
Cell-Free Synthesis: Expediting Biomanufacturing of Chemical and Biological Molecules

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
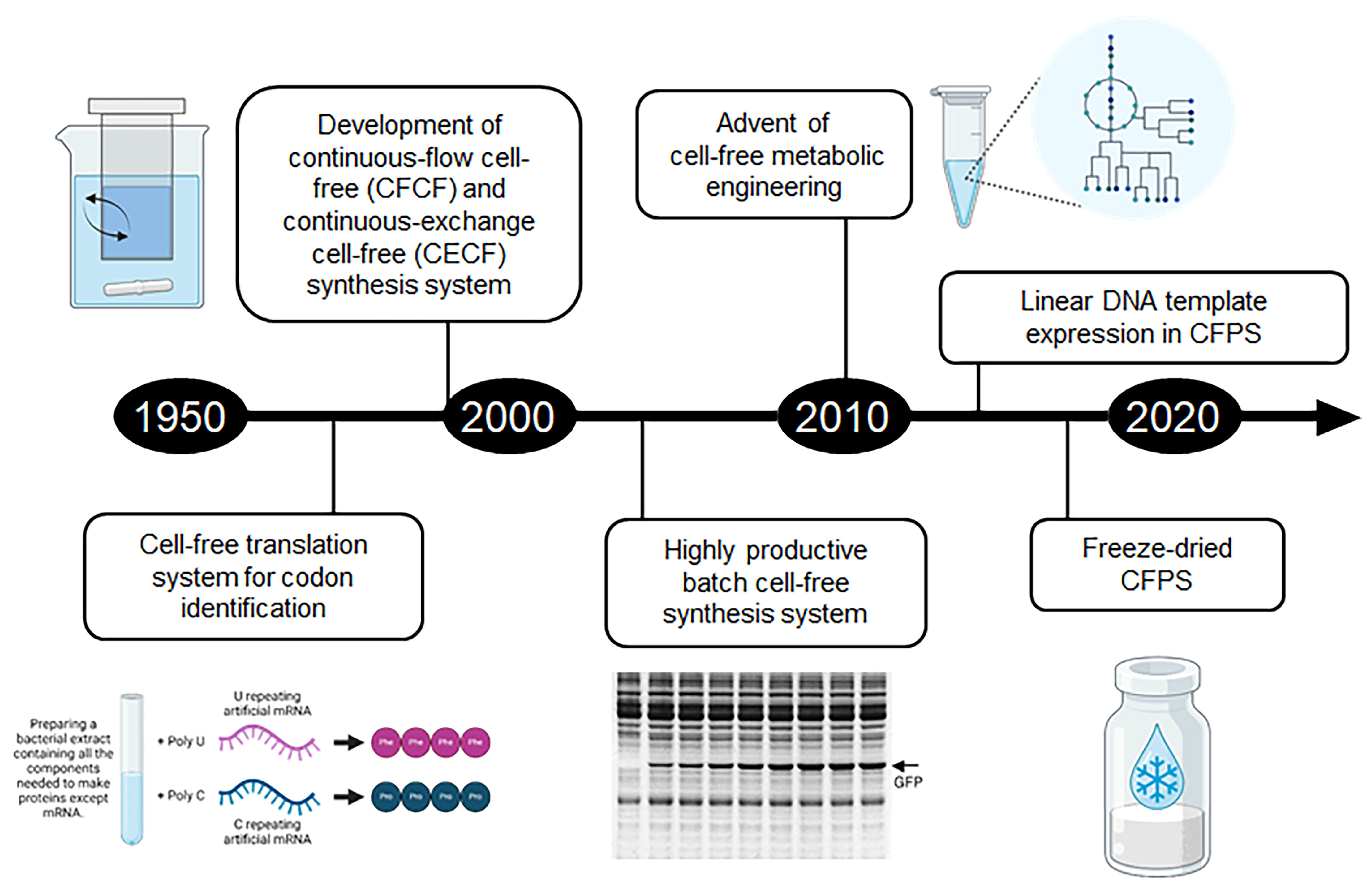
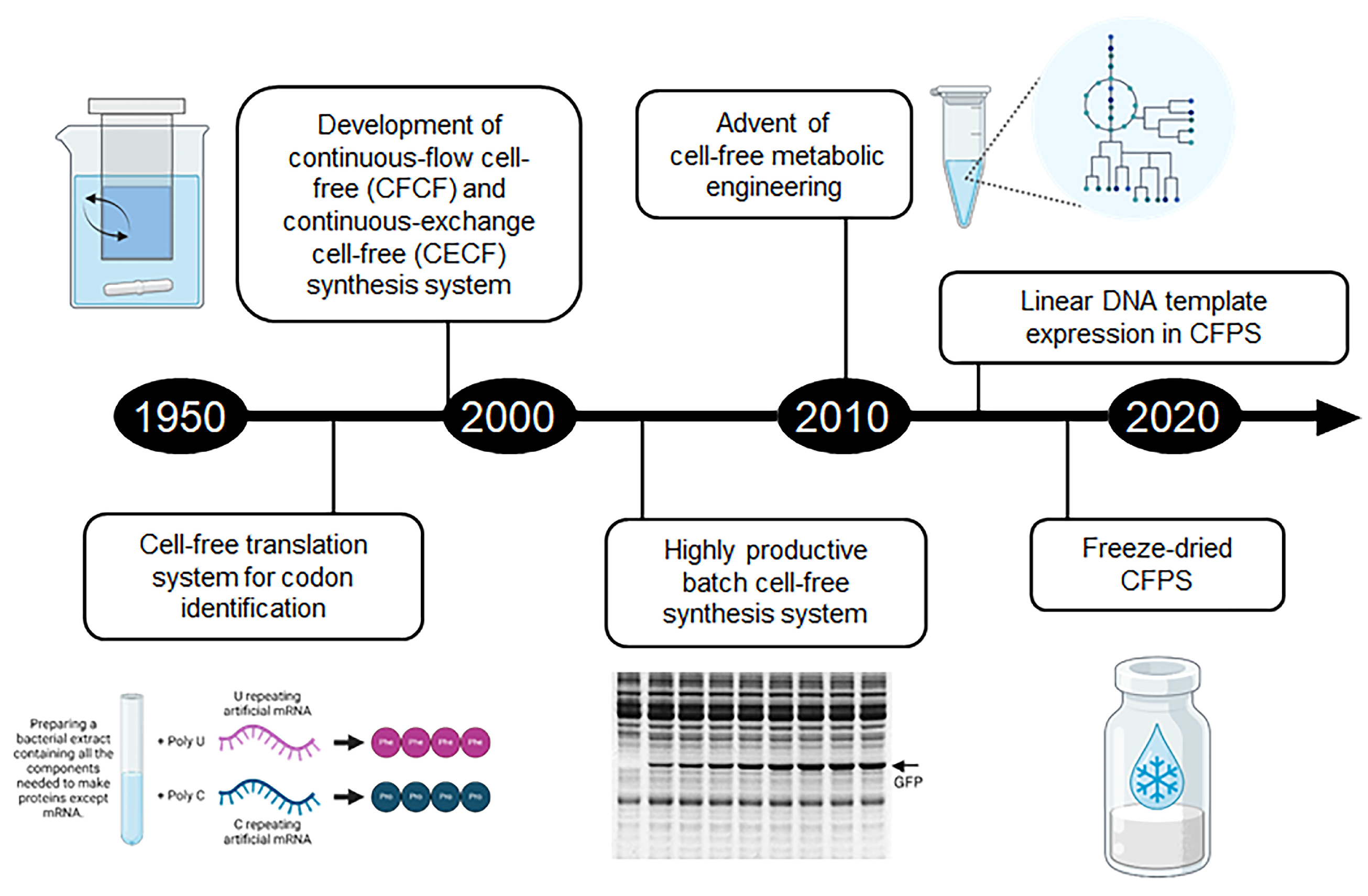
2. Development and Evolution of Cell-Free Synthesis Systems
3. Classification of Cell-Free Synthesis Systems
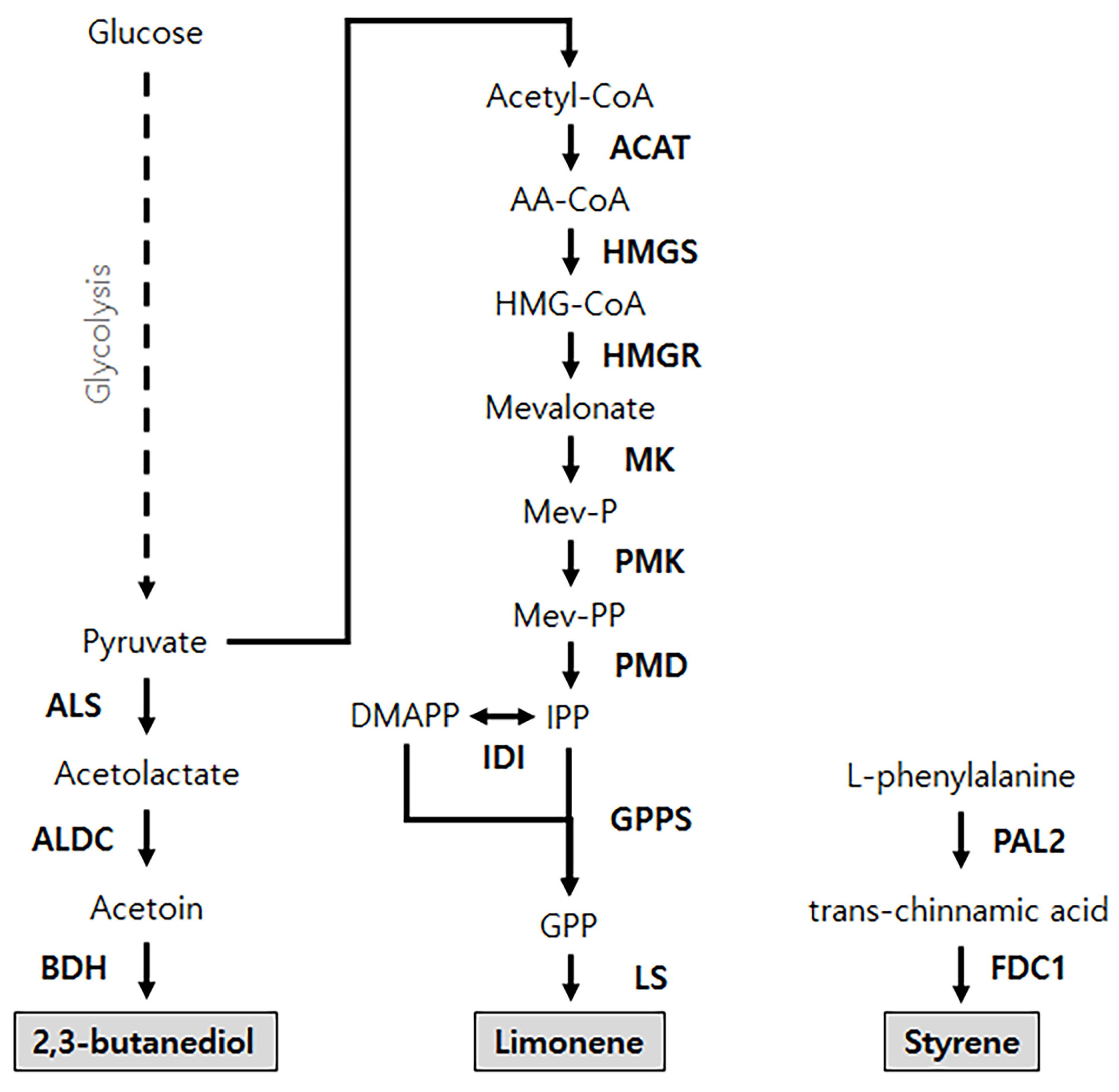
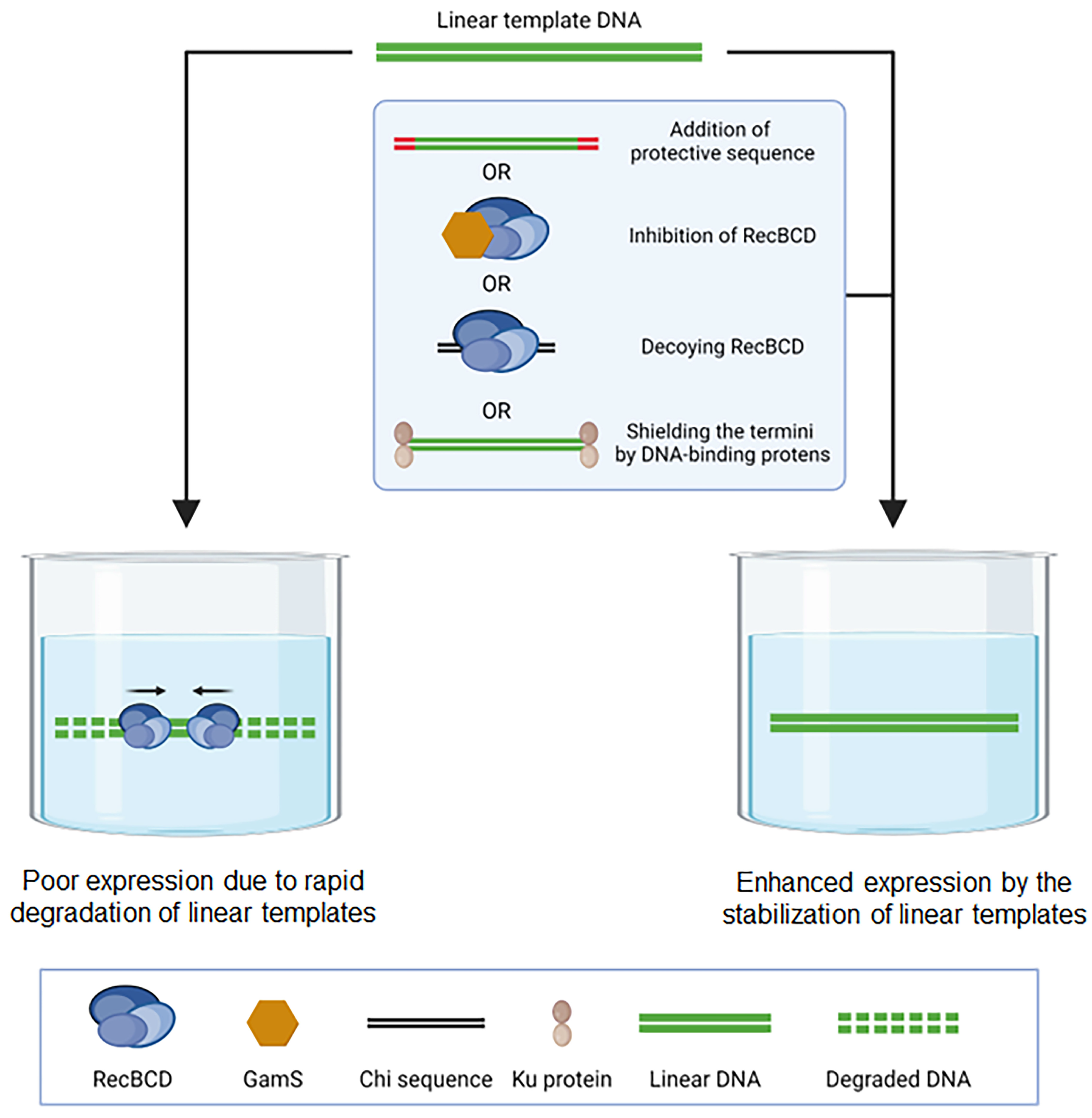
4. Deploying Synthetic Pathways into Cell-Free Synthesis Systems
5. Harnessing Cell-Free Synthesis for Decentralized Biomanufacturing
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mengal, P.; Wubbolts, M.; Zika, E.; Ruiz, A.; Brigitta, D.; Pieniadz, A.; Black, S. Bio-based Industries Joint Undertaking: The catalyst for sustainable bio-based economic growth in Europe. New Biotech. 2018, 40, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Grangeia, C.; Santos, L.; Lazaro, L.L.B. The Brazilian biofuel policy (RenovaBio) and its uncertainties: An assessment of technical, socioeconomic and institutional aspects. Energy Conv. Manag.-X 2022, 13, 100156. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, C.; Shao, M.-W.; Chen, Y.-L.; Liu, P.; Chen, G.-Q. The roadmap of bioeconomy in China. Eng. Biol. 2022, 6, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Pascoli, D.U.; Aui, A.; Frank, J.; Therasme, O.; Dixon, K.; Gustafson, R.; Kelly, B.; Volk, T.A.; Wright, M.M. The US bioeconomy at the intersection of technology, policy, and education. Biofuel. Bioprod. Bior. 2022, 16, 9–26. [Google Scholar] [CrossRef]
- Stock, M.; Gorochowski, T.E. Open-endedness in synthetic biology: A route to continual innovation for biological design. Sci. Adv. 2024, 10, eadi3621. [Google Scholar] [CrossRef] [PubMed]
- Cravens, A.; Payne, J.; Smolke, C.D. Synthetic biology strategies for microbial biosynthesis of plant natural products. Nat. Commun. 2019, 10, 2142. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, X.; Zhao, C.; Chen, G.-Q. Applications of synthetic biology in medical and pharmaceutical fields. Signal Transduct. Target. Ther. 2023, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Peplow, M. Around the world in a month. Nat. Biotechnol. 2015, 33, 1123–1125. [Google Scholar] [PubMed]
- Karim, A.S.; Dudley, Q.M.; Juminaga, A.; Yuan, Y.; Crowe, S.A.; Heggestad, J.T.; Garg, S.; Abdalla, T.; Grubbe, W.S.; Rasor, B.J.; et al. In vitro prototyping and rapid optimization of biosynthetic enzymes for cell design. Nat. Chem. Biol. 2020, 16, 912–919. [Google Scholar] [CrossRef]
- Gurdo, N.; Volke, D.C.; Nikel, P.I. Merging automation and fundamental discovery into the design-build-test-learn cycle of nontraditional microbes. Trends Biotechnol. 2022, 40, 1148–1159. [Google Scholar] [CrossRef]
- Mohsenin, H.; Wagner, H.J.; Rosenblatt, M.; Kemmer, S.; Drepper, F.; Huesgen, P.; Timmer, J.; Weber, W. Design of a Biohybrid Materials Circuit with Binary Decoder Functionality. Adv. Mater. 2023, 36, e2308092. [Google Scholar] [CrossRef]
- Tanaka, K.; Bamba, T.; Kondo, A.; Hasunuma, T. Metabolomics-based development of bioproduction processes toward industrial-scale production. Curr. Opin. Biotechnol. 2024, 85, 103057. [Google Scholar] [CrossRef] [PubMed]
- Tellechea-Luzardo, J.; Otero-Muras, I.; Goñi-Moreno, A.; Carbonell, P. Fast biofoundries: Coping with the challenges of biomanufacturing. Trends Biotechnol. 2022, 40, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.N.; Cho, N.; Lee, K.; Gwon, D.A.; Lee, J.W.; Lee, J. Programmable Synthesis of Biobased Materials Using Cell-Free Systems. Adv. Mater. 2023, 35, 24. [Google Scholar] [CrossRef]
- Gu, Y.Q.; Fan, F.; Liu, Y.; Chai, Y.F.; Yuan, Y.F.; Chen, X.F. Cell-free protein synthesis system for bioanalysis: Advances in methods and applications. Trac-Trends Anal. Chem. 2023, 161, 13. [Google Scholar] [CrossRef]
- Ullah, M.W.; Manan, S.; Ul-Islam, M.; Khattak, W.A.; Khan, K.A.; Liu, J.; Yang, G.; Sun, J.Z. Cell-free systems for biosynthesis: Towards a sustainable and economical approach. Green Chem. 2023, 25, 4912–4940. [Google Scholar] [CrossRef]
- Yue, K.; Chen, J.Y.; Li, Y.Q.; Kai, L. Advancing synthetic biology through cell-free protein synthesis. Comp. Struct. Biotechnol. J. 2023, 21, 2899–2908. [Google Scholar] [CrossRef]
- Harrison, R.P.; Ruck, S.; Medcalf, N.; Rafiq, Q.A. Decentralized manufacturing of cell and gene therapies: Overcoming challenges and identifying opportunities. Cytotherapy 2017, 19, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.P.; Ruck, S.; Rafiq, Q.A.; Medcalf, N. Decentralised manufacturing of cell and gene therapy products: Learning from other healthcare sectors. Biotechnol. Adv. 2018, 36, 345–357. [Google Scholar] [CrossRef]
- Bazmi, A.A.; Zahedi, G.; Hashim, H. Progress and challenges in utilization of palm oil biomass as fuel for decentralized electricity generation. Renew. Sust. Energ. Rev. 2011, 15, 574–583. [Google Scholar] [CrossRef]
- Hu, V.T.; Kamat, N.P. Cell-free protein synthesis systems for vaccine design and production. Curr. Opin. Biotechnol. 2023, 79, 102888. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, S.J.; Kim, D.M. The Use of Cell-free Protein Synthesis to Push the Boundaries of Synthetic Biology. Biotechnol. Bioprocess Eng. 2023, 28, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, M.W.; Matthaei, J.H. The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. Natl. Acad. Sci. USA 1961, 47, 1588–1602. [Google Scholar] [CrossRef]
- Endo, Y.; Otsuzuki, S.; Ito, K.; Miura, K.-i. Production of an enzymatic active protein using a continuous flow cell-free translation system. J. Biotechnol. 1992, 25, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-M.; Choi, C.-Y. A Semicontinuous Prokaryotic Coupled Transcription/Translation System Using a Dialysis Membrane. Biotechnol. Prog. 1996, 12, 645–649. [Google Scholar] [CrossRef]
- Kim, T.-W.; Kim, D.-M.; Choi, C.-Y. Rapid production of milligram quantities of proteins in a batch cell-free protein synthesis system. J. Biotechnol. 2006, 124, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, K.A.; Swartz, J.R. Energizing cell-free protein synthesis with glucose metabolism. Biotechnol. Bioeng. 2005, 90, 606–613. [Google Scholar] [CrossRef]
- Yunker, P.J.; Asahara, H.; Hung, K.-C.; Landry, C.; Arriaga, L.R.; Akartuna, I.; Heyman, J.; Chong, S.; Weitz, D.A. One-pot system for synthesis, assembly, and display of functional single-span membrane proteins on oil–water interfaces. Proc. Natl. Acad. Sci. USA 2016, 113, 608–613. [Google Scholar] [CrossRef]
- Peruzzi, J.A.; Galvez, N.R.; Kamat, N.P. Engineering transmembrane signal transduction in synthetic membranes using two-component systems. Proc. Natl. Acad. Sci. USA 2023, 120, e2218610120. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.L.; Boyd, M.A.; Kamat, N.P. Diblock copolymers enhance folding of a mechanosensitive membrane protein during cell-free expression. Proc. Natl. Acad. Sci. USA 2019, 116, 4031–4036. [Google Scholar] [CrossRef]
- Dudley, Q.M.; Karim, A.S.; Jewett, M.C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnol. J. 2015, 10, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.S.; Liu, W.Q.; Huang, S.H.; Xu, H.L.; Lu, H.F.; Wu, C.Z.; Li, J. Cell-free metabolic engineering enables selective biotransformation of fatty acids to value-added chemicals. Metab. Eng. Commun. 2023, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Swartz, J.R. Expanding biological applications using cell-free metabolic engineering: An overview. Metab. Eng. 2018, 50, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.E.; Jewett, M.C. Lysate of engineered Escherichia coli supports high-level conversion of glucose to 2,3-butanediol. Metab. Eng. 2015, 32, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Grubbe, W.S.; Rasor, B.J.; Krüger, A.; Jewett, M.C.; Karim, A.S. Cell-free styrene biosynthesis at high titers. Metab. Eng. 2020, 61, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Dudley, Q.M.; Karim, A.S.; Nash, C.J.; Jewett, M.C. In vitro prototyping of limonene biosynthesis using cell-free protein synthesis. Metab. Eng. 2020, 61, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Dudley, Q.M.; Anderson, K.C.; Jewett, M.C. Cell-Free Mixing of Escherichia coli Crude Extracts to Prototype and Rationally Engineer High-Titer Mevalonate Synthesis. ACS Synth. Biol. 2016, 5, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.S.; Jewett, M.C. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery. Metab. Eng. 2016, 36, 116–126. [Google Scholar] [CrossRef]
- Kelwick, R.; Ricci, L.; Chee, S.M.; Bell, D.; Webb, A.J.; Freemont, P.S. Cell-free prototyping strategies for enhancing the sustainable production of polyhydroxyalkanoates bioplastics. Synth. Biol. 2018, 3, ysy016. [Google Scholar] [CrossRef]
- Niu, F.-X.; Huang, Y.-B.; Shen, Y.-P.; Ji, L.-N.; Liu, J.-Z. Enhanced Production of Pinene by Using a Cell-Free System with Modular Cocatalysis. J. Agric. Food Chem. 2020, 68, 2139–2145. [Google Scholar] [CrossRef]
- Niu, F.-X.; Yan, Z.-B.; Huang, Y.-B.; Liu, J.-Z. Cell-free Biosynthesis of Chlorogenic Acid Using a Mixture of Chassis Cell Extracts and Purified Spy-Cyclized Enzymes. J. Agric. Food Chem. 2021, 69, 7938–7947. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Huang, S.; Liu, W.-Q.; Karim, A.S.; Jewett, M.C.; Li, J. Total in vitro biosynthesis of the nonribosomal macrolactone peptide valinomycin. Metab. Eng. 2020, 60, 37–44. [Google Scholar] [CrossRef]
- Ditzel, A.; Zhao, F.; Gao, X.; Phillips, G.N., Jr. Utilizing a cell-free protein synthesis platform for the biosynthesis of a natural product, caffeine. Synth. Biol. 2023, 8, ysad017. [Google Scholar] [CrossRef] [PubMed]
- Sitaraman, K.; Esposito, D.; Klarmann, G.; Le Grice, S.F.; Hartley, J.L.; Chatterjee, D.K. A novel cell-free protein synthesis system. J. Biotechnol. 2004, 110, 257–263. [Google Scholar] [CrossRef]
- Yim, S.S.; Johns, N.I.; Noireaux, V.; Wang, H.H. Protecting Linear DNA Templates in Cell-Free Expression Systems from Diverse Bacteria. ACS Synth. Biol. 2020, 9, 2851–2855. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, M.; Panfilov, S.; Pardee, K. High-Efficiency Protection of Linear DNA in Cell-Free Extracts from Escherichia coli and Vibrio natriegens. ACS Synth. Biol. 2021, 10, 1615–1624. [Google Scholar] [CrossRef]
- Sun, Z.Z.; Yeung, E.; Hayes, C.A.; Noireaux, V.; Murray, R.M. Linear DNA for Rapid Prototyping of Synthetic Biological Circuits in an Escherichia coli Based TX-TL Cell-Free System. ACS Synth. Biol. 2014, 3, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Niederholtmeyer, H.; Sun, Z.Z.; Hori, Y.; Yeung, E.; Verpoorte, A.; Murray, R.M.; Maerkl, S.J. Rapid cell-free forward engineering of novel genetic ring oscillators. eLife 2015, 4, e09771. [Google Scholar] [CrossRef] [PubMed]
- Schinn, S.-M.; Broadbent, A.; Bradley, W.T.; Bundy, B.C. Protein synthesis directly from PCR: Progress and applications of cell-free protein synthesis with linear DNA. New Biotech. 2016, 33, 480–487. [Google Scholar] [CrossRef]
- Rajput, H.; Changotra, R.; Rajput, P.; Gautam, S.; Gollakota, A.R.K.; Arora, A.S. A shock like no other: Coronavirus rattles commodity markets. Environ. Dev. Sustain. 2021, 23, 6564–6575. [Google Scholar] [CrossRef]
- Kis, Z.; Kontoravdi, C.; Shattock, R.; Shah, N.I. Resources, Production Scales and Time Required for Producing RNA Vaccines for the Global Pandemic Demand. Vaccines 2021, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Wilding, K.M.; Zhao, E.L.; Earl, C.C.; Bundy, B.C. Thermostable lyoprotectant-enhanced cell-free protein synthesis for on-demand endotoxin-free therapeutic production. New Biotech. 2019, 53, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Ding, X.; Lu, Y. Development of a robust Escherichia coli-based cell-free protein synthesis application platform. Biochem. Eng. J. 2021, 165, 107830. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.K.; Alam, K.K.; Verosloff, M.S.; Capdevila, D.A.; Desmau, M.; Clauer, P.R.; Lee, J.W.; Nguyen, P.Q.; Pastén, P.A.; Matiasek, S.J.; et al. Cell-free biosensors for rapid detection of water contaminants. Nat. Biotechnol. 2020, 38, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Pardee, K.; Green, A.A.; Ferrante, T.; Cameron, D.E.; DaleyKeyser, A.; Yin, P.; Collins, J.J. Paper-based synthetic gene networks. Cell 2014, 159, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Chavez, F.; Arce, A.; Adhikari, A.; Vadhin, S.; Pedroza-Garcia, J.A.; Gandini, C.; Ajioka, J.W.; Molloy, J.; Sanchez-Nieto, S.; Varner, J.D.; et al. Constructing Cell-Free Expression Systems for Low-Cost Access. ACS Synth. Biol. 2022, 11, 1114–1128. [Google Scholar] [CrossRef]
- Warfel, K.F.; Williams, A.; Wong, D.A.; Sobol, S.E.; Desai, P.; Li, J.; Chang, Y.F.; DeLisa, M.P.; Karim, A.S.; Jewett, M.C. A Low-Cost, Thermostable, Cell-Free Protein Synthesis Platform for On-Demand Production of Conjugate Vaccines. ACS Synth. Biol. 2023, 12, 95–107. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-J.; Kim, D.-M. Cell-Free Synthesis: Expediting Biomanufacturing of Chemical and Biological Molecules. Molecules 2024, 29, 1878. https://doi.org/10.3390/molecules29081878
Lee S-J, Kim D-M. Cell-Free Synthesis: Expediting Biomanufacturing of Chemical and Biological Molecules. Molecules. 2024; 29(8):1878. https://doi.org/10.3390/molecules29081878
Chicago/Turabian StyleLee, So-Jeong, and Dong-Myung Kim. 2024. "Cell-Free Synthesis: Expediting Biomanufacturing of Chemical and Biological Molecules" Molecules 29, no. 8: 1878. https://doi.org/10.3390/molecules29081878
APA StyleLee, S.-J., & Kim, D.-M. (2024). Cell-Free Synthesis: Expediting Biomanufacturing of Chemical and Biological Molecules. Molecules, 29(8), 1878. https://doi.org/10.3390/molecules29081878