

Preliminary Study on EGCG-Enhanced Vanadium Toxicity in Cells: Impact on Oxidative Stress

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Effect of EGCG on V Toxicity in Cells
2.2. Effect of V–EGCG Cotreatment on MMP
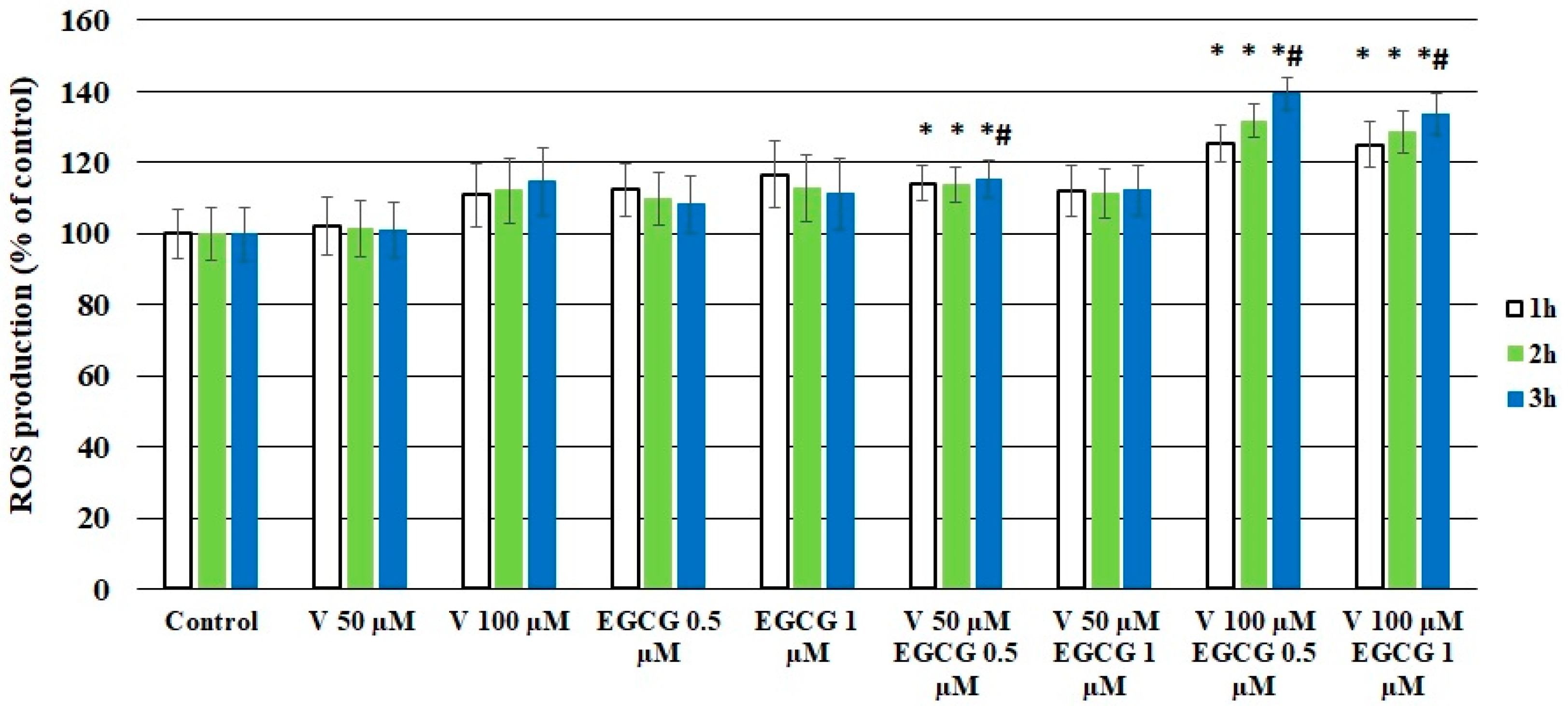
2.3. EGCG Influenced the ROS Production in CHO-K1 Cells Treated with V
2.4. Caspase-3 Activity
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. V and EGCG Concentrations
4.4. Reagents Preparations
4.5. Cell Viability Assessment with Resazurin
4.6. Mitochondrial Membrane Potential (MMP) Assessment with MitoPT TMRM Assay
4.7. Determination of Changes in ROS Levels Using an Intracellular Total ROS Activity Assay Kit
4.8. Determination of Caspase-3 Activity
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barceloux, D.G. Vanadium. Clin. Toxicol. 1999, 37, 265–278. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.S.; Martins, H.; Gutiérrez-Merino, C.; Aureliano, M. Vanadium and cadmium in vivo effects in teleost cardiac muscle: Metal accumulation and oxidative stress markers. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I. Vanadium detoxification and resistance in yeast: A minireview. Ann. Microbiol. 2001, 51, 1–9. [Google Scholar]
- Kamika, I.; Momba, M.N.B. Effect of vanadium toxicity at its different oxidation states on selected bacterial and protozoan isolates in wastewater systems. Environ. Technol. 2014, 35, 2075–2085. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, B.; Yuan, H.; Cheng, Y.; Wang, S.; He, Z. Microbial reduction of vanadium (V) in groundwater: Interactions with coexisting common electron acceptors and analysis of microbial community. Environ. Pollut. 2017, 231, 1362–1369. [Google Scholar] [CrossRef]
- Arcega-Cabrera, F.; Gold-Bouchot, G.; Lamas-Cosío, E.; Dótor-Almazán, A.; Ceja-Moreno, V.; Zapata-Pérez, O.; Oceguera-Vargas, I. Vanadium and Cadmium in Water from the Perdido Area, Northwest of the Gulf of Mexico: 2 years’ Monitoring and Current Status. Bull. Environ. Contam. Toxicol. 2022, 108, 37–42. [Google Scholar] [CrossRef]
- Peng, H.; Guo, J.; Li, B.; Huang, H. Vanadium properties, toxicity, mineral sources and extraction methods: A review. Environ. Chem. Lett. 2022, 20, 1249–1263. [Google Scholar] [CrossRef]
- Zwolak, I.; Wnuk, E. Sodium pyruvate as a protective compound against vanadyl sulphate-induced reactive species generation and mitochondrial destabilisation in CHO-K1 cells. Int. J. Mol. Sci. 2022, 11, 909. [Google Scholar] [CrossRef]
- Tian, Y.; Qi, H.; Wang, G.; Li, L.; Zhou, D. Anticancer effect of sodium metavanadate on murine breast cancer both in vitro and in vivo. BioMetals 2021, 34, 557–571. [Google Scholar] [CrossRef]
- Mukherjee, B.; Patra, B.; Mahapatra, S.; Banerjee, P.; Tiwari, A.; Chatterjee, M. Vanadium—An element of atypical biological significance. Toxicol. Lett. 2004, 150, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xiao, Z.; Wang, H.; Yang, K. Toxic effects of vanadium (V) on a combined autotrophic denitrification system using sulfur and hydrogen as electron donors. Bioresour. Technol. 2018, 264, 319–326. [Google Scholar] [CrossRef]
- Capella, L.S.; Gefé, M.R.; Silva, E.F.; Affonso-Mitidieri, O.; Lopes, A.G.; Rumjanek, V.M.; Capella, M.A.M. Mechanisms of vanadate-induced cellular toxicity: Role of cellular glutathione and NADPH. Arch. Biochem. Biophys. 2002, 406, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Aureliano, M.; Gândara, R.M.C. Decavanadate effects in biological systems. J. Inorg. Biochem. 2005, 99, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.S.; Martins, H.; Duarte, R.O.; Moura, J.J.G.; Coucelo, J.; Gutiérrez-Merino, C.; Aureliano, M. Vanadium distribution, lipid peroxidation and oxidative stress markers upon decavanadate in vivo administration. J. Inorg. Biochem. 2007, 101, 80–88. [Google Scholar] [CrossRef]
- Smith, D.M.; Pickering, R.M.; Lewith, G.T. A systematic review of vanadium oral supplements for glycaemic control in type 2 diabetes mellitus. QJM Int. J. Med. 2008, 101, 351–358. [Google Scholar] [CrossRef]
- Bartosikova, L.; Necas, J. Epigallocatechin gallate: A review. Vet. Med. 2018, 63, 443–467. [Google Scholar] [CrossRef]
- Weinreb, O.; Amit, T.; Mandel, S.; Youdim, M.B.H. Neuroprotective molecular mechanisms of (−)-epigallocatechin-3-gallate: A reflective outcome of its antioxidant, iron chelating and neuritogenic properties. Genes Nutr. 2009, 4, 283–296. [Google Scholar] [CrossRef]
- Pervin, M.; Unno, K.; Takagaki, A.; Isemura, M.; Nakamura, Y. Function of green tea catechins in the brain: Epigallocatechin gallate and its metabolites. Int. J. Mol. Sci. 2019, 20, 3630. [Google Scholar] [CrossRef]
- Carlson, J.R.; Bauer, B.A.; Vincent, A.; Limburg, P.J.; Wilson, T. Reading the tea leaves: Anticarcinogenic properties of (-)- epigallocatechin-3-gallate. Mayo Clin. Proc. 2007, 82, 725–732. [Google Scholar] [CrossRef]
- Lin, Y.S.; Tsai, Y.J.; Tsay, J.S.; Lin, J.K. Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J. Agric. Food Chem. 2003, 51, 1864–1873. [Google Scholar] [CrossRef] [PubMed]
- Kucera, O.; Mezera, V.; Moravcova, A.; Endlicher, R.; Lotkova, H.; Drahota, Z.; Cervinkova, Z. In vitro toxicity of epigallocatechin gallate in rat liver mitochondria and hepatocytes. Oxid. Med. Cell. Longev. 2015, 2015, 476180. [Google Scholar] [CrossRef]
- Wu, C.D.; Wei, G. Tea as a functional food for oral health. Nutrition 2002, 18, 443–444. [Google Scholar] [CrossRef]
- Cai, Z.Y.; Li, X.M.; Liang, J.P.; Xiang, L.P.; Wang, K.R.; Shi, Y.L.; Yang, R.; Shi, M.; Ye, J.H.; Lu, J.L.; et al. Bioavailability of tea catechins and its improvement. Molecules 2018, 23, 2346. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.Y.; Chen, C.Y.; Chiou, Y.H.; Shyu, H.W.; Lin, K.H.; Chou, M.C.; Huang, M.H.; Wang, Y.F. Epigallocatechin-3-gallate suppresses human herpesvirus 8 replication and induces ROS leading to apoptosis and autophagy in primary effusion lymphoma cells. Int. J. Mol. Sci. 2018, 19, 16. [Google Scholar] [CrossRef]
- Kuriya, K.; Itoh, S.; Isoda, A.; Tanaka, S.; Nishio, M.; Umekawa, H. Green tea polyphenol EGCg induces cell fusion via reactive oxygen species. Biochem. Biophys. Rep. 2023, 35, 101536. [Google Scholar] [CrossRef] [PubMed]
- Sukanya, P.; Reddy, C.V.R. Structural investigation, DNA interactions and in vitro anticancer studies of transition metal complexes of 3-(2-(2,4-dihydroxy benzylidene) hydrazinyl) quinoxalin-2(1H)-one. J. Biomol. Struct. Dyn. 2022, 40, 6151–6162. [Google Scholar] [CrossRef]
- García-Fernández, A.J.; Bayoumi, A.E.; Pérez-Pertejo, Y.; Motas, M.; Reguera, R.M.; Ordóñez, C.; Balaña-Fouce, R.; Ordóñez, D. Alterations of the glutathione-redox balance induced by metals in CHO-K1 cells. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 132, 365–373. [Google Scholar] [CrossRef]
- Bayoumi, A.; Garcfa-femandez, A.J.; Navas, I.; Balana-Fouce, R.; Ordonez, D. Cytotoxic evaluation of heavy metals on mammalian and fish cellular model. Toxicol. Lett. 1996, 88, 12. [Google Scholar] [CrossRef]
- Ferrer, E.; Juan-García, A.; Font, G.; Ruiz, M.J. Reactive oxygen species induced by beauvericin, patulin and zearalenone in CHO-K1 cells. Toxicol. In Vitro 2009, 23, 1504–1509. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Foldbjerg, R.; Miclaus, T.; Wang, L.; Singh, R.; Hayashi, Y.; Sutherland, D.; Chen, C.; Autrup, H.; Beer, C. Multi-platform genotoxicity analysis of silver nanoparticles in the model cell line CHO-K1. Toxicol. Lett. 2013, 222, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Plewa, M.J.; Wagner, E.D.; Muellner, M.G.; Hsu, K.-M.; Richardson, S.D. Comparative mammalian cell toxicity of N-DBPs and C-DBPs. In Disinfection By-Products in Drink Water; American Chemical Society: Washington, DC, USA, 2008; Volume 995, pp. 36–50. [Google Scholar] [CrossRef]
- Wang, Y.; Plewa, M.J.; Mukherjee, U.K.; Verma, V. Assessing the cytotoxicity of ambient particulate matter (PM) using Chinese hamster ovary (CHO) cells and its relationship with the PM chemical composition and oxidative potential. Atmos. Environ. 2018, 179, 132–141. [Google Scholar] [CrossRef]
- Souza, T.A.J.; Franchi, L.P.; Rosa, L.R.; da Veiga, M.A.M.S.; Takahashi, C.S. Cytotoxicity and genotoxicity of silver nanoparticles of different sizes in CHO-K1 and CHO-XRS5 cell lines. Mutat. Res.—Genet. Toxicol. Environ. Mutagen. 2016, 795, 70–83. [Google Scholar] [CrossRef]
- Zhang, H.X.; Du, G.H.; Zhang, J.T. Assay of mitochondrial functions by resazurin in vitro. Acta Pharmacol. Sin. 2004, 25, 385–389. [Google Scholar] [PubMed]
- Nakagawa, K.; Miyazawa, T. Absorption and distribution of tea catechin, (−)-epigallocatechin-3-gallate, in the rat. J. Nutr. Sci. Vitaminol. 1997, 43, 679–684. [Google Scholar] [CrossRef]
- Lee, M.J.; Maliakal, P.; Chen, L.; Meng, X.; Bondoc, F.Y.; Prabhu, S.; Lambert, G.; Mohr, S.; Yang, C.S. Pharmacokinetics of tea catechins after ingestion of green tea and (−)-epigallocatechin-3-gallate by humans: Formation of different metabolites and individual variability. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1025–1032. [Google Scholar]
- Naumovski, N.; Blades, B.L.; Roach, P.D. Food inhibits the oral bioavailability of the major green tea antioxidant epigallocatechin gallate in humans. Antioxidants 2015, 4, 373–393. [Google Scholar] [CrossRef]
- Kim, S.; Lee, M.J.; Hong, J.; Li, C.; Smith, T.J.; Yang, G.Y.; Seril, D.N.; Yang, C.S. Plasma and tissue levels of tea catechins in rats and mice during chronic consumption of green tea polyphenols. Nutr. Cancer 2000, 37, 41–48. [Google Scholar] [CrossRef]
- Cuesta, S.; Francés, D.; García, G.B. ROS formation and antioxidant status in brain areas of rats exposed to sodium metavanadate. Neurotoxicol. Teratol. 2011, 33, 297–302. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, C.; Li, J.; Leonard, S.S.; Lanciotti, R.; Butterworth, L.; Shi, X.; Zhang, Z.; Shi, X. Vanadate-induced cell growth regulation and the role of reactive oxygen species. Arch. Biochem. Biophys. 2001, 392, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, T.; Ito, K.; Ikeda, Y.; Kizaki, M. Green tea component, catechin, induces apoptosis of human malignant B cells via production of reactive oxygen species. Clin. Cancer Res. 2005, 11, 6040–6049. [Google Scholar] [CrossRef]
- Kim, J.Y.; Choi, J.Y.; Lee, H.J.; Byun, C.J.; Park, J.H.; Park, J.H.; Cho, H.S.; Cho, S.J.; Jo, S.A.; Jo, I. The green tea component (-)-epigallocatechin-3-gallate sensitizes primary endothelial cells to Arsenite-induced apoptosis by decreasing C-Jun n-Terminal Kinase-mediated Catalase activity. PLoS ONE 2015, 10, e0138590. [Google Scholar] [CrossRef]
- Yu, H.N.; Shen, S.R.; Yin, J.J. Effects of interactions of EGCG and Cd2+ on the growth of PC-3 cells and their mechanisms. Food Chem. Toxicol. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.C.; Yu, H.N.; Sun, S.; Yang, J.G.; He, G.Q.; Ruan, H.; Shen, S.R. Investigations of the cytotoxicity of epigallocatechin-3-gallate against PC-3 cells in the presence of Cd2+ in vitro. Toxicol. In Vitro 2008, 22, 953–960. [Google Scholar] [CrossRef]
- Banki, K.; Hutter, E.; Gonchoroff, N.J.; Perl, A. Elevation of Mitochondrial Transmembrane Potential and Reactive Oxygen Intermediate Levels Are Early Events and Occur Independently from Activation of Caspases in Fas Signaling. J. Immunol. 1999, 162, 1466–1479. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alcazar, J.A.; Ault, J.G.; Khodjakov, A.; Schneider, E. Increased mitochondrial cytochrome c levels and mitochondrial hyperpolarization precede camptothecin-induced apoptosis in Jurkat cells. Cell Death Differ. 2000, 7, 1090–1100. [Google Scholar] [CrossRef]
- Giovannini, C.; Matarrese, P.; Scazzocchio, B.; Sanchez, M.; Masella, R.; Malorni, W. Mitochondria hyperpolarization is an early event in oxidized low-density lipoprotein-induced apoptosis in Caco-2 intestinal cells. FEBS Lett. 2002, 523, 200–206. [Google Scholar] [CrossRef]
- Almeida, A.; Almeida, J.; Bolañ, J.P.; Moncada, S. Different responses of astrocytes and neurons to nitric oxide: The role of glycolytically generated ATP in astrocyte protection. Proc. Natl. Acad. Sci. USA 2001, 98, 15294–15299. [Google Scholar] [CrossRef]
- Bondad, S.E.C.; Kurasaki, M. Analysis of Cadmium, Epigallocatechin Gallate, and Vitamin C Co-exposure on PC12 Cellular Mechanisms. Biol. Trace Elem. Res. 2020, 198, 627–635. [Google Scholar] [CrossRef]
- Bertram, B.; Bollow, U.; Rajaee-Behbahani, N.; Bürkle, A.; Schmezer, P. Induction of poly(ADP-ribosyl)ation and DNA damage in human peripheral lymphocytes after treatment with (-)-epigallocatechin-gallate. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2003, 534, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Hasumi, K.; Takami, M.; Aida-Hyugaji, S.; Woo, J.T.; Nagai, K.; Ishikawa, T.; Wachi, M. Identification of two biologically crucial hydroxyl groups of (-)-epigallocatechin gallate in osteoclast culture. Biochem. Pharmacol. 2007, 73, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, T.; Ogo, T.; Eto, T.; Asano, Y.; Suganuma, M.; Niho, Y. Growth inhibition of leukemic cells by (−)-epigallocatechin gallate, the main constitient of green tea. Life Sci. 1998, 63, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.T.; Ha, J.; Park, I.J.; Lee, S.K.; Baik, H.W.; Kim, Y.M.; Park, O.J. Apoptotic effect of EGCG in HT-29 colon cancer cells via AMPK signal pathway. Cancer Lett. 2007, 247, 115–121. [Google Scholar] [CrossRef]
- Lambert, J.D.; Sang, S.; Hong, J.; Kwon, S.J.; Lee, M.J.; Ho, C.T.; Yang, C.S. Peracetylation as a means of enhancing in vitro bioactivity and bioavailability of epigallocatechin-3-gallate. Drug Metab. Dispos. 2006, 34, 2111–2116. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wnuk, E.; Zwolak, I. Preliminary Study on EGCG-Enhanced Vanadium Toxicity in Cells: Impact on Oxidative Stress. Molecules 2025, 30, 2114. https://doi.org/10.3390/molecules30102114
Wnuk E, Zwolak I. Preliminary Study on EGCG-Enhanced Vanadium Toxicity in Cells: Impact on Oxidative Stress. Molecules. 2025; 30(10):2114. https://doi.org/10.3390/molecules30102114
Chicago/Turabian StyleWnuk, Ewa, and Iwona Zwolak. 2025. "Preliminary Study on EGCG-Enhanced Vanadium Toxicity in Cells: Impact on Oxidative Stress" Molecules 30, no. 10: 2114. https://doi.org/10.3390/molecules30102114
APA StyleWnuk, E., & Zwolak, I. (2025). Preliminary Study on EGCG-Enhanced Vanadium Toxicity in Cells: Impact on Oxidative Stress. Molecules, 30(10), 2114. https://doi.org/10.3390/molecules30102114