
Transcriptome, Plant Hormone, and Metabolome Analysis Reveals the Mechanism of Purple Pericarp Formation in ‘Zihui’ Papaya (Carica papaya L.)


Abstract
1. Introduction
2. Results
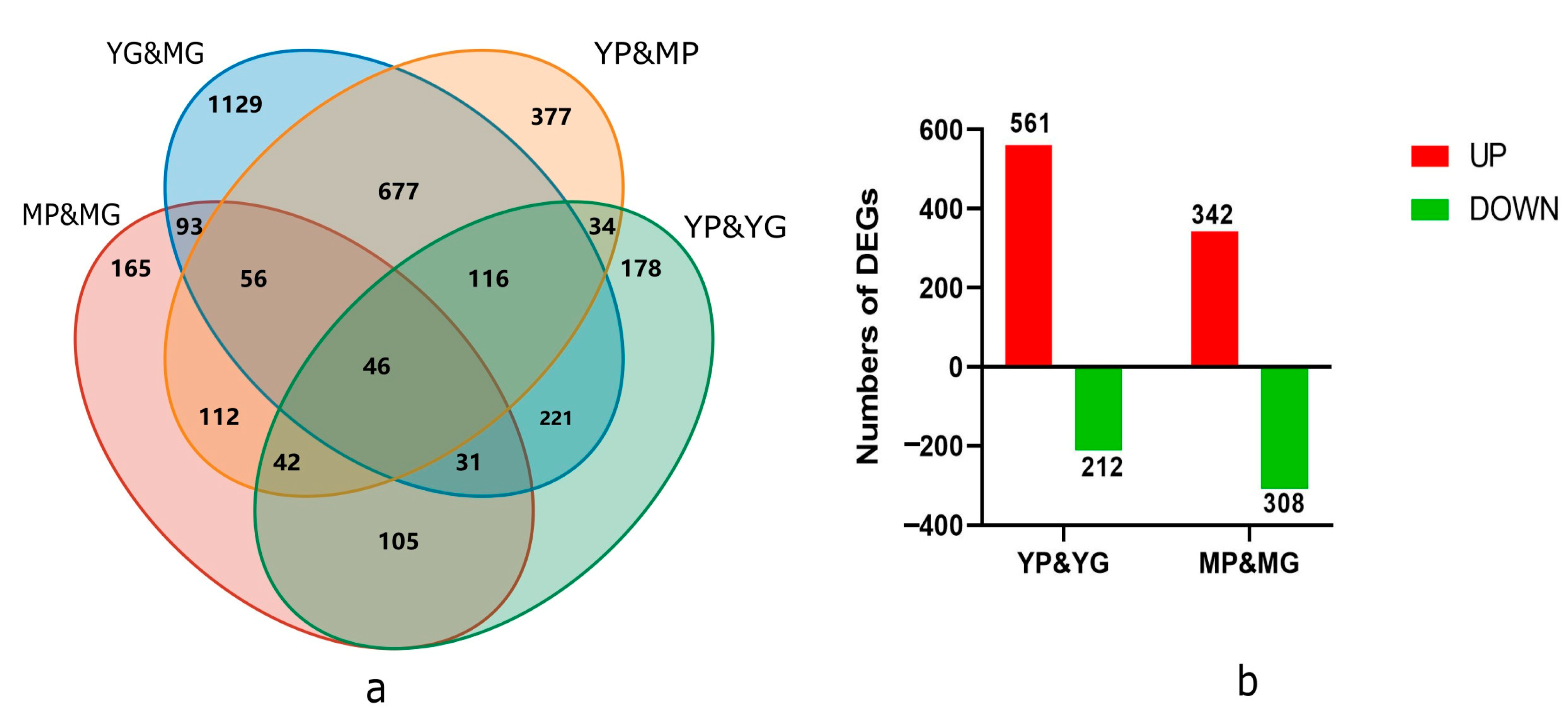
2.1. Sequencing Data Analysis of 12 RNA Libraries from Different Colored Papaya Pericarp
2.2. Different Concentrations of Plant Hormones between Purple and Green Papaya Pericarp
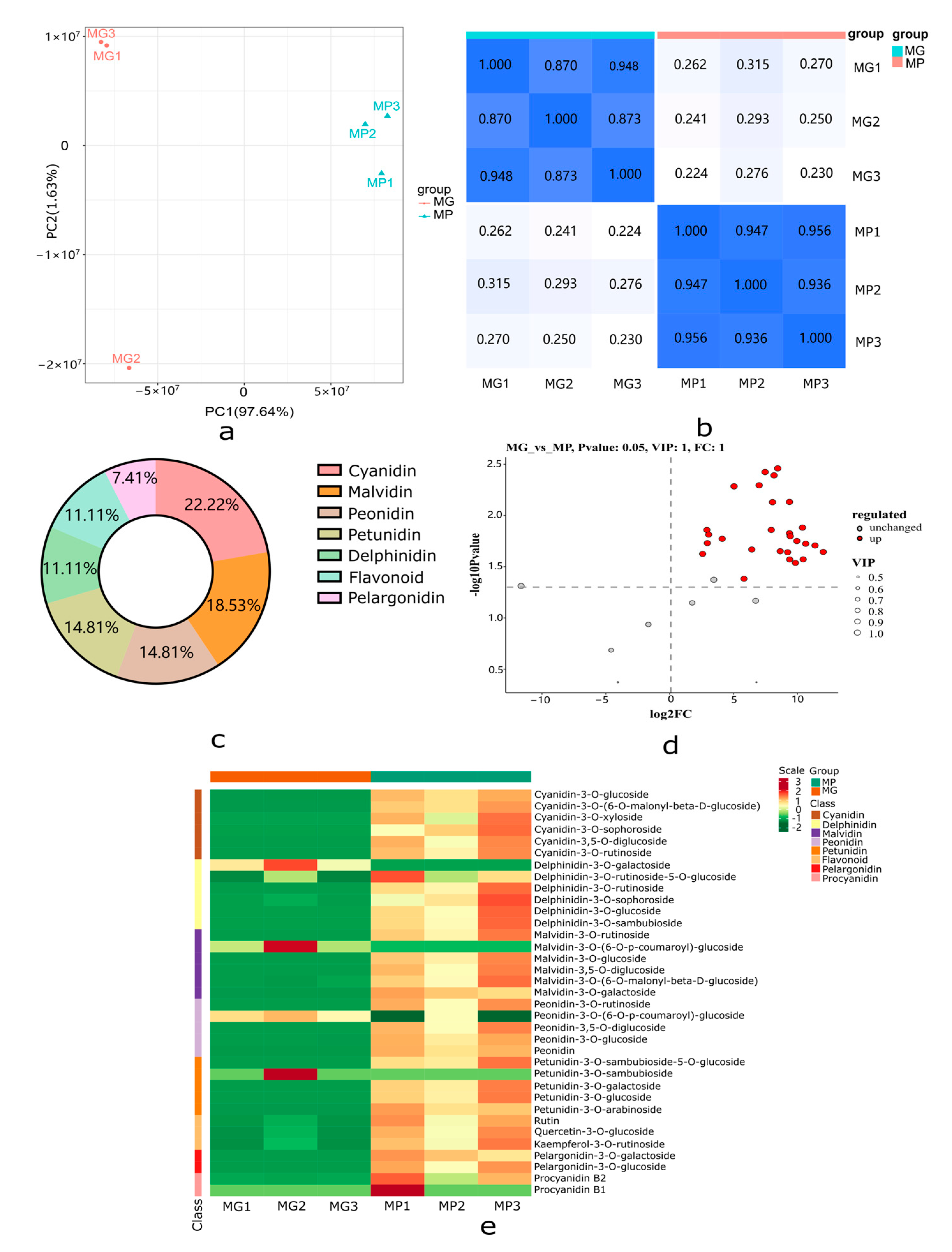
2.3. Metabolome Profiling of the Papaya Pericarp Related to Anthocyanin Metabolism
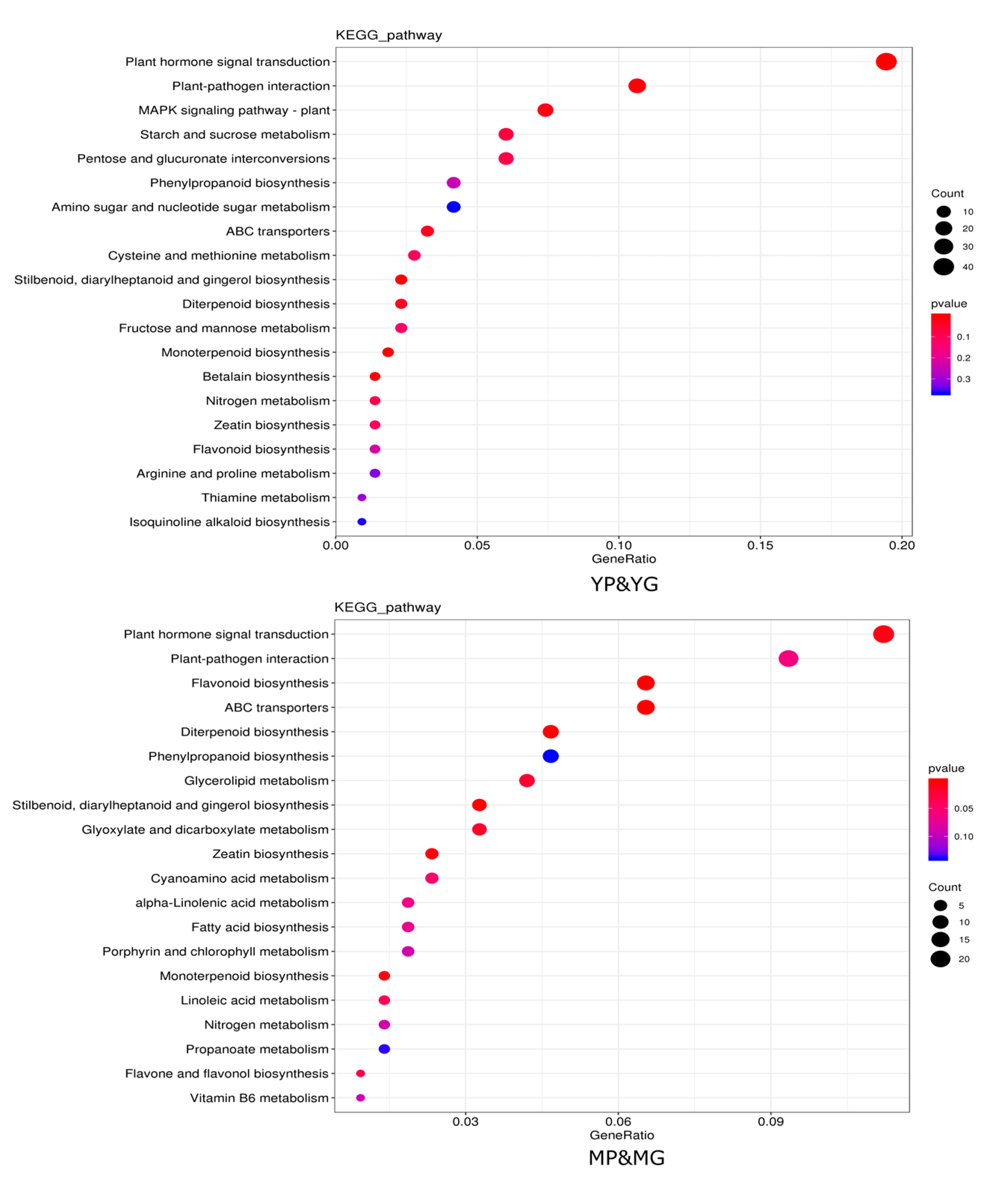
2.4. Integrated Analysis of DAMs and DEGs in Response to Different Colors of Papaya Pericarp
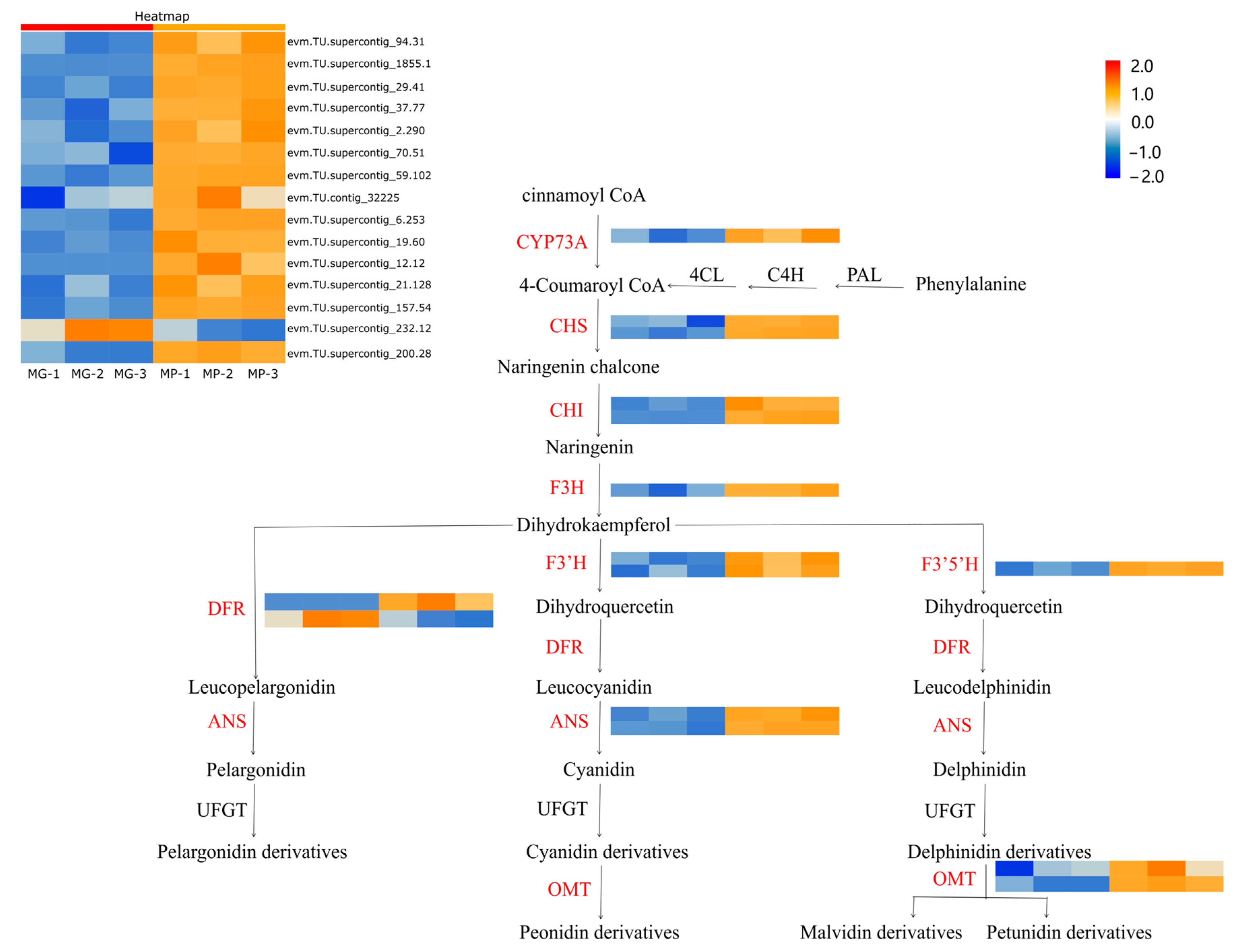
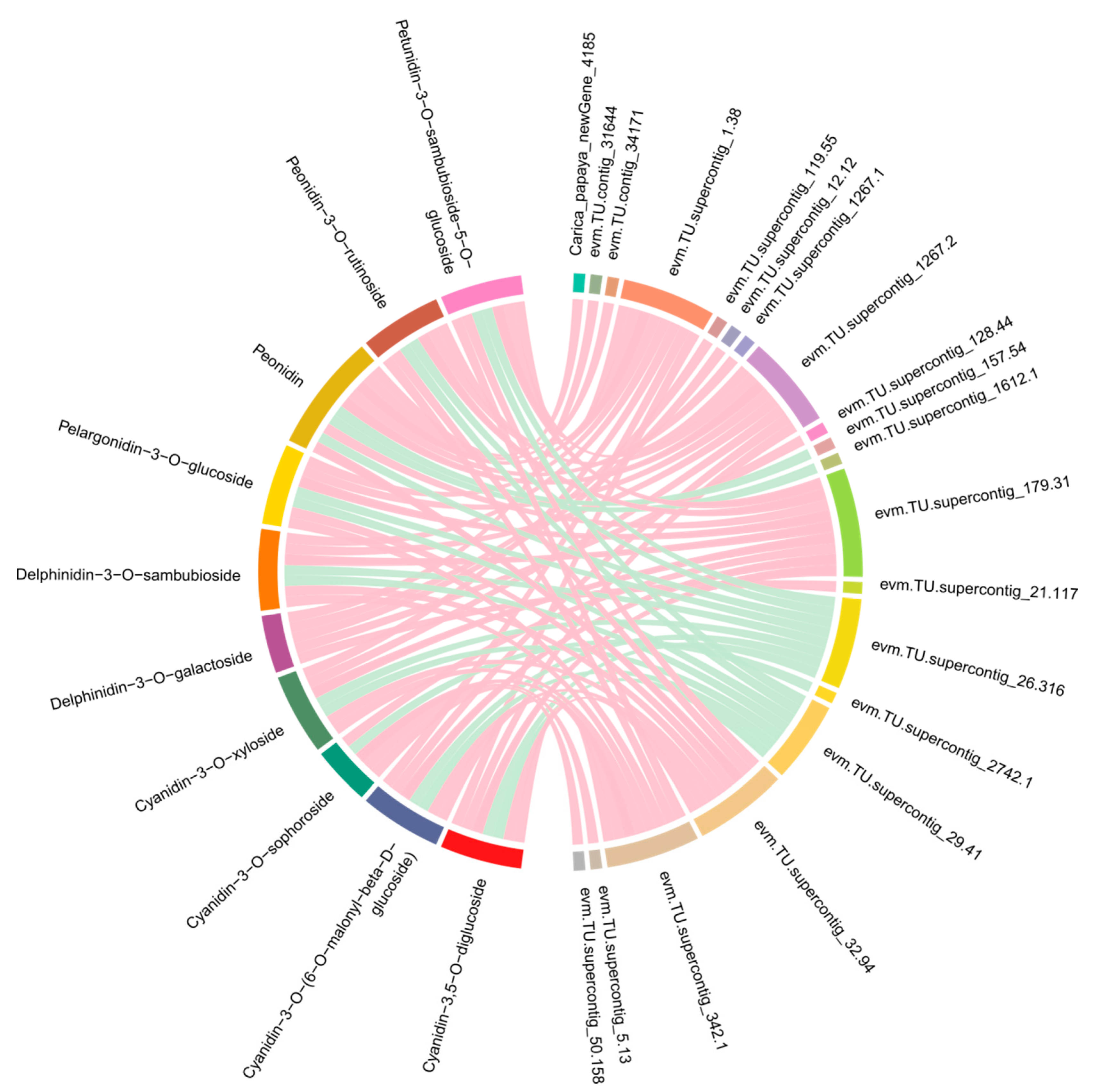
2.5. Correlation Analysis between Transcripts and Anthocyanidins
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Sequencing and Data Analyses
4.3. Differentially Expressed Gene Identification
4.4. Metabolites Extraction
4.5. UPLC-MS-MS Analysis
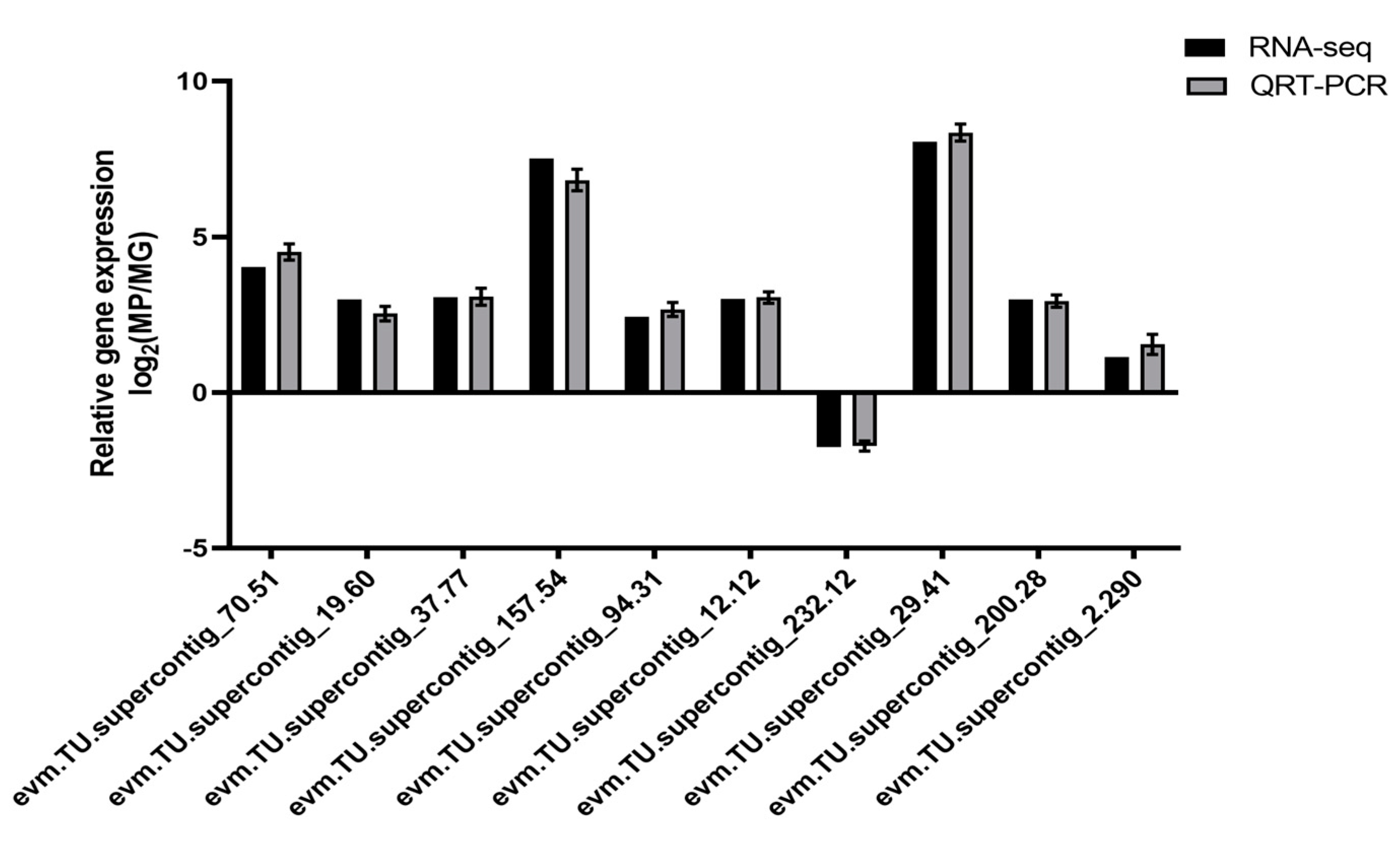
4.6. Quantitative Reverse-Transcriptase PCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vieria, W.; Nunes, A.; Veloso, J.; Machado, A.; Balbino, V.; Silva, A.; Gomes, A.; Doyle, V.; Camara, M. Colletotrichum truncatum causing anthracnose on papaya fruit (Carica papaya) in Brazil. Australas. Plant Dis. 2020, 15, 2. [Google Scholar] [CrossRef]
- Dar, J.; Wani, A.; Ahmed, M.; Nazir, R.; Zargar, S.M.; Javaid, K. Peel colour in apple (Malus × domestica Borkh.): An economic quality parameter in fruit market. Sci. Hortic. 2019, 244, 50–60. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef]
- Davies, K.M. Plant pigments and their manipulation. Annu. Rev. Plant Biol. 2004, 60, 92. [Google Scholar]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, M.; Nakatsuka, T. Genetic engineering of flavonoid pigments to modify flower color in floricultural plants. Biotechnol. Lett. 2011, 33, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.J.; Wang, T.T.; Zhang, M.; Zhang, Y.B.; Irfan, M.; Chen, L.J.; Zhang, L. Role of core structural genes for flavonoid biosynthesis and transcriptional factors in flower color of plants. Biotechnol. Biotec. Eq. 2021, 35, 1214–1229. [Google Scholar] [CrossRef]
- Timoneda, A.; Feng, T.; Sheehan, H. The evolution of betalain biosynthesis in Caryophyllales. New Phytol. 2019, 224, 71–85. [Google Scholar] [CrossRef]
- Miodzinska, E. Survey of plant pigments: Molecular and environmental determinants of plant colors. Acta Biol. Cracoviensia Ser. Bot. 2009, 51, 7–16. [Google Scholar]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Chemistry, pharmacology and health benefits of anthocyanins. Phytother. Res. 2016, 30, 1265–1286. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotech. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, S.; Spelt, C.; Hosokawa, S.; Nakamura, N.; Brugliera, F.; Demelis, L.; Geerke, D.; Schubert, A.; Tanaka, Y.; Quattrocchio, F.; et al. Genetic control and evolution of anthocyanin methylation. Plant Physiol. 2014, 165, 962–977. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Nakayama, T. Achievements and perspectives in biochemistry concerning anthocyanin modification for blue flower coloration. Plant Cell Physiol. 2015, 56, 28–40. [Google Scholar] [CrossRef]
- Meng, X.; Wang, X. Regulation of flower development and anthocyanin accumulation in Gerbera hybrida. J. Hortic. Sci. Biotech. 2004, 79, 131–137. [Google Scholar] [CrossRef]
- Faraco, M.; Spelt, C.; Bliek, M.; Verweij, W.; Hoshino, A.; Espen, L.; Prinsi, B.; Jaarsma, R.; Tarhan, E.; Boer, A.; et al. Hyperacidification of vacuoles by the combined action of two different P-ATPases in the tonoplast determines flower color. Cell Rep. 2014, 6, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.B.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Zhang, X.B.; Abrahan, C.; Colquhoun, T.A.; Liu, C.J. A proteolytic regulator controlling chalcone synthase stability and flavonoid biosynthesis in Arabidopsis. Plant Cell. 2017, 29, 1157–1174. [Google Scholar] [CrossRef]
- Song, B.; Xu, H.; Chen, L.Z.; Fan, X.X.; Jing, Z.G.; Chen, S.; Xu, Z.G. Study of the relationship between leaf color formation and anthocyanin metabolism among different purple pakchoi lines. Molecules 2020, 25, 4809. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, Y.T.; Wang, H.; Tian, Z.D.; Zhu, P.F. The dihydroflavonol 4-reductase BoDFR1 drives anthocyanin accumulation in pink-leaved ornamental kale. Theor. Appl. Genet. 2021, 134, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.P.; Pezhmanmehr, M.; Speirs, J.; McDavid, D.A.J.; Hooper, L.C.; Rinaldo, A.R.; Bogs, J.; Ebadi, A.; Walker, A.R. Grape and wine flavonoid composition in transgenic grapevines with altered expression of flavonoid hydroxylase genes. Aust. J. Grape Wine Res. 2019, 25, 293–306. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The flavonoid biosynthesis network in plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Al Sane, K.O.; Hesham, E.L. Biochemical and genetic evidences of anthocyanin biosynthesis and accumulation in a selected tomato mutant. Rend. Lincei 2015, 26, 293–306. [Google Scholar] [CrossRef]
- Kim, E.; Kim, C.; Kim, S. Identification of two novel mutant ANS alleles responsible for inactivation of anthocyanidin synthase and failure of anthocyanin production in onion (Allium cepa L.). Euphytica 2016, 212, 427–437. [Google Scholar] [CrossRef]
- Feng, X.; Gao, G.; Yu, C.; Zhu, A.; Chen, J.; Chen, K.; Wang, X.; Abubakar, A.S.; Chen, P. Transcriptome and metabolome analysis reveals anthocyanin biosynthesis pathway associated with ramie (Boehmeria nivea (L.) Gaud.) leaf color formation. BMC Genomics. 2021, 22, 684. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.Y.; Ma, X.; Gao, X.; Wu, W.L.; Zhou, B. Light induced regulation pathway of anthocyanin biosynthesis in plants. Int. J. Mol. Sci. 2021, 22, 11116. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lv, J.; Liu, Z.; Wang, J.; Yang, B.; Chen, W.; Ou, L.; Dai, X.; Zhang, Z.; Zou, X. Integrative analysis of metabolome and transcriptome reveals the mechanism of color formation in pepper fruit (Capsicum annuum L.). Food Chem. 2020, 306, 125629. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, H.; Zhou, C.; Kuang, R.; Huang, B.; Yang, M. Breeding of a new papaya cultivar Zihui with good quality and high yield. J. Fruit Sci. 2022, 39, 1129–1132. [Google Scholar]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Su, W.; Cai, Z.; Dong, L.; Li, C.; Xin, M.; Fang, W.; Liu, Y.; Wang, X.; Huang, Z.; et al. Combined analysis of transcriptome and metabolome reveals the potential mechanism of coloration and fruit quality in yellow and purple Passiflora edulis Sims. J. Agric. Food Chem. 2020, 68, 12096–12106. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; et al. Development of “purple endosperm rice” by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Lao, F.; Giusti, M.M. Quantification of purple corn (Zea mays L.) anthocyanins using spectrophotometric and HPLC approaches: Method comparison and correlation. Food Anal. Method 2016, 9, 1367–1380. [Google Scholar] [CrossRef]
- Bellido, G.G.; Beta, T. Anthocyanin composition and oxygen radical scavenging capacity (ORAC) of milled and pearled purple, black, and common barley. J. Agric. Food Chem. 2009, 57, 1022–1028. [Google Scholar] [CrossRef]
- Li, Z.; Ahammed, G.J. Hormonal regulation of anthocyanin biosynthesis for improved stress tolerance in plants. Plant Physiol. Bioch. 2023, 201, 107835. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Rothstein, S. Auxin-responsive SAUR39 gene modulates auxin level in rice. Plant Signal. Behav. 2009, 4, 1174–1175. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Thwe, A.A.; Kim, S.J.; Park, J.S.; Arasu, M.; Al-Dhabi, N.A.; Park, N.; Park, S.U. Effect of auxins on anthocyanin accumulation in hairy root cultures of Tartary buckwheat cultivar Hokkai T10. Nat. Prod. Commun. 2016, 11, 1283–1286. [Google Scholar] [CrossRef] [PubMed]
- Craker, L.E.; Standley, L.A.; Starbuck, M.J. Ethylene control of anthocyanin synthesis in Sorghum. Plant Physiol. 1971, 48, 349–352. [Google Scholar] [CrossRef]
- Banerjee, A.; Singh, A.; Roychoudhury, A. Spermidine application reduces fluoride uptake and ameliorates physiological injuries in a susceptible rice cultivar by activating diverse regulators of the defense machinery. Environ. Sci. Pollut. Res. 2019, 26, 36598–36614. [Google Scholar] [CrossRef]
- Jin, S.W.; Rahim, M.A.; Jung, H.J.; Afrin, K.S.; Kim, H.T.; Park, J.I.; Kang, J.G.; Nou, I.S. Abscisic acid and ethylene biosynthesis-related genes are associated with anthocyanin accumulation in purple ornamental cabbage (Brassica oleracea var. Acephala). Genome 2019, 62, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Liu, Y.; Wang, A.; Liu, Y.; Li, X.; Liu, Z.; Li, X.; Yang, Y.; Wang, J. The response of tartary buckwheat and 19 bZIP genes to abscisic acid (ABA). Mol. Biol. Rep. 2021, 48, 4341–4350. [Google Scholar] [CrossRef] [PubMed]
- Piazza, P.; Procissi, A.; Jenkins, G.I.; Tonelli, C. Members of the c1/pl1 regulatory gene family mediate the response of maize aleurone and mesocotyl to different light qualities and cytokinins. Plant Physiol. 2002, 128, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, A.; Paul, L.K.; Sharma, E.; Jha, S.; Jain, M.; Khurana, J.P. OsRR6, a type-A response regulator in rice, mediates cytokinin, light and stress responses when over-expressed in Arabidopsis. Plant Physiol. Bioch. 2021, 161, 98–112. [Google Scholar] [CrossRef]
- Jeon, E.H.; Pak, J.H.; Kim, M.J.; Kim, H.J.; Shin, S.H.; Lee, J.H.; Kin, D.H.; Oh, J.; On, B.; Jung, H.W.; et al. Ectopic expression of ubiquitin-conjugating enzyme gene from wild rice, ogubc1, confers resistance against uv-b radiation and botrytis infection in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2012, 427, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Yan, F.; Chen, G.; Hu, Z.; Wei, S.; Lai, J. Accumulation of anthocyanin and its associated gene expression in purple tumorous stem mustard (Brassica juncea var. tumida Tsen et Lee) sprouts when exposed to light, dark, sugar and methyl jasmonate. J. Agric. Food Chem. 2019, 67, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Xiang, F.; Yang, P.; Li, X.; Zhong, M.; He, R.; Li, X.; Peng, W.; Liu, X.; Zhao, X. Overexpression of BnGA2ox2, a rapeseed gibberellin 2-oxidase, causes dwarfism and increased chlorophyll and anthocyanin accumulation in Arabidopsis and rapeseed. Plant Growth Regul. 2021, 93, 65–77. [Google Scholar] [CrossRef]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.; Sederoff, R.; Zhao, X. MYB-mediated regulation of anthocyanin biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Chu, G.; Huang, C.; Tian, S.; Zhao, Z.; Chen, G. Anthocyanin accumulation and molecular analysis of anthocyanin biosynthesis-associated genes in eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 2906–2912. [Google Scholar] [CrossRef]
- Xiao, X.; Wenqiu, L.; Wei, L.; Gao, X.; Lv, L.; Ma, F.; Liu, Y. The analysis of physiological variations in M2 generation of Solanum melongena L. mutagenized by ethyl methane sulfonate. Front. Plant Sci. 2017, 8, 17. [Google Scholar]
- Wang, Y.; Luo, Z.; Lu, C.; Zhou, R.; Zhang, H.; Zhao, L.; Yu, W.; Zhao, T. Transcriptome profiles reveal new regulatory factors of anthocyanin accumulation in a novel purple-colored cherry tomato cultivar Jinling Moyu. Plant Growth Regul. 2019, 87, 9–18. [Google Scholar] [CrossRef]
- Tang, B.; Li, L.; Hu, Z.; Chen, Y.; Tan, T.; Jia, Y.; Xie, Q.; Chen, G. Anthocyanin accumulation and transcriptional regulation of anthocyanin biosynthesis in purple pepper. J. Agric. Food Chem. 2020, 68, 12152–12163. [Google Scholar] [CrossRef]
- Liu, Y.; Lin-Wang, K.; Deng, C.; Warran, B.; Wang, L.; Yu, B.; Yang, H.; Wang, J.; Espley, R.V.; Zhang, J.; et al. Comparative transcriptome analysis of white and purple potato to identify genes involved in anthocyanin biosynthesis. PLoS ONE 2015, 10, e0129148. [Google Scholar] [CrossRef]
- Liu, F.; Yang, Y.; Gao, J.; Ma, C.; Bi, Y. A comparative transcriptome analysis of a wild purple potato and its red mutant provides insight into the mechanism of anthocyanin transformation. PLoS ONE 2018, 13, e0191406. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Singh, R.K.; Navarre, D.A. Transcription factors, sucrose, and sucrose metabolic genes interact to regulate potato phenylpropanoid metabolism. J. Exp. Bot. 2013, 64, 5115–5131. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Barragán, A.; Ochoa-Alejo, N. Virus-induced silencing of MYB and WD40 transcription factor genes affects the accumulation of anthocyanins in chilli pepper fruit. Biol. Plant. 2014, 58, 567–574. [Google Scholar] [CrossRef]
- Yuan, Y.; Chiu, L.W.; Li, L. Transcriptional regulation of anthocyanin biosynthesis in red cabbage. Planta 2009, 230, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.W.; Zhou, X.; Burke, S.; Wu, X.; Prior, R.L.; Li, L. The purple cauliflower arises from activation of a MYB transcription factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef]
- Hsu, C.C.; Chen, Y.Y.; Tsai, W.C.; Chen, W.H.; Chen, H.H. Three R2R3-MYB transcription factors regulate distinct floral pigmentation patterning in Phalaenopsis spp. Plant Physiol. 2015, 168, 175–191. [Google Scholar] [CrossRef]
- Xu, H.F.; Yang, G.X.; Zhang, J.; Wang, Y.C.; Zhang, T.L.; Wang, N.; Jiang, S.H.; Zhang, Z.Y.; Chen, X.S. Overexpression of a repressor MdMYB15L negatively regulates anthocyanin and cold tolerance in red-fleshed calus, Biochem. Biophys. Res. Commun. 2018, 500, 405–410. [Google Scholar] [CrossRef]
- Sakai, M.; Yamagishi, M.; Matsuyama, K. Repression of anthocyanin biosynthesis by R3-MYB transcription factors in lily (Lilium spp.). Plant Cell Rep. 2019, 38, 609–622. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Z.; Li, L.X.; Wang, H.B.; Zhou, H.; Chen, X.S.; Feng, S.Q. Apple (Malus pumila) MdMYB306-like inhibits anthocyanin synthesis by directly interacting with MdMYB17 and MdbHLH33. Plant J. 2022, 110, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; He, Y.; Li, J.; Liu, Y.; Chen, H. CBFs function in anthocyanin biosynthesis by interacting with MYB113 in eggplant (Solanum melongena L.). Plant Cell Physiol. 2020, 61, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; He, K.; Zhang, Z.; Hu, Y. Molecular mechanism of phosphorous signaling inducing anthocyanin accumulation in Arabidopsis. Plant Physiol. Bioch. 2023, 196, 121–129. [Google Scholar] [CrossRef]
- Zhu, L.; Liao, Y.; Lin, K.; Wu, W.; Duan, L.; Wang, P.; Xiao, X.; Zhang, T.; Chen, X.; Wang, J.; et al. Cytokinin promotes anthocyanin biosynthesis via regulating sugar accumulation and MYB113 expression in Eucalyptus. Tree Physiol. 2023, 44, tpad154. [Google Scholar] [CrossRef] [PubMed]
- An, G.; Chen, J. Frequent gain-and loss-of-function mutations of the BjMYB113 gene accounted for leaf color variation in Brassica juncea. BMC Plant Biol. 2021, 21, 301. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.Q.; Cheng, S.Y.; Zhang, J.Q.; Lin, H.F.; Chen, Y.Y.; Yue, S.J.; Tian, M.; Tang, Y.P.; Zhao, Y.C. Morus alba L. leaves—Integration of their transcriptome and metabolomics dataset: Investigating potential genes involved in flavonoid biosynthesis at different harvest times. Front. Plant Sci. 2021, 12, 736332. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Hou, H.; Ma, X.; Sun, S.; Wang, H.; Kong, L. Metabolomics and gene expression analysis reveal the accumulation patterns of phenylpropanoids and flavonoids in different colored-grain wheats (Triticum aestivum L.). Food Res. Int. 2020, 138, 109711. [Google Scholar] [CrossRef]
- Wu, X.M.; Wang, B.; Xie, F.G.; Zhang, L.P.; Gong, J.; Zhu, W.; Li, X.Q.; Feng, F.Q.; Huang, J. QTL mapping and transcriptome analysis identify candidate genes regulating pericarp thickness in sweet corn. BMC Plant Biol. 2020, 20, 117. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Reads | Mapped Reads | Mapping Rate% | Unique Mapping Rate% | Multiple Mapping Reads% | GC% | Q30/% |
|---|---|---|---|---|---|---|---|
| MG-1 | 38,351,726 | 33,980,621 | 88.60 | 86.43 | 2.17 | 44.86 | 95.79 |
| MG-2 | 42,073,822 | 36,849,432 | 87.58 | 85.40 | 2.18 | 44.68 | 95.07 |
| MG-3 | 41,554,166 | 36,600,415 | 88.08 | 85.89 | 2.19 | 44.88 | 94.10 |
| MP-1 | 41,276,198 | 36,194,284 | 87.69 | 85.83 | 1.86 | 44.81 | 93.73 |
| MP-2 | 41,227,370 | 35,387,347 | 85.83 | 83.89 | 1.95 | 44.92 | 95.10 |
| MP-3 | 41,212,436 | 36,040,076 | 87.45 | 85.32 | 2.13 | 44.43 | 95.01 |
| YG-1 | 41,863,196 | 36,964,877 | 88.30 | 86.24 | 2.06 | 45.01 | 93.90 |
| YG-2 | 41,476,052 | 36,522,639 | 88.06 | 86.19 | 1.86 | 44.29 | 95.10 |
| YG-3 | 43,072,194 | 37,717,196 | 87.57 | 85.77 | 1.80 | 44.28 | 94.12 |
| YP-1 | 41,486,398 | 36,277,749 | 87.44 | 85.25 | 2.20 | 45.23 | 93.97 |
| YP-2 | 41,419,874 | 36,460,280 | 88.03 | 85.70 | 2.32 | 44.52 | 95.14 |
| YP-3 | 41,542,488 | 36,476,648 | 87.81 | 85.45 | 2.35 | 44.81 | 94.83 |
| Plant Hormone Types | Metabolite Concentration (nmol/kg) | |
|---|---|---|
| MG | MP | |
| (+)-Abscisic acid | 216.56 ± 65.88 | 2633.46 ± 194.76 * |
| 1-Aminocyclopropanecarboxylic acid | 2598.37 ± 396.12 * | 0.00 ± 0.00 |
| Salicylic acid | 135.98 ± 23.66 | 218.01 ± 109.51 |
| Indole-3-carboxaldehyde | 99.94 ± 25.51 | 97.22 ± 7.43 |
| Indole-3-acetic acid | 82.21 ± 11.92 | 94.96 ± 20.32 |
| N6-isopentenyladenosine | 23.09 ± 8.43 | 27.83 ± 7.92 |
| Dihydrojasmonic Acid | 18.03 ± 1.74 | 13.49 ± 6.93 |
| N-((-)-jasmonoyl)-S-isoleucine | 14.97 ± 2.49 * | 0.00 ± 0.00 |
| trans-Zeatin-riboside | 10.65 ± 1.79 | 16.20 ± 1.75 |
| trans-Zeatin | 1.02 ± 0.17 | 7.66 ± 2.02 * |
| N6-(delta 2-Isopentenyl)-adenine | 1.15 ± 0.31 | 0.92 ± 0.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Yang, M.; Liu, C.; Kuang, R.; He, H.; Zhou, C.; Wei, Y. Transcriptome, Plant Hormone, and Metabolome Analysis Reveals the Mechanism of Purple Pericarp Formation in ‘Zihui’ Papaya (Carica papaya L.). Molecules 2024, 29, 1485. https://doi.org/10.3390/molecules29071485
Wu X, Yang M, Liu C, Kuang R, He H, Zhou C, Wei Y. Transcriptome, Plant Hormone, and Metabolome Analysis Reveals the Mechanism of Purple Pericarp Formation in ‘Zihui’ Papaya (Carica papaya L.). Molecules. 2024; 29(7):1485. https://doi.org/10.3390/molecules29071485
Chicago/Turabian StyleWu, Xiaming, Min Yang, Chuanhe Liu, Ruibing Kuang, Han He, Chenping Zhou, and Yuerong Wei. 2024. "Transcriptome, Plant Hormone, and Metabolome Analysis Reveals the Mechanism of Purple Pericarp Formation in ‘Zihui’ Papaya (Carica papaya L.)" Molecules 29, no. 7: 1485. https://doi.org/10.3390/molecules29071485
APA StyleWu, X., Yang, M., Liu, C., Kuang, R., He, H., Zhou, C., & Wei, Y. (2024). Transcriptome, Plant Hormone, and Metabolome Analysis Reveals the Mechanism of Purple Pericarp Formation in ‘Zihui’ Papaya (Carica papaya L.). Molecules, 29(7), 1485. https://doi.org/10.3390/molecules29071485