
Lipidomic Analysis of Liver and Adipose Tissue in a High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Mice Model Reveals Alterations in Lipid Metabolism by Weight Loss and Aerobic Exercise

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
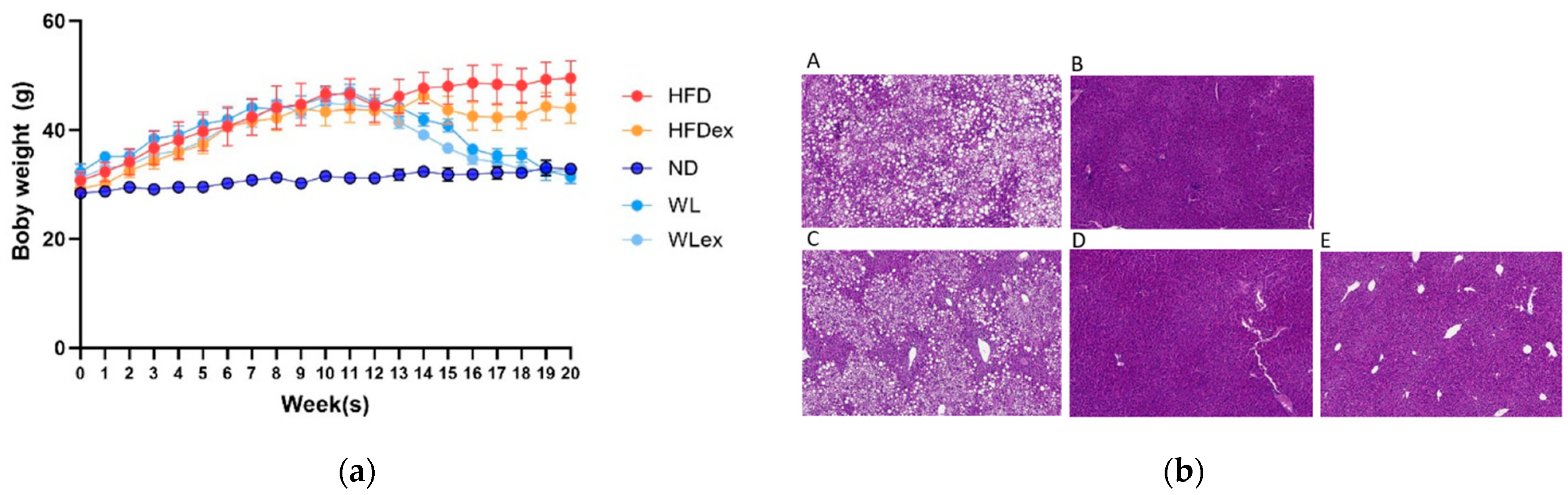
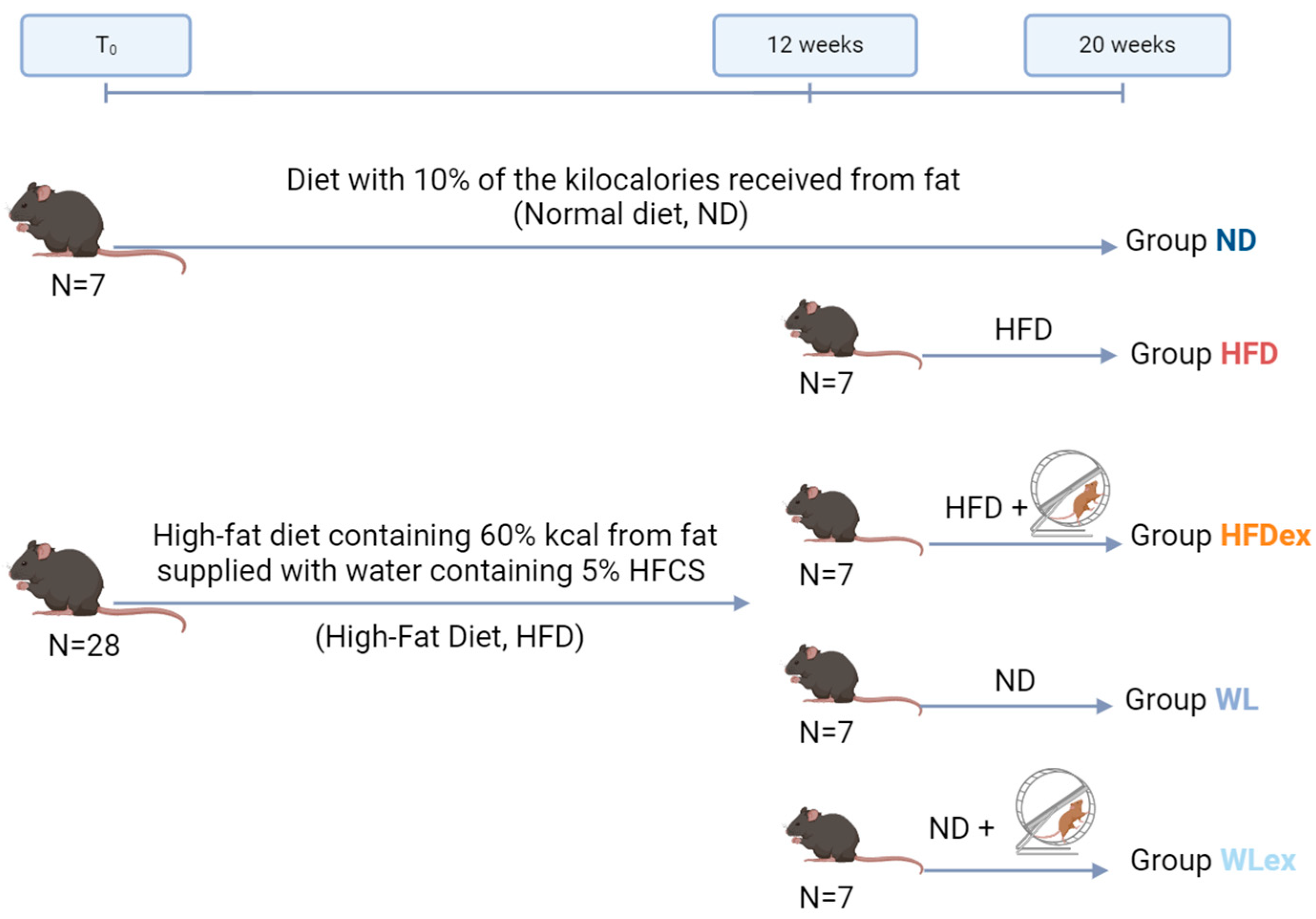
2.1. Animal Study
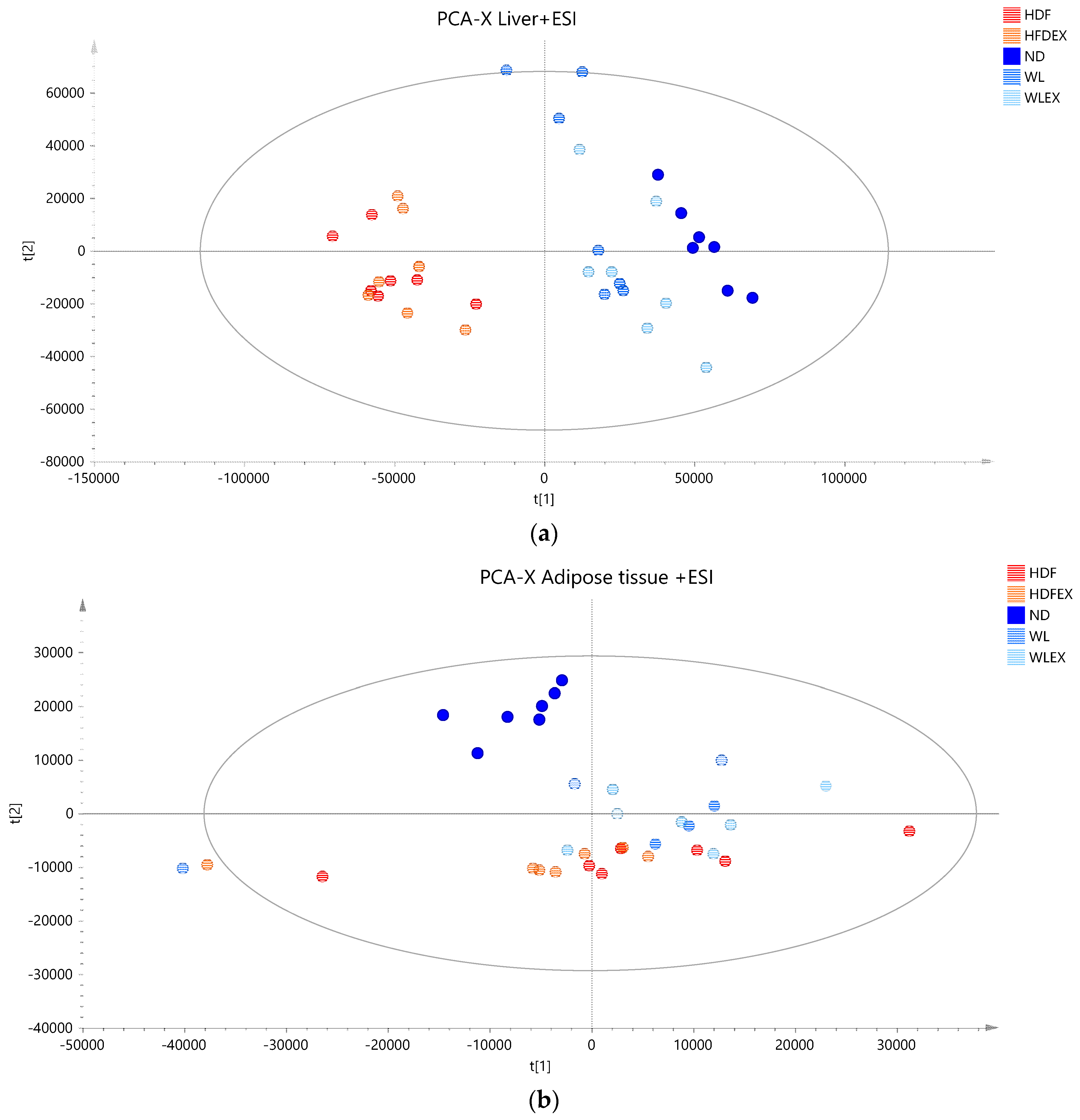
2.2. LC–TOF–MS Lipidomics Data
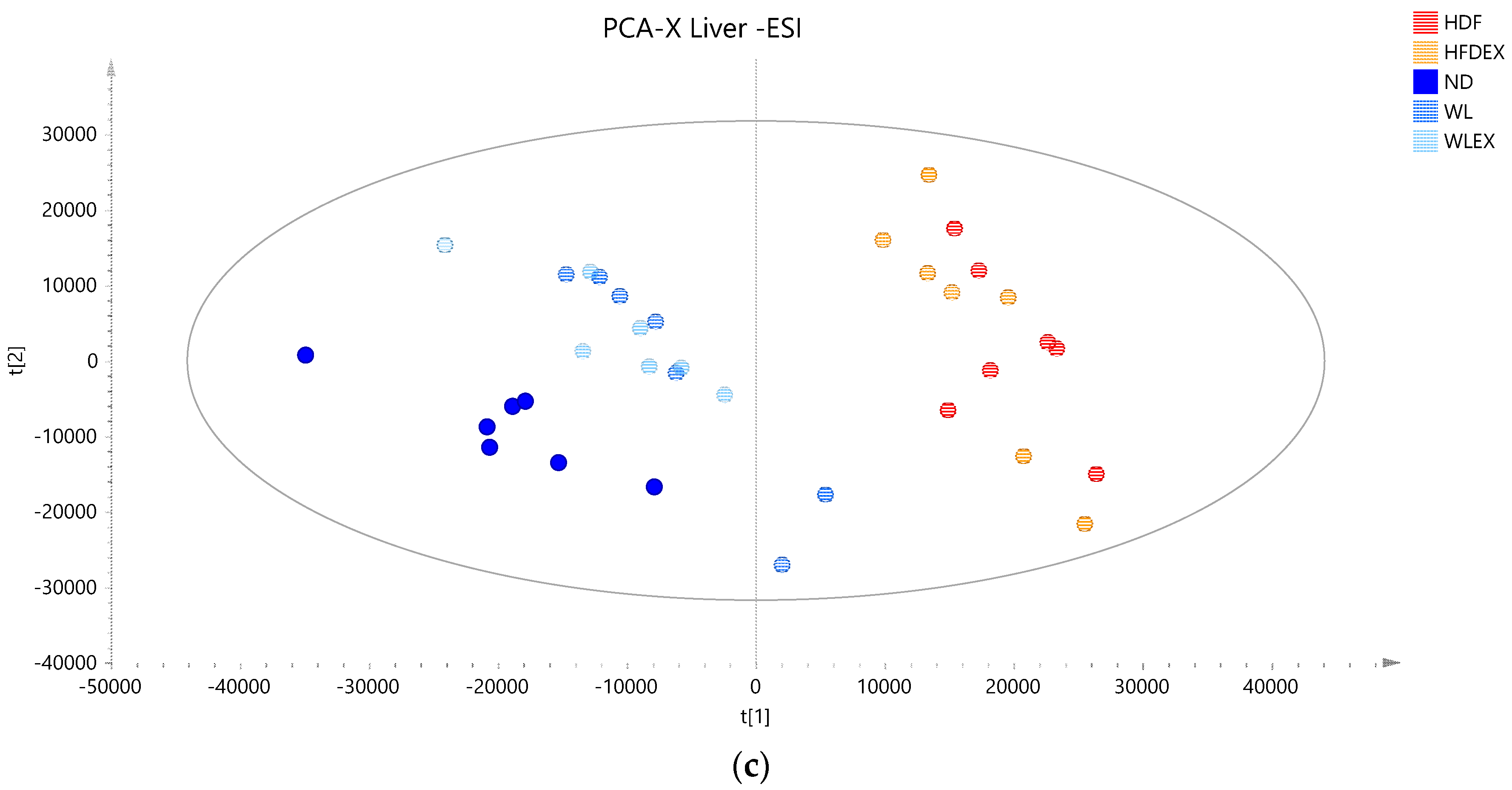
2.3. Hepatic Lipids Profile Reveal Alterations in Major Lipid Classes with Diet
2.3.1. Hepatic Phospholipids
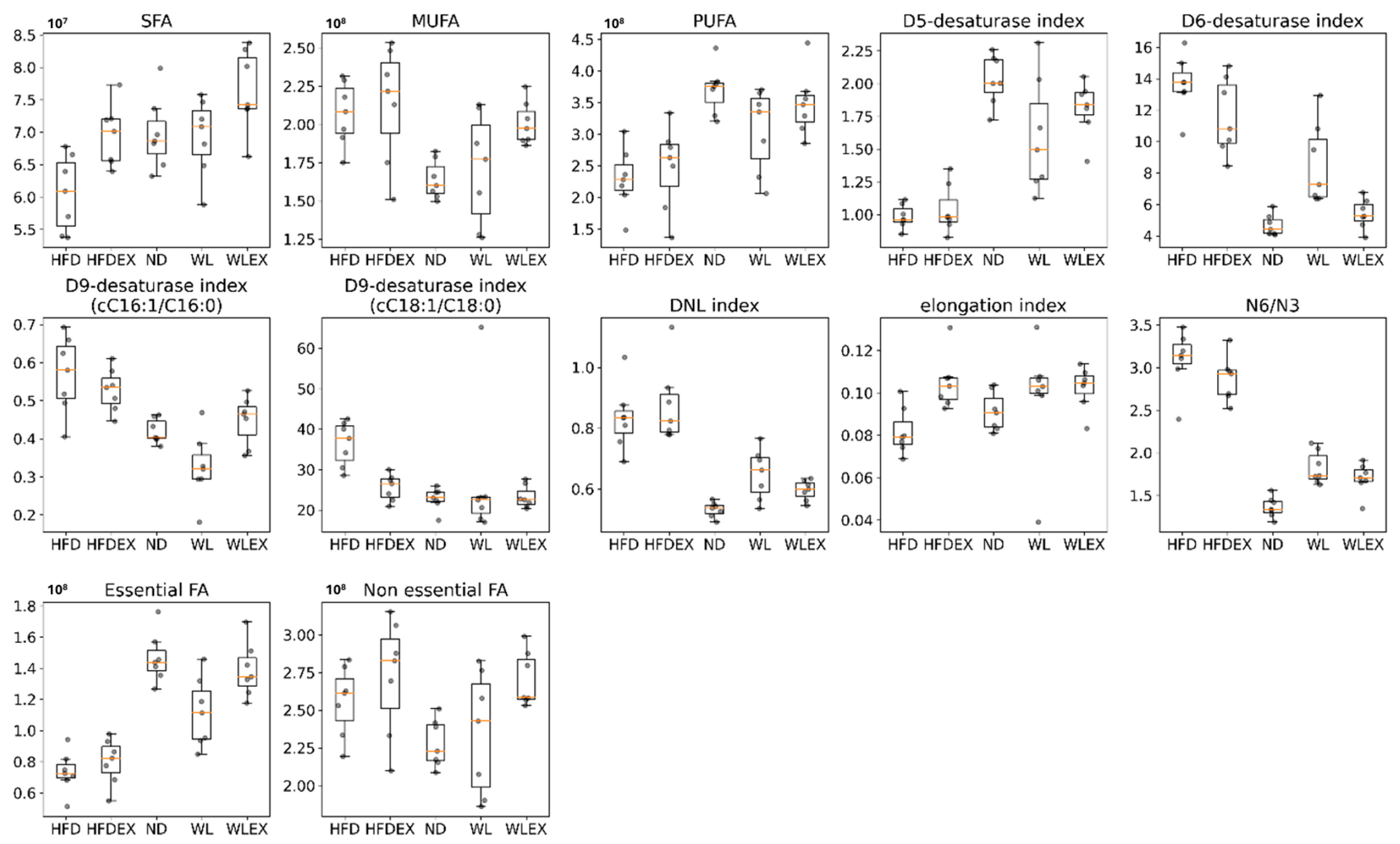
2.3.2. Fatty Acids Dysregulation in Hepatic Tissue
2.3.3. Glycerolipid and Sphingomyelins Dysregulation
2.4. Visceral Adipose Tissue Triglyceride Profile
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Animal Study
4.3. Lipidomic Analysis
4.3.1. Liver and Visceral Adipose Tissue Extraction
4.3.2. Analytical Instrumentation and Conditions
4.3.3. Data Analysis
4.3.4. Lipids’ Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lazarus, J.V.; Mark, H.E.; Anstee, Q.M.; Arab, J.P.; Batterham, R.L.; Castera, L.; Cortez-Pinto, H.; Crespo, J.; Cusi, K.; Dirac, M.A.; et al. Advancing the Global Public Health Agenda for NAFLD: A Consensus Statement. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Huby, T.; Gautier, E.L. Immune Cell-Mediated Features of Non-Alcoholic Steatohepatitis. Nat. Rev. Immunol. 2022, 22, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Han, S.K.; Baik, S.K.; Kim, M.Y. Non-Alcoholic Fatty Liver Disease: Definition and Subtypes. Clin. Mol. Hepatol. 2023, 29, S5–S16. [Google Scholar] [CrossRef] [PubMed]
- Nseir, W.; Hellou, E.; Assy, N. Role of Diet and Lifestyle Changes in Nonalcoholic Fatty Liver Disease. World J. Gastroenterol. 2014, 20, 9338–9344. [Google Scholar] [PubMed]
- van der Windt, D.J.; Sud, V.; Zhang, H.; Tsung, A.; Huang, H. The Effects of Physical Exercise on Fatty Liver Disease. Gene Expr. 2018, 18, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Zarghamravanbakhsh, P.; Frenkel, M.; Poretsky, L. Metabolic Causes and Consequences of Nonalcoholic Fatty Liver Disease (NAFLD). Metab. Open 2021, 12, 100149. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Lang, S.; Goeser, T.; Demir, M.; Steffen, H.-M.; Kasper, P. Management of Dyslipidemia in Patients with Non-Alcoholic Fatty Liver Disease. Curr. Atheroscler. Rep. 2022, 24, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Lui, D.T.; Lam, K.S. Non-alcoholic Fatty Liver Disease and Type 2 Diabetes: An Update. J. Diabetes Investig. 2022, 13, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Francque, S.M.A.; Dirinck, E. NAFLD Prevalence and Severity in Overweight and Obese Populations. Lancet Gastroenterol. Hepatol. 2023, 8, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Loomba, R. From NAFLD to MASLD: Implications of the New Nomenclature for Preclinical and Clinical Research. Nat. Metab. 2024, 1–3. [Google Scholar] [CrossRef]
- Eng, J.M.; Estall, J.L. Diet-Induced Models of Non-Alcoholic Fatty Liver Disease: Food for Thought on Sugar, Fat, and Cholesterol. Cells 2021, 10, 1805. [Google Scholar] [CrossRef] [PubMed]
- Sigala, D.M.; Hieronimus, B.; Medici, V.; Lee, V.; Nunez, M.V.; Bremer, A.A.; Cox, C.L.; Price, C.A.; Benyam, Y.; Chaudhari, A.J.; et al. Consuming Sucrose- or HFCS-Sweetened Beverages Increases Hepatic Lipid and Decreases Insulin Sensitivity in Adults. J. Clin. Endocrinol. Metab. 2021, 106, 3248–3264. [Google Scholar] [CrossRef]
- Jensen, T.; Abdelmalek, M.F.; Sullivan, S.; Nadeau, K.J.; Green, M.; Roncal, C.; Nakagawa, T.; Kuwabara, M.; Sato, Y.; Kang, D.-H.; et al. Fructose and Sugar: A Major Mediator of Nonalcoholic Fatty Liver Disease. J. Hepatol. 2018, 68, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, G.; Legaki, A.-I.; Georgila, K.; Vorkas, P.; Giannousi, E.; Stamatakis, G.; Moustakas, I.I.; Petrocheilou, M.; Pyrina, I.; Gercken, B.; et al. Integrated Omics Analysis for Characterization of the Contribution of High Fructose Corn Syrup to Non-Alcoholic Fatty Liver Disease in Obesity. Metabolism 2023, 144, 155552. [Google Scholar] [CrossRef] [PubMed]
- Finer, N. Weight Loss Interventions and Nonalcoholic Fatty Liver Disease: Optimizing Liver Outcomes. Diabetes Obes. Metab. 2022, 24, 44–54. [Google Scholar] [CrossRef]
- Wong, V.W.-S.; Wong, G.L.-H.; Chan, R.S.-M.; Shu, S.S.-T.; Cheung, B.H.-K.; Li, L.S.; Chim, A.M.-L.; Chan, C.K.-M.; Leung, J.K.-Y.; Chu, W.C.-W.; et al. Beneficial Effects of Lifestyle Intervention in Non-Obese Patients with Non-Alcoholic Fatty Liver Disease. J. Hepatol. 2018, 69, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Vilar-Gomez, E.; Martinez-Perez, Y.; Calzadilla-Bertot, L.; Torres-Gonzalez, A.; Gra-Oramas, B.; Gonzalez-Fabian, L.; Friedman, S.L.; Diago, M.; Romero-Gomez, M. Weight Loss Through Lifestyle Modification Significantly Reduces Features of Nonalcoholic Steatohepatitis. Gastroenterology 2015, 149, 367–378.e5, quiz e14–15. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Lindor, K.; St Saver, J.; Lymp, J.; Mendes, F.; Muto, A.; Okada, T.; Angulo, P. Effect of Changes on Body Weight and Lifestyle in Nonalcoholic Fatty Liver Disease. J. Hepatol. 2005, 43, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Hickman, I.J.; Jonsson, J.R.; Prins, J.B.; Ash, S.; Purdie, D.M.; Clouston, A.D.; Powell, E.E. Modest Weight Loss and Physical Activity in Overweight Patients with Chronic Liver Disease Results in Sustained Improvements in Alanine Aminotransferase, Fasting Insulin, and Quality of Life. Gut 2004, 53, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Shama, S.; Jang, H.; Wang, X.; Zhang, Y.; Shahin, N.N.; Motawi, T.K.; Kim, S.; Gawrieh, S.; Liu, W. Phosphatidylethanolamines Are Associated with Nonalcoholic Fatty Liver Disease (NAFLD) in Obese Adults and Induce Liver Cell Metabolic Perturbations and Hepatic Stellate Cell Activation. Int. J. Mol. Sci. 2023, 24, 1034. [Google Scholar] [CrossRef]
- Knebel, B.; Fahlbusch, P.; Dille, M.; Wahlers, N.; Hartwig, S.; Jacob, S.; Kettel, U.; Schiller, M.; Herebian, D.; Koellmer, C.; et al. Fatty Liver Due to Increased de Novo Lipogenesis: Alterations in the Hepatic Peroxisomal Proteome. Front. Cell Dev. Biol. 2019, 7, 248. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, S.; Binienda, A.; Jabłkowski, M.; Szemraj, J.; Czarny, P. The Interplay between Insulin Resistance, Inflammation, Oxidative Stress, Base Excision Repair and Metabolic Syndrome in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2021, 22, 11128. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Xu, J.-W.; Li, S.; Ng, X.E.; Tung, Y.-T. Effects of Exercise on High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease and Lipid Metabolism in ApoE Knockout Mice. Nutr. Metab. 2022, 19, 10. [Google Scholar] [CrossRef]
- Poole, D.C.; Copp, S.W.; Colburn, T.D.; Craig, J.C.; Allen, D.L.; Sturek, M.; O’Leary, D.S.; Zucker, I.H.; Musch, T.I. Guidelines for Animal Exercise and Training Protocols for Cardiovascular Studies. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, H1100–H1138. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Ezagouri, S.; Dandavate, V.; Asher, G. Monitoring Daytime Differences in Moderate Intensity Exercise Capacity Using Treadmill Test and Muscle Dissection. STAR Protoc. 2021, 2, 100331. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Herrera, J.J.; Guseh, J.S.; Atlason, B.; Houstis, N.E.; Abdul Kadir, A.; Li, H.; Sheffield, C.; Singh, A.P.; Roh, J.D.; et al. Animal Models of Exercise from Rodents to Pythons. Circ. Res. 2022, 130, 1994–2014. [Google Scholar] [CrossRef] [PubMed]
- PubMed. Physical Activity Protocols in Non-Alcoholic Fatty Liver Disease Management: A Systematic Review of Randomized Clinical Trials and Animal Models. Available online: https://pubmed.ncbi.nlm.nih.gov/37510432/ (accessed on 19 November 2023).
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The Critical Role of Phosphatidylcholine and Phosphatidylethanolamine Metabolism in Health and Disease. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Béland-Bonenfant, S.; Rouland, A.; Petit, J.-M.; Vergès, B. Concise Review of Lipidomics in Nonalcoholic Fatty Liver Disease. Diabetes Metab. 2023, 49, 101432. [Google Scholar] [CrossRef] [PubMed]
- Ščupáková, K.; Soons, Z.; Ertaylan, G.; Pierzchalski, K.A.; Eijkel, G.B.; Ellis, S.R.; Greve, J.W.; Driessen, A.; Verheij, J.; De Kok, T.M.; et al. Spatial Systems Lipidomics Reveals Nonalcoholic Fatty Liver Disease Heterogeneity. Anal. Chem. 2018, 90, 5130–5138. [Google Scholar] [CrossRef]
- Orešič, M.; Hyötyläinen, T.; Kotronen, A.; Gopalacharyulu, P.; Nygren, H.; Arola, J.; Castillo, S.; Mattila, I.; Hakkarainen, A.; Borra, R.J.H.; et al. Prediction of Non-Alcoholic Fatty-Liver Disease and Liver Fat Content by Serum Molecular Lipids. Diabetologia 2013, 56, 2266–2274. [Google Scholar] [CrossRef]
- Masoodi, M.; Gastaldelli, A.; Hyötyläinen, T.; Arretxe, E.; Alonso, C.; Gaggini, M.; Brosnan, J.; Anstee, Q.M.; Millet, O.; Ortiz, P.; et al. Metabolomics and Lipidomics in NAFLD: Biomarkers and Non-Invasive Diagnostic Tests. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 835–856. [Google Scholar] [CrossRef]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.-K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The Plasma Lipidomic Signature of Nonalcoholic Steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar] [CrossRef] [PubMed]
- Gorden, D.L.; Myers, D.S.; Ivanova, P.T.; Fahy, E.; Maurya, M.R.; Gupta, S.; Min, J.; Spann, N.J.; McDonald, J.G.; Kelly, S.L.; et al. Biomarkers of NAFLD Progression: A Lipidomics Approach to an Epidemic1[S]. J. Lipid Res. 2015, 56, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Catalá, A. Five Decades with Polyunsaturated Fatty Acids: Chemical Synthesis, Enzymatic Formation, Lipid Peroxidation and Its Biological Effects. J. Lipids 2013, 2013, e710290. [Google Scholar] [CrossRef] [PubMed]
- Araya, J.; Rodrigo, R.; Pettinelli, P.; Araya, A.V.; Poniachik, J.; Videla, L.A. Decreased Liver Fatty Acid Delta-6 and Delta-5 Desaturase Activity in Obese Patients. Obesity 2010, 18, 1460–1463. [Google Scholar] [CrossRef]
- Mäkelä, T.N.K.; Tuomainen, T.-P.; Hantunen, S.; Virtanen, J.K. Associations of Serum N-3 and n-6 Polyunsaturated Fatty Acids with Prevalence and Incidence of Nonalcoholic Fatty Liver Disease. Am. J. Clin. Nutr. 2022, 116, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Samovski, D.; Smith, G.I.; Cifarelli, V.; Farabi, S.S.; Yoshino, J.; Pietka, T.; Chang, S.-W.; Ghosh, S.; Myckatyn, T.M.; et al. Associations Among Adipose Tissue Immunology, Inflammation, Exosomes and Insulin Sensitivity in People with Obesity and Nonalcoholic Fatty Liver Disease. Gastroenterology 2021, 161, 968–981.e12. [Google Scholar] [CrossRef]
- Guerra, S.; Mocciaro, G.; Gastaldelli, A. Adipose Tissue Insulin Resistance and Lipidome Alterations as the Characterizing Factors of Non-alcoholic Steatohepatitis. Eur. J. Clin. Investig. 2022, 52, e13695. [Google Scholar] [CrossRef] [PubMed]
- Rosso, C.; Kazankov, K.; Younes, R.; Esmaili, S.; Marietti, M.; Sacco, M.; Carli, F.; Gaggini, M.; Salomone, F.; Møller, H.J.; et al. Crosstalk between Adipose Tissue Insulin Resistance and Liver Macrophages in Non-Alcoholic Fatty Liver Disease. J. Hepatol. 2019, 71, 1012–1021. [Google Scholar] [CrossRef]
- Lange, M.; Angelidou, G.; Ni, Z.; Criscuolo, A.; Schiller, J.; Blüher, M.; Fedorova, M. AdipoAtlas: A Reference Lipidome for Human White Adipose Tissue. Cell Rep. Med. 2021, 2, 100407. [Google Scholar] [CrossRef]
- Cockcroft, S. Mammalian Lipids: Structure, Synthesis and Function. Essays Biochem. 2021, 65, 813–845. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, M.; Patel, P.; Dunn-Valadez, S.; Dao, C.; Khan, V.; Ali, H.; El-Serag, L.; Hernaez, R.; Sisson, A.; Thrift, A.P.; et al. Women Have Lower Risk of Nonalcoholic Fatty Liver Disease but Higher Risk of Progression vs Men: A Systematic Review and Meta-Analysis. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2021, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Illana, Á.; Piñeiro-Ramos, J.D.; Sanjuan-Herráez, J.D.; Vento, M.; Quintás, G.; Kuligowski, J. Evaluation of Batch Effect Elimination Using Quality Control Replicates in LC-MS Metabolite Profiling. Anal. Chim. Acta 2018, 1019, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Goracci, L.; Tortorella, S.; Tiberi, P.; Pellegrino, R.M.; Di Veroli, A.; Valeri, A.; Cruciani, G. Lipostar, a Comprehensive Platform-Neutral Cheminformatics Tool for Lipidomics. Anal. Chem. 2017, 89, 6257–6264. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HFD-ND | HFD-WL | HFD-WLEX | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Lipids | p-Value | Log2FC | VIP | p-Value | Log2FC | VIP | p-Value | Log2FC | VIP | CV% |
| DG 36:2 | 8.03 × 10−5 | −3.61 | 4.2 | 9.75 × 10−3 | −1.43 | 4.1 | 2.44 × 10−1 | −0.96 | 3.4 | 1.53 |
| DG 36:3 | 2.55 × 10−2 | 0.27 | 6.8 | 1.00 × 100 | −0.04 | 3.6 | 7.42 × 10−2 | 0.24 | 7.2 | 0.89 |
| DG 36:4 | 3.78 × 10−5 | 2.13 | 11 | 1.00 × 100 | 0.94 | 5.9 | 3.81 × 10−2 | 1.56 | 9.9 | 1.03 |
| DG 38:2 | 3.24 × 10−5 | −1.81 | 4.8 | 3.12 × 10−2 | −1.31 | 5.2 | 1.63 × 10−1 | −1.17 | 4.7 | 1.79 |
| DG 38:3 | 3.45 × 10−2 | −0.96 | 4.2 | 3.24 × 10−5 | −1.58 | 5.9 | 1.50 × 10−1 | −0.88 | 4.5 | 6.21 |
| DG 38:4 | 5.07 × 10−4 | −1.66 | 4.3 | 3.44 × 10−3 | −1.58 | 5.1 | 1.77 × 10−1 | −0.98 | 4.1 | 2.06 |
| DG 40:7 | 1.51 × 10−2 | −0.51 | 5.3 | 1.00 × 100 | −0.11 | 2.9 | 7.61 × 10−1 | −0.29 | 8.4 | 18.7 |
| DG 40:8 | 2.37 × 10−5 | 5.03 | 8.0 | 3.07 × 10−1 | 3.50 | 4.5 | 5.61 × 10−2 | 4.12 | 6.2 | 18.3 |
| FA 16:0 | 1.92 × 10−1 | 0.18 | 4.6 | 2.64 × 10−1 | 0.17 | 5.4 | 2.11 × 10−3 | 0.31 | 7.2 | 4.68 |
| FA 16:1 | 5.85 × 10−1 | −0.23 | 3.3 | 2.55 × 10−2 | −0.60 | 6.6 | 1.00 × 100 | −0.02 | 1.1 | 3.43 |
| FA 18:1 | 2.55 × 10−2 | −0.32 | 9.5 | 9.17 × 10−1 | −0.18 | 8.8 | 1.00 × 100 | −0.01 | 5.4 | 5.26 |
| FA 18:2 | 8.64 × 10−4 | 0.82 | 12 | 2.85 × 10−1 | 0.53 | 11 | 4.37 × 10−3 | 0.77 | 14 | 8.73 |
| FA 18:3 | 1.08 × 10−4 | 2.38 | 8.1 | 4.76 × 10−1 | 1.32 | 5.7 | 2.70 × 10−3 | 2.13 | 8.4 | 3.38 |
| FA 18:4 | 1.92 × 10−4 | 3.26 | 2.5 | 4.76 × 10−1 | 2.11 | 1.8 | 1.64 × 10−3 | 3.18 | 2.7 | 3.51 |
| FA 20:1 | 1.64 × 10−3 | −1.06 | 4.2 | 6.95 × 10−3 | −1.05 | 5.2 | 9.72 × 10−2 | −0.81 | 4.3 | 12.4 |
| FA 20:2 | 7.57 × 10−4 | −1.27 | 3.7 | 2.30 × 10−2 | −1.06 | 4.2 | 6.16 × 10−2 | −0.94 | 3.8 | 2.78 |
| FA 20:3 | 2.55 × 10−4 | −1.36 | 3.7 | 1.38 × 10−1 | −0.85 | 3.8 | 1.92 × 10−1 | −0.69 | 3.3 | 3.99 |
| FA 20:5 | 5.96 × 10−5 | 2.95 | 11 | 1.77 × 10−1 | 2.07 | 9.1 | 2.82 × 10−2 | 2.56 | 11 | 2.58 |
| FA 22:3 | 3.78 × 10−5 | −2.86 | 2.4 | 1.16 × 10−1 | −1.79 | 2.7 | 4.20 × 10−2 | −2.08 | 2.6 | 5.01 |
| FA 22:4 | 9.75 × 10−3 | −1.03 | 3.7 | 5.61 × 10−2 | −0.96 | 4.4 | 2.30 × 10−2 | −0.95 | 4.4 | 3.68 |
| FA 22:5 | 3.05 × 10−3 | 1.54 | 6.5 | 5.61 × 10−2 | 1.31 | 6.4 | 3.05 × 10−3 | 1.54 | 7.6 | 3.86 |
| FA 22:6 | 7.57 × 10−4 | 0.87 | 10 | 6.76 × 10−2 | 0.63 | 10 | 6.76 × 10−2 | 0.65 | 9.9 | 2.43 |
| LPC 18:0 | 1.00 × 100 | 0.60 | 2.2 | 3.12 × 10−2 | 1.07 | 5.0 | 1.00 × 100 | 0.34 | 2.2 | 4.23 |
| LPC 18:2 | 8.12 × 10−2 | 1.25 | 5.2 | 6.20 × 10−3 | 1.41 | 7.2 | 2.39 × 10−3 | 1.44 | 6.5 | 3.09 |
| LPC 20:4 | 1.64 × 10−3 | −1.06 | 4.2 | 3.12 × 10−2 | −0.94 | 4.5 | 1.00 × 100 | −0.44 | 2.8 | 4.30 |
| LPE 18:2 | 6.20 × 10−3 | 2.13 | 2.9 | 2.30 × 10−2 | 1.85 | 3.2 | 5.61 × 10−2 | 1.77 | 2.7 | 2.68 |
| PC 32:0 | 1.64 × 10−3 | 0.81 | 5.5 | 3.45 × 10−2 | 0.65 | 5.8 | 1.50 × 10−1 | 0.44 | 3.3 | 12.0 |
| PC 32:1 | 2.85 × 10−1 | 0.22 | 2.5 | 1.87 × 10−2 | 0.37 | 4.4 | 4.63 × 10−2 | 0.29 | 3.3 | 4.38 |
| PC 34:1 | 6.20 × 10−3 | −0.45 | 3.8 | 1.00 × 100 | −0.01 | 0.5 | 6.26 × 10−1 | −0.23 | 2.9 | 1.98 |
| PC 34:2 | 1.26 × 10−5 | 0.88 | 5.5 | 7.42 × 10−2 | 0.64 | 5.5 | 1.26 × 10−1 | 0.61 | 4.9 | 2.66 |
| PC 34:3 | 3.45 × 10−2 | 0.64 | 2.9 | 1.00 × 100 | −0.04 | 0.5 | 1.00 × 100 | 0.29 | 1.3 | 3.60 |
| PC 36:2 | 1.12 × 10−3 | 0.87 | 4.7 | 4.91 × 10−3 | 0.85 | 5.7 | 4.13 × 10−1 | 0.55 | 3.7 | 1.63 |
| PC 36:3 | 1.09 × 10−2 | 0.91 | 3.3 | 1.00 × 100 | 0.46 | 1.6 | 8.71 × 10−3 | 0.96 | 3.9 | 2.63 |
| PC 36:4 | 2.03 × 10−5 | 1.99 | 4.7 | 3.45 × 10−2 | 1.30 | 3.8 | 3.57 × 10−1 | 0.98 | 2.8 | 4.42 |
| PC 36:5 | 1.07 × 10−5 | 3.58 | 2.7 | 1.26 × 10−1 | 2.00 | 2.0 | 8.12 × 10−2 | 2.43 | 2.0 | 2.69 |
| PC 38:3 | 2.93 × 10−4 | −1.58 | 8.0 | 1.00 × 100 | −0.24 | 3.1 | 4.20 × 10−2 | −0.89 | 6.9 | 1.50 |
| PC 38:5 | 5.07 × 10−4 | −0.94 | 5.5 | 3.81 × 10−2 | −0.46 | 5.4 | 2.30 × 10−2 | −0.62 | 5.3 | 3.27 |
| PC 40:5 | 6.76 × 10−2 | 1.25 | 2.8 | 8.71 × 10−3 | 1.33 | 3.3 | 1.35 × 10−2 | 1.36 | 3.4 | 3.35 |
| PC 40:6 | 1.00 × 100 | 0.07 | 3.6 | 9.75 × 10−3 | 0.20 | 7.1 | 1.00 × 100 | 0.09 | 3.9 | 2.97 |
| PC 40:8 | 3.12 × 10−2 | 0.96 | 5.5 | 3.05 × 10−3 | 1.09 | 7.4 | 1.51 × 10−2 | 0.99 | 6.3 | 1.27 |
| PE 34:2 | 5.80 × 10−4 | 1.57 | 5.1 | 8.89 × 10−2 | 1.19 | 5.4 | 4.44 × 10−1 | 0.95 | 3.8 | 1.39 |
| PE 36:2 | 3.78 × 10−5 | 1.34 | 5.2 | 1.87 × 10−2 | 1.07 | 5.4 | 2.26 × 10−1 | 0.94 | 4.7 | 1.57 |
| PE 36:3 | 9.31 × 10−5 | 1.35 | 4.6 | 3.57 × 10−1 | 0.81 | 3.9 | 1.26 × 10−1 | 0.80 | 3.1 | 1.25 |
| PE 36:4 | 6.92 × 10−5 | 2.26 | 11 | 9.72 × 10−2 | 2.06 | 12 | 3.45 × 10−2 | 2.07 | 11 | 0.95 |
| PE 36:5 | 1.21 × 10−2 | 2.23 | 2.8 | 2.39 × 10−3 | 2.33 | 3.6 | 2.07 × 10−2 | 2.16 | 3.2 | 3.64 |
| PE 38:5 | 6.68 × 10−1 | 1.05 | 2.6 | 3.86 × 10−4 | 1.64 | 4.9 | 1.35 × 10−2 | 1.32 | 3.8 | 1.08 |
| PE 38:6 | 3.86 × 10−4 | 0.79 | 5.9 | 1.77 × 10−1 | 0.42 | 4.8 | 1.63 × 10−1 | 0.42 | 4.4 | 1.83 |
| PE 38:7 | 9.31 × 10−5 | 1.22 | 2.8 | 3.31 × 10−1 | 0.67 | 2.1 | 1.38 × 10−1 | 0.74 | 2.1 | 1.30 |
| PE 40:6 | 3.45 × 10−2 | 0.60 | 2.9 | 2.30 × 10−2 | 0.50 | 3.1 | 1.16 × 10−1 | 0.43 | 2.5 | 1.43 |
| PE O-38:5 | 1.00 × 100 | −0.36 | 1.5 | 1.51 × 10−2 | −0.93 | 2.7 | 1.35 × 10−2 | −0.83 | 2.5 | 2.50 |
| PI 34:2 | 2.78 × 10−5 | 1.81 | 3.5 | 9.72 × 10−2 | 1.30 | 3.4 | 6.16 × 10−2 | 1.29 | 3.2 | 4.91 |
| PI 38:5 | 1.87 × 10−2 | 2.84 | 2.1 | 7.57 × 10−4 | 3.08 | 2.8 | 3.45 × 10−2 | 2.65 | 2.2 | 2.58 |
| PS 38:6 | 4.43 × 10−4 | 1.70 | 3.3 | 7.13 × 10−1 | 0.77 | 2.0 | 1.00 × 100 | 0.35 | 1.2 | 2.88 |
| SM 40:1;O2 | 2.55 × 10−4 | 1.48 | 4.8 | 5.10 × 10−1 | 0.79 | 3.4 | 1.00 × 100 | 0.50 | 2.5 | 4.90 |
| SM 41:1;O2 | 3.44 × 10−3 | 1.20 | 4.3 | 2.70 × 10−3 | 1.25 | 5.2 | 1.63 × 10−1 | 0.98 | 3.7 | 1.00 |
| SM 42:1;O2 | 2.78 × 10−5 | 1.23 | 6.7 | 2.85 × 10−1 | 0.98 | 6.6 | 1.68 × 10−2 | 1.07 | 6.6 | 6.28 |
| TG 52:2 | 4.43 × 10−4 | −1.35 | 4.1 | 2.85 × 10−1 | −0.71 | 3.6 | 4.44 × 10−1 | −0.69 | 2.7 | 12.6 |
| TG 52:4 | 6.20 × 10−3 | 0.53 | 4.3 | 1.00 × 100 | 0.23 | 2.3 | 2.08 × 10−1 | 0.45 | 3.9 | 1.65 |
| TG 56:8 | 2.08 × 10−1 | 0.92 | 3.2 | 7.79 × 10−3 | 1.12 | 4.2 | 6.76 × 10−2 | 0.90 | 3.3 | 5.10 |
| TG 56:9 | 3.24 × 10−5 | 1.63 | 3.9 | 4.13 × 10−1 | 1.00 | 2.6 | 1.38 × 10−1 | 1.24 | 3.4 | 3.78 |
| HFD-ND | HFD-WL | HFD-WLex | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compounds | p-Values | Log2FC | VIP | p-Values | Log2FC | VIP | p-Values | Log2FC | VIP | CV% |
| TG 38:1 | 1.00 × 100 | −0.01 | 0.3 | 7.44 × 10−3 | 2.87 | 2.6 | 2.60 × 10−1 | 1.19 | 1.0 | 5.13 |
| TG 38:2 | 2.82 × 10−1 | 1.52 | 0.6 | 5.94 × 10−4 | 3.57 | 3.0 | 5.71 × 10−2 | 2.00 | 1.3 | 4.75 |
| TG 40:1 | 1.00 × 100 | −0.01 | 0.2 | 5.17 × 10−3 | 3.04 | 2.8 | 1.87 × 10−1 | 1.42 | 0.9 | 5.77 |
| TG 40:3 | 1.32 × 10−1 | 2.01 | 0.7 | 6.79 × 10−4 | 3.33 | 3.0 | 1.17 × 10−2 | 2.42 | 1.3 | 3.92 |
| TG 42:2 | 1.00 × 100 | 0.43 | 0.2 | 2.74 × 10−3 | 2.93 | 2.8 | 2.27 × 10−2 | 1.89 | 1.0 | 6.83 |
| TG 50:4 | 2.40 × 10−4 | 1.72 | 3.1 | 2.19 × 10−1 | 0.91 | 1.6 | 4.14 × 10−1 | 0.82 | 2.4 | 4.12 |
| TG 50:5 | 4.51 × 10−5 | 3.23 | 1.2 | 1.21 × 10−1 | 1.95 | 2.6 | 2.60 × 10−1 | 1.68 | 0.8 | 6.50 |
| TG 51:3 | 5.63 × 10−4 | 1.20 | 1.8 | 1.00 × 10⁰ | 0.49 | 1.2 | 5.94 × 10−1 | 0.55 | 1.4 | 5.03 |
| TG 52:1 | 1.11 × 10−3 | −1.65 | 3.7 | 1.00 × 10⁰ | −0.26 | 0.9 | 9.44 × 10−1 | −0.56 | 3.6 | 3.97 |
| TG 52:2 | 1.66 × 10−3 | −0.90 | 8.1 | 1.00 × 10⁰ | −0.17 | 0.8 | 1.00 × 100 | −0.14 | 5.7 | 2.40 |
| TG 52:4 | 3.20 × 10−4 | 1.42 | 8.0 | 8.20 × 10−1 | 0.49 | 1.2 | 7.78 × 10−1 | 0.57 | 5.7 | 4.88 |
| TG 52:5 | 3.29 × 10−5 | 2.33 | 4.6 | 1.45 × 10−1 | 1.07 | 1.7 | 3.56 × 10−1 | 0.95 | 2.8 | 4.21 |
| TG 52:6 | 1.14 × 10−4 | 3.48 | 1.6 | 1.91 × 10−1 | 1.78 | 2.5 | 3.10 × 10−1 | 1.63 | 1.1 | 9.23 |
| TG 53:4 | 8.51 × 10−4 | 1.28 | 1.6 | 1.00 × 10⁰ | 0.34 | 1.1 | 6.36 × 10−1 | 0.59 | 1.2 | 4.42 |
| TG 54:2 | 1.66 × 10−3 | −2.04 | 5.3 | 1.00 × 10⁰ | −0.50 | 1.0 | 1.00 × 100 | −0.33 | 4.3 | 4.06 |
| TG 54:3 | 3.12 × 10−2 | −1.42 | 8.9 | 1.00 × 10⁰ | −0.13 | 0.7 | 1.00 × 100 | 0.21 | 5.6 | 2.94 |
| TG 54:4 | 2.02 × 10−5 | 2.22 | 1.6 | 2.43 × 10−1 | 1.09 | 1.7 | 1.11 × 10−1 | 1.19 | 1.4 | 3.68 |
| TG 54:5 | 1.14 × 10−4 | 1.84 | 7.2 | 1.00 × 10⁰ | 0.48 | 1.1 | 2.03 × 10−1 | 0.80 | 5.2 | 3.41 |
| TG 54:6 | 2.02 × 10−5 | 2.98 | 4.5 | 1.84 × 10−1 | 1.28 | 1.8 | 1.44 × 10−1 | 1.32 | 3.0 | 4.10 |
| TG 54:7 | 7.18 × 10−5 | 4.84 | 1.8 | 2.09 × 10−1 | 2.61 | 2.9 | 2.62 × 10−1 | 2.44 | 1.1 | 10.2 |
| TG 56:2 | 2.81 × 10−2 | −1.05 | 1.9 | 1.00 × 10⁰ | −0.06 | 0.6 | 1.00 × 100 | 0.07 | 1.1 | 3.19 |
| TG 56:3 | 2.38 × 10−5 | 2.63 | 2.2 | 1.99 × 10−1 | 1.22 | 1.8 | 1.72 × 10−1 | 1.24 | 1.6 | 3.54 |
| TG 56:4 | 1.66 × 10−3 | 0.95 | 1.6 | 6.35 × 10−1 | 0.55 | 1.1 | 1.21 × 10−1 | 0.70 | 1.9 | 3.21 |
| TG 56:5 | 2.44 × 10−3 | 1.81 | 1.5 | 8.91 × 10−1 | 1.06 | 1.4 | 1.72 × 10−1 | 1.48 | 1.9 | 6.52 |
| TG 56:6 | 2.38 × 10−5 | 2.63 | 2.2 | 1.99 × 10−1 | 1.22 | 1.8 | 1.72 × 10−1 | 1.24 | 1.6 | 3.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mouskeftara, T.; Deda, O.; Papadopoulos, G.; Chatzigeorgiou, A.; Gika, H. Lipidomic Analysis of Liver and Adipose Tissue in a High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Mice Model Reveals Alterations in Lipid Metabolism by Weight Loss and Aerobic Exercise. Molecules 2024, 29, 1494. https://doi.org/10.3390/molecules29071494
Mouskeftara T, Deda O, Papadopoulos G, Chatzigeorgiou A, Gika H. Lipidomic Analysis of Liver and Adipose Tissue in a High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Mice Model Reveals Alterations in Lipid Metabolism by Weight Loss and Aerobic Exercise. Molecules. 2024; 29(7):1494. https://doi.org/10.3390/molecules29071494
Chicago/Turabian StyleMouskeftara, Thomai, Olga Deda, Grigorios Papadopoulos, Antonios Chatzigeorgiou, and Helen Gika. 2024. "Lipidomic Analysis of Liver and Adipose Tissue in a High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Mice Model Reveals Alterations in Lipid Metabolism by Weight Loss and Aerobic Exercise" Molecules 29, no. 7: 1494. https://doi.org/10.3390/molecules29071494
APA StyleMouskeftara, T., Deda, O., Papadopoulos, G., Chatzigeorgiou, A., & Gika, H. (2024). Lipidomic Analysis of Liver and Adipose Tissue in a High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Mice Model Reveals Alterations in Lipid Metabolism by Weight Loss and Aerobic Exercise. Molecules, 29(7), 1494. https://doi.org/10.3390/molecules29071494