil, N.Y.; Alaraj, M.; Altameen, K.K.A.; Alasheqi, M.Q.; Hamoody, A.-H.M.; Alkhafaji, A.T.; Shakir, M.N.; et al. Kaempferol: Unveiling its anti-inflammatory properties for therapeutic innovation. Cytokine 2025, 186, 156846. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, J.; Lv, M.; Wang, F.; Zhang, H.; Tang, B.; Li, Y. Thymol improves autism-like behavior in VPA-induced ASD rats through the Pin1/p38 MAPK pathway. Int. Immunopharmacol. 2023, 117, 109885. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yan, H.; Chen, X.; Han, L.; Liu, G.; Yang, H.; Lu, D.; Liu, W.; Che, C. Thymol ameliorates Aspergillus fumigatus keratitis by downregulating the TLR4/MyD88/NF/IL-1β signal expression and reducing necroptosis and pyroptosis. J. Microbiol. Biotechnol. 2023, 33, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Liu, J.; Liu, H.; Chen, X.; Yu, N.; Li, X.; Xu, F. Integration of network pharmacology and molecular docking to analyse the mechanism of action of oregano essential oil in the treatment of bovine mastitis. Vet. Sci. 2023, 10, 350. [Google Scholar] [CrossRef]
- Chavda, V.P.; Feehan, J.; Apostolopoulos, V. Inflammation: The cause of all diseases. Cells 2024, 13, 1906. [Google Scholar] [CrossRef] [PubMed]
- Orlando, F.A.; Mainous III, A.G. Editorial: Inflammation and chronic disease. Front. Med. 2024, 11, 1434533. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- El-Sayed, E.M.; Abd-Allah, A.R.; Mansour, A.M.; El-Arabey, A.A. Thymol and carvacrol prevent cisplatin-induced nephrotoxicity by abrogation of oxidative stress, inflammation, and apoptosis in rats. J. Biochem. Mol. Toxicol. 2015, 29, 165–172. [Google Scholar]
- El-Sayed, E.M.; Mansour, A.M.; Abdul-Hameed, M.S. Thymol and carvacrol prevent doxorubicin-induced cardiotoxicity by abrogation of oxidative stress, inflammation, and apoptosis rats. J. Biochem. Mol. Toxicol. 2016, 30, 37–44. [Google Scholar] [CrossRef]
- Fouad, A.A.; Moussa, N.A.; Kareem, M.M.A.; Akl, U.I.; Abdelghany, M.I.; Abdel-Aziz, A.M. Thymol exerts antioxidant, anti-inflammatory, and anti-apoptotic protective effects against gentamicin nephrotoxicity in rats. Pharmacia 2022, 69, 181–186. [Google Scholar] [CrossRef]
- Hussein, R.M.; Arafa, E.-S.A.; Raheem, S.A.; Mohamed, W.R. Thymol protects against bleomycin-induced pulmonary fibrosis via abrogation of oxidative stress, inflammation, and modulation of miR-29a/TGF-β and PI3K/Akt signaling in mice. Life Sci. 2023, 314, 121256. [Google Scholar] [CrossRef] [PubMed]
- Eräsalo, H.; Hämäläien, M.; Leppänen, T.; Mäki-Opas, I.; Laavola, M.; Haavikko, R.; Yli-Kauhaluoma, J.; Moilanen, E. Natural stilbenoids have anti-inflammatory properties in vivo and down-regulate the production of inflammatory mediators NO, IL6, and MCP1 possibly in a PI3K/Akt-dependent manner. J. Nat. Prod. 2018, 81, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Al-Khrashi, L.A.; Badr, A.M.; Al-Amin, M.A.; Mahran, Y.F. Thymol ameliorates 5-fluouracil-induced intestinal mucositis: Evidence of down-regulatory effect on TGF-β/MAPK pathways through NF-κB. J. Biochem. Mol. Toxicol. 2022, 36, e22932. [Google Scholar] [CrossRef] [PubMed]
- Badr, A.M.; Alkharashi, L.A.; Sherif, I.O.; Alanteet, A.A.; Alotaibi, H.N.; Mahran, Y.F. IL-17/Notch1/STAT3 pathway contributes to 5-fluorouracil-induced intestinal mucositis in rats: Amelioration by thymol treatment. Pharmaceuticals 2022, 15, 1412. [Google Scholar] [CrossRef] [PubMed]
- Gholijani, N.; Gharagozloo, M.; Farjadian, S.; Amirghofran, Z. Modulatory effects of thymol and carvacrol on inflammatory transcription factors in lipopolysaccharide-treated macrophages. J. Immunotoxicol. 2016, 13, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.R.S.; Diniz, P.B.F.; Pinheiro, M.S.; Albuquerque-Júnior, R.L.C.; Thomazzi, S.M. Gastroprotective effects of thymol on acute and chronic ulcers in rats. The role of prostaglandins, ATP-sensitive K+ channels, and gastric mucus secretion. Chem. Biol. Interact. 2016, 244, 121–128. [Google Scholar] [CrossRef]
- Geyikoglu, F.; Yilmaz, E.G.; Erol, H.S.; Koc, K.; Cerig, S.; Ozek, N.S.; Aysin, F. Hepatoprotective role of thymol in drug-induced gastric ulcer model. Annals Hepatol. 2018, 17, 980–991. [Google Scholar] [CrossRef]
- Ashraf, Z.; Alamgeer; Kanwal, M.; Hassan, M.; Abdullah, S.; Waheed, M.; Ahsan, H.; Kim, S.J. Flurbiprofen-antioxidant mutual prodrugs as safer nonsteroidal anti-inflammatory drugs: Synthesis, pharmacological investigation, and computational molecular modeling. Drug Des. Devel. Ther. 2016, 10, 2401–2419. [Google Scholar]
- Abdelgawad, F.E.; El-Rahman, G.I.A.; Behairy, A.; Abd-Elhakim, Y.M.; Saber, T.M.; Metwally, M.M.M.; El-Fatah, S.S.A.; Samaha, M.M.; Saber, T.; Aglan, M.A. Thymol’s modulation of cellular macromolecules, oxidative stress, DNA damage, and NF-κB/caspase-3 signaling in the liver of imidacloprid-exposed rats. Environ. Toxicol. Pharmacol. 2024, 109, 104492. [Google Scholar] [CrossRef]
- Abd-Elhakim, Y.M.; Saber, T.M.; Metwally, M.M.M.; Abd-Allah, N.A.; Mohamed, R.M.S.M.; Ahmed, G.A. Thymol abates the detrimental impacts of imidacloprid on rat brains by lessening oxidative damage and apoptotic and inflammatory reactions. Chem. Biol. Interact. 2023, 383, 110690. [Google Scholar] [CrossRef]
- Li, D.; Yang, M.; Ma, Z.; Che, L.; Feng, B.; Xu, S.; Zhuo, Y.; Li, J.; Wang, J.; Zhang, Z.; et al. Glycerol monolaurate complex improved antioxidant, anti-inflammation, and gut microbiota composition of offspring in a sow-piglet model. Vet. Sci. 2025, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, X.; Zhang, K.; Tian, G.; Ding, X.; Bai, S.; Zeng, Q. Effects of thymol and carvacrol eutectic on growth performance, serum biochemical parameters, and intestinal health in broiler chickens. Animals 2023, 13, 2242. [Google Scholar] [CrossRef] [PubMed]
- Ghiselli, F.; Yu, L.-E.; Piva, A.; Grilli, E.; Li, Y. Evaluating protective effects of botanicals under inflammation and oxidative stress in chicken apical-out enteroids. Poultry Sci. 2023, 102, 102821. [Google Scholar] [CrossRef]
- Ibrahim, D.; Shahin, S.E.; Alqahtani, L.S.; Hassan, Z.; Althobaiti, F.; Albogami, S.; Soliman, M.M.; El-Malt, R.M.S.; Al-Harthi, H.F.; Alqadri, N.; et al. Exploring the interactive effects of thymol and thymoquinone: Moving towards an enhanced performance, gross margin, immunity and Aeromonas sobria resistance of Nile tilapia (Oreochromis niloticus). Animals 2022, 12, 3034. [Google Scholar] [CrossRef] [PubMed]
- Pivetta, T.P.; Simões, S.; Araújo, M.M.M.; Carvalho, T.; Arruda, C.; Marcato, P.D. Development of nanoparticles from natural lipids for topical delivery of thymol: Investigation of its anti-inflammatory properties. Colloids Surf. B. Biointerfaces 2018, 164, 281–290. [Google Scholar] [CrossRef]
- García-Salinas, S.; Evangelopoulos, M.; Gámez-Herrera, E.; Arruebo, M.; Irusta, S.; Taraballi, F.; Mendoza, G.; Tasciotti, E. Electrospun anti-inflammatory patch loaded with essential oils for wound healing. Int. J. Pharm. 2020, 577, 119067. [Google Scholar] [CrossRef] [PubMed]
- Folle, C.; Díaz-Garrido, N.; Sánchez-López, E.; Marqués, A.M.; Badia, J.; Baldomà, L.; Espina, M.; Calpena, A.C.; García, M.L. Surface-modified multifunctional thymol-loaded biodegradable nanoparticles for topical acne treatment. Pharmaceutics 2021, 13, 1501. [Google Scholar] [CrossRef] [PubMed]
- Ahmady, A.R.; Razmjooee, K.; Saber-Samandari, S.; Toghraie, D. fabrication of chitosan-gelatin films incorporated with thymol-loaded alginate microparticles for controlled drug delivery, antibacterial activity and wound healing: In vitro and in vivo studies. Int. J. Biol. Macromol. 2022, 223, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Sheorain, J.; Mehra, M.; Thakur, R.; Grewal, S.; Kumari, S. In vitro anti-inflammatory and antioxidant potential of thymol loaded bipolymeric (tragacanth gum/chitosan) nanocarrier. Int. J. Biol. Macromol. 2019, 125, 1069–1074. [Google Scholar] [CrossRef]
- Safarbalou, A.; Abbasi, A. Oral administration of liposome-encapsulated thymol could alleviate the inflammatory parameters in serum and hippocampus in a rat model of Alzheimer’s disease. Exper. Gerontol. 2024, 193, 112473. [Google Scholar] [CrossRef]
- Zheng, M.; Huang, Y.; Hu, W.; Li, R.; Wang, J.; Han, M.; Li, Z. Evaluation of the antibacterial, anti-inflammatory, and bone-promoting capacity of UiO-66 loaded with thymol or carvacrol. Appl. Mater. Interfaces 2024, 16, 36017–36029. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.N.; Arsenault, R.J.; Piva, A.; Grilli, E.; Swaggerty, C.L. A microencapsulated feed additive containing organic acids and botanicals has a distinct effect on proliferative and metabolic related signaling in the jejunum and ileum of broiler chickens. Front. Physiol. 2023, 14, 1147483. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, D.-L.; Xie, L.-N.; Ma, Y.-r.; Wu, P.-P.; Li, C.; Liu, W.-F.; Zhang, K.; Zhou, R.-P.; Xu, X.-T.; et al. Synergistic anti-inflammatory effects of silibinin and thymol combination on LPS-induced RAW264.7 cells by inhibition of NF-κB and MAPK activation. Phytomedicine 2020, 78, 153309. [Google Scholar] [CrossRef] [PubMed]
- Golbahari, S.; Meysam, S.; Froushani, A. Synergistic benefits of nicotine and thymol in alleviating experimental rheumatoid arthritis. Life Sci. 2019, 239, 117037. [Google Scholar] [CrossRef] [PubMed]
- Frota, R.M.; Gallani, S.U.; dos Santos, P.D.P.; Pereira, C.S.; Oishi, C.A.; Gonçalves, L.U.; Valladão, G.M.R. Effects of thymol:carvacrol association on health and zootechnical performance of tambaqi (Colossoma macropomum). Bol. Inst. Pesca 2022, 48, e725. [Google Scholar] [CrossRef]
- Islam, M.T.; Bappi, M.H.; Bhuia, M.S.; Ansari, S.A.; Ansari, I.A.; Shill, M.C.; Albayouk, T.; Saleh, N.; El-Shazly, M.; El-Nashar, H.A.S. Anti-inflammatory effects of thymol: An emphasis on the molecular interactions through in vivo approach and molecular dynamic simulations. Front. Chem. 2024, 12, 1376783. [Google Scholar] [CrossRef]
- Islam, M.T.; Khalipha, A.B.R.; Bagchi, R.; Mondal, M.; Smrity, S.Z.; Uddin, S.J.; Shilpi, J.A.; Rouf, R. Anticancer activity of thymol: A literature-based review and docking study with emphasis on its anticancer mechanisms. IUBMB Life 2019, 71, 9–19. [Google Scholar] [CrossRef]
- del Prado-Audelo, M.L.; Cortés, H.; Caballero-Florán, I.H.; González-Torres, M.; Escutia-Guadarrama, L.; Bernal-Chávez, S.A.; Giraldo-Gomez, D.M.; Magaña, J.J.; Leyva-Gómez, G. Therapeutic applications of terpenes on inflammatory diseases. Front. Pharmacol. 2021, 12, 704197. [Google Scholar] [CrossRef]
- Araruna, M.E.; Serafim, C.; Júnior, E.A.; Hiruma-Lima, C.; Diniz, M.; Batista, L. Intestinal anti-inflammatory activity of terpenes in experimental models (2010–2020): A review. Molecules 2020, 25, 5430. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front. Endocrinol. 2023, 14, 1217165. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, X.; Sharma, G.; Dai, C. Thymol as a potential neuroprotective agent: Mechanisms, efficacy, and future prospects. J. Agric. Food Chem. 2024, 72, 6803–6814. [Google Scholar] [CrossRef] [PubMed]
- Begh, M.Z.A.; Khan, J.; Amin, M.A.; Sweilam, S.H.; Dharmamoorthy, G.; Gupta, J.K.; Sangeetha, J.; Nafady, M.H.; Ahmad, I.; Alshehri, M.A.; et al. Monoterpenoid synergy: A new frontier in biological applications. Naunyn Schmiedebergs Arch. Pharmacol. 2025, 398, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Vyhlídalová, B.; Ondrová, K.; Zůvalová, I. Dietary monoterpenoids and human health: Unlocking the potential for therapeutic use. Biochimie 2025, 228, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Shokoohizadeh, L.; Taheri, M. Natural Products in the treatment of diabetic foot infection. Eur. J. Med. Res. 2025, 30, 8. [Google Scholar] [CrossRef]
- El-Hack, M.E.A.; Alagawany, M.; Farag, M.R.; Tiwari, R.; Karthik, K.; Dharma, K.; Zorriehzahra, J.; Adel, M. Beneficial impacts of thymol essential oil on health and production of animals, fish and poultry: A review. J. Essent. Oil Res. 2016, 28, 365–382. [Google Scholar] [CrossRef]
- Huang, C.M.; Lee, T.T. Immunomodulatory effects of phytogenics in chickens and pigs—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Gholami-Ahangaran, M.; Ahmadi-Dastgerdi, A.; Azizi, S.; Basiratpour, A.; Zikaei, M.; Derakhshan, M. Thymol and carvacrol supplementation in poultry health and performance. Vet. Med. Sci. 2022, 8, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single components of botanicals and nature-identical compounds as a non-antibiotic strategy to ameliorate health status and improve performance in poultry and pigs. Nutr. Res. Rev. 2020, 33, 218–234. [Google Scholar] [CrossRef]
- Meeran, M.F.N.; Javed, H.; Taee, H.A.; Azimullah, S.; Ojha, S.K. Pharmacological properties and molecular mechanisms of thymol: Prospects for its therapeutic potential and pharmaceutical development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef]
- Pérez-Recalde, M.; Arias, I.E.R.; Hermida, E.B. Could essential oil enhance biopolymers performance for wound healing? A systematic review. Phytomedicine 2018, 38, 57–65. [Google Scholar] [CrossRef]
- Romo-Rico, J.; Krishna, S.M.; Bazaka, K.; Golledge, J. Potential of plant secondary metabolite-based polymers to enhance wound healing. Acta Biomater. 2022, 147, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Özen, I.; Demir, A.; Bahtiyari, M.I.; Wang, X.; Nilghaz, A.; Wu, P.; Shirvanimoghaddam, K.; Naebe, M. Multifaceted applications of thymol/carvacrol-containing polymeric fibrous structures. Adv. Ind. Eng. Polym. Res. 2024, 7, 182–200. [Google Scholar] [CrossRef]
- Sajed, H.; Sahebkar, A.; Iranshahi, M. Zataria multiflora Boiss. (Shirazi thyme)—An ancient condiment with modern pharmaceutical uses. J. Ethnopharmacol. 2013, 145, 686–698. [Google Scholar] [CrossRef] [PubMed]
- Khazdair, M.R.; Ghorani, V.; Alavinezhad, A.; Boskabady, M.H. Pharmacological effects of Zataria multiflora Boiss L. and its constituents focus on their anti-inflammatory, antioxidant, and immunomodulatory effects. Fund. Clin. Pharmacol. 2018, 32, 26–50. [Google Scholar] [CrossRef] [PubMed]
- Gou, Y.; Fan, R.; Pei, S.; Wang, Y. Before it disappeared: Ethnobotanical study of fleagrass (Adenosma buchneroides), a traditional aromatic plant used by the Akha people. Ethnobiol. Ethnomed. 2018, 14, 79. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Khaneghah, A.M.; Gavahian, M.; Marszałek, K.; Eş, I.; Munekata, P.E.S.; Ferreira, I.C.F.R.; Barba, F.J. Understanding the potential benefits of thyme and its derived products for food industry and consumer health: From extraction of value-added compounds to the evaluation of bioaccessibility, bioavailability, anti-inflammatory, and antimicrobial activities. Crit. Rev. Food Sci. Nutr. 2019, 59, 2879–2895. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, E.; Awoleye, O.; Davis, A.; Mishra, S. Anti-inflammatory and antimicrobial properties of thyme oil and its main constituents. Int. J. Mol. Sci. 2023, 24, 6936. [Google Scholar] [CrossRef]
- Kianmehr, M.; Khazdair, M.R.; Abbasnezhad, A.; Akram, M. Effects of Lamiaceae family plants and their bioactive ingredients on coronavirus-induced lung inflammation. Food Sci. Nutr. 2024, 12, 1528–1544. [Google Scholar] [CrossRef]
- Anwar, F.; Mahrye; Khan, R.; Qadir, R.; Saadi, S.; Gruczynska-Sekowska, E.; Saari, N.; Brishti, F.H. Exploring the biochemical and nutra-pharmaceutical prospects of some Thymus species—A review. Chem. Biodiver. 2024, 21, 500. [Google Scholar] [CrossRef]
- Waheed, M.; Hussain, M.B.; Saeed, F.; Afzaal, M.; Ahmed, A.; Irfan, R.; Akram, N.; Ahmed, F.; Hailu, G.G. Phytochemical profiling and therapeutic potential of thyme (Thymus spp.): A medicinal herb. Food Sci. Nutr. 2024, 12, 9893–9912. [Google Scholar] [CrossRef]
- Silva, A.S.; Rewari, D.; Sureda, A.; Suntar, I.; Belwal, T.; Battino, M.; Nabavi, S.M.; Nabavi, S.F. The evidence of health benefits and food applications of Thymus vulgaris L. Trends Food Sci. Technol. 2021, 117, 218–227. [Google Scholar] [CrossRef]
- Halat, D.H.; Krayem, M.; Khaled, S.; Younes, S. A focused insight into thyme: Biological, chemical, and therapeutic properties of an indigenous Mediterranean herb. Nutrients 2022, 14, 2014. [Google Scholar] [CrossRef] [PubMed]
- Nadi, A.; Shiravi, A.A.; Mohammadi, Z.; Aslani, A.; Zeinalian, M. Thymus vulgaris, a natural pharmacy against COVID-19: A molecular review. J. Herb. Med. 2023, 38, 100635. [Google Scholar] [CrossRef]
- Stojanović, N.M.; Randelović, P.J.; Simonović, M.; Radić, M.; Todorović, S.; Corrigan, M.; Harkin, A.; Boylan, F. Essential oil constituents as anti-inflammatory and neuroprotective agents: An insight through microglia modulation. Int. J. Mol. Sci. 2024, 25, 5168. [Google Scholar] [CrossRef]
- Kim, M.; Sowndhararajan, K.; Kim, S. The chemical composition and biological activities of essential oil from Korean native thyme Bak-Ri-Hyang (Thymus quinquecostatus Celak.). Molecules 2022, 27, 4251. [Google Scholar] [CrossRef] [PubMed]
- Saoulajan, C.; Boujida, N.; Mihyaoui, E.E.; Baakili, A.E.; Alshahrani, M.M.; Lee, L.-H.; Bouyahya, A. Phytochemistry, pharmacological investigations, industrial applications, and encapsulation of Thymbra capitata L. A review. Trends Food Sci. Technol. 2022, 129, 463–491. [Google Scholar] [CrossRef]
- Bautista, I.-B.; Aguilar, C.N.; Martínez-Ávida, G.C.G.; Torres-León, C.; Ilina, A.; Flores-Gallegos, A.C.; Verma, D.K.; Chávez-González, L.L. Mexican oregano (Lippia graveolens Kunth) as source of bioactive compounds: A review. Molecules 2021, 26, 5156. [Google Scholar] [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.-M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C.; et al. A recent insight regarding the phytochemistry and bioactivity of Origanum vulgare L. essential oil. Int. J. Mol. Sci. 2020, 21, 9653. [Google Scholar] [CrossRef]
- Aziz, A.A.E.; Elfadadny, A.; Ghanima, M.A.; Cavallini, D.; Fusaro, I.; Giammarco, M.; Buonaiuto, G.; El-Sabrout, K. Nutritional value of oregano-based products and its effect on rabbit performance and health. Animals 2024, 14, 3021. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, C.; Su, K.; Fan, T.; Chen, L.; Yang, Z.; Zhang, M.; Li, J.; Zhang, Y.; Liu, J. Oregano essential oil in livestock and veterinary medicine. Animals 2024, 14, 1532. [Google Scholar] [CrossRef]
- Słabęska, N.; Czajkowska, K.; Łstaś, P.; Śniarowki, P.; Ciosek, L.; Jastrzębski, K.; Pietrzyk, B. Sol-gel SiO2 coatings with curcumin and thymol on 3D printouts manufactured from Ti6Al4V ELI. Coatings 2024, 14, 1149. [Google Scholar] [CrossRef]
- Almeida, C.R.; Serra, T.; Oliveira, M.I.; Planell, J.A.; Barbosa, M.A.; Navarro, M. Impacto f 3-D printed PLA- and chitosan-based scaffolds on human monocyte/macrophage responses: Unraveling the effect of 3-D structures on inflammation. Acta Biomater. 2014, 10, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Periferakis, A.; Periferakis, A.-T.; Troumpata, L.; Dragosloveanu, S.; Timofticiuc, I.-A.; Georgatos-Garcia, S.; Scheau, A.-E.; Periferakis, K.; Caruntu, A.; Badarau, I.A.; et al. Use of biomaterials in 3D printing as a solution to microbial infections in arthroplasty and osseous reconstruction. Biomimetics 2024, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, R.; Beltrami, B.; Regondi, S.; Lunetta, C. Polymeric biomaterials for 3D printing in medicine: An overview. Ann. 3D Print. Med. 2021, 2, 100011. [Google Scholar] [CrossRef]
- Tappa, K.; Jammalamadaka, U. Novel biomaterials used in medical 3D printing techniques. J. Funct. Biomat. 2018, 9, 17. [Google Scholar] [CrossRef]
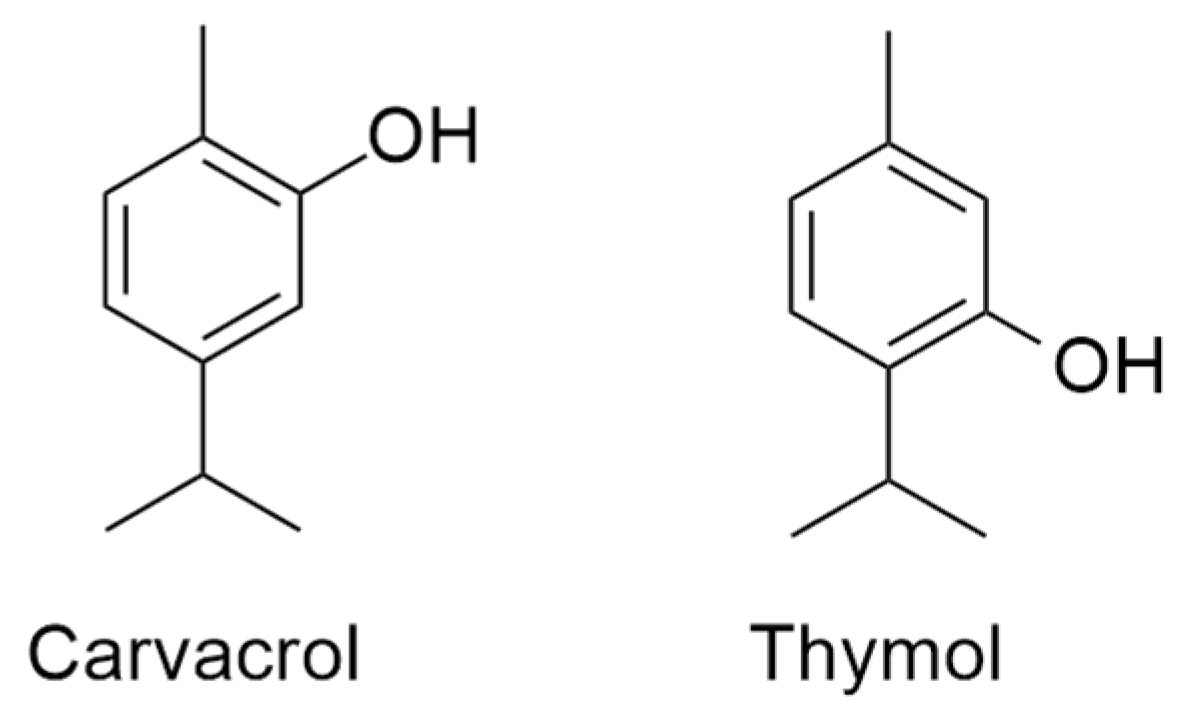

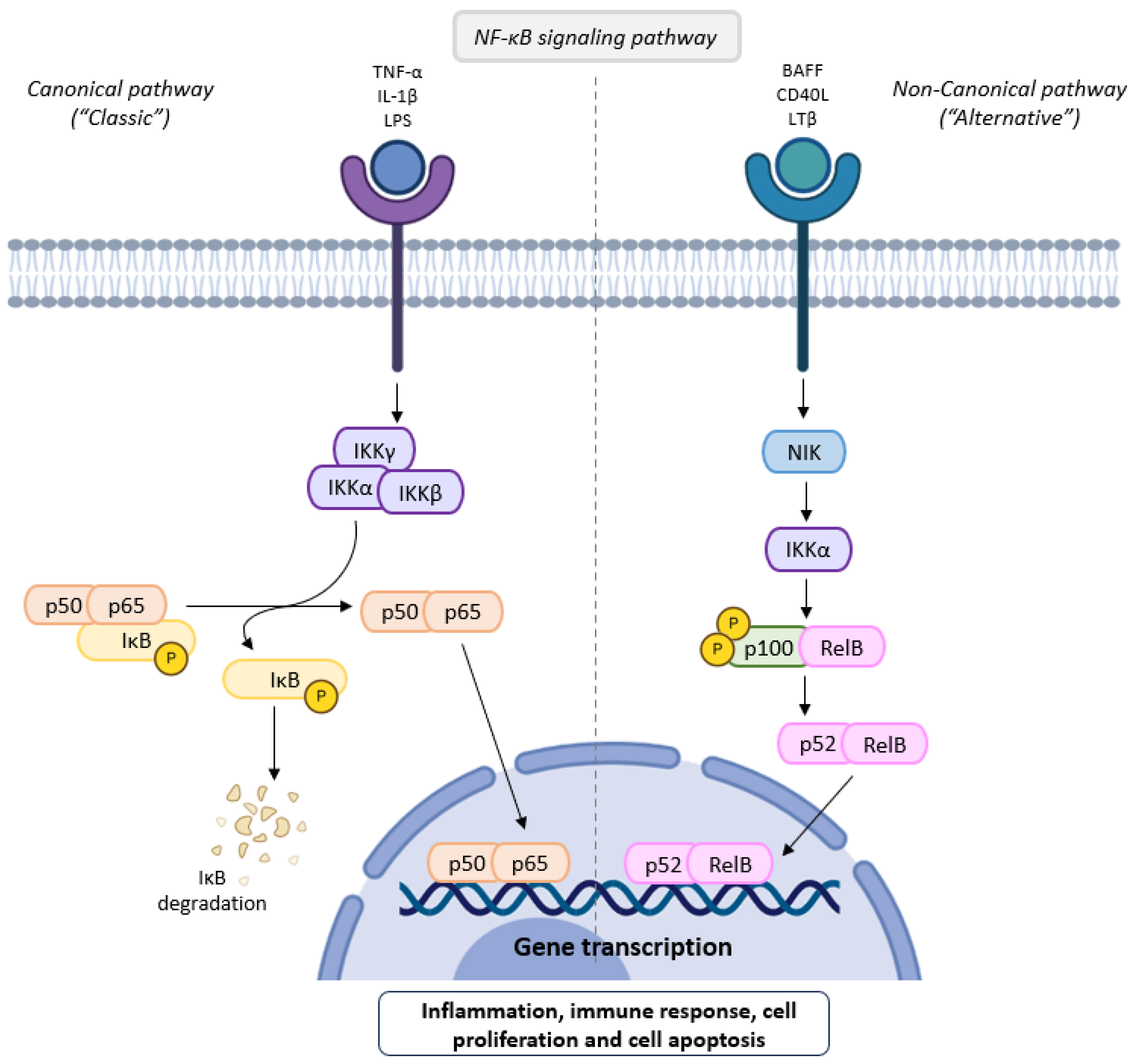
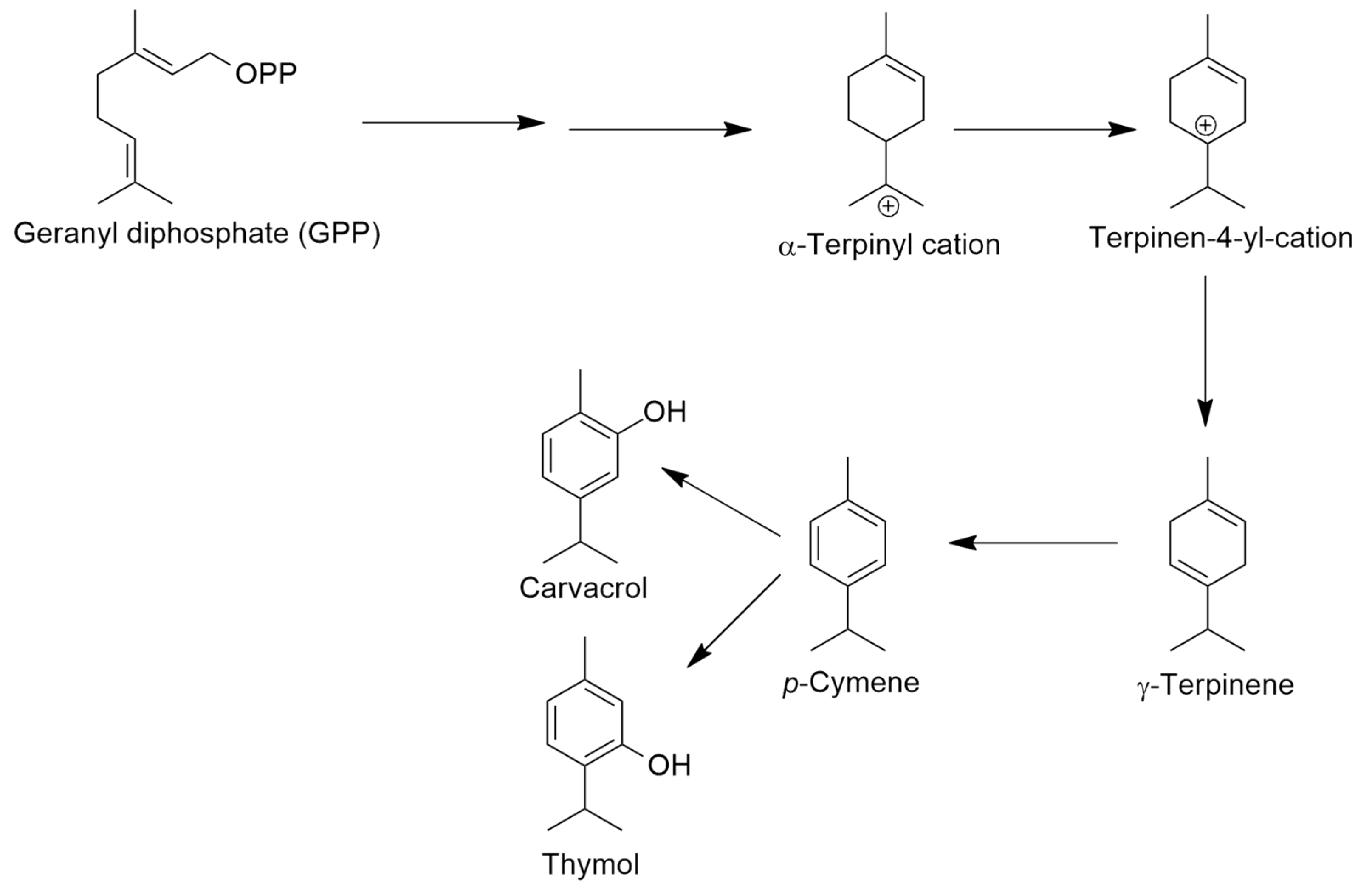
 inhibition;
inhibition;  stimulation.
inhibition; stimulation.
stimulation.
inhibition; stimulation.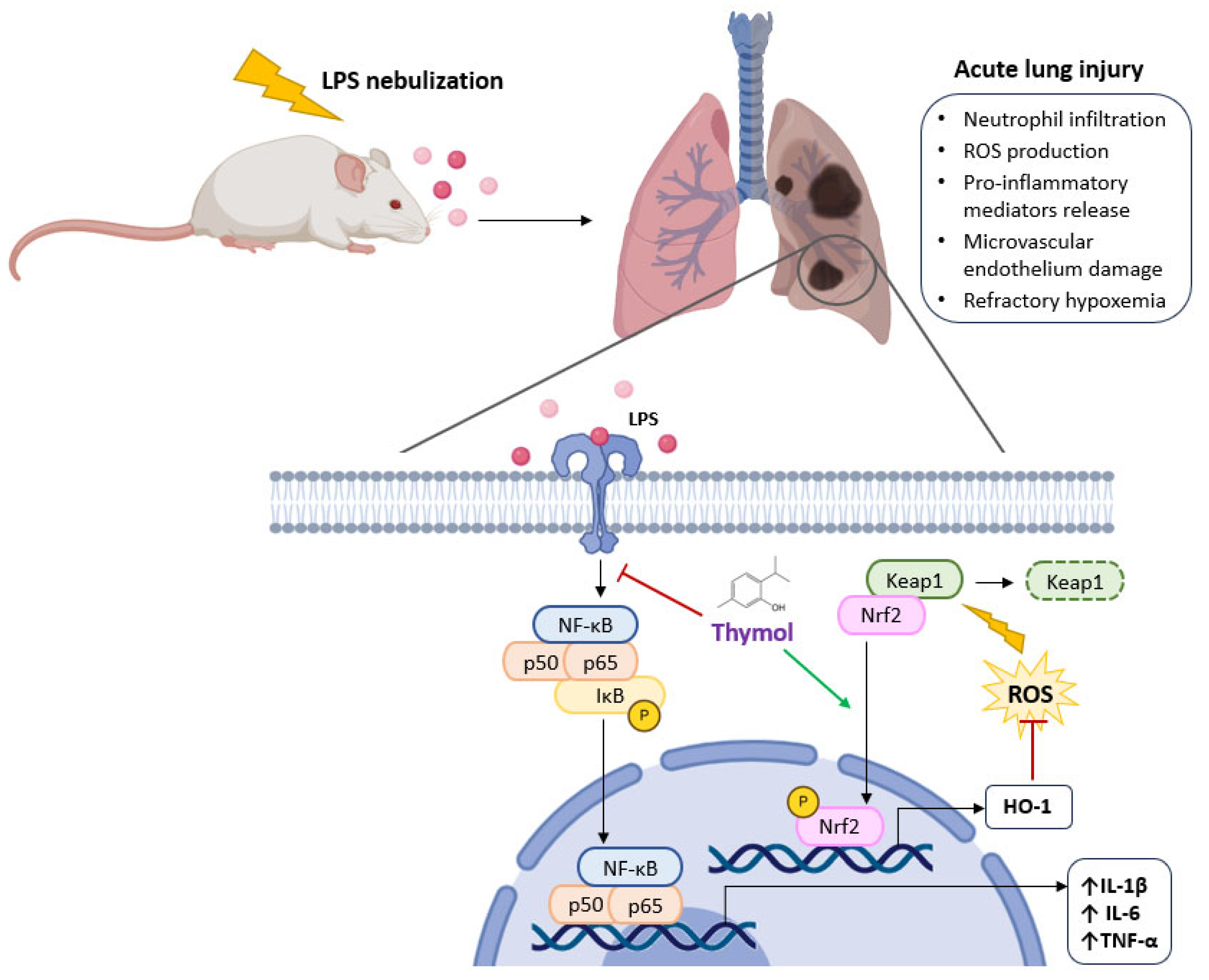
 inhibition;
inhibition;  potential stimulation.
inhibition; potential stimulation.
potential stimulation.
inhibition; potential stimulation.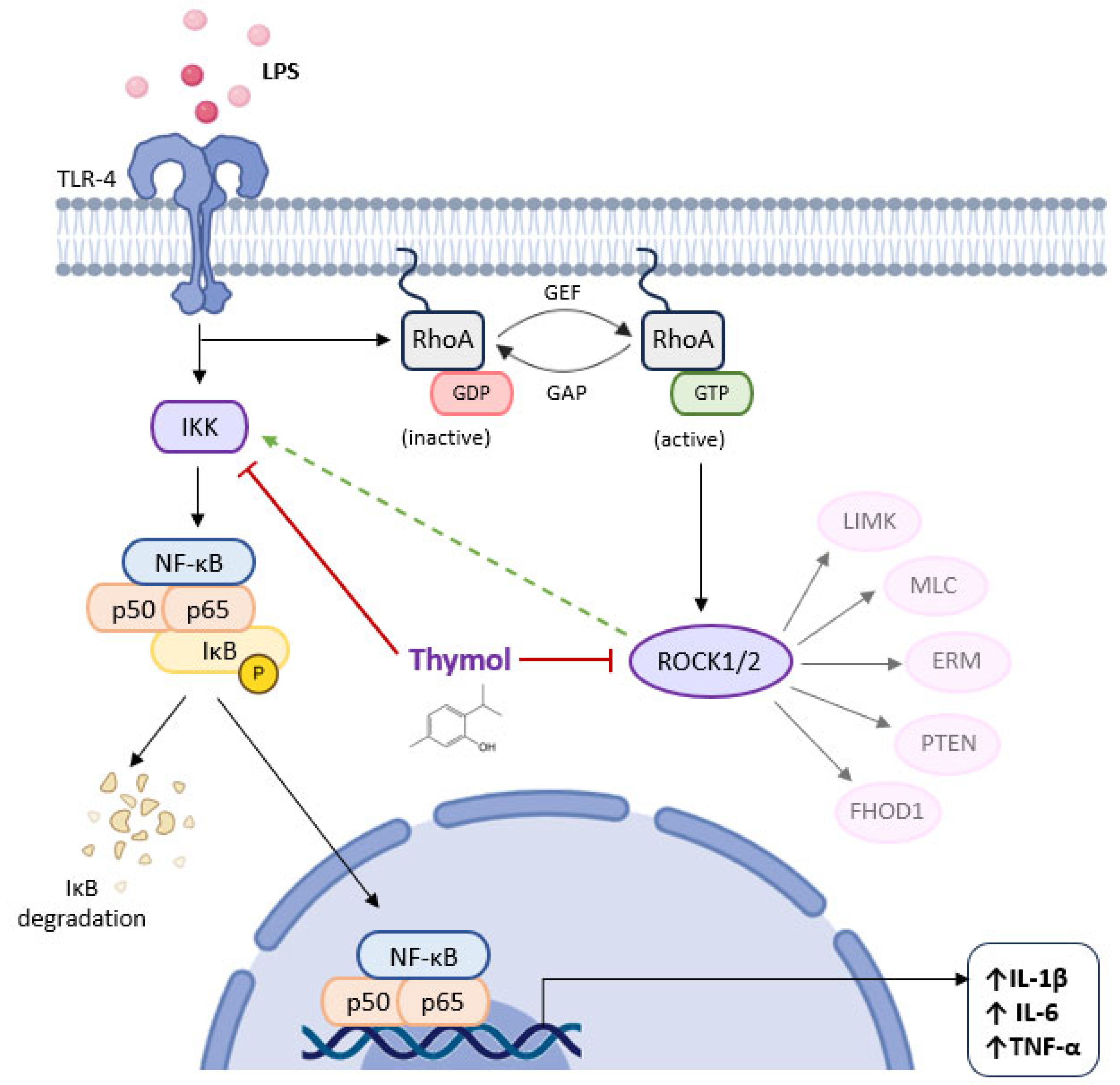
 inhibition.
inhibition.
inhibition.
inhibition.
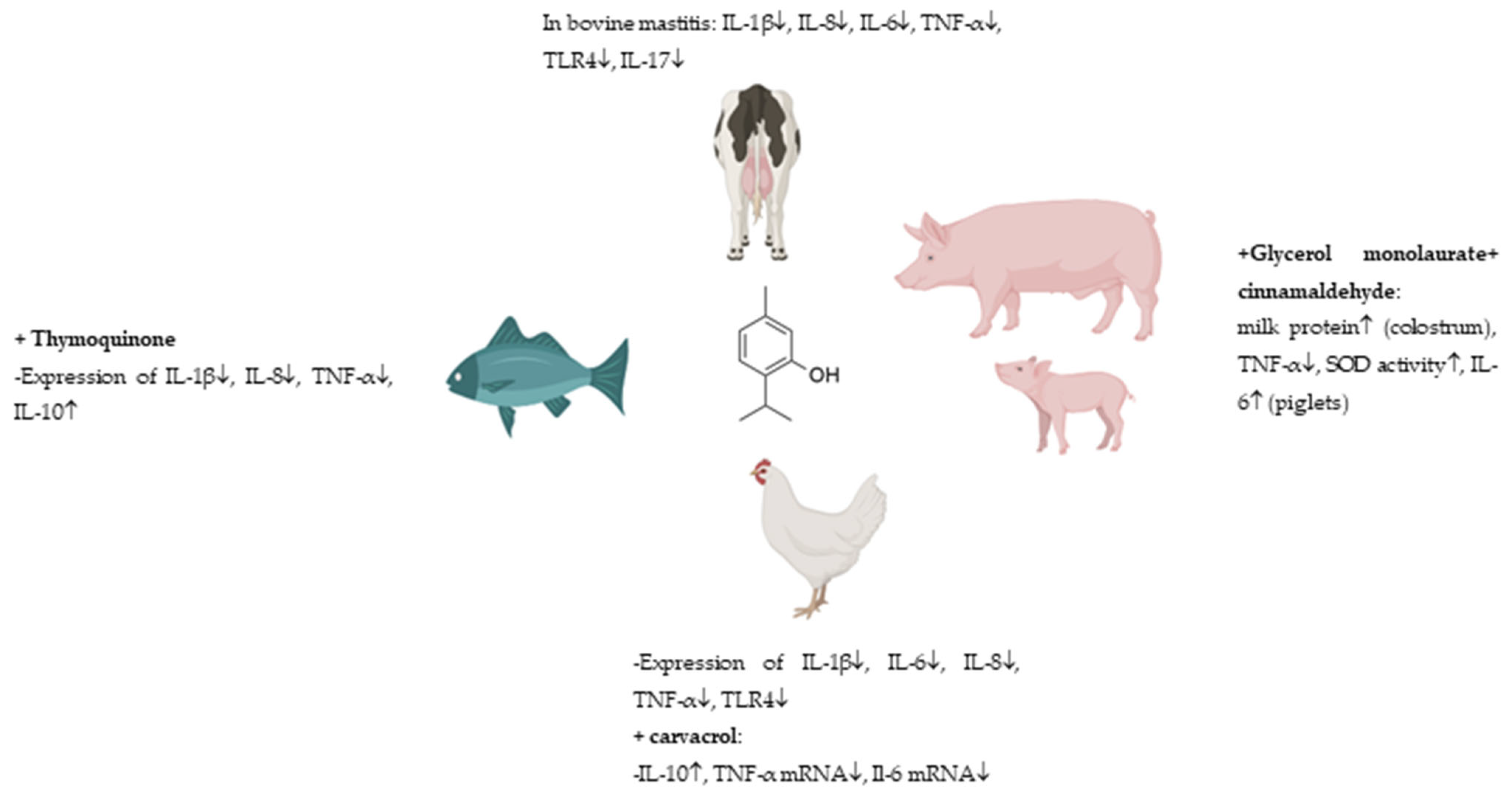
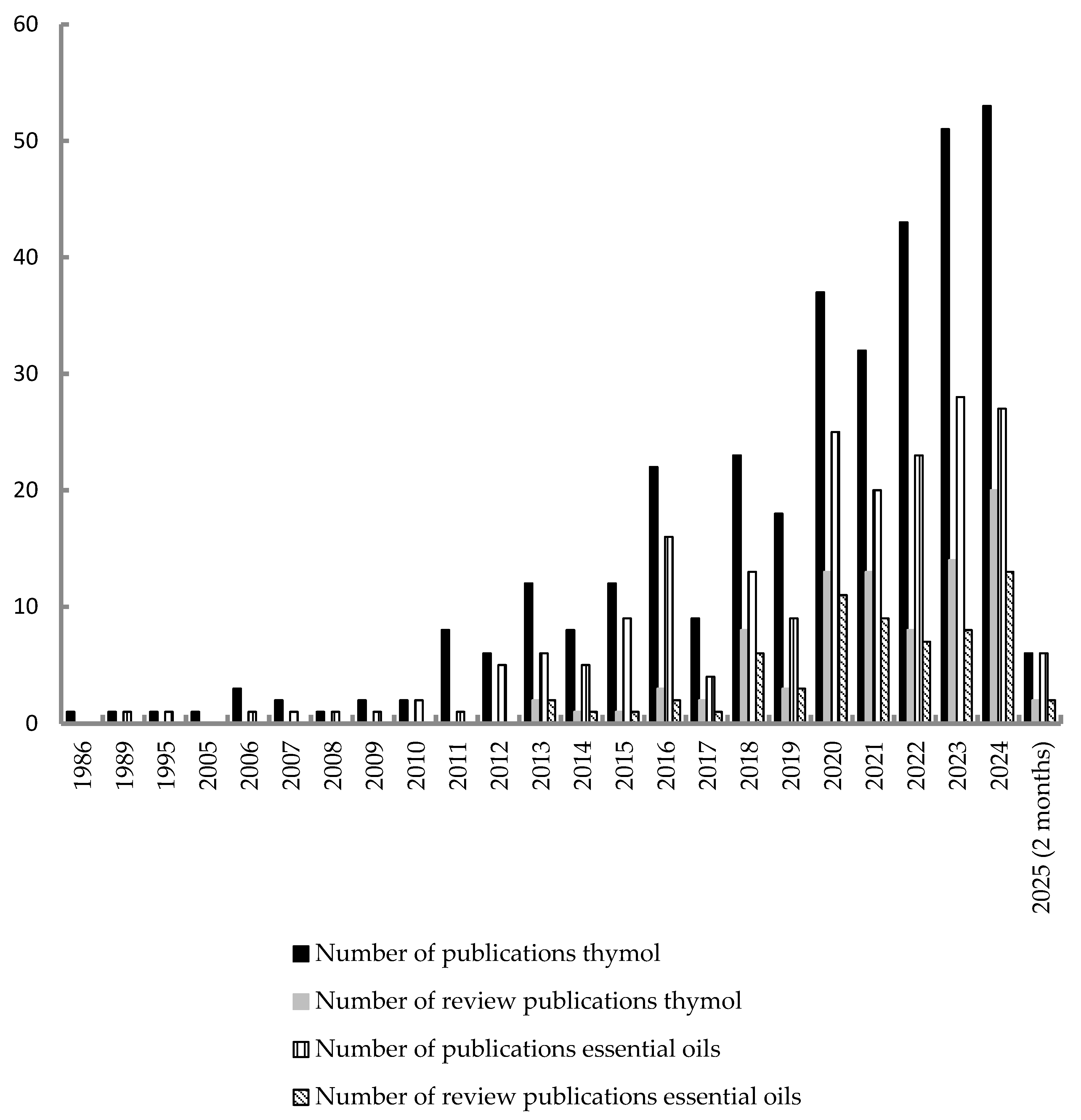
| Clade | Order | Family | Species | Reference |
|---|---|---|---|---|
| Angiosperms, Eudicots | Apiales | Apiaceae | Achillea millefolium | [6] |
| Angiosperms, Eudicots | Apiales | Apiaceae | Anethum graveolens | [6] |
| Angiosperms, Eudicots | Apiales | Apiaceae | Ferulago angulata | [7] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Ocimum basilicum | [8] |
| Angiosperms, Eudicots | Apiales | Apiaceae | Oliveria decumbens | [9] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Origanum floribundum | [10] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Origanum heracleoticum | [11,12] |
| Angiosperms, Eudicots | Celastrales | Celastraceae | Salacia pallescens | [13] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus atlanticus | [14] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus caespitius | [15] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus linearis | [16] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus sipyleus | [17] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus vulgaris | [18,19,20,21,22] |
| Angiosperms, Eudicots | Lamiales | Lamiaceae | Thymus zygis | [23,24] |
| Angiosperms, Eudicots | Apiales | Apiaceae | Trachyspermum ammi | [6,25,26,27,28] |
| Angiosperms, Eudicots | Apiales | Apiaceae | Trachyspermum roxburghianum | [29] |
| Species (Origin) | Plant Part | % of Thymol and Carvacrol and Main Compound (>5%) | Method | Values | References |
|---|---|---|---|---|---|
| Achillea millefolium Ilam, Iran Anethum graveolens Ilam, Iran Carum copticum (=Trachyspermum ammi) Ilam, Iran | Aerial parts Seeds Seeds | Thymol 26, carvacrol 10, borneol 16, limonene 15, α-pinene 10 Thymol 20, carvacrol 8, limonene 16, α-pinene 9 Thymol 23, carvacrol 6, sabinene 18, borneol 10 | Inhibition of nitric oxide (NO) production in in vitro model of lipopolysaccharide (LPS)-stimulated RAW-264.7 macrophages Inhibition of NO production in in vitro model of LPS-stimulated RAW-264.7 macrophages Inhibition of NO production in in vitro model of LPS-stimulated RAW-264.7 macrophages | Production of nitrites 15.04 µM; LPS (33.98 µM) Production of nitrites 41.04 µM; LPS (33.98 µM) Production of nitrites 26.02 µM; LPS (33.98 µM) | [6] |
| Carum copticum (=Trachyspermum ammi) Unknown | Seeds | Thymol 46, carvacrol 3, γ-terpinene 20, p-cymene 11, β-pinene 6 | Inhibition of NO production in in vitro model of LPS-stimulated RAW-264.7 macrophages | Essential oil: % NO inhibition (80% at 45.0 µg/mL) p-cymene: % NO inhibition (48.05% at 45.0 µg/mL) γ-terpinene: % NO inhibition (60.20% at 45.0 µg/mL) β-pinene: % NO inhibition (45.20% at 45.0 µg/mL) thymol: % NO inhibition (85.12% at 45.0 µg/mL) | [25] |
| Ferulago angulata Yasouj, Iran | Aerial parts | Thymol 8, carvacrol 3, spathulenol 7, trans-anetole 6, p-menth-2-en-1-ol 5, myristicin 5 | In vivo anti-inflammatory activity through the croton oil-induced male Swiss mice (20–30 g) ear edema | Both 200 and 100 μL/kg of essential oil produced 66% inhibition of ear edema. Indomethacin (positive control): 82% inhibition at 10 mg/kg) | [7] |
| Ocimum basilicum/Kairouan, Tunisia | Seeds | Solid-liquid Soxhlet extraction using n-hexane: Concentration (µg/g): Thymol 33, luteolin 9 2-Methyltetrahydro-furan (MeTHF) Concentration (µg/g): Thymol 128, rosmarinic acid 21, luteolin 9, gallic acid 7, chlorogenic acid 7, ellagic acid 6, circimaritin 6 | Inhibition of NO production in LPS-induced murine macrophage RAW 264.7 cell line | At 150 µg/mL, NO production decreased by 60% At 150 µg/mL, NO production decreased by 64% | [8] |
| Oliveria decumbens/Dil village, Iran | Aerial parts | Extraction with ethanol:H2O (70:30) and then fractionation: n-hexane: thymol 56, carvacrol 38 dichloromethane: thymol 53, carvacrol 41 | Oxidative burst assay using luminol-enhanced chemiluminescence technique with blood HBSS++ (Hanks Balanced Salt Solution), serum opsonized zymosan (SOZ) and intracellular reactive oxygen species | Crude extract: *IC50 = 21.7 µg/mL n-Hexane: IC50 = 22.6 µg/mL Dichloromethane: IC50 = 15.8 µg/mL Positive control (Ibuprofen): IC50 = 11.2 µg/mL Thymol: not active (>100 µg/mL) Carvacrol: not active (>100 µg/mL) | [9] |
| Origanum floribundum/ Lakhdaria, Algeria | Aerial parts at flowering stage (July 2011) | Thymol 34, carvacrol 9, γ-terpinene 20, p-cymene 16, | In vitro spectrophotometrically lipoxygenase inhibition | IC50: 125.7 µg/mL **NDGA (positive control): IC50 = 63.4 µg/mL | [10] |
| Origanum heracleoticum (5 samples) /Agrigento, Italy | Leaves and flowers | Thymol 47–65, carvacrol 3–5, γ-terpinene 13–22, p-cymene 4–5, carvacrol methyl ether 3–4 | Gene expression analysis of ***NF-κB pathway in Caco-2 cells treated with ****TNF-α | An increase trend of *****IL-1α, IL-6 and IL-8 gene expression was observed when compared to the treatment with TNF-α, without statistical significance It is advisable to read the article, data presented as graphics | [11] |
| Origanum heracleoticum (6 samples)/Calabria, Italy | Not identified plant part used | Thymol 9104–54,459, carvacrol 3311–68,379, o-cymene 3304–48,176, γ-terpinene 24–19,133, carvacrol-methyl ether 2658–19,844, β-caryophyllene 1182–16,107, γ-muurolene 1364–10,824 (unknown unities, the compounds were chosen if at least one value is >10,000 | Inhibition of NO production in lipopolysaccharide (LPS)-induced murine macrophage RAW 264.7 cell line | IC50 = 32.77–170.9 μg/mL. Positive controls (indomethacin and L-NAME): IC50 = 58.00 and 45.86, respectively) | [12] |
| Salacia pallescens/Idi-Ayunre, Ibadan, Nigeria | Leaves | Extraction with methanol 50%: Thymol 30, 3-carene 16, p-cymene 12, caffeine 8, hexadecanoic acid 6, bicycle[3.1.1]hept-2-ene, 2,6-dimethyl-6-(4-methyl-3-pentenyl) 6, caryophyllene 5 | In vitro inhibition of nitrite oxide (NO) production Inhibition of interleukin 6 (IL-6) production in in vitro model of LPS-stimulated RAW-264.7 macrophages | IC50 = 49.49 µg/mL IC50 = 48.74 µg/mL (positive control, ascorbic acid) Nine-fold reduction in LPS induced IL-6 production in RAW-264.7 macrophages pre-treated with 400 µg/mL of the extract. 1.4-fold reduction in LPS induced IL-6 production in RAW-264.7 macrophages pre-treated with 50 µg/mL of the extract | [13] |
| Siddhalepa Asamodagam Spirit (water distillate derived from Trachyspermum roxburghianum seeds/Pakistan | Seeds | The water distillate was then extracted with ethyl acetate: Thymol 93, carvacrol 1 | Heat induced hemolysis principle using human red blood cell stabilization method | IC50 = 0.57 mg/mL Positive control (aspirin): IC50 = 0.24 mg/mL | [29] |
| Thymus atlanticus/Errachidia region, Morocco | Aerial parts | Thymol 24, carvacrol 23, γ-terpinene 21, p-cymene 19 | In vivo phenol induced ear edema in Wister albino rats of both sexes weighing 100–120 g | The application of phenol alone has developed in the control group an ear inflammation of 77.59% The topical application of essential oil (1 mg/ear) has reduced the ear edema with a percentage of 19.39%. Indomethacin (positive control) (1 mg/ear) has reduced the ear edema with a percentage of 14.71% | [14] |
| Thymus caespitius/Azores, Portugal | Plant material was collected during the flowering phase | Thymol 34, carvacrol 11, p-cymene 12, thymol acetate 8, γ-terpinene 5 | NO scavenging activity In vitro spectrophotometrically lipoxygenase inhibition | IC50 = 0.3 mg/mL IC50 = 0.1 mg/mL | [15] |
| Thymus linearis/Harinagar and Dhanachuli, India | Aerial parts | Thymol 67. carvacrol 3, p-cymene 10 | In vivo anti-inflammatory activity through the carrageenan-induced paw edema of Swiss albino mice In vivo sub-acute anti-inflammatory activity through the formaldehyde induced arthritis in the right hind paw of Swiss albino mice (formaldehyde solution 1% injected on the first day of the experiment. Samples administered orally every day of the experiment. The results were recorded every day till the end (10 days). For knowing all results it is advisable to read the reference | At 5%: Inhibition % after 4 h and 24 h = 3.83 and 6.38, respectively At 10%: Inhibition % after 4 h and 24 h = 5.19 and 9.52, respectively At 20%: Inhibition % after 4 h and 24 h = 9.73 and 14.6, respectively Positive control (Ibuprofen): At 0.004%: Inhibition % after 4 h and 24 h = 26.07 and 37.18, respectively Volume of inflammation (mm3) (5%): day 0, day 5, day 10 = 2.26, 2.47, 2.51 Volume of inflammation (mm3) (10%): day 0, day 5, day 10 = 2.21, 2.38, 2.38 Volume of inflammation (mm3) (20%): day 0, day 5, day 10 = 2.19, 2.32, 2.26 Volume of inflammation (mm3) saline water (control): day 0, day 5, day 10 = 2.13, 2.39, 2.37 Volume of inflammation (mm3) positive control (Ibuprofen) (0.001%): day 0, day 5, day 10 = 2.11, 2.19, 2.15 | [16] |
| Thymus sipyleus Boiss. subsp. sipyleus var. sipyleus/Sivas, Turkey | Aerial parts | Thymol 66, carvacrol 3, p-cymene 9, γ-terpinene 9 | In vitro spectrophotometrically lipoxygenase inhibition | 12% inhibition for a concentration of 100 μg/mL. NDGA (positive control) 100% inhibition for a concentration of 100 μg/mL | [17] |
| Thymus vulgaris/Szigetvár city, Hungary | Not identified the part of plant used | Beginning of flowering: Thymol 56, carvacrol 2, p-cymene 13, γ-terpinene 15 End of flowering: Thymol 54, carvacrol 3, p-cymene 21, γ-terpinene 6 | Inhibition of cytokine production through P. aeruginosa LPS-activated THP-1 macrophage cells | Inhibition of the production of IL-6, IL-8, IL-1β, TNF-α. Thymol (positive control) had better activity These results are based on the figures’ observation. It is advisable to read the article There is not a decrease of the four examined proinflammatory cytokine IL-6, IL-8, IL-β, TNF-α expression both at the mRNA and protein levels These results are based on the figures’ observation. It is advisable to read the article | [18] |
| Thymus vulgaris (purchased) | Not reported | Thymol 49, carvacrol 3, p-cymene 29 | Aging-related and pro-inflammatory cytokine gene expression in the liver, hippocampus, cerebellum, and cerebral cortex of mice fed with the thyme EO 250 mg/kg/day (0.2% (w/w) Aging-related and pro-inflammatory cytokine gene expression in age-accelerated NIH-3T3 cells. Thyme oil concentrations: 30, 60, 120 µg/mL | Lower levels of p16INK4A in the hippocampus than control; lower expression of Il-1b in the liver and cerebellum, and Il6 in the hippocampus than the control; lower expression of p65 and p50 (two transcription factors of NF-κB) than the control Lower levels of Il6 and Ccl2 mRNA expression (dose-dependent) It is advisable to read the article since the results are presented under graphics | [19] |
| Thymus vulgaris/Póvoa de Lanhoso, Portugal | Aerial parts | Thymol 41, carvacrol 5, o-cymene 25, p-cymene 14 | Inhibition of NO production in in vitro model of LPS-stimulated RAW-264.7 macrophages | IC50 = 8 µg/mL Dexamethasone (positive control): IC50 = 16 µM | [20] |
| Thymus vulgaris (diplod)/Prague, Czech Republic Thymus vulgaris (tetraploid)/Prague, Czech Republic | Aerial parts Aerial parts | Thymol 51, p-cymene 20, γ-terpinene 6 Thymol 54, γ-terpinene 22, p-cymene 8, α-cadinol 8, caryophyllene 6 | In vitro inhibitory activity against ******COX-2 In vitro inhibitory activity against COX-2 | Inhibition percentage: 80.96 (500 µg/mL) 70.53 (50 µg/mL) 2.02 (5 µg/mL) Inhibition percentage: 85.57 (500 µg/mL) 83.74 (50 µg/mL) 6.74 (5 µg/mL) Ibuprofen (positive control): 74.52 (5 µg/mL) | [21] |
| Thymus vulgaris/Tlemcen, Algeria Thymus vulgaris/Mostaganem, Algeria | Aerial parts Aerial parts | Thymol 67, carvacrol < 0.05, γ-terpinene 10, p-cymene 6 Thymol 60, carvacrol < 0.05, γ-terpinene 9, α-pinene 6, p-cymene 6, linalool 5 | In vivo anti-inflammatory activity through the carrageenan-induced paw edema of Swiss albino mice of both sexes (weighing 25–30 g) In vivo anti-inflammatory activity through the carrageenan-induced paw edema of Swiss albino mice of both sexes (weighing 25–30 g) | Paw thickness (mm): (100 mg/kg–400 mg/kg, between 1–6 h): 2.76–2.36, respectively Paw thickness (mm): (100 mg/kg–400 mg/kg, between 1–6 h): 2.83–2.54, respectively Paw thickness (mm): Diclofenac (positive control) (10 mg/kg, between 1 h–6 h): 2.84–2.07, respectively Paw thickness (mm): Vehicle Tween 80 (Control) (10 mg/kg, between 1 h–6 h): 2.86–3.09, respectively | [22] |
| Thymus zygis chemotype thymol (4 samples)/Murcia, Spain Thymus zygis chemotype linalool (2 samples)/Murcia, Spain | Not identified the plant part used Not identified the plant part used | Thymol 30–54, carvacrol 0.4–3, p-cymene 14–27, γ-terpinene 8–28, linalool 0.1–5 Thymol 0.1–1, carvacrol 0.4–3, linalool 41–43, myrcene 7–8, terpinen- 4-ol 13–13, γ-terpinene 6–8, | In vitro spectrophotometrically lipoxygenase inhibition In vitro spectrophotometrically lipoxygenase inhibition | IC50 (μL/L): 54–73 IC50 (μL/L): 299–402 | [23] |
| Thymus zygis subsp. sylvestris/Parque Natural das Serras de Aire e Candeeiros, Portugal | Aerial parts -collected in flowering stage | Thymol 20, carvacrol 16, p-cymene 22, γ-terpinene 7, linalool 6 | Production of NO in in vitro model of LPS-stimulated RAW-264.7 macrophages Production of NO in in vitro model of LPS-stimulated in microglia | Production of NO in the presence of four concentrations of the essential oil: 42.67 (0.64 μL/mL), 62.67 (0.32 μL/mL), 91.67 0.16 μL/mL) and 88.00 (0.08 μL/mL Thymol, carvacrol had stronger activity than p-cymene and the essential oil (without numeric results, only observed from te figure), particularly in the concentrations ranging from 0.08 to 0.32 μL/mL Production of NO in the presence of four concentrations of the essential oil: 22.00 (0.64 μL/mL), 24.34 (0.32 μL/mL), 26.71 (0.16 μL/mL), 38.94 (0.08 μL/mL) The inhibitory profile triggered by the main compounds was more pronounced in microglia relative to macrophages. All the concentrations of p-cymene, thymol and carvacrol tested decreased NO production, quite similar to those obtained in the presence of the essential oil (only based in the observation of the figure | [24] |
| Trachyspermum ammi/Tiruvannamalai, Tamil Nadu, India | Seeds | Unprocessed: extraction with ethanol 90% Thymol 42, 3-methoxybutyric acid 5, 9,12-octadecadienoic acid (Z,Z) 6 Processed: lime treatment followed by extraction with ethanol 90% Thymol 34, 9,12-octadecadienoic acid (Z,Z) 42, | In vitro inhibitory activity through the protein (egg albumin) denaturation method | Inhibition percentage (1000 µg/mL): 74 Positive control (dichlofenac) (1000 µg/mL): 80 Inhibition percentage (1000 µg/mL): 78 Positive control (dichlofenac) (1000 µg/mL): 80 | [26] |
| Trachyspermum ammi/Jorhat, Assam, India | Seeds | Thymol 50, carvacrol 0.1, γ-terpinene 25, p-cymene 22 | In vitro inhibitory activity through the protein (bovine serum albumin, BSA) denaturation method | IC50 = 93.12 µL/mL IC50 = 108.61 µL/mL (positive control, diclofenac) | [27] |
| Trachyspermum ammi/Salem, Tamil Nadu, India | Seeds | Not reported, discussion made by the authors is based on the chemical composition of the essential oil of this species found in other articles. According to the authors, the main components of the essential oils of Trachyspermum ammi are thymol and carvacrol | In vitro COX-2 inhibition assay Production of nitrite oxide (NO) in in vitro model of LPS-stimulated RAW-264.7 macrophages Production of prostaglandin E2 (PGE2) in in vitro model of LPS-stimulated RAW-264.7 macrophages | IC50 = 4.49 µg/mL, IC50 = 1 µM (thymol) IC50 = 0.8 µM (carvcrol) NO production: 82.54 µM (0.5 µg/mL) 73.94 µM (1.0 µg/mL) 99.64 µM (without essential oil) (PGE2) production: 699.89 pg/mL (0.5 µg/mL) 585.56 pg/mL (1 µg/mL) 776.21 pg/mL (without essential oil) | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gago, C.; Serralheiro, A.; Miguel, M.d.G. Anti-Inflammatory Activity of Thymol and Thymol-Rich Essential Oils: Mechanisms, Applications, and Recent Findings. Molecules 2025, 30, 2450. https://doi.org/10.3390/molecules30112450
Gago C, Serralheiro A, Miguel MdG. Anti-Inflammatory Activity of Thymol and Thymol-Rich Essential Oils: Mechanisms, Applications, and Recent Findings. Molecules. 2025; 30(11):2450. https://doi.org/10.3390/molecules30112450
Chicago/Turabian StyleGago, Custódia, Ana Serralheiro, and Maria da Graça Miguel. 2025. "Anti-Inflammatory Activity of Thymol and Thymol-Rich Essential Oils: Mechanisms, Applications, and Recent Findings" Molecules 30, no. 11: 2450. https://doi.org/10.3390/molecules30112450
APA StyleGago, C., Serralheiro, A., & Miguel, M. d. G. (2025). Anti-Inflammatory Activity of Thymol and Thymol-Rich Essential Oils: Mechanisms, Applications, and Recent Findings. Molecules, 30(11), 2450. https://doi.org/10.3390/molecules30112450