
The Evolution and Application of a Novel DNA Aptamer Targeting Bone Morphogenetic Protein 2 for Bone Regeneration


Abstract
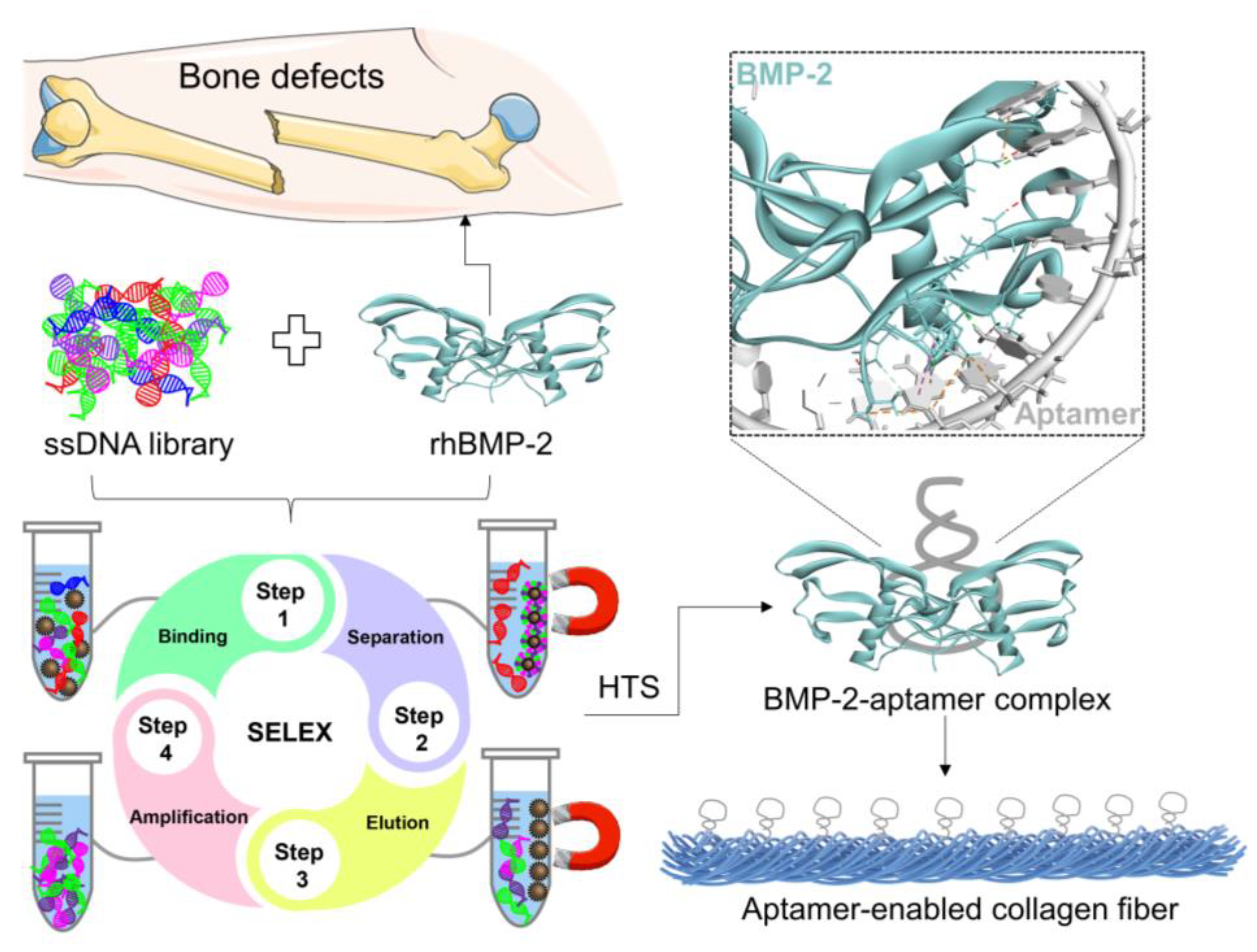
1. Introduction
2. Results and Discussions
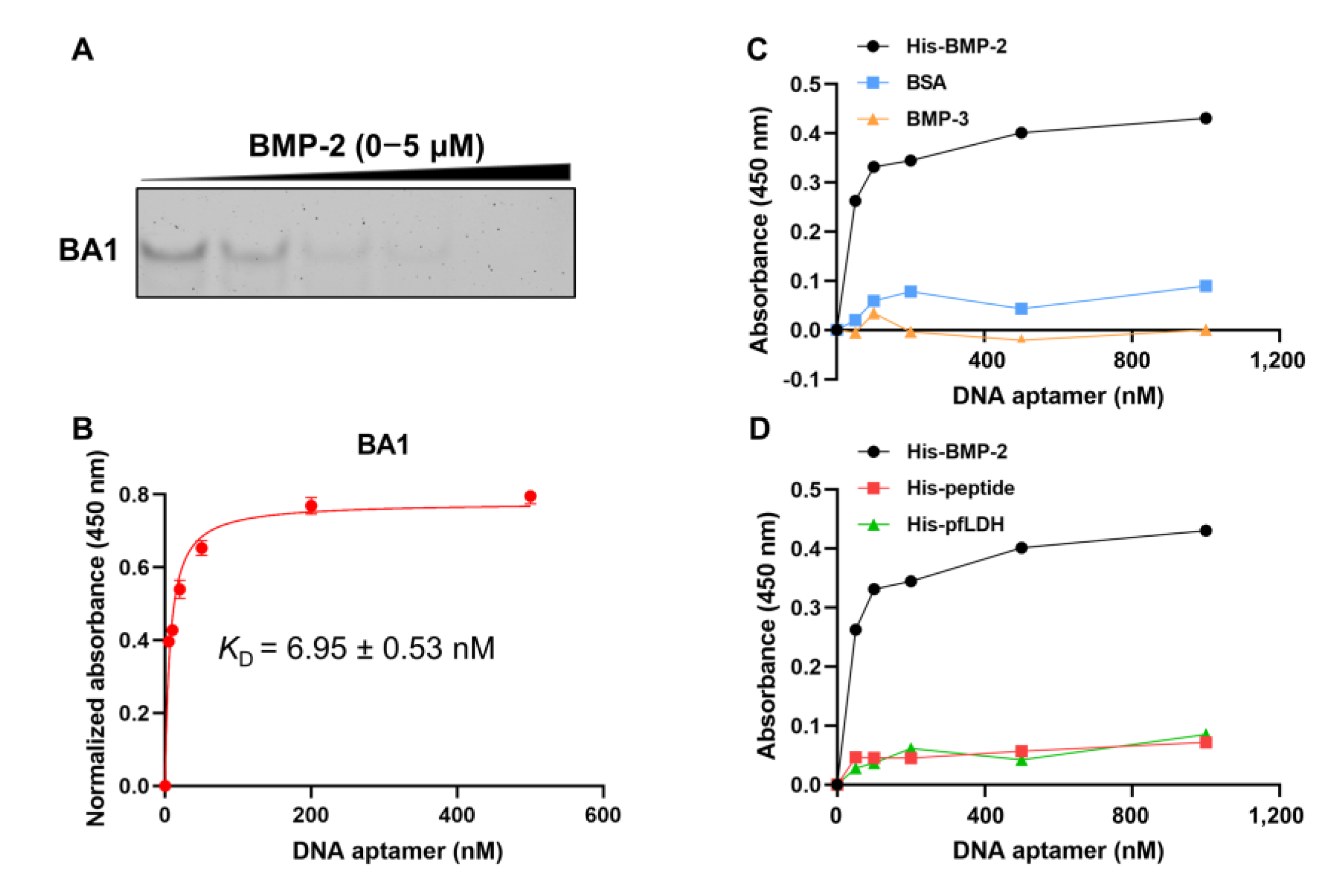
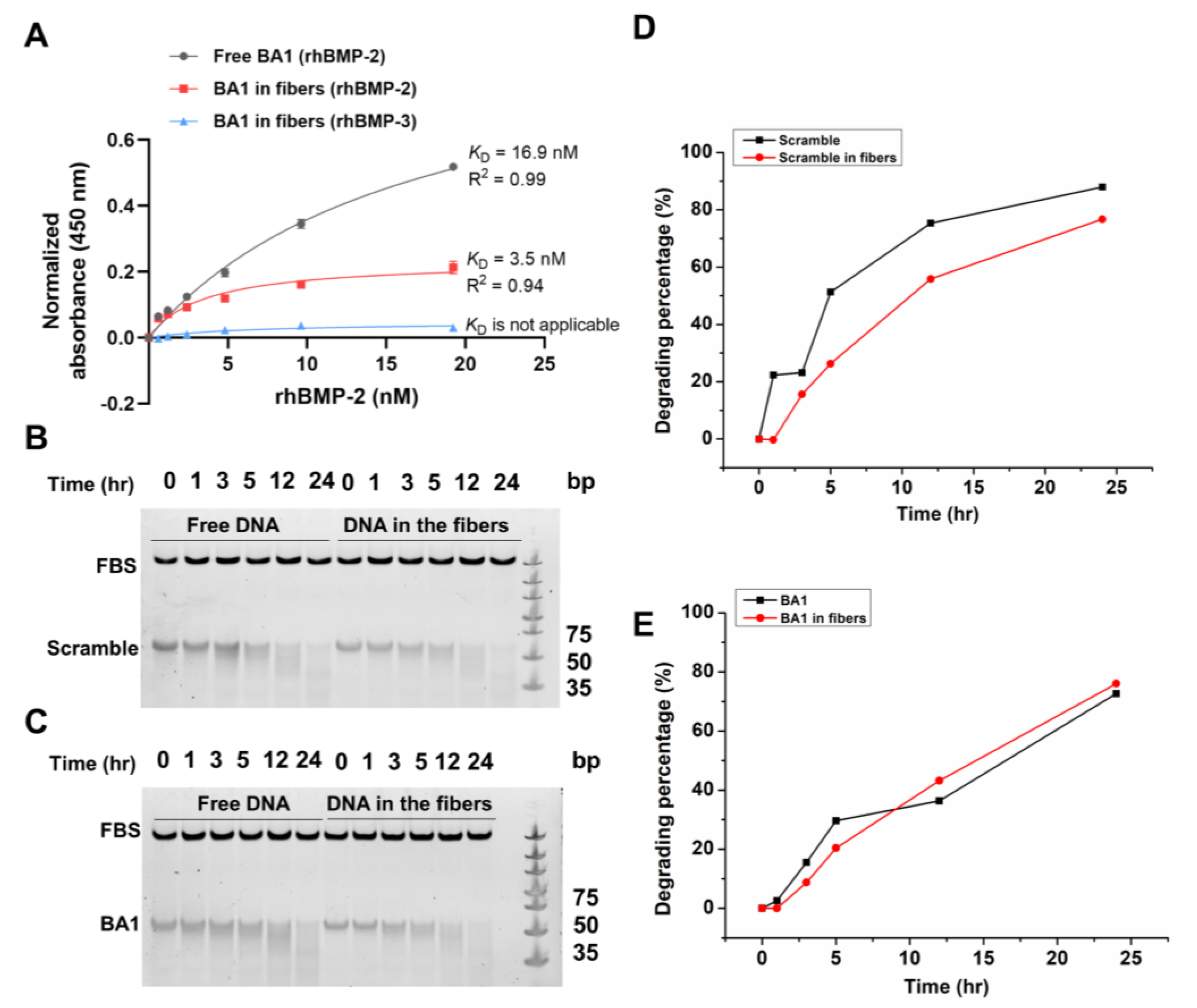
2.1. Selected DNA Aptamers Demonstrate Strong Affinity and Specificity to rhBMP-2
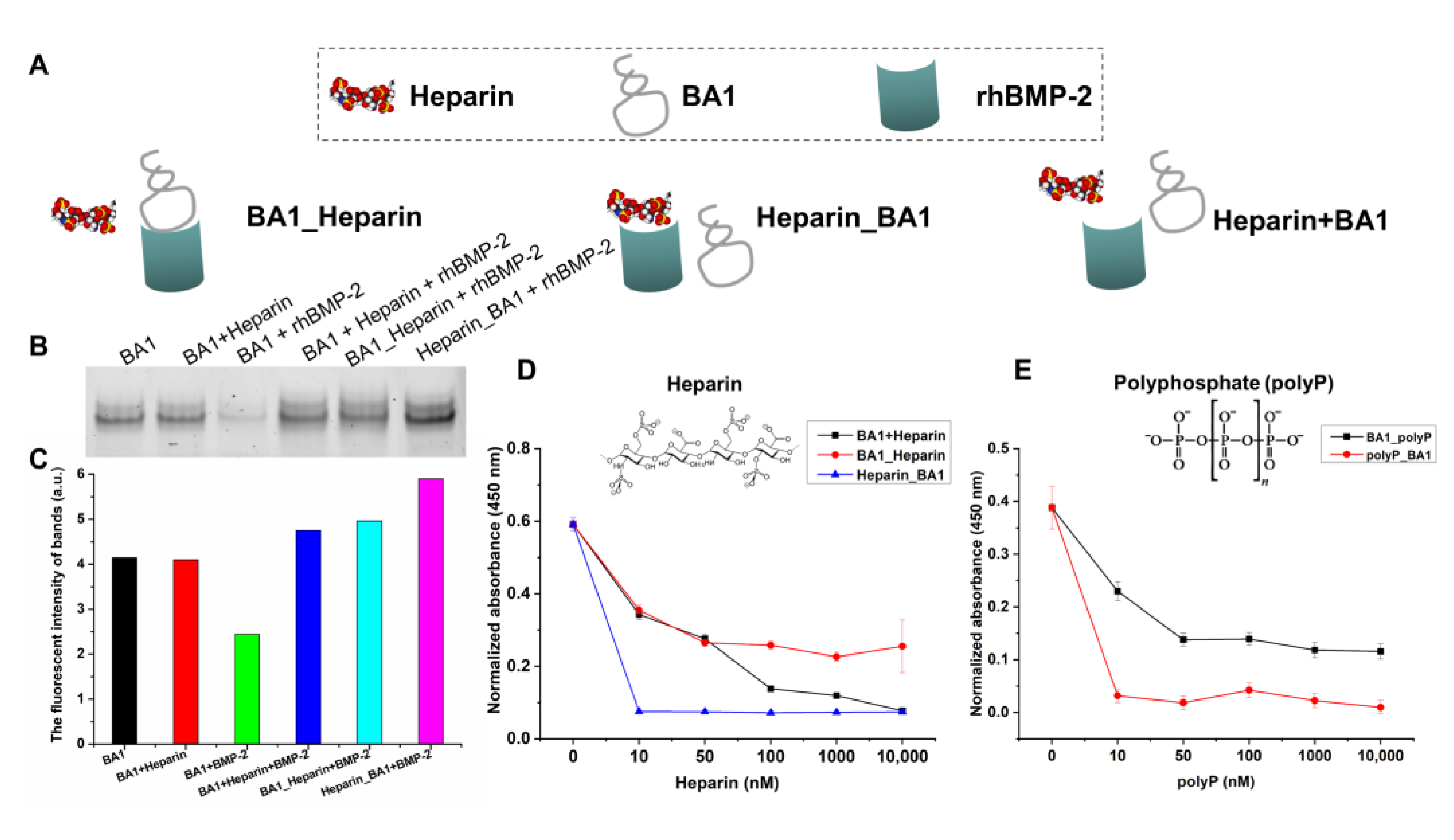
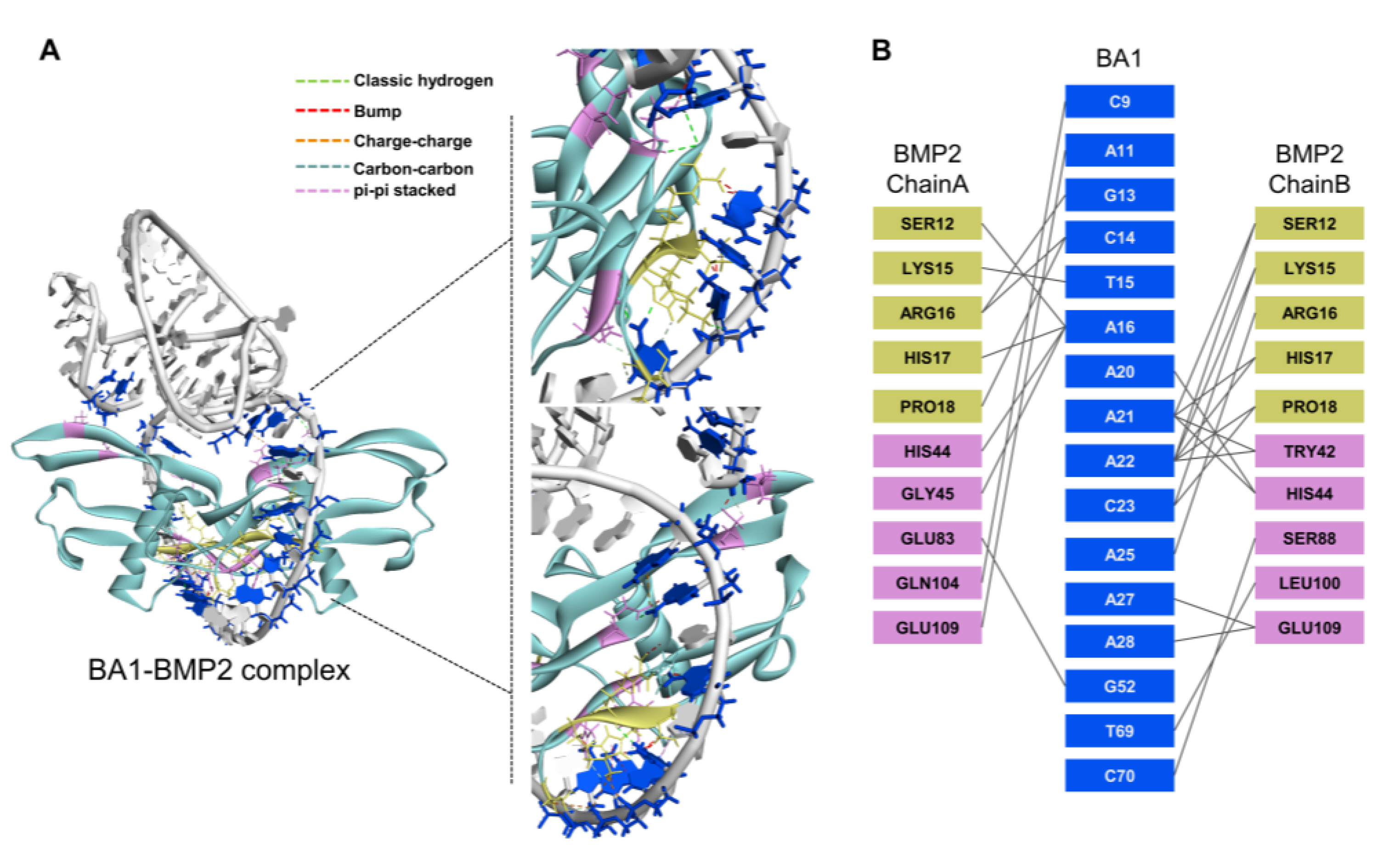
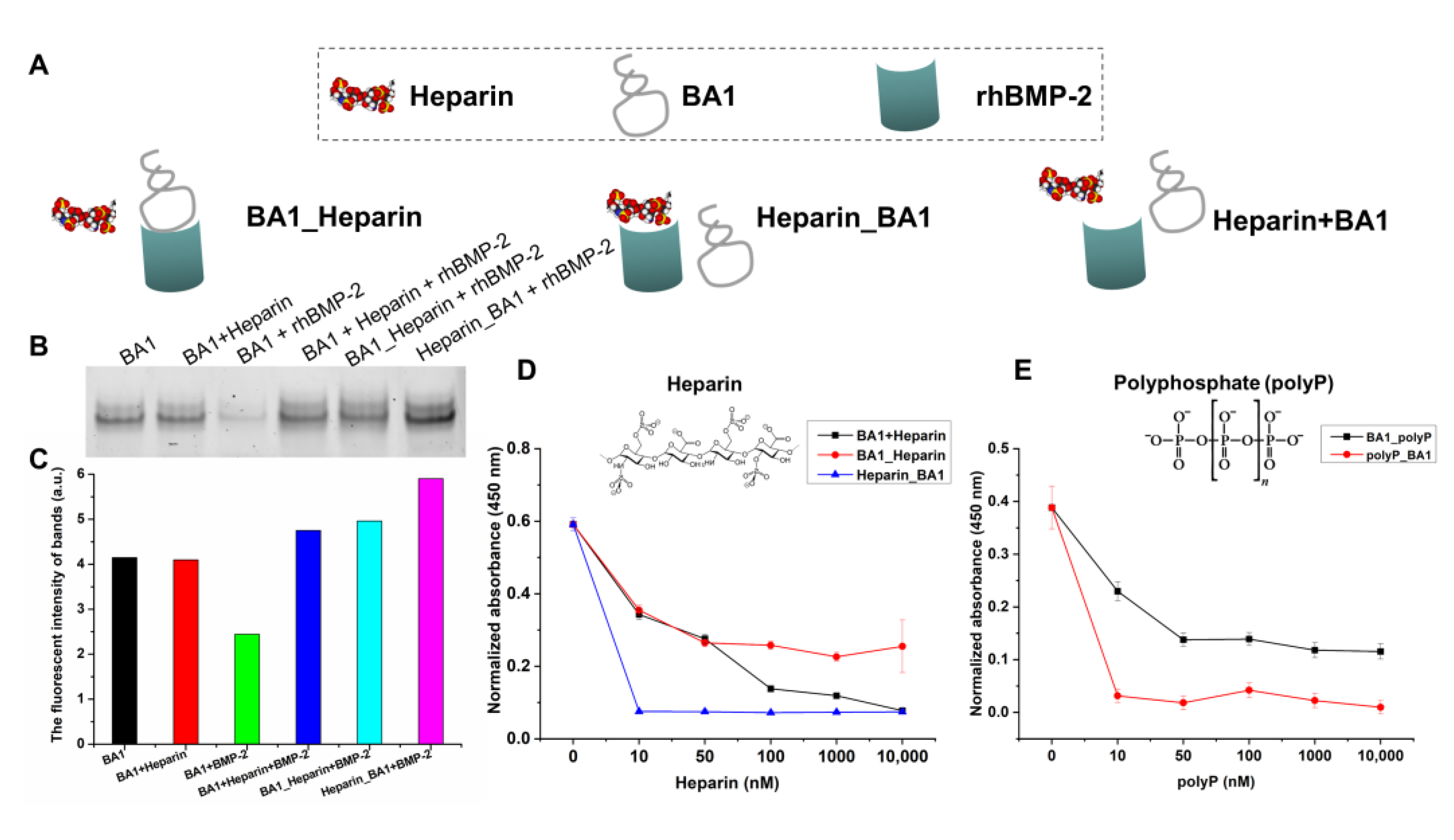
2.2. Aptamers May Interact with rhBMP-2 at Heparin-Binding Domains
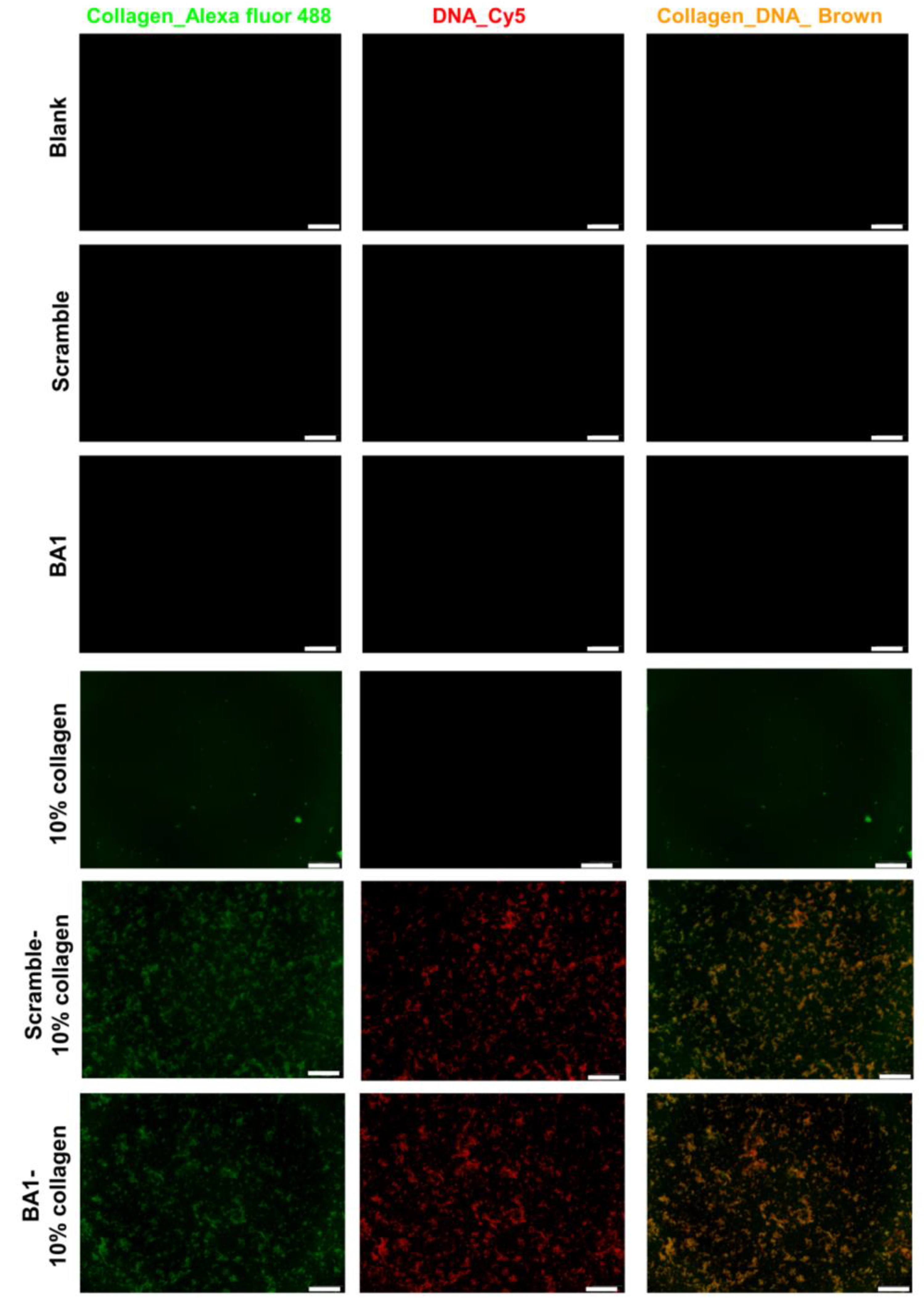
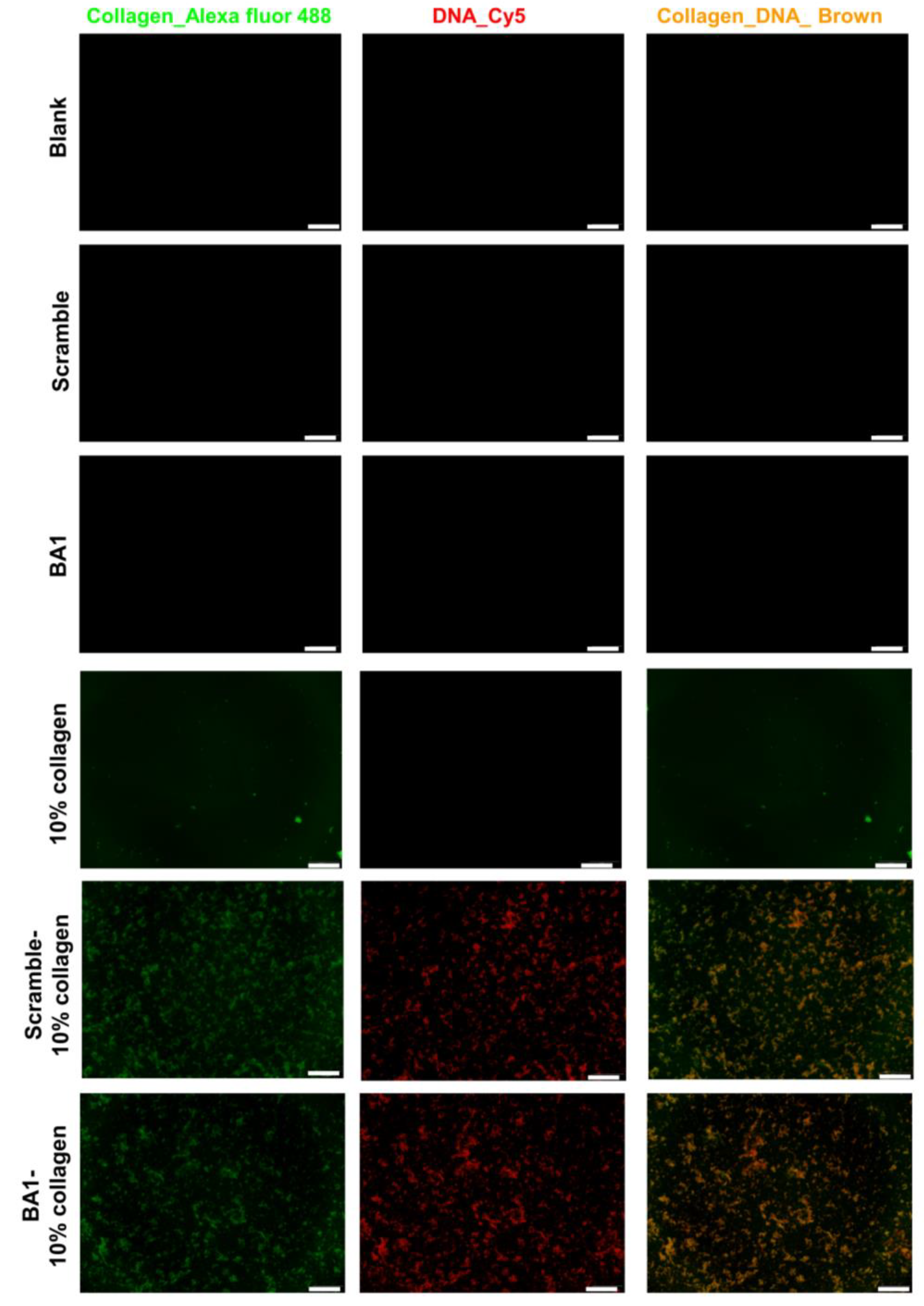
2.3. Assembly of Aptamer-Modified Collagen Fibers as rhBMP-2′s Scaffolds
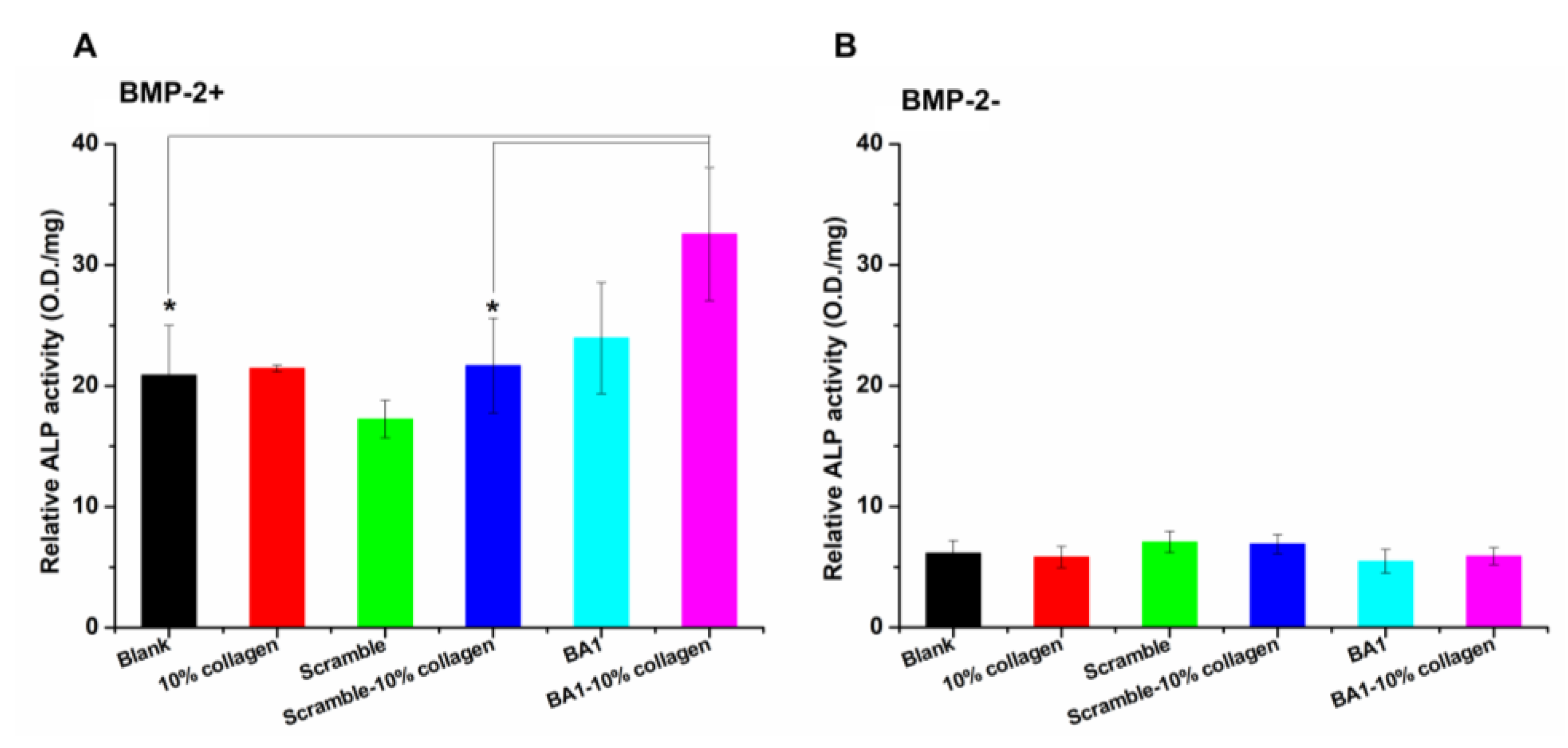
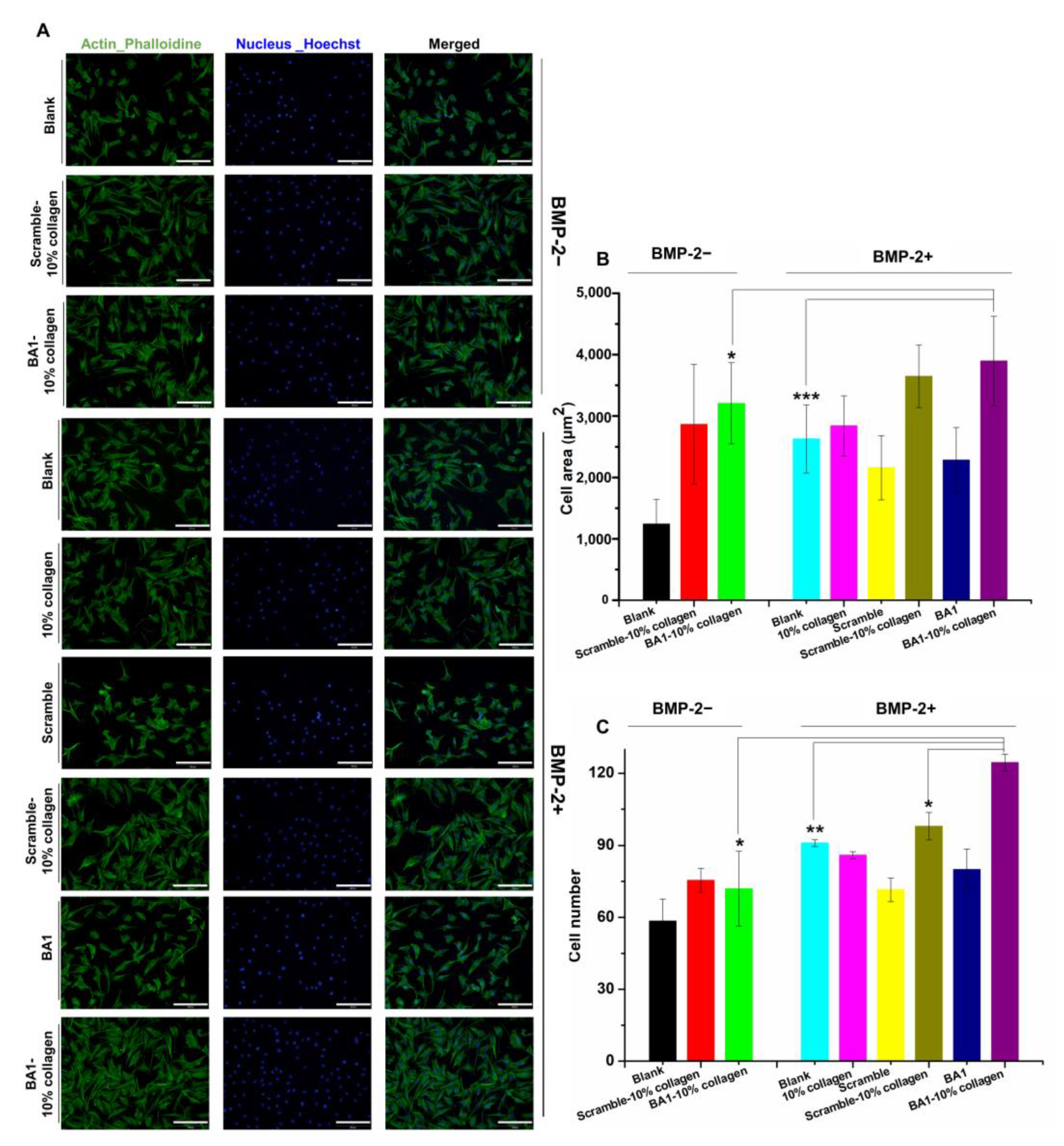
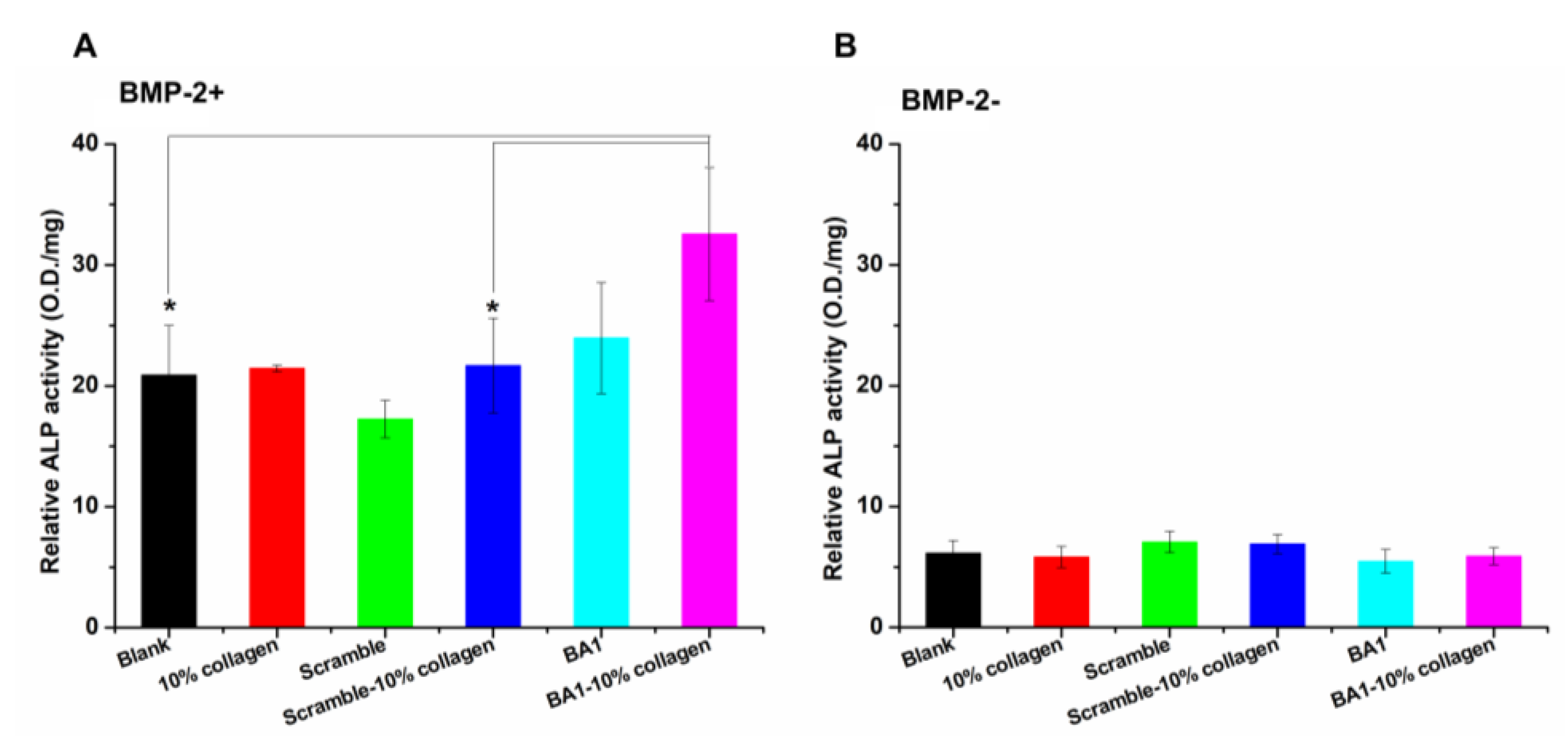
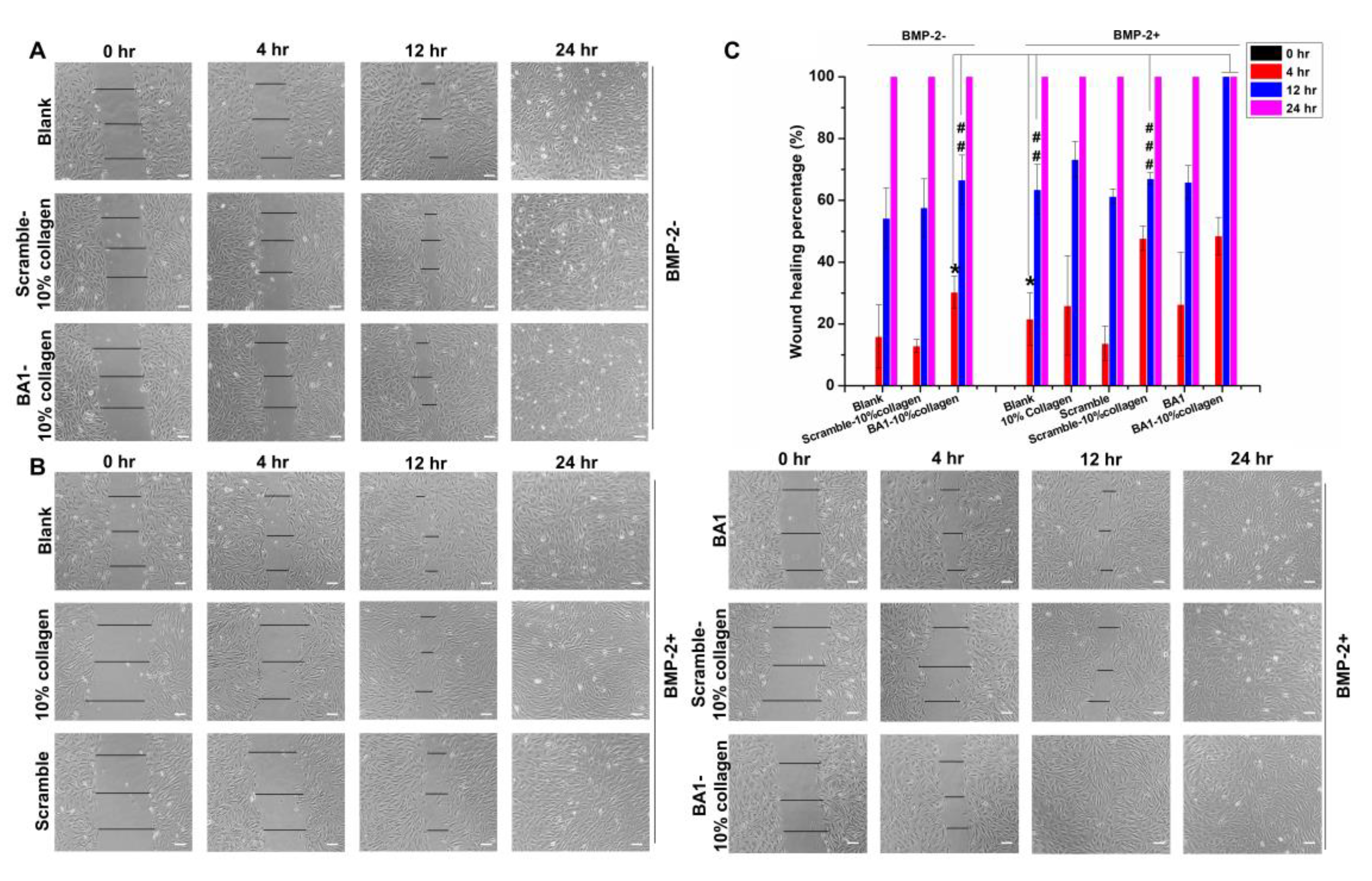
2.4. Aptamer-Enabled Scaffolds Tended to Enhance rhBMP-2-Mediated Osteogenesis
3. Materials and Methods
3.1. Materials
3.2. Expression, Renaturation, and Purification of His-Tagged rhBMP-2 Targets
3.3. Aptamer SELEX
3.4. High-Throughput Sequencing (HTS)
3.5. EMSA
3.6. ELONA
3.7. Molecular Docking
3.8. Assembly of Aptamer–Collagen Fibrous Scaffolds
3.9. Surface Immobilization of Aptamer–Collagen Fibers
3.10. Elemental Mapping of Aptamer–Collagen Scaffolds
3.11. Enzyme-Linked Immunosorbent Assay (ELISA)
3.12. Alkaline Phosphatase (ALP) Assay
3.13. Cell Adhesion Assay
3.14. Wound Healing Assay
3.15. Calcium Nodule Detection
3.16. Statistics and Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wildemann, B.; Ignatius, A.; Leung, F.; Taitsman, L.A.; Smith, R.M.; Pesántez, R.; Stoddart, M.J.; Richards, R.G.; Jupiter, J.B. Non-union bone fractures. Nat. Rev. Dis. Primers 2021, 7, 57. [Google Scholar] [CrossRef] [PubMed]
- Bosse, M.J.; MacKenzie, E.J.; Kellam, J.F.; Burgess, A.R.; Webb, L.X.; Swiontkowski, M.F.; Sanders, R.W.; Jones, A.L.; McAndrew, M.P.; Patterson, B.M. An analysis of outcomes of reconstruction or amputation after leg-threatening injuries. N. Engl. J. Med. 2002, 347, 1924–1931. [Google Scholar] [CrossRef] [PubMed]
- Sattui, S.E.; Saag, K.G. Fracture mortality: Associations with epidemiology and osteoporosis treatment. Nat. Rev. Endocrinol. 2014, 10, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, P.; Li, D.J.; Auston, D.A.; Mir, H.S.; Yoon, R.S.; Koval, K.J. Autograft, Allograft, and Bone Graft Substitutes: Clinical Evidence and Indications for Use in the Setting of Orthopaedic Trauma Surgery. J. Orthop. Trauma 2019, 33, 203–213. [Google Scholar] [CrossRef]
- Brydone, A.; Meek, D.; Maclaine, S. Bone grafting, orthopaedic biomaterials, and the clinical need for bone engineering. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2010, 224, 1329–1343. [Google Scholar] [CrossRef] [PubMed]
- Seeherman, H.J.; Berasi, S.P.; Brown, C.T.; Martinez, R.X.; Juo, Z.S.; Jelinsky, S.; Cain, M.J.; Grode, J.; Tumelty, K.E.; Bohner, M.; et al. A BMP/activin A chimera is superior to native BMPs and induces bone repair in nonhuman primates when delivered in a composite matrix. Sci. Transl. Med. 2019, 11, eaar4953. [Google Scholar] [CrossRef] [PubMed]
- Briquez, P.S.; Tsai, H.-M.; Watkins, E.A.; Hubbell, J.A. Engineered bridge protein with dual affinity for bone morphogenetic protein-2 and collagen enhances bone regeneration for spinal fusion. Sci. Adv. 2021, 7, eabh4302. [Google Scholar] [CrossRef]
- Friess, W.; Uludag, H.; Foskett, S.; Biron, R.; Sargeant, C. Characterization of absorbable collagen sponges as rhBMP-2 carriers. Int. J. Pharm. 1999, 187, 91–99. [Google Scholar] [CrossRef]
- Jones, A.L.; Bucholz, R.W.; Bosse, M.J.; Mirza, S.K.; Lyon, T.R.; Webb, L.X.; Pollak, A.N.; Golden, J.D.; Valentin-Opran, A. Recombinant human BMP-2 and allograft compared with autogenous bone graft for reconstruction of diaphyseal tibial fractures with cortical defects: A randomized, controlled trial. JBJS 2006, 88, 1431–1441. [Google Scholar] [CrossRef]
- Tannoury, C.A.; An, H.S. Complications with the use of bone morphogenetic protein 2 (BMP-2) in spine surgery. Spine J. 2014, 14, 552–559. [Google Scholar] [CrossRef]
- Bakshi, R.; Hokugo, A.; Khalil, D.; Wang, L.; Shibuya, Y.; Zhou, S.; Zhang, Z.; Rezzadeh, K.; McClendon, M.; Stupp, S.I. A Chemotactic Functional Scaffold with VEGF-Releasing Peptide Amphiphiles Facilitates Bone Regeneration by BMP-2 in a Large-Scale Rodent Cranial Defect Model. Plast. Reconstr. Surg. 2021, 147, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Toftdal, M.S.; Le Friec, A.; Dong, M.; Han, X.; Chen, M. 3D Electrospun Synthetic Extracellular Matrix for Tissue Regeneration. Small Sci. 2021, 1, 2100003. [Google Scholar] [CrossRef]
- Lee, S.S.; Hsu, E.L.; Mendoza, M.; Ghodasra, J.; Nickoli, M.S.; Ashtekar, A.; Polavarapu, M.; Babu, J.; Riaz, R.M.; Nicolas, J.D. Gel scaffolds of BMP-2-binding peptide amphiphile nanofibers for spinal arthrodesis. Adv. Healthc. Mater. 2015, 4, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Huang, B.J.; Kaltz, S.R.; Sur, S.; Newcomb, C.J.; Stock, S.R.; Shah, R.N.; Stupp, S.I. Bone regeneration with low dose BMP-2 amplified by biomimetic supramolecular nanofibers within collagen scaffolds. Biomaterials 2013, 34, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Behanna, H.A.; Donners, J.J.; Gordon, A.C.; Stupp, S.I. Coassembly of amphiphiles with opposite peptide polarities into nanofibers. J. Am. Chem. Soc. 2005, 127, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Svintradze, D.V.; Mrevlishvili, G.M.; Metreveli, N.; Jariashvili, K.; Namicheishvili, L.; Skopinska, J.; Sionkowska, A. Collagen–DNA complex. Biomacromolecules 2008, 9, 21–28. [Google Scholar] [CrossRef]
- Toh, W.S.; Loh, X.J. Advances in hydrogel delivery systems for tissue regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 45, 690–697. [Google Scholar] [CrossRef]
- Di, Y.; Wang, P.; Li, C.; Xu, S.; Tian, Q.; Wu, T.; Tian, Y.; Gao, L. Design, Bioanalytical, and Biomedical Applications of Aptamer-Based Hydrogels. Front. Med. 2020, 7, 456. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, C.; Yang, C.; Wu, Y.; Yu, F.; Chen, Y.; Du, J. Aptamer-Patterned Hydrogel Films for Spatiotemporally Programmable Capture and Release of Multiple Proteins. ACS Appl. Mater. Interfaces 2018, 10, 8546–8554. [Google Scholar] [CrossRef]
- Zhao, N.; Suzuki, A.; Zhang, X.; Shi, P.; Abune, L.; Coyne, J.; Jia, H.; Xiong, N.; Zhang, G.; Wang, Y. Dual Aptamer-Functionalized in Situ Injectable Fibrin Hydrogel for Promotion of Angiogenesis via Codelivery of Vascular Endothelial Growth Factor and Platelet-Derived Growth Factor-BB. ACS Appl. Mater. Interfaces 2019, 11, 18123–18132. [Google Scholar] [CrossRef] [PubMed]
- James, B.D.; Guerin, P.; Iverson, Z.; Allen, J.B. Mineralized DNA-collagen complex-based biomaterials for bone tissue engineering. Int. J. Biol. Macromol. 2020, 161, 1127–1139. [Google Scholar] [CrossRef]
- James, B.D.; Allen, J.B. Self-assembled VEGF-R2 targeting DNA aptamer-collagen fibers stimulate an angiogenic-like endothelial cell phenotype. Mater. Sci. Eng. C 2021, 120, 111683. [Google Scholar] [CrossRef]
- Long, S.; Truong, L.; Bennett, K.; Phillips, A.; Wong-Staal, F.; Ma, H. Expression, purification, and renaturation of bone morphogenetic protein-2 from Escherichia coli. Protein Expr. Purif. 2006, 46, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Tapp, M.; Dennis, P.; Naik, R.R.; Milam, V.T. Competition-Enhanced Ligand Selection to Screen for DNA Aptamers for Spherical Gold Nanoparticles. Langmuir 2021, 37, 9043–9052. [Google Scholar] [CrossRef] [PubMed]
- Nastasijevic, B.; Becker, N.A.; Wurster, S.E.; Maher, L.J., III. Sequence-specific binding of DNA and RNA to immobilized Nickel ions. Biochem. Biophys. Res. Commun. 2008, 366, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, A.M.; Wan, L.; Zheng, L.; Eisenstein, M.; Soh, H.T. A system for multiplexed selection of aptamers with exquisite specificity without counterselection. Proc. Natl. Acad. Sci. USA 2022, 119, e2119945119. [Google Scholar] [CrossRef]
- Isaksson, J.; Acharya, S.; Barman, J.; Cheruku, P.; Chattopadhyaya, J. Single-stranded adenine-rich DNA and RNA retain structural characteristics of their respective double-stranded conformations and show directional differences in stacking pattern. Biochemistry 2004, 43, 15996–16010. [Google Scholar] [CrossRef]
- Capobianco, A.; Caruso, T.; Peluso, A. Hole delocalization over adenine tracts in single stranded DNA oligonucleotides. Phys. Chem. Chem. Phys. 2015, 17, 4750–4756. [Google Scholar] [CrossRef]
- Biesiada, M.; Purzycka, K.J.; Szachniuk, M.; Blazewicz, J.; Adamiak, R.W. Automated RNA 3D structure prediction with RNAComposer. In RNA Structure Determination; Springer: Cham, Swizterland, 2016; pp. 199–215. [Google Scholar]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Pawar, S.S.; Rohane, S.H. Review on discovery studio: An important tool for molecular docking. Asian J. Res. Chem 2021, 14, 86–88. [Google Scholar] [CrossRef]
- Yarizadeh, K.; Behbahani, M.; Mohabatkar, H.; Noorbakhsh, A. Computational analysis and optimization of carcinoembryonic antigen aptamers and experimental evaluation. J. Biotechnol. 2019, 306, 1–8. [Google Scholar] [CrossRef]
- De Vries, S.J.; Van Dijk, M.; Bonvin, A.M. The HADDOCK web server for data-driven biomolecular docking. Nat. Protoc. 2010, 5, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Scheufler, C.; Sebald, W.; Hülsmeyer, M. Crystal structure of human bone morphogenetic protein-2 at 2.7 A resolution. J. Mol. Biol. 1999, 287, 103–115. [Google Scholar] [CrossRef]
- Ruppert, R.; Hoffmann, E.; Sebald, W. Human bone morphogenetic protein 2 contains a heparin-binding site which modifies its biological activity. Eur. J. Biochem. 1996, 237, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Kawazoe, Y.; Katoh, S.; Onodera, Y.; Kohgo, T.; Shindoh, M.; Shiba, T. Activation of the FGF signaling pathway and subsequent induction of mesenchymal stem cell differentiation by inorganic polyphosphate. Int. J. Biol. Sci. 2008, 4, 37. [Google Scholar] [CrossRef]
- Mrevlishvili, G.M.; Svintradze, D.V. DNA as a matrix of collagen fibrils. Int. J. Biol. Macromol. 2005, 36, 324–326. [Google Scholar] [CrossRef]
- James, B.D.; Saenz, S.; van Gent, A.; Allen, J.B. Oligomer length defines the self-assembly of single-stranded DNA–collagen complex fibers. ACS Biomater. Sci. Eng. 2019, 6, 213–218. [Google Scholar] [CrossRef]
- Fetz, A.E.; Fantaziu, C.A.; Smith, R.A.; Radic, M.Z.; Bowlin, G.L. Surface area to volume ratio of electrospun polydioxanone templates regulates the adsorption of soluble proteins from human serum. Bioengineering 2019, 6, 78. [Google Scholar] [CrossRef]
- Nedachi, T.; Kanzaki, M. Regulation of glucose transporters by insulin and extracellular glucose in C2C12 myotubes. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E817–E828. [Google Scholar] [CrossRef]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. Part B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Miyauchi, M.; Takata, T. Initial attachment of osteoblasts to various guided bone regeneration membranes: An in vitro study. J. Periodontal. Res. 2002, 37, 340–344. [Google Scholar] [CrossRef]
- Daley, W.P.; Peters, S.B.; Larsen, M. Extracellular matrix dynamics in development and regenerative medicine. J. Cell Sci. 2008, 121, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Büth, H.; Luigi Buttigieg, P.; Ostafe, R.; Rehders, M.; Dannenmann, S.R.; Schaschke, N.; Stark, H.J.; Boukamp, P.; Brix, K. Cathepsin B is essential for regeneration of scratch-wounded normal human epidermal keratinocytes. Eur. J. Cell Biol. 2007, 86, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Budiraharjo, R.; Neoh, K.G.; Kang, E.-T. Enhancing bioactivity of chitosan film for osteogenesis and wound healing by covalent immobilization of BMP-2 or FGF-2. J. Biomater. Sci. Polym. Ed. 2013, 24, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Seltana, A.; Basora, N.; Beaulieu, J.F. Intestinal epithelial wound healing assay in an epithelial-mesenchymal co-culture system. Wound Repair Regen. 2010, 18, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Benders, K.E.; van Weeren, P.R.; Badylak, S.F.; Saris, D.B.; Dhert, W.J.; Malda, J. Extracellular matrix scaffolds for cartilage and bone regeneration. Trends Biotechnol. 2013, 31, 169–176. [Google Scholar] [CrossRef]
- Ni, S.; Zhuo, Z.; Pan, Y.; Yu, Y.; Li, F.; Liu, J.; Wang, L.; Wu, X.; Li, D.; Wan, Y.; et al. Recent Progress in Aptamer Discoveries and Modifications for Therapeutic Applications. ACS Appl. Mater. Interfaces 2021, 13, 9500–9519. [Google Scholar] [CrossRef]
- Ulrich, T.A.; Jain, A.; Tanner, K.; MacKay, J.L.; Kumar, S. Probing cellular mechanobiology in three-dimensional culture with collagen–agarose matrices. Biomaterials 2010, 31, 1875–1884. [Google Scholar] [CrossRef]
- Chen, S.; Shi, Y.; Zhang, X.; Ma, J. Evaluation of BMP-2 and VEGF loaded 3D printed hydroxyapatite composite scaffolds with enhanced osteogenic capacity in vitro and in vivo. Mater. Sci. Eng. C 2020, 112, 110893. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Meng, C.; Ding, Q.; Yu, K.; Zhang, X.; Zhang, W.; Tian, W.; Zhang, Q.; Guo, X.; Wu, B. In situ bone regeneration with sequential delivery of aptamer and BMP2 from an ECM-based scaffold fabricated by cryogenic free-form extrusion. Bioact. Mater. 2021, 6, 4163–4175. [Google Scholar] [CrossRef] [PubMed]
- Fuerkaiti, S.N.; Çakmak, A.S.; Karaaslan, C.; Gümüşderelioğlu, M. Enhanced osteogenic effect in reduced BMP-2 doses with siNoggin transfected pre-osteoblasts in 3D silk scaffolds. Int. J. Pharm. 2022, 612, 121352. [Google Scholar] [CrossRef]
- Cheung, Y.-W.; Kwok, J.; Law, A.W.; Watt, R.M.; Kotaka, M.; Tanner, J.A. Structural basis for discriminatory recognition of Plasmodium lactate dehydrogenase by a DNA aptamer. Proc. Natl. Acad. Sci. USA 2013, 110, 15967–15972. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ → 3′) | KD (ELONA) (nM) |
|---|---|---|
| BNA1 | CGTACGGTCGACGCTAGCCGCGGTCCTAGAGCGGACGCCGGGGGGGTGCCCGACACGTGGAGCTCGGATCC | NA |
| BNA2 | CGTACGGTCGACGCTAGCATCAGCTCTCTGGGCTGAGTGGGTGGTGTGGCACGCACGTGGAGCTCGGATCC | 22.30 ± 1.97 |
| BNA3 | CGTACGGTCGACGCTAGCTTTCTAAAATTTTCAAAAAAGATCTCGAAAAGCAACACGTGGAGCTCGGATCC | NA |
| BA1 | CGTACGGTCGACGCTAGCAAAACAAAAAAAAATAAAAAAAAAAAGACTAAAGACACGTGGAGCTCGGATCC | 6.95 ± 0.53 |
| BA2 | CGTACGGTCGACGCTAGCAAAAAAAAATCTAAAAACAAGAAAAGAAAAACAAACACGTGGAGCTCGGATCC | 20.33 ± 4.73 |
| BA3 | CGTACGGTCGACGCTAGCAAAAAAGAAAAATAAAAAACAAGAGAAACTAAAAACACGTGGAGCTCGGATCC | NA |
| BA4 | CGTACGGTCGACGCTAGCATAATAAGACAAGAAAAAAAGAACAAAAAAAAGAACACGTGGAGCTCGGATCC | NA |
| BA5 | CGTACGGTCGACGCTAGCAAAAACAAAAAGGAAAA-ACCAAAAAAACATAAAAACACGTGGAGCTCGGATCC | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Kinghorn, A.B.; Wang, L.; Bhuyan, S.K.; Shiu, S.C.-C.; Tanner, J.A. The Evolution and Application of a Novel DNA Aptamer Targeting Bone Morphogenetic Protein 2 for Bone Regeneration. Molecules 2024, 29, 1243. https://doi.org/10.3390/molecules29061243
Liu M, Kinghorn AB, Wang L, Bhuyan SK, Shiu SC-C, Tanner JA. The Evolution and Application of a Novel DNA Aptamer Targeting Bone Morphogenetic Protein 2 for Bone Regeneration. Molecules. 2024; 29(6):1243. https://doi.org/10.3390/molecules29061243
Chicago/Turabian StyleLiu, Mengping, Andrew B. Kinghorn, Lin Wang, Soubhagya K. Bhuyan, Simon Chi-Chin Shiu, and Julian A. Tanner. 2024. "The Evolution and Application of a Novel DNA Aptamer Targeting Bone Morphogenetic Protein 2 for Bone Regeneration" Molecules 29, no. 6: 1243. https://doi.org/10.3390/molecules29061243
APA StyleLiu, M., Kinghorn, A. B., Wang, L., Bhuyan, S. K., Shiu, S. C.-C., & Tanner, J. A. (2024). The Evolution and Application of a Novel DNA Aptamer Targeting Bone Morphogenetic Protein 2 for Bone Regeneration. Molecules, 29(6), 1243. https://doi.org/10.3390/molecules29061243