


Bioassay-Guided Isolation and Active Compounds Identification of the AntiDiabetic Fractions of Centaurea calcitrapa Extract and the Predicted Interaction Mechanism


 ,
,
Abstract

1. Introduction
2. Results
2.1. Centaurea calcitrapa Extracts Ameliorate Renal, Hepatic, and Oxidative Stress Changes Induced by Streptozotocin
2.2. Antidiabetic Properties of C. calcitrapa Extracts on Diabetic Rats Induced by Streptozotocin
2.3. Antidiabetic Properties of E-2 Subfractions on Diabetic Rats Induced by Streptozotocin
2.4. Purification of E2-VIII Subfraction by HPLC
2.5. Calibration of Nepetin, Kaempferide, and Luteolin Compounds by HPLC
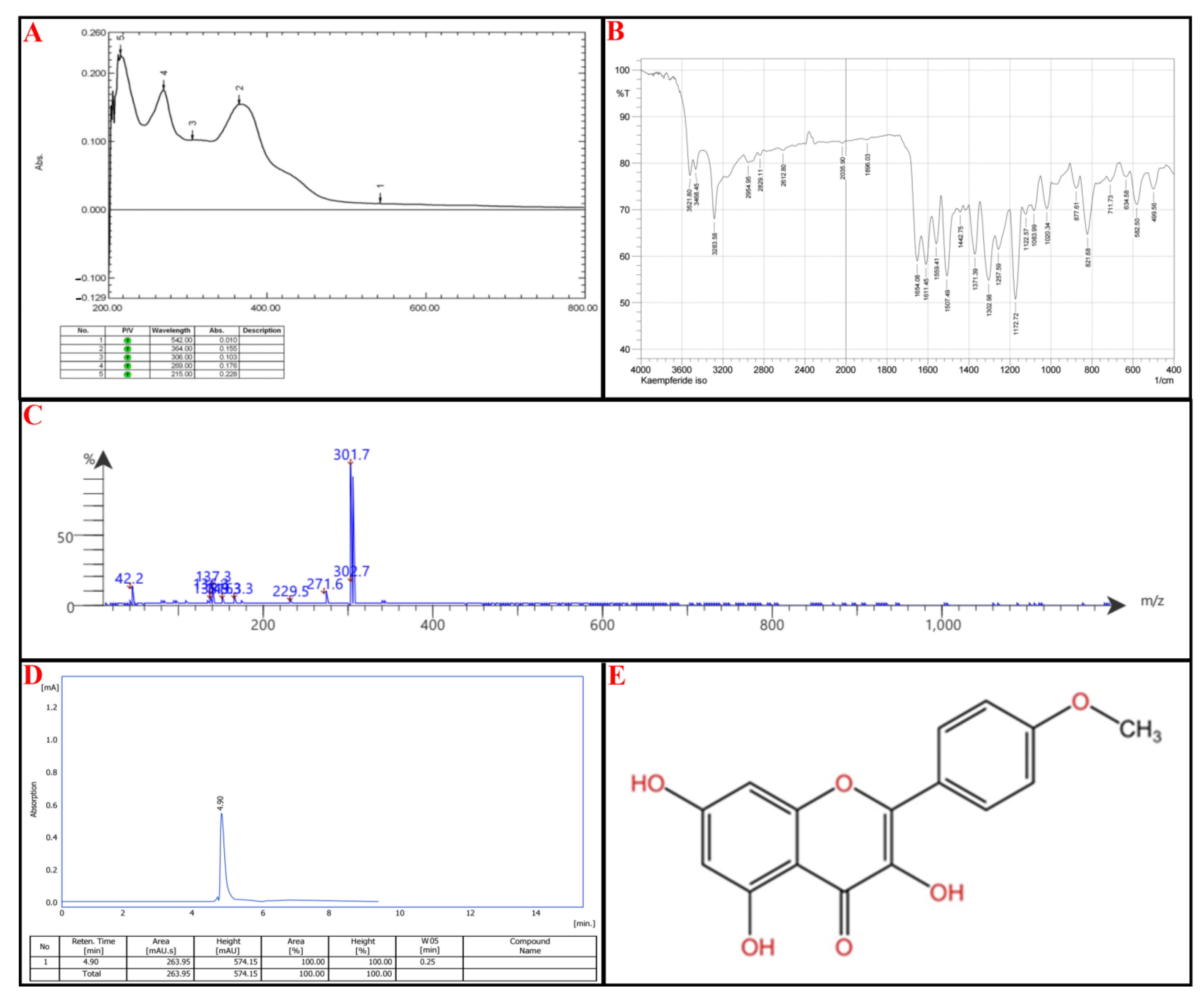
2.6. Structural Analysis of E2-VIII Subfractions
2.7. Molecular Docking
2.8. In Vitro α-Amylase Inhibitory Activity of Nepetin
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents
4.2. Plant Materials
4.3. Extraction
4.4. Column Chromatography
4.5. Purification of E2-VIII Subfraction by High-Performance Liquid Chromatography (HPLC)
4.6. Identification of the Isolated Compounds
4.6.1. UV Spectrometry
4.6.2. Fourier-Transform Infrared Spectroscopy (FTIR)
4.6.3. Mass Spectrophotometry (MS)
4.6.4. Calibration Curve
4.7. In Vitro α-Amylase Inhibitory Assay
4.8. Experimental Design and Settings
4.8.1. Diabetic Induction
4.8.2. Special Considerations to Minimize the Suffering and Distress of Animals
4.8.3. Plant Extracts Experimental Design
4.8.4. C. calcitrapa Ethyl Acetate Extract (E2) Fractions Experimental Design
4.9. Clinical and Laboratory Assessment
4.9.1. Body Weight Monitoring
4.9.2. Monitoring the Blood Glucose Level
4.9.3. Biochemical Analysis (Conducted on Stage 1)
4.10. In Silico Molecular Docking Studies
4.11. Ethical Considerations
4.12. Statistical Analysis and Sample Size Calculation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Fattaheian-Dehkordi, S.; Hojjatifard, R.; Saeedi, M.; Khanavi, M. A Review on Antidiabetic Activity of Centaurea spp.: A New Approach for Developing Herbal Remedies. Evid.-Based Complement. Altern. Med. 2021, 2021, 5587938. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Numan, A.T.; Jawad, N.K.; Fawzi, H.A. Biochemical study of the effect of lead exposure in nonobese gasoline station workers and risk of hyperglycemia: A retrospective case-control study. Medicine 2024, 103, e39152. [Google Scholar] [CrossRef] [PubMed]
- Numan, A.T.; Jawad, N.K.; Fawzi, H.A. Biochemical study of the risk of diabetes, prediabetic and insulin resistance in car painters and its association with mercury exposure: A retrospective case-control study. Toxicol. Res. 2024, 13, tfae221. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Cerf, M.E. Beta cell dysfunction and insulin resistance. Front. Endocrinol. 2013, 4, 37. [Google Scholar] [CrossRef]
- Roca-Rivada, A.; Castelao, C.; Senin, L.L.; Landrove, M.O.; Baltar, J.; Belén Crujeiras, A.; Seoane, L.M.; Casanueva, F.F.; Pardo, M. FNDC5/irisin is not only a myokine but also an adipokine. PLoS ONE 2013, 8, e60563. [Google Scholar] [CrossRef]
- Dali-Youcef, N.; Mecili, M.; Ricci, R.; Andrès, E. Metabolic inflammation: Connecting obesity and insulin resistance. Ann. Med. 2013, 45, 242–253. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Jansen, C.; Baker, J.D.; Kodaira, E.; Ang, L.; Bacani, A.J.; Aldan, J.T.; Shimoda, L.M.N.; Salameh, M.; Small-Howard, A.L.; Stokes, A.J.; et al. Medicine in motion: Opportunities, challenges and data analytics-based solutions for traditional medicine integration into western medical practice. J. Ethnopharmacol. 2021, 267, 113477. [Google Scholar] [CrossRef]
- Dimkić, I.; Petrović, M.; Gavrilović, M.; Gašić, U.; Ristivojević, P.; Stanković, S.; Janaćković, P. New perspectives of purple starthistle (Centaurea calcitrapa) leaf extracts: Phytochemical analysis, cytotoxicity and antimicrobial activity. AMB Express 2020, 10, 183. [Google Scholar] [CrossRef]
- Al-Joboury, K.R.; Aliwy, S.A. Survey with revised checklist of compositae in the herbarium of iraq natural history research center and museum. Bull. Iraq Nat. Hist. Mus. 2023, 17, 375–407. [Google Scholar] [CrossRef]
- Khammar, A.; Djeddi, S. Pharmacological and biological properties of some Centaurea species. Eur. J. Sci. Res. 2012, 84, 398–416. [Google Scholar]
- Pieroni, A.; Janiak, V.; Dürr, C.M.; Lüdeke, S.; Trachsel, E.; Heinrich, M. In vitro antioxidant activity of non-cultivated vegetables of ethnic Albanians in southern Italy. Phytother. Res. 2002, 16, 467–473. [Google Scholar] [CrossRef]
- Lentini, F. The role of ethnobotanics in scientific research. State of ethnobotanical knowledge in Sicily. Fitoterapia 2000, 71 (Suppl. 1), S83–S88. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Dumlu, M.U.; Gürkan, E. A new active compound from Centaurea species. Z. Naturforsch C 2006, 61, 44–46. [Google Scholar] [CrossRef]
- Csupor, D.; Blazsó, G.; Balogh, A.; Hohmann, J. The traditional Hungarian medicinal plant Centaurea sadleriana Janka accelerates wound healing in rats. J. Ethnopharmacol. 2010, 127, 193–195. [Google Scholar] [CrossRef]
- Sarker, S.D.; Kumarasamy, Y.; Shoeb, M.; Celik, S.; Eucel, E.; Middleton, M.; Nahar, L. Antibacterial and antioxidant activities of three Turkish species of the genus Centaurea. Adv. Tradit. Med. 2005, 5, 246–250. [Google Scholar]
- Erol-Dayi, Ö.; Pekmez, M.; Bona, M.; Aras-Perk, A.; Arda, N. Total phenolic contents, antioxidant activities cytotoxicity of three Centaurea species: C. calcitrapa subsp. calcitrapa, C. ptosimopappa C. spicata. Free Radic. Antioxid. 2011, 1, 31–36. [Google Scholar] [CrossRef]
- Kitouni, R.; Benayache, F.; Benayache, S. Flavonoids of the Exudate of Centaurea calcitrapa. Chem. Nat. Compd. 2015, 51, 762–763. [Google Scholar] [CrossRef]
- Bruno, M.; Bancheva, S.; Rosselli, S.; Maggio, A. Sesquiterpenoids in subtribe Centaureinae (Cass.) Dumort (tribe Cardueae, Asteraceae): Distribution, (13)C NMR spectral data and biological properties. Phytochemistry 2013, 95, 19–93. [Google Scholar] [CrossRef] [PubMed]
- Formisano, C.; Rigano, D.; Senatore, F.; Bancheva, S.; Maggio, A.; Rosselli, S.; Bruno, M. Flavonoids in subtribe Centaureinae (Cass.) Dumort. (tribe Cardueae, Asteraceae): Distribution and (13)C-NMR spectral data. Chem. Biodivers. 2012, 9, 2096–2158. [Google Scholar] [CrossRef] [PubMed]
- Kaskoos, R.A. In-vitro α-glucosidase inhibition and antioxidant activity of methanolic extract of Centaurea calcitrapa from Iraq. Am. J. Essent. Oils Nat. Prod. 2013, 1, 122–125. [Google Scholar]
- Pham, E.C.; Truong, T.N.; Dong, N.H.; Vo, D.D.; Hong Do, T.T. Synthesis of a Series of Novel 2-Amino-5-substituted 1,3,4-oxadiazole and 1,3,4-thiadiazole Derivatives as Potential Anticancer, Antifungal and Antibacterial Agents. Med. Chem. 2022, 18, 558–573. [Google Scholar] [CrossRef]
- Ibraheem, H.H.; Queen, B.K.; Al-Sabti, M.D.; Issa, A.A.; Al-Majedy, Y.K.; Jabir, M.S.; Sulaiman, G.M.; Hasoon, B.A.; Eshaq, M.M.; Jawad, K.H.; et al. Insights into the pharmaceutical properties and in silico study of novel hydrazone derivatives. Sci. Rep. 2024, 14, 29912. [Google Scholar] [CrossRef]
- Feng, W.; Hao, Z.; Li, M. Isolation and Structure Identification of Flavonoids. In Flavonoids—From Biosynthesis to Human Health; Justino, G., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Date, K.; Satoh, A.; Iida, K.; Ogawa, H. Pancreatic α-Amylase Controls Glucose Assimilation by Duodenal Retrieval through N-Glycan-specific Binding, Endocytosis, and Degradation. J. Biol. Chem. 2015, 290, 17439–17450. [Google Scholar] [CrossRef]
- de Souza, P.M.; de Oliveira Magalhães, P. Application of microbial α-amylase in industry—A review. Braz. J. Microbiol. 2010, 41, 850–861. [Google Scholar] [CrossRef]
- Kenza, B.; Djihane, A.; Mouad, B.; Ratiba, M.; Samir, B.; Fadila, B.; Souad, A. Renoprotective Effect of Centaurea choulettiana Pomel (Asteraceae) Leaves on Cisplatin-induced Oxidative Stress and Renal dysfunction in Mice. J. Appl. Pharm. Sci. 2017, 7, 147–154. [Google Scholar]
- Bensaad, M.; Dassamiour, S.; Hambaba, L.; Saidi, A.; Melakhsou, M.; Nouicer, F.; Baghiani, A.; Khennouf, S.; Kahoul, M.; Kadrine, N. In vivo investigation of antidiabetic, hepatoprotective, anti-inflammatory and antipyretic activities of Centaurea tougourensis Boiss. & Reut. J. Physiol. Pharmacol. 2021, 72, 439–449. [Google Scholar]
- Hajleh, M.N.A.; Khleifat, K.M.; Alqaraleh, M.; Al-Hraishat, E.a.; Al-Limoun, M.O.; Qaralleh, H.; Al-Dujaili, E.A. Antioxidant and antihyperglycemic effects of ephedra foeminea aqueous extract in streptozotocin-induced diabetic rats. Nutrients 2022, 14, 2338. [Google Scholar] [CrossRef] [PubMed]
- Erendor, F.; Eksi, Y.E.; Sahin, E.O.; Balci, M.K.; Griffith, T.S.; Sanlioglu, S. Lentivirus Mediated Pancreatic Beta-Cell-Specific Insulin Gene Therapy for STZ-Induced Diabetes. Mol. Ther. 2021, 29, 149–161. [Google Scholar] [CrossRef]
- Maiti, R.; Das, U.K.; Ghosh, D. Attenuation of hyperglycemia and hyperlipidemia in streptozotocin-induced diabetic rats by aqueous extract of seed of Tamarindus indica. Biol. Pharm. Bull. 2005, 28, 1172–1176. [Google Scholar] [CrossRef]
- LaMoia, T.E.; Shulman, G.I. Cellular and Molecular Mechanisms of Metformin Action. Endocr. Rev. 2021, 42, 77–96. [Google Scholar] [CrossRef]
- Abd-Alhussain, G.K.; Alatrakji, M.; Ahmed, S.J.; Fawzi, H.A. Efficacy of oral insulin nanoparticles for the management of hyperglycemia in a rat model of diabetes induced with streptozotocin. J. Med. Life 2024, 17, 217–225. [Google Scholar] [CrossRef]
- Gomathi, D.; Kalaiselvi, M.; Ravikumar, G.; Devaki, K.; Uma, C. Evaluation of antioxidants in the kidney of streptozotocin induced diabetic rats. Indian J. Clin. Biochem. IJCB 2014, 29, 221–226. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef]
- Ji, N.; Kim, S.-G.; Park, H.-H.; Lee, E.; Lee, Y.J.; Jin, M.; Lee, E. Nepetin, a natural compound from Inulae flos, suppresses degranulation and eicosanoid generation through PLCγ1 and Akt signaling pathways in mast cells. Arch. Pharmacal Res. 2020, 43, 224–232. [Google Scholar] [CrossRef]
- Yoon, S.Y.; Ahn, D.; Kim, J.K.; Seo, S.O.; Chung, S.J. Nepetin Acts as a Multi-Targeting Inhibitor of Protein Tyrosine Phosphatases Relevant to Insulin Resistance. Chem. Biodivers. 2022, 19, e202100600. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Han, R.; Hao, P.; Wang, L.; Liu, M.; Jin, M.; Kong, D.; Li, X. Nepetin inhibits IL-1β induced inflammation via NF-κB and MAPKs signaling pathways in ARPE-19 cells. Biomed. Pharmacother. 2018, 101, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Clavin, M.; Gorzalczany, S.; Macho, A.; Muñoz, E.; Ferraro, G.; Acevedo, C.; Martino, V. Anti-inflammatory activity of flavonoids from Eupatorium arnottianum. J. Ethnopharmacol. 2007, 112, 585–589. [Google Scholar] [CrossRef]
- Dumon, M.F.; Freneix-Clerc, M.; Carbonneau, M.A.; Thomas, M.J.; Perromat, A.; Clerc, M. In vitro demonstration of the 3′-5,7-trihydroxy-4′-methoxy flavone rutinoside antilipoperoxidant activity. Ann. Biol. Clin. 1994, 52, 265–270. [Google Scholar]
- Tang, H.; Zeng, Q.; Ren, N.; Wei, Y.; He, Q.; Chen, M.; Pu, P. Kaempferide improves oxidative stress and inflammation by inhibiting the TLR4/IκBα/NF-κB pathway in obese mice. Iran. J. Basic Med. Sci. 2021, 24, 493–498. [Google Scholar] [CrossRef]
- Nguyen, V.-S.; Shi, L.; Luan, F.-Q.; Wang, Q.-A. Synthesis of kaempferide Mannich base derivatives and their antiproliferative activity on three human cancer cell lines. Acta Biochim. Pol. 2015, 62, 547–552. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, X.; Li, D.; Hao, W.; Meng, F.; Wang, B.; Han, J.; Zheng, Q. Kaempferide protects against myocardial ischemia/reperfusion injury through activation of the PI3K/Akt/GSK-3β pathway. Mediat. Inflamm. 2017, 2017, 5278218. [Google Scholar] [CrossRef]
- Tang, H.; Zeng, Q.; Tang, T.; Wei, Y.; Pu, P. Kaempferide improves glycolipid metabolism disorder by activating PPARγ in high-fat-diet-fed mice. Life Sci. 2021, 270, 119133. [Google Scholar] [CrossRef]
- He, X.-W.; Yu, D.; Li, W.-L.; Zheng, Z.; Lv, C.-L.; Li, C.; Liu, P.; Xu, C.-Q.; Hu, X.-F.; Jin, X.-P. Anti-atherosclerotic potential of baicalin mediated by promoting cholesterol efflux from macrophages via the PPARγ-LXRα-ABCA1/ABCG1 pathway. Biomed. Pharmacother. 2016, 83, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Bossé, Y.; Weisnagel, S.J.; Bouchard, C.; Després, J.P.; Pérusse, L.; Vohl, M.C. Combined effects of PPARγ2 P12A and PPARα L162V polymorphisms on glucose and insulin homeostasis: The Québec Family Study. J. Hum. Genet. 2003, 48, 614–621. [Google Scholar] [CrossRef]
- Shyni, G.L.; Sasidharan, K.; Francis, S.K.; Das, A.A.; Nair, M.S.; Raghu, K.G. Licarin B from Myristica fragrans improves insulin sensitivity via PPARγ and activation of GLUT4 in the IRS-1/PI3K/AKT pathway in 3T3-L1 adipocytes. RSC Adv. 2016, 6, 79859–79870. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Stalin, A.; Balakrishna, K.; Ignacimuthu, S.; Paulraj, M.G.; Vishal, R. Insulin sensitization via partial agonism of PPARγ and glucose uptake through translocation and activation of GLUT4 in PI3K/p-Akt signaling pathway by embelin in type 2 diabetic rats. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 2243–2255. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef]
- Gong, L.; Feng, D.; Wang, T.; Ren, Y.; Liu, Y.; Wang, J. Inhibitors of α-amylase and α-glucosidase: Potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci. Nutr. 2020, 8, 6320–6337. [Google Scholar] [CrossRef]
- Sangeetha, R. Luteolin in the management of type 2 diabetes mellitus. Curr. Res. Nutr. Food Sci. J. 2019, 7, 393–398. [Google Scholar] [CrossRef]
- Kahksha; Alam, O.; Al-Keridis, L.A.; Khan, J.; Naaz, S.; Alam, A.; Ashraf, S.A.; Alshammari, N.; Adnan, M.; Beg, M.A. Evaluation of Antidiabetic Effect of Luteolin in STZ Induced Diabetic Rats: Molecular Docking, Molecular Dynamics, In Vitro and In Vivo Studies. J. Funct. Biomater. 2023, 14, 126. [Google Scholar] [CrossRef]
- Sheng, Y.; Zheng, S.; Ma, T.; Zhang, C.; Ou, X.; He, X.; Xu, W.; Huang, K. Mulberry leaf alleviates streptozotocin-induced diabetic rats by attenuating NEFA signaling and modulating intestinal microflora. Sci. Rep. 2017, 7, 12041. [Google Scholar] [CrossRef]
- Bogers, R.J.; Craker, L.E.; Lange, D. Medicinal and Aromatic Plants: Agricultural, Commercial, Ecological, Legal, Pharmacological and Social Aspects; Springer: Berlin/Heidelberg, Germany, 2006; Volume 17. [Google Scholar]
- Prasedya, E.S.; Frediansyah, A.; Martyasari, N.W.R.; Ilhami, B.K.; Abidin, A.S.; Padmi, H.; Fahrurrozi; Juanssilfero, A.B.; Widyastuti, S.; Sunarwidhi, A.L. Effect of particle size on phytochemical composition and antioxidant properties of Sargassum cristaefolium ethanol extract. Sci. Rep. 2021, 11, 17876. [Google Scholar] [CrossRef]
- Subramaniam, S.; Vaughn, K.; Carrier, D.J.; Clausen, E.C. Pretreatment of milk thistle seed to increase the silymarin yield: An alternative to petroleum ether defatting. Bioresour. Technol. 2008, 99, 2501–2506. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K.M.; Yoga Latha, L. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 1–10. [Google Scholar] [CrossRef]
- Jandera, P.; Churacek, J. Gradient Elution in Column Liquid Chromatography: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Garayev, E.; Di Giorgio, C.; Herbette, G.; Mabrouki, F.; Chiffolleau, P.; Roux, D.; Sallanon, H.; Ollivier, E.; Elias, R.; Baghdikian, B. Bioassay-guided isolation and UHPLC-DAD-ESI-MS/MS quantification of potential anti-inflammatory phenolic compounds from flowers of Inula montana L. J. Ethnopharmacol. 2018, 226, 176–184. [Google Scholar] [CrossRef]
- Colowick, S.P.; Kaplan, N.O. Amylase, α and β in Methods in Enzymology; Bernfeld, P., Ed.; Academic Press: New York, NY, USA, 1955; pp. 149–158. [Google Scholar]
- Rao, P.S.; Mohan, G.K. In vitro alpha-amylase inhibition and in vivo antioxidant potential of Momordica dioica seeds in streptozotocin-induced oxidative stress in diabetic rats. Saudi J. Biol. Sci. 2017, 24, 1262–1267. [Google Scholar] [CrossRef]
- Buchwald, P. A single unified model for fitting simple to complex receptor response data. Sci. Rep. 2020, 10, 13386. [Google Scholar] [CrossRef]
- .Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.A.; Meyer, R.; Miller, D.; et al. AVMA Guidelines for the Euthanasia of Animals: 2020 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, A.; Jeddi, S. Streptozotocin as a tool for induction of rat models of diabetes: A practical guide. EXCLI J. 2023, 22, 274–294. [Google Scholar] [CrossRef]
- Ramírez, K.; Quesada-Yamasaki, D.; Jaime, F.-T.C. A Protocol to Perform Systemic Lipopolysacharide (LPS) Challenge in Rats. Odovtos 2019, 21, 53–66. [Google Scholar] [CrossRef]
- Khafaji, A.W.; Al-Zubaidy, A.A.; Farhood, I.G.; Fawzi, H.A. Effects of topical isoxsuprine ointment on imiquimod-induced psoriasiform skin inflammation in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 398, 1545–1556. [Google Scholar] [CrossRef]
- Bouhrim, M.; Bencheikh, N.; Imtara, H.; Daoudi, N.E.; Mechchate, H.; Ouassou, H.; Kharchoufa, L.; Elachouri, M.; Mekhfi, H.; Ziyyat, A.; et al. Protective Effect of Opuntia dillenii (Ker Gawl.) Haw. Seed Oil on Gentamicin-Induced Nephrotoxicity: A Biochemical and Histological Analysis. Sci. World J. 2021, 2021, 2173012. [Google Scholar] [CrossRef]
- Han, X.; Tao, Y.L.; Deng, Y.P.; Yu, J.W.; Cai, J.; Ren, G.F.; Sun, Y.N.; Jiang, G.J. Metformin ameliorates insulitis in STZ-induced diabetic mice. PeerJ 2017, 5, e3155. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Mostofa, M.; Hoque, M.; Das, S.; Sarkar, A. Comparative efficacy of Neem (Azadirachta indica) and Metformin hydrochloride (Comet®) in streptozotocin induced diabetes melitus in rats. Bangl. J. Vet. Med. 2010, 8, 75–80. [Google Scholar] [CrossRef]
- Heinegård, D.; Tiderström, G. Determination of serum creatinine by a direct colorimetric method. Clin. Chim. Acta Int. J. Clin. Chem. 1973, 43, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Bergmeyer, H.-U. Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Talke, H.; Schubert, G.E. Enzymatic urea determination in the blood and serum in the warburg optical test. Klin. Wochenschr. 1965, 43, 174–175. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Charan, J.; Kantharia, N.D. How to calculate sample size in animal studies? J. Pharmacol. Pharmacother. 2013, 4, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Festing, M.F.W. Design and Statistical Methods in Studies Using Animal Models of Development. ILAR J. 2006, 47, 5–14. [Google Scholar] [CrossRef]
- García-Pérez, M.A. Use and misuse of corrections for multiple testing. Methods Psychol. 2023, 8, 100120. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analysis | P-I | P-II | P-III |
|---|---|---|---|
| Benzoyl peak | |||
| Cinnamoyl peak [29] | 273 | 269 | 268 |
| Extended conjugation | 351 | 364 | 352 |
| FT-IR analysis | |||
| O-H stretch | 3407 | 3522–3284 | 3397 and 3219 |
| Aromatic C-H stretching | 2947 | 2964–2829 | 2923 |
| C=O (carbonyl) and C=C (aromatic) stretches | 1656 and 1607 | 1654–1611 | 1659 and 1612 |
| C-H bending vibrations | 1456 and 1377 | 1371 | 1443 and 1351 |
| C-O stretching | 1271–1037 | 1258–1020 | 1260–1030 |
| MS analysis | |||
| M + H + 1 | 317.8 | 302.7 | 288.7 |
| M + H | 316.8 | 301.7 | 287.7 |
| M + H-CH2O | 271.6 | ||
| M + H-C3H6O | 229.5 | ||
| M + H-C9H6O2 | 171.4 | 171.4 | |
| M + H-C7H4O5 | 149.3 | ||
| C7H4O3 | 136.3 | 136.3 | |
| C7H5O3 | 137.3 | ||
| C8H9O | 121.3 | ||
| C2H2O | 42.2 | 42.2 | 42.3 |
| Group | Details | Procedure |
|---|---|---|
| G-1 | The control group | Did not receive any treatment |
| G-2 | The induction group | Received 60 mg/kg STZ and, after three days, received 10 mL/kg distilled water orally via gastric gavage daily for 30 days [75] |
| G-3 | The metformin group | Received 60 mg/kg STZ and, after three days, received 250 mg/kg metformin orally via gastric gavage daily for 30 days [72,76,77]. |
| G-4 | C. calcitrapa chloroform extract (E1) group | Received 60 mg/kg STZ and, after three days, received 250 mg/kg of C. calcitrapa chloroform extract daily for 30 days. |
| G-5 | C calcitrapa ethyl acetate extract (E2) group | Received 60 mg/kg STZ and, after three days, received 250 mg/kg of C. calcitrapa ethyl acetate extract daily for 30 days. |
| G-6 | C calcitrapa methanol extract (E3) group | Received 60 mg/kg STZ and, after three days, received 250 mg/kg of C. calcitrapa methanol extract daily for 30 days. |
| Group | Details | Procedure |
|---|---|---|
| S-1 | The control group | Did not receive any treatment |
| S-2 | The induction group | Received 60 mg/kg STZ and, after three days, received 10 mL/kg distilled water orally via gastric gavage daily for 30 days [75] |
| S-3 | The metformin group | Received 60 mg/kg STZ and, after three days, received 250 mg/kg metformin orally via gastric gavage daily for 30 days [72,76,77] |
| S-4 | C calcitrapa ethyl acetate extract fraction I (E2-I) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-I) fraction orally via gastric gavage daily for 30 days |
| S-5 | C calcitrapa ethyl acetate extract fraction II (E2-II) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-II) fraction orally via gastric gavage daily for 30 days |
| S-6 | C calcitrapa ethyl acetate extract fraction III (E2-III) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-III) fraction orally via gastric gavage daily for 30 days |
| S-7 | C calcitrapa ethyl acetate extract fraction IV (E2-IV) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-IV) fraction orally via gastric gavage daily for 30 days |
| S-8 | C calcitrapa ethyl acetate extract fraction V (E2-V) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-V) fraction orally via gastric gavage daily for 30 days |
| S-9 | C calcitrapa ethyl acetate extract fraction VI (E2-VI) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-VI) fraction orally via gastric gavage daily for 30 days |
| S-10 | C calcitrapa ethyl acetate extract fraction VII (E2-VII) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-VII) fraction orally via gastric gavage daily for 30 days |
| S-11 | C calcitrapa ethyl acetate extract fraction VIII (E2-VIII) group | Received 60 mg/kg STZ and, after three days, received 100 mg/kg (E2-VIII) fraction orally via gastric gavage daily for 30 days |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadhim, H.M.; Kadhim, Y.M.; Fawzi, H.A.; Abdul Khalik, Z.M.; Jawad, A.M.; Ghédira, K. Bioassay-Guided Isolation and Active Compounds Identification of the AntiDiabetic Fractions of Centaurea calcitrapa Extract and the Predicted Interaction Mechanism. Molecules 2025, 30, 2394. https://doi.org/10.3390/molecules30112394
Kadhim HM, Kadhim YM, Fawzi HA, Abdul Khalik ZM, Jawad AM, Ghédira K. Bioassay-Guided Isolation and Active Compounds Identification of the AntiDiabetic Fractions of Centaurea calcitrapa Extract and the Predicted Interaction Mechanism. Molecules. 2025; 30(11):2394. https://doi.org/10.3390/molecules30112394
Chicago/Turabian StyleKadhim, Hayder Mohammed, Yasir M. Kadhim, Hayder Adnan Fawzi, Zaid M. Abdul Khalik, Ali Mohammed Jawad, and Kamel Ghédira. 2025. "Bioassay-Guided Isolation and Active Compounds Identification of the AntiDiabetic Fractions of Centaurea calcitrapa Extract and the Predicted Interaction Mechanism" Molecules 30, no. 11: 2394. https://doi.org/10.3390/molecules30112394
APA StyleKadhim, H. M., Kadhim, Y. M., Fawzi, H. A., Abdul Khalik, Z. M., Jawad, A. M., & Ghédira, K. (2025). Bioassay-Guided Isolation and Active Compounds Identification of the AntiDiabetic Fractions of Centaurea calcitrapa Extract and the Predicted Interaction Mechanism. Molecules, 30(11), 2394. https://doi.org/10.3390/molecules30112394