
Sesquiterpenoids from Meliaceae Family and Their Biological Activities


Abstract
1. Introduction
2. Methodology and Botany
3. Phytochemistry
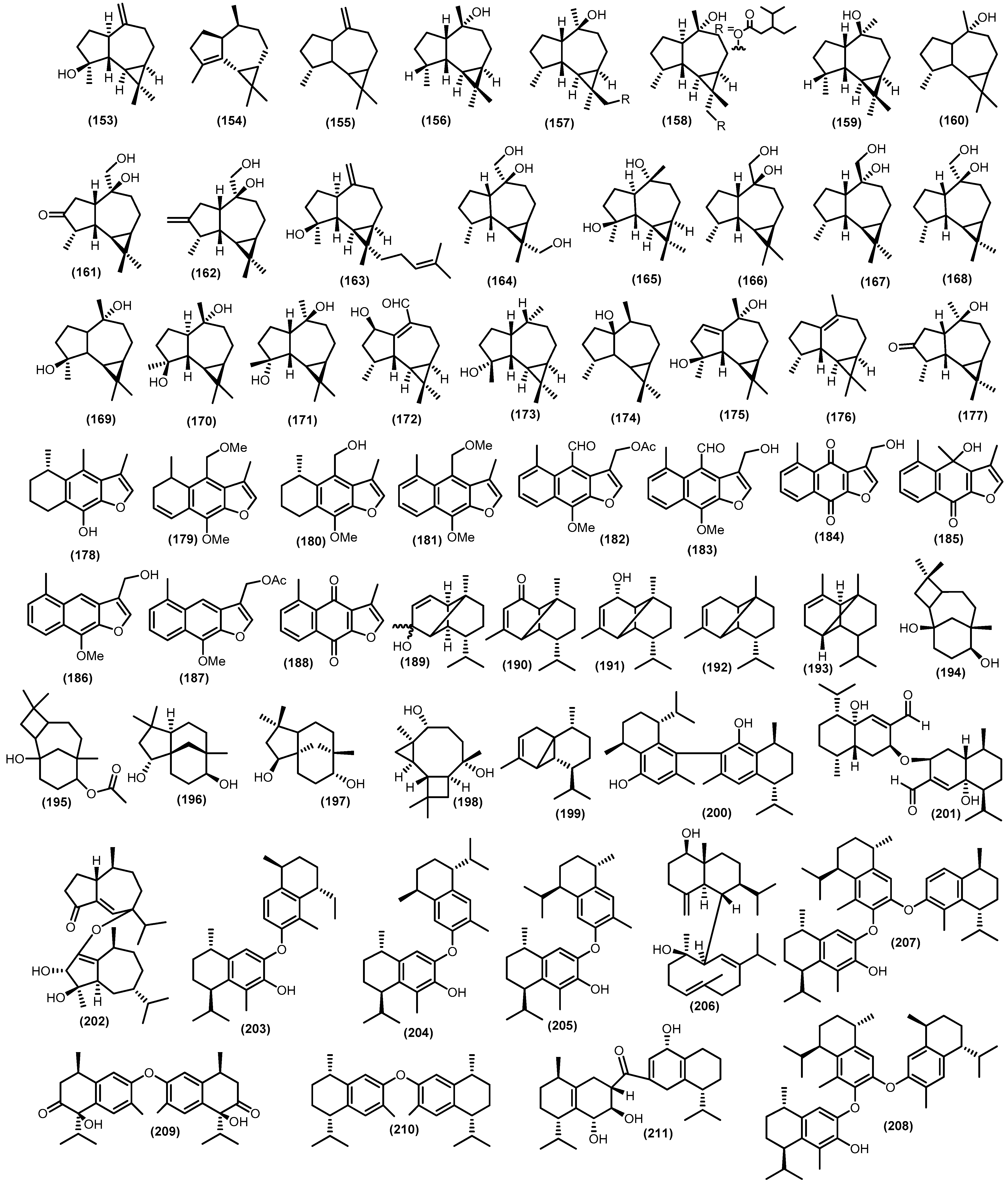
3.1. Overview of the Sesquiterpenoids Isolated from Meliaceae Family
3.2. Isolated Sesquiterpenoids
3.2.1. Acyclic Sesquiterpenoids
3.2.2. Monocyclic Sesquiterpenoids
3.2.3. Bicyclic Sesquiterpenoids
3.2.4. Tricyclic and Other Sesquiterpenoids
3.3. Volatile Sesquiterpenoids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
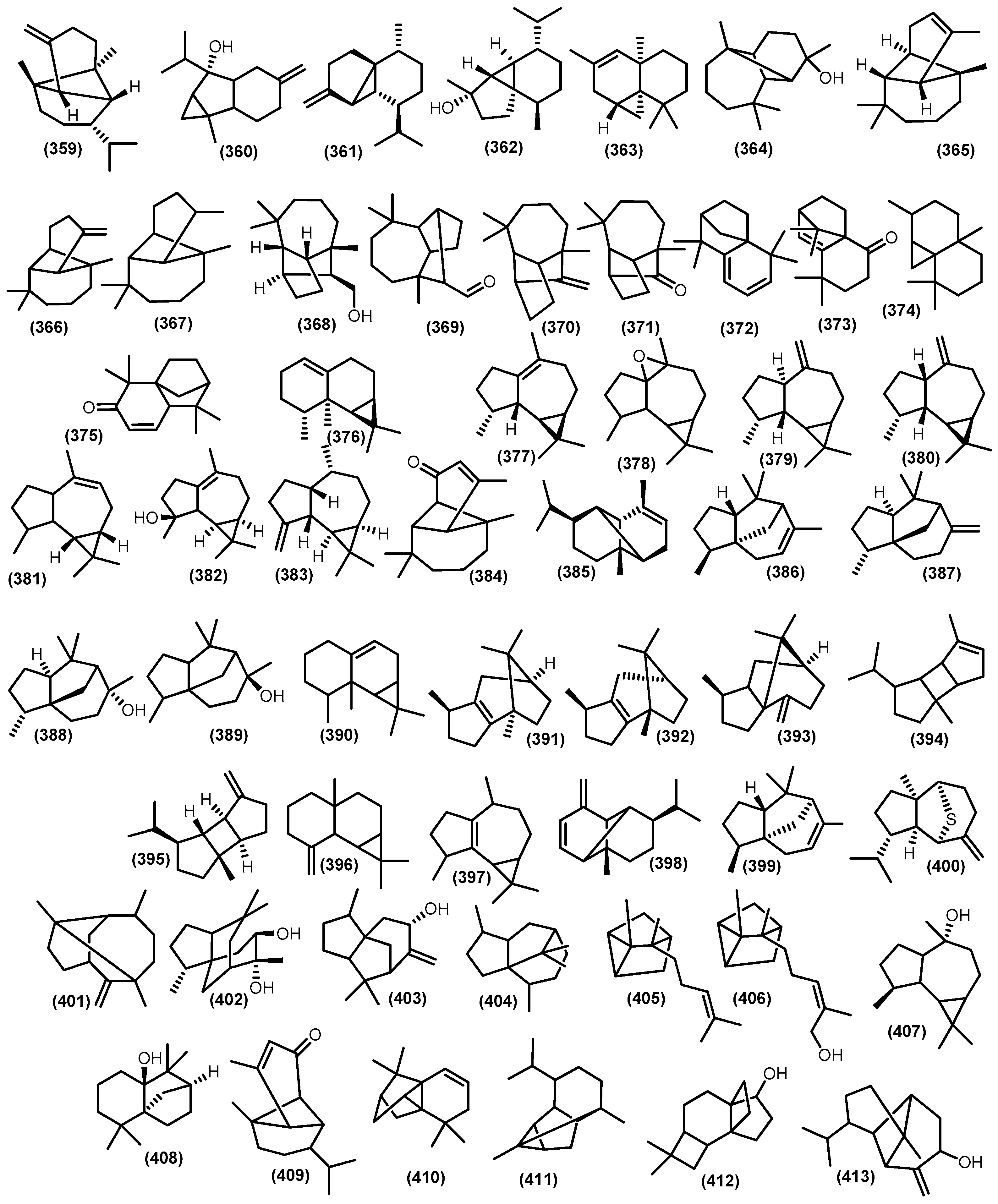{kind=link}
| Species | Part of Plant | Volatile Sesquiterpenoids | Biological Activity of Essential Oil | Major Compounds | Ref. |
|---|---|---|---|---|---|
| N. alata Wight and Arn. | Root | 214, 222, 13, 226, 228, 25, 230, 233, 251, 131, 132, 259, 264, 150, 270, 275, 276, 277, 278, 119, 118, 114, 286, 287, 193, 361, 363, 367, 368, 369, 374 | Antimicrobial activity against Sa, Bs, Pv, Ec, and Kp with inhibition zone Sa (11 mm); Bs (12 mm); Pv (10 mm); Ec (13 mm); Kp (11 mm), respectively | 259, 131, 256 | [121] |
| N. alata Wight and Arn. | Whole plant | 224, 241, 242, 251, 252, 131, 132, 272, 277, 278, 284, 285, 3, 291, 293, 296, 298, 358, 361, 364, 373, 380, 381, 384 | Not reported | 132, 251, 131, 3 | [124] |
| T. sinensis (A.Juss.) M. Roem. | Leaves | 215, 220, 221, 13, 4, 240, 131, 132, 262, 263, 280, 281, 104, 3, 34, 329, 193, 199, 361, 155, 153, 389, 395 | Antimicrobial activity against Sa, Sp, Ec, Pa, Sf, and St with MIC values Sa (1.57 µg/mL); Sp (1.57 µg/mL); Ec (3.13 µg/mL); Pa (3.13 µg/mL); Sf (12.5 µg/mL); St (6.25 µg/mL) | 131 | [125] |
| T. sinensis (A.Juss.) M. Roem. | Roots | 213, 13, 232, 237, 254, 131, 132, 260, 261, 294, 305, 153, 386, 388, 159 | Cytotoxic effect against 786-O and Caki-1 cell lines for 786-O (%viability cell 41.86 at 250 ppm) and Caki-1 (%viability cell 44.73% at 250 ppm) | 153 | [138] |
| T. sinensis (A.Juss.) M. Roem. | Leaves | 220, 131, 118, 199, 370, 155, 385, 390, 391 | Not reported | 131 | [142] |
| T. sinensis (A.Juss.) M. Roem. | Leaves | 219, 220, 221, 222, 236, 4, 253, 131, 132, 147, 282, 104, 283, 307, 310, 315, 316, 193, 199, 361, 372, 379, 392, 395, 410 | Antibacterial activity against MSSA (MIC 0.125 mg/mL) and MRSA (MIC 1 mg/mL) Cytotoxic activity against SGC7902 (IC50 70.38 μg/mL); HepG2 (IC50 82.2 μg/mL); HT29 (IC50 99.94 μg/mL) | 131 | [127] |
| T. sinensis (A.Juss.) M. Roem. | Leaves | 215, 220, 221, 227, 228, 25, 229, 131, 305, 307, 332, 153, 160 | Antidiabetic properties effect on medium glucose consumption in the 3T3-L1 cells. The essential oil could increase the use of medium glucose to approximately 50% at 50 µg/mL. | 227 | [129] |
| T. ciliata M. Roem. | Leaves | 220, 13, 227, 25, 4, 255, 131, 259, 261, 265, 266, 268, 119, 118, 104, 114, 317, 330, 361, 362, 370, 155, 153, 156, 160, 154, 383, 385, 395, 159, 401 | Not reported | 314, 131, 25 | [136] |
| T. ciliata M. Roem. | Leaves | 219, 220, 221, 245, 253, 263, 280, 281, 331, 199, 361, 380, 379, 385, | Antidepressant activity using FST and TST immobility as a result of FST (decrease immobility duration > 50 s); TST (decrease immobility duration > 50 s) | 220, 361, 222 | [143] |
| A. odorata Lour. | Stems | 13, 25, 243, 131, 257 | Antimicrobial activity against Bo, Po, and Rs. Bo (MIC values 0.0625–0.5 mg/mL), Po (MIC values 0.0625–0.5 mg/mL), Rs (MIC values 0.0625–0.5 mg/mL); Bo (MFC values 0.25–1 mg/mL), Po (MFC values 0.25–1 mg/mL), Rs (MFC values 0.25–1 mg/mL). | 25 | [144] |
| A. indica (Hook.fil) Harms | Flowers | 221, 13, 147, 148, 119, 118, 104, 283, 110, 317, 126, 321, 193, 199, 155, 380, 174, 156, 160, 154, 176, 159 | Antimicrobial activity against Bs, Ca, and Mg with inhibition zone Bs (10.5 ± 0.5 mm); Ca (14 ± 0.5 mm); Mg (11.0 ± 1.0 mm) | 104, 193 | [145] |
| A. indica (Hook.fil) Harms | Flowers | 228, 245, 257, 396 | Not reported | 241 | [146] |
| C. fissilis Vell. | Leaves | 220, 13, 227, 25, 4, 131, 132, 147, 148, 104, 114, 110, 296, 38, 34, 317, 319, 193, 199, 155, 153, 156, 160, 159 | Antibacterial activity against Sa, Ec, Pa with inhibition zone Sa (9.3 ± 0.6 mm); Ec (6.7 ± 0.6 mm); Pa (inactive) | 314, 131 | [137] |
| C. mexicana M.Roem | Leaves and Stembarks | 221, 13, 227, 228, 25, 230, 231, 243, 245, 90, 131, 132, 282, 104, 283, 291, 305, 321, 324, 99, 155, 380, 153, 160, 397, 398 | Not reported | 131 | [141] |
| C. odorata L. | Leaves | 220, 13, 223, 225, 4, 253, 131, 132, 271, 104, 289, 290, 304, 318, 325, 326, 193, 199, 399, 400, 405 | Not reported | 405 | [135] |
| C. odorata L. | Essential oil | 221, 13, 228, 4, 238, 131, 147, 267, 104, 283, 305, 34, 317, 319, 128, 321, 193, 199, 376, 377, 380 | Not reported | - | [128] |
| C. tabularis A.Juss | Leaves | 218, 220, 13, 245, 248, 247, 253, 131, 132, 261, 267, 118, 104, 3, 296, 38, 34, 128, 327, 193, 199, 338, 380, 156, 154, 383, 395, 159 | Not reported | 132 | [134] |
| C. baccifera (Roth) Miq | Leaves | 220, 13, 230, 231, 132, 258, 261, 104, 303, 311, 312, 313, 314, 320, 360, 367, 375, 155, 380, 153, 382, 397 | Not reported | 131 | [131] |
| T. connaroides (Wight and Arn.) Bentv. | Roots | 131, 279, 104, 114, 38, 301, 302, 128, 328, 193, 386 | Not reported | 328 | [132] |
| T. connaroides (Wight and Arn.) Bentv. | Barks, Leaves, Roots | 219, 220, 25, 4, 131, 147, 267, 282, 104, 283, 288, 305, 308, 126, 328, 193, 199, 361, 365, 155, 154, 383, 385, 387, 392, 394, 397, 405, 411 | Antiplasmodial activity as a result of weak antiplasmodial with IC50 range 2–22 µg/mL. | 193, 131, 328 | [130] |
| K. grandifolia C.DC. | Flowers | 220, 13, 224, 228, 253, 132, 256, 272, 273, 274, 119, 285, 294, 128, 328, 193, 361, 365, 371, 160, 154, 393, 176, 159, 411 | Cytotoxic activity against HepG-2 (IC50 21.6 μg/mL); MCF-7 (IC50 26.1 μg/mL); HCT-116 (37.6 μg/mL) | 132, 250, 13 | [133] |
| K. senegalensis (Desv.) A.Juss. | Flowers | 221, 13, 224, 131, 132, 261, 119, 114, 296, 335, 361, 371, 153, 160, 387, 394, 159, 403 | Cytotoxic activity against HepG-2 (IC50 61.1 μg/mL); MCF-7 (IC50 79.7 μg/mL); HCT-116 (61.0 μg/mL) | 132, 131 | |
| M. azedarach L. | Leaves | 253, 131, 132, 317 | Not reported | 131 | [123] |
| M. azedarach L. | Flowers | 213, 253, 131, 291, 317 | Antimicrobial activity against Sa, Pv, Pa, Ec, Se, and Kp with MIC values Sa (150 µL/mL); Pv (150 µL/mL); Pa (150 µL/mL); Ec (175 µL/mL); Se (175 µL/mL); Kp (175 µL/mL). | 210, 209 | [126] |
| S. macrophylla G.King | Flowers | 25, 132, 148, 104, 126, 333, 334, 193, 199, 361, 155, 153, 395 | Not reported | 131 | [122] |
| S. macrophylla G.King | Leaves | 227, 25, 238, 4, 13, 259, 283, 308, 317, 193, 361, 380, 383, 395 | Not reported | 25 | [140] |
| S. macrophylla G.King | Leaves | 220, 13, 224, 226, 227, 25, 131, 265, 147, 272, 273, 119, 280, 281, 104, 283, 285, 294, 317, 319, 128, 193, 199, 361, 380, 153, 154, 383, 385, 395 | Not reported | 25 | [147] |
| G. convergen T.D.Penn. | Branches | 356, 357, 358, 409 | Not reported | 405, 193 | [148] |
| G. kunthiana A.Juss | Leaves | 153, 182, 220, 193, 230, 231, 254 | Antimicrobial activity against Ec, Pa, Se, Pm, Kp, Sa, Ef, Se, Bs, and Ca with MIC values Ec (Not active); Pa (7000 mg/mL); Se (7000 mg/mL); Pm (7000 mg/mL); Kp (inactive); Sa (13.6 mg/mL); Ef (437.5 mg/mL); Se (3500 mg/mL); Bs (875 mg/mL); Ca (1750 mg/mL) Antioxidant activity using DPPH scavenging capacity with IC50 17.54 ± 0.18 µg/mL | 231, 256 | [149] |
| G. macrophylla G.King | Leaves | 156, 131, 148, 119, 110 | Not reported | 89 | [32] |
| G. macrophylla G.King | Leaves | 347, 348, 349, 350, 351, 352, 353, 354, 355 | Anti-inflammatory activity using macrophages BALB/c mice with CC 17.7 > 100 µg/mL. | 264 | [150] |
| G. macrophylla G.King | Fruits | 193, 361, 153, 160, 176 | Not reported | 193, 176, 283 | [151] |
| G. cedreta (A.Chev.) Pellegr. | Barks | 193, 370, 380, 160, 385, 176, 131, 147, 267, 118, 104, 114, 296 | Not reported | 131 | [152] |
| G. macrophylla G.King | Leaves | 359, 388, 13, 227, 147, 267, 118, 281, 104, 297, 346, 385 | Not reported | 297, 385, 104, 267, 131 | [153] |
4. Ethnobotany and Medicinal Uses
5. Biological Activity
5.1. Cytotoxic Activity
5.2. Anti-Inflammatory
5.3. Antioxidant
5.4. Antidiabetic
5.5. Antimicrobial
5.6. Antiviral
5.7. Antiplasmodial
5.8. Antidepressant-Like Activity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muellner-Riehl, A.N.; Rojas-Andrés, B.M. Biogeography of Neotropical Meliaceae: Geological Connections, Fossil and Molecular Evidence Revisited. Rev. Bras. Bot. 2022, 45, 527–543. [Google Scholar] [CrossRef]
- Luo, J.; Sun, Y.; Li, Q.; Kong, L. Research Progress of Meliaceous Limonoids from 2011 to 2021. Nat. Prod. Rep. 2022, 39, 1325–1365. [Google Scholar] [CrossRef]
- Naini, A.A.; Mayanti, T.; Harneti, D.; Darwati; Nurlelasari; Maharani, R.; Farabi, K.; Herlina, T.; Supratman, U.; Fajriah, S.; et al. Sesquiterpenoids and Sesquiterpenoid Dimers from the Stem Bark of Dysoxylum parasiticum (Osbeck) Kosterm. Phytochemistry 2023, 205, 113477. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Inoue, A.; Sastrapradja, S.; Hayashi, Y. (+)-8-Hydroxycalamenene: A Fish-Poison Principle of Dysoxylum acutangulum and D. alliaceum. Phytochemistry 1983, 22, 2083–2085. [Google Scholar] [CrossRef]
- Traore, M.; Zhai, L.; Chen, M.; Olsen, C.E.; Odile, N.; Pierre, G.; Bosco, O.; Robert, G.; Christensen, S.B. Cytotoxic Kurubasch Aldehyde from Trichilia emetica. Nat. Prod. Res. 2007, 21, 13–17. [Google Scholar] [CrossRef]
- Harneti, D.; Ayu Permatasari, A.; Anisshabira, A.; Arofatus Naini, A.; Mayanti, T.; Maharani, R.; Safari, A.; Tatang Hidayat, A.; Farabi, K.; Supratman, U.; et al. Sesquiterpenoids from the Stem Bark of Aglaia grandis. Nat. Prod. Sci. 2022, 28, 6–12. [Google Scholar]
- Ara, I.; Siddiqui, B.S.; Faizi, S.; Siddiqui, S. Structurally Novel Diterpenoid Constituents from the Stem Bark of Azadirachta indica (Meliaceae). J. Chem. Soc. Perkin Trans. 1 1989, 2, 343–345. [Google Scholar] [CrossRef]
- Mulholland, D.A.; Monkhe, T.V.; Pegel, K.H.; Taylor, D.A.H. Limonoids and Diterpenoids from Dysoxylum spectabile (Meliaceae). Biochem. Syst. Ecol. 1999, 27, 1999–2001. [Google Scholar]
- Gu, J.; Qian, S.Y.; Zhao, Y.L.; Cheng, G.G.; Hu, D.B.; Zhang, B.H.; Li, Y.; Liu, Y.P.; Luo, X.D. Prenyleudesmanes, Rare Natural Diterpenoids from Dysoxylum densiflorum. Tetrahedron 2014, 70, 1375–1382. [Google Scholar] [CrossRef]
- Fang, F.H.; Huang, W.J.; Zhou, S.Y.; Han, Z.Z.; Li, M.Y.; Liu, L.F.; Wu, X.Z.; Yao, X.J.; Li, Y.; Yuan, C.S. Aphapolins A and B: Two Nemoralisin Diterpenoids Isolated from Aphanamixis polystachya (Wall.) R. Parker. Eur. J. Org. Chem. 2017, 2017, 4429–4433. [Google Scholar] [CrossRef]
- Supratman, U.; Naibaho, W.; Salam, S.; Maharani, R.; Hidayat, A.T.; Harneti, D.; Nurlelasari; Shiono, Y. Cytotoxic Triterpenoids from the Bark of Chisocheton patens Blume (Meliaceae). Phytochem. Lett. 2019, 30, 81–87. [Google Scholar] [CrossRef]
- Kurimoto, S.I.; Takaishi, Y.; Ahmed, F.A.; Kashiwada, Y. Triterpenoids from the Fruits of Azadirachta indica (Meliaceae). Fitoterapia 2014, 92, 200–205. [Google Scholar] [CrossRef]
- Harneti, D.; Tjokronegoro, R.; Safari, A.; Supratman, U.; Loong, X.M.; Mukhtar, M.R.; Mohamad, K.; Awang, K.; Hayashi, H. Cytotoxic Triterpenoids from the Bark of Aglaia smithii (Meliaceae). Phytochem. Lett. 2012, 5, 496–499. [Google Scholar] [CrossRef]
- Happi, G.M.; Talontsi, F.M.; Laatsch, H.; Zühlke, S.; Ngadjui, B.T.; Spiteller, M.; Kouam, S.F. Seco-Tiaminic Acids B and C: Identification of Two Novel 3,4-Seco-Tirucallane Triterpenoids Isolated from the Root of Entandrophragma congoënse (Meliaceae). Fitoterapia 2018, 124, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Gu, Z.M.; Chang, C.J.; Wood, K.V.; McLaughlin, J.L. Meliavolkenin, a New Bioactive Triterpenoid from Melia volkensii (Meliaceae). Bioorganic Med. Chem. 1995, 3, 383–390. [Google Scholar] [CrossRef]
- Tsamo, A.T.; Melong, R.; Mkounga, P.; Nkengfack, A.E. Rubescins I and J, Further Limonoid Derivatives from the Stem Bark of Trichilia rubescens (Meliaceae). Nat. Prod. Res. 2019, 33, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Kowa, T.K.; Tchokouaha, L.R.Y.; Cieckiewicz, E.; Philips, T.J.; Dotse, E.; Wabo, H.K.; Tchinda, A.T.; Tane, P.; Frédérich, M. Antileishmanial and Cytotoxic Activities of a New Limonoid and a New Phenyl Alkene from the Stem Bark of Trichilia gilgiana (Meliaceae). Nat. Prod. Res. 2020, 34, 3182–3188. [Google Scholar] [CrossRef] [PubMed]
- Miranda, R.N.C., Jr.; Dolabela, M.F.; Da Silva, M.N.; Póvoa, M.M.; Maia, J.G.S. Antiplasmodial Activity of the Andiroba (Carapa guianensis Aubl., Meliaceae) Oil and Its Limonoid-Rich Fraction. J. Ethnopharmacol. 2012, 142, 679–683. [Google Scholar] [CrossRef]
- Supriatno; Nurlelasari; Herlina, T.; Harneti, D.; Maharani, R.; Hidayat, A.T.; Mayanti, T.; Supratman, U.; Azmi, M.N.; Shiono, Y. A New Limonoid from Stem Bark of Chisocheton pentandrus (Meliaceae). Nat. Prod. Res. 2018, 32, 2610–2616. [Google Scholar] [CrossRef]
- Liu, J.; Yang, S.P.; Su, Z.S.; Lin, B.D.; Wu, Y.; Yue, J.M. Limonoids from the Stems of Toona ciliata Var. Henryi (Meliaceae). Phytochemistry 2011, 72, 2189–2196. [Google Scholar] [CrossRef]
- Peng, L.; Fu, W.X.; Zeng, C.X.; Zhou, L.; Bao, M.F.; Cai, X.H. Two New Lignans from Twigs of Aglaia odorata. J. Asian Nat. Prod. Res. 2016, 18, 147–152. [Google Scholar] [CrossRef]
- Wang, L.; Li, F.; Yang, C.Y.; Khan, A.A.; Liu, X.; Wang, M.K. Neolignans, Lignans and Glycoside from the Fruits of Melia toosendan. Fitoterapia 2014, 99, 92–98. [Google Scholar] [CrossRef]
- Greger, H.; Pacher, T.; Brem, B.; Bacher, M.; Hofer, O. Insecticidal Flavaglines and Other Compounds from Fijian aglaia Species. Phytochemistry 2001, 57, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.D.; Weiss, U.; Silverton, J.V. The Structure of Rohitukine, the Main Alkaloid of Amoora rohituka (Syn. Aphanamixis polystachya) (Meliaceae). Tetrahedron Lett. 1979, 20, 721–724. [Google Scholar] [CrossRef]
- Kumar, V.; Gupta, M.; Gandhi, S.G.; Bharate, S.S.; Kumar, A.; Vishwakarma, R.A.; Bharate, S.B. Anti-Inflammatory Chromone Alkaloids and Glycoside from Dysoxylum binectariferum. Tetrahedron Lett. 2017, 58, 3974–3978. [Google Scholar] [CrossRef]
- Naini, A.A.; Mayanti, T.; Supratman, U. Triterpenoids from Dysoxylum Genus and Their Biological Activities. Arch. Pharm. Res. 2022, 45, 63–89. [Google Scholar] [CrossRef]
- Hilmayanti, E.; Nurlelasari; Supratman, U.; Kabayama, K.; Shimoyama, A.; Fukase, K. Limonoids with Anti-Inflammatory Activity: A Review. Phytochemistry 2022, 204, 11346. [Google Scholar] [CrossRef]
- Harneti, D.; Supratman, U. Phytochemistry and Biological Activities of Aglaia Species. Phytochemistry 2021, 181, 112540. [Google Scholar] [CrossRef]
- Modzelewska, A.; Sur, S.; Kumar, S.K.; Khan, S.R. Sesquiterpenes: Natural Products That Decrease Cancer Growth. Curr. Med. Chem. Anti-Cancer Agents 2005, 5, 477–499. [Google Scholar] [CrossRef] [PubMed]
- Abidullah, S.; Rauf, A.; Khan, S.W.; Ayaz, A.; Liaquat, F.; Saqib, S. A Comprehensive Review on Distribution, Paharmacological Uses and Biological Activities of Argyrolobium roseum (Cambess.). Jaub. Spach. Acta Ecol. Sin. 2022, 42, 198–205. [Google Scholar] [CrossRef]
- Nagasampagi, B.A.; Yankov, L.; Dev, S. Sesquiterpenoids from the Wood of Cedrela toona Roxb; Partial Synthesis of t-Muurolol, t-Cadinol and Cubenol; Structures of δ-Cadinene and δ-Cadinol. Tetrahedron Lett. 1968, 9, 1913–1918. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Soares, M.G.; Batista-Pereira, L.G.; Silva, M.F.G.F.; Corrêa, A.G.; Fernandes, J.B.; Vieira, P.C.; Roque, N.F. Volatile Oil from Guarea macrophylla Ssp. Tuberculata: Seasonal Variation and Electroantennographic Detection by Hypsipyla grandella. Phytochemistry 2006, 67, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Panshin, A.J. Comparative Anatomy of the Woods of the Meliaceae, Sub-Family Swietenioideae. Am. J. Bot. 1933, 20, 638–668. [Google Scholar] [CrossRef]
- Shu, H.Z.; Peng, C.; Bu, L.; Guo, L.; Liu, F.; Xiong, L. Bisabolane-Type Sesquiterpenoids: Structural Diversity and Biological Activity. Phytochemistry 2021, 192, 112927. [Google Scholar] [CrossRef]
- Kumeta, Y.; Ito, M. Characterization of α-Humulene Synthases Responsible for the Production of Sesquiterpenes Induced by Methyl Jasmonate in Aquilaria Cell Culture. J. Nat. Med. 2016, 70, 452–459. [Google Scholar] [CrossRef]
- Gaikwad, N.W.; Madyastha, K.M. Biosynthesis of β-Substituted Furan Skeleton in the Lower Furanoterpenoids: A Model Study. Biochem. Biophys. Res. Commun. 2002, 290, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Zhang, P.; Tang, P.; Wang, C.; Kong, L.; Luo, J. Acyclic Diterpene and Norsesquiterpene from the Seed of Aphanamixis polystachya. Fitoterapia 2020, 142, 104518. [Google Scholar] [CrossRef]
- Zhang, R.; He, H.P.; Di, Y.T.; Li, S.L.; Zuo, G.Y.; Zhang, Y.; Hao, X.J. Chemical Constituents from Aphanamixis grandifolia. Fitoterapia 2013, 92, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Krishnappa, S.; Dev, S. Sesquiterpenes from Lansium Anamalayanum. Phytochemistry 1973, 12, 823–825. [Google Scholar] [CrossRef]
- Mulholland, D.A.; Iourine, S.; Taylor, D.A.H. Sesquiterpenoids from Dysoxylum Schiffneri. Phytochemistry 1998, 47, 1421–1422. [Google Scholar] [CrossRef]
- Mulholland, D.A.; McFarland, K.; Randrianarivelojosia, M. Sesquiterpenoid Derivatives from Cipadessa boiviniana (Meliaceae). Biochem. Syst. Ecol. 2006, 34, 365–369. [Google Scholar] [CrossRef]
- Liu, H.B.; Zhang, C.R.; Dong, S.H.; Yang, S.P.; Sun, Q.; Geng, M.Y.; Yue, J.M. Sesquiterpenes from Dysoxylum oliganthum and Dysoxylum excelsum. J. Asian Nat. Prod. Res. 2012, 14, 224–234. [Google Scholar] [CrossRef]
- Febriandari, R. Tukiran Suatu Senyawa Seskuiterpen Dari Ekstrak Kloroform Kulit Batang Tumbuhan Aglaia elaeagnoidea (A. Juss) Benth. UNESA J. Chem. 2013, 2, 100–102. [Google Scholar]
- Xia, M.J.; Zhang, M.; Li, S.W.; Cai, Z.F.; Zhao, T.S.; Liu, A.H.; Luo, J.; Zhang, H.Y.; Li, J.; Guo, Y.W.; et al. Anti-Inflammatory and PTP1B Inhibitory Sesquiterpenoids from the Twigs and Leaves of Aglaia lawii. Fitoterapia 2022, 162, 105260. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Brochini, C.B.; Roque, N.F. Terpenes from Leaves of Guarea macrophylla (Meliaceae). Phytochemistry 2000, 55, 727–731. [Google Scholar] [CrossRef]
- Núñez, C.V.; Roque, N.F. Sesquiterpenes from the Stem Bark of Guarea guidonia (L.) Sleumer (Meliaceae). J. Essent. Oil Res. 1999, 11, 439–440. [Google Scholar] [CrossRef]
- Pupo, M.T.; Adorno, M.A.T.; Vieira, P.C.; Fernandes, J.B.; Da Silva, M.F.D.G.F.; Piranic, J.R. Terpenoids and Steroids from Trichilia Species. J. Braz. Chem. Soc. 2002, 13, 382–388. [Google Scholar] [CrossRef]
- Garcez, F.R.; Núñez, C.V.; Garcez, W.S.; Almeida, R.M.; Roque, N.F. Sesquiterpenes, Limonoid and Coumarin from the Wood Bark of Guarea guidonia. Planta Med. 1998, 64, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Mahdzir, M.A.; Shilpi, J.A.; Mahmud, N.; Ramasamy, S.; Awang, K. Chemical Constituents from Walsura pinnata (Meliaceae). Nat. Prod. Commun. 2017, 12, 1934578X1701200. [Google Scholar] [CrossRef]
- Nangmo, K.P.; Tsamo, T.A.; Zhen, L.; Mkounga, P.; Akone, S.H.; Tsabang, N.; Müller, W.E.G.; Marat, K.; Proksch, P.; Nkengfack, A.E. Chemical Constituents from Leaves and Root Bark of Trichilia monadelpha (Meliaceae). Phytochem. Lett. 2018, 23, 120–126. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Reis, A.A.; Roque, N.F. Chemical Composition from Volatile Oil of the Stem Bark of Guarea macrophylla Vahl. Ssp. Tuberculata vellozo (Meliaceae). Flavour Fragr. J. 2002, 17, 255–257. [Google Scholar] [CrossRef]
- Ragasa, C.Y.; Labrador, P.; Rideout, J.A. Antimicrobial Terpenoids from Lansium domesticum. Philipp. Agric. Sci. 2006, 89, 101–105. [Google Scholar]
- Sofian, F.F.; Subarnas, A.; Hakozaki, M.; Uesugi, S.; Koseki, T.; Shiono, Y. Bidysoxyphenols A–C, Dimeric Sesquiterpene Phenols from the Leaves of Dysoxylum parasiticum (Osbeck) Kosterm. Fitoterapia 2022, 158, 105157. [Google Scholar] [CrossRef]
- Zeng, Q.; Ye, J.; Cheng, X.; Qin, J.; Jin, H.; Zhang, W.D. Chemical Constituents from Aphanamixis grandifolia. Chem. Nat. Compd. 2013, 49, 486–492. [Google Scholar]
- D’Ambola, M.; Malafronte, N.; Gualtieri, M.; Hernández, V.; Vassallo, A.; Severino, L. A Novel Tirucallane-Type Triterpene and Sesquiterpene from Trichilia Maynasiana. Nat. Prod. Commun. 2016, 11, 447–448. [Google Scholar] [CrossRef]
- Fadhilah, K.; Wahyuono, S.; Astuti, P. A Sesquiterpene Aldehyde Isolated from Ethyl Acetate Extract of Lansium Domesticum Fruit Peel. Indones. J. Pharm. 2021, 32, 394–398. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Brochini, C.B.; Roque, N.F. Terpenoids from Guarea guidonia. Phytochemistry 2002, 60, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Brochini, C.B.; Roque, N.F.; Lago, J.H.G. Minor Sesquiterpenes from the Volatile Oil from Leaves of Guarea guidonia Sleumer (Meliaceae). Nat. Prod. Res. 2009, 23, 1615–1620. [Google Scholar] [CrossRef]
- Kurniasih, N.; Milawati, H.; Fajar, M.; Hidayat, A.T.; Abdulah, R.; Harneti, D.; Supratman, U.; Azmi, M.N. Sesquiterpenoid Compounds from The Stembark of Aglaia minahassae (Meliaceae). Molekul 2018, 13, 56–62. [Google Scholar] [CrossRef]
- Fu, L.R.; Ma, Q.Y.; Huang, S.Z.; Dai, H.F.; Guo, Z.K.; Yu, Z.F.; Zhao, Y.X. Terpenoids and Their Anti-Feedant Activity from Cipadessa cinerascens. J. Asian Nat. Prod. Res. 2014, 16, 1054–1059. [Google Scholar] [CrossRef]
- Lin, L.G.; Tang, C.P.; Ke, C.Q.; Zhang, Y.; Ye, Y. Terpenoids from the Stems of Cipadessa baccifera. J. Nat. Prod. 2008, 71, 628–632. [Google Scholar] [CrossRef]
- Soares, L.R.; De Queiroz E Silva, A.C.; Freire, T.V.; Garcez, F.R.; Garcez, W.S. Sesquiterpenos de Sementes de Guarea guidonia (Meliaceae). Quim. Nova 2012, 35, 323–326. [Google Scholar] [CrossRef]
- Chen, H.D.; Yang, S.P.; Liao, S.G.; Zhang, B.; Wu, Y.; Yue, J.M. Limonoids and Sesquiterpenoids from Amoora tsangii. J. Nat. Prod. 2008, 71, 93–97. [Google Scholar] [CrossRef]
- Sinaga, S.E.; Mayanti, T.; Naini, A.A.; Harneti, D.; Nurlelasari, N.; Maharani, R.; Farabi, K.; Supratman, U.; Fajriah, S.; Azmi, M.N. Sesquiterpenoids from the Stem Bark of Lansium domesticum Corr. Cv. Kokossan and Their Cytotoxic Activity against MCF-7 Breast Cancer Cell Lines. Indones. J. Chem. 2022, 22, 1035–1042. [Google Scholar] [CrossRef]
- Xie, B.J.; Yang, S.P.; Yue, J.M. Terpenoids from Dysoxylum densiflorum. Phytochemistry 2008, 69, 2993–2997. [Google Scholar] [CrossRef]
- Chenda, L.B.N.; Kouam, S.F.; Lamshöft, M.; Kusari, S.; Talontsi, F.M.; Ngadjui, B.T.; Spiteller, M. Isolation and Characterization of Six Labdane Diterpenes and One Pregnane Steroid of Turraeanthus africanus. Phytochemistry 2014, 103, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.C.; Ambrozin, A.R.P.; Fernandes, J.B.; Vieira, P.C.; Da Silva, M.F.D.G.F.; De Albuquerque, S. Trypanocidal Activity of Limonoids and Triterpenes from Cedrela fissilis. Planta Med. 2008, 74, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, D.; Liu, X.; Liu, F.; Xuan, L.; Tang, Y.; Li, W. Cytotoxicity and Polyol Pathway Inhibitory Activities of Chemical Constituents Isolated from the Pericarp of Toona sinensis. Nat. Prod. Res. 2022, 36, 1593–1598. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, F.; Ji, C.; Liu, D.; Liu, X.; Wang, R.; Li, W. Chemical Constituents of the Pericarp of Toona sinensis and Their Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 104, 104458. [Google Scholar] [CrossRef]
- Oktaviani, D.; Sukmawati, W.; Farabi, K.; Harneti, D.; Mahari, R.; Mayanti, T.; Safari, A.; Supratman, U. Terpenoids from The Stem Bark of Aglaia elaeagnoidea and Their Cytotoxic Activity against HeLa and DU145 Cancer Cell Lines. Molekul 2022, 17, 76–84. [Google Scholar] [CrossRef]
- Liu, S.; Liu, S.B.; Zuo, W.J.; Guo, Z.K.; Mei, W.L.; Dai, H.F. New Sesquiterpenoids from Aglaia odorata Var Microphyllina and Their Cytotoxic Activity. Fitoterapia 2014, 92, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Parulian, S.S.; Nurlelasari; Naini, A.A.; Hilmayanti, E.; Mayanti, T.; Harneti, D.; Maharani, R.; Farabi, K.; Supratman, U.; Anwar, R.; et al. Sesquiterpenoids from Stem Bark of Chisocheton lasiocarpus and Their Cytotoxic Activity against MCF-7 Breast Cancer Cell. Molekul 2022, 17, 413–420. [Google Scholar] [CrossRef]
- Pan, L.; Kardono, L.B.S.; Riswan, S.; Chai, H.; Carcache De Blanco, E.J.; Pannell, C.M.; Soejarto, D.D.; McCloud, T.G.; Newman, D.J.; Kinghorn, A.D. Isolation and Characterization of Minor Analogues of Silvestrol and Other Constituents from a Large-Scale Re-Collection of Aglaia foveolata. J. Nat. Prod. 2010, 73, 1873–1878. [Google Scholar] [CrossRef]
- Gu, J.; Qian, S.Y.; Cheng, G.G.; Li, Y.; Liu, Y.P.; Luo, X.D. Chemical Components of Dysoxylum densiflorum. Nat. Products Bioprospect. 2013, 3, 66–69. [Google Scholar] [CrossRef][Green Version]
- Djemgou, P.C.; Gatsing, D.; Hegazy, M.E.F.; El-Hamd Mohamed, A.H.; Ngandeu, F.; Tane, P.; Ngadjui, B.T.; Fotso, S.; Laatsch, H. Turrealabdane, Turreanone and an Antisalmonellal Agent from Turraeanthus africanus. Planta Med. 2010, 76, 165–171. [Google Scholar] [CrossRef]
- Yang, M.H.; Wang, J.S.; Kong, L.Y. Chemical Constituents of Chisocheton cumingianus. Chin. J. New Drugs 2012, 21, 555–558. [Google Scholar]
- Naini, A.A.; Mayanti, T.; Nurlelasari; Harneti, D.; Maharani, R.; Safari, A.; Hidayat, A.T.; Farabi, K.; Lesmana, R.; Supratman, U.; et al. Cytotoxic Sesquiterpenoids from Dysoxylum parasiticum (Osbeck) Kosterm. Stem Bark. Phytochem. Lett. 2022, 47, 102–106. [Google Scholar] [CrossRef]
- Rodrigues, V.F.; Carmo, H.M.; Oliveira, R.R.; Filho, R.B.; Mathias, L.; Vieira, I.J.C. Isolation of Terpenoids from Trichilia quadrijuga (Meliaceae) by Droplet Counter-Current Chromatography. Chromatographia 2009, 70, 1191–1195. [Google Scholar] [CrossRef]
- Zhang, S.N.; Huang, L.; Ma, R.J.; Yang, M.F.; Wei, B.F.; Song, H.Z.; Wang, H.S.; Tan, Q.G. Chemical Constituents from the Barks of Melia Azedarach and Their PTP1B Inhibitory Activity. Nat. Prod. Res. 2021, 35, 4442–4447. [Google Scholar] [CrossRef] [PubMed]
- Thị, N.; Ly, T.; Dũng, N.; Thị, C.; Trang, K.; Thu, L.; Diệu, N.; Hoa, L. Sesqui-và Triterpenoid Từ vỏ Trái Bòn Bon (Lansium Domesticum). Tập 2018, 21, 47–52. [Google Scholar]
- Hoffmann, J.J.; Cole, J.R.; Arora, S.K.; Bates, R.B.; Kriek, G.R. Voleneol Diacetate: A New Sesquiterpenoid from Lepidotrichilia volensii Leroy (Meliaceae). J. Org. Chem. 1978, 43, 1254–1256. [Google Scholar] [CrossRef]
- Hua, W.; Wang, X.; Li, Q.; Li, Y.; Cui, L.; Li, Y.; Kong, L.; Luo, J. Rare Noreudesmane Sesquiterpenoids from Fruits of Dysoxylum densiflorum. Fitoterapia 2023, 166, 105426. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Hasan, C.M.; Rashid, M.A. Guaiane Sesquiterpenes from Amoora rohituka. Phytochemistry 2003, 62, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Wu, S.; Wu, D. The Chemical Constituent of Amoora yunnanensis. J. Integr. Plant Biol. 2001, 43, 426–430. [Google Scholar]
- Garcez, F.R.; Garcez, W.S.; Da Silva, A.F.G.; De Cássia Bazzo, R.; Resende, U.M. Terpenoid Constituents from Leaves of Guarea kunthiana. J. Braz. Chem. Soc. 2004, 15, 767–772. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Roque, N.F. Cycloartane Triterpenoids from Guarea macrophylla. Phytochemistry 2002, 60, 329–332. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Roque, N.F. Estudo Fitoquímico Da Madeira de Guarea macrophylla (Meliaceae). Quim. Nova 2009, 32, 2351–2354. [Google Scholar] [CrossRef]
- Roux, D.; Martin, M.T.; Adeline, M.T.; Sevenet, T.; Hadi, A.H.A.; Païs, M. Foveolins A and B, Dammarane Triterpenes from Aglaia foveolata. Phytochemistry 1998, 49, 1745–1748. [Google Scholar] [CrossRef]
- Pan, L.; Acuña, U.M.; Li, J.; Jena, N.; Ninh, T.N.; Pannell, C.M.; Chai, H.; Fuchs, J.R.; De Carcache Blanco, E.J.; Soejarto, D.D.; et al. Bioactive Flavaglines and Other Constituents Isolated from Aglaia perviridis. J. Nat. Prod. 2013, 76, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Tang, G.H.; Zhang, Y.; Hao, X.J.; Zhao, Q.; He, H.P. A New Carotane Sesquiterpene from Walsura robusta. Chin. J. Nat. Med. 2013, 11, 84–86. [Google Scholar] [CrossRef]
- Huang, P.Q.; Deng, J.W.; Li, Y.; Liao, Z.B.; Zhao, E.; Tian, Y.C.; Tu, Y.D.; Li, D.L.; Jin, J.W.; Zhou, C.X.; et al. Terpenoids from the Twigs and Leaves of Aglaia elaeagnoidea and Their Chemotaxonomic Significance. Biochem. Syst. Ecol. 2022, 103, 104427. [Google Scholar] [CrossRef]
- De Paula, J.R.; Castro-Gamboa, I.; Neto, J.O.; Da Silva, M.F.D.G.F.; Fo, E.R.; Fernandes, J.B.; Vieira, P.C.; Pinheiro, A.L. Chemistry of Cedrela odorata Graft and Speculations on the Induced Resistance against Hypsipyla grandella. An. Acad. Bras. Cienc. 1998, 70, 1449–1454. [Google Scholar] [CrossRef]
- Dharmayani, N.K.T.; Yoshimura, T.; Hermawati, E.; Juliawaty, L.D.; Syah, Y.M. Antibacterial and Antifungal Two Phenolic Sesquiterpenes from Dysoxylum densiflorum. Z. Für Nat. C 2020, 75, 1–5. [Google Scholar] [CrossRef]
- Daniewski, W.M.; Anczewski, W.; Gumułka, M.; Danikiewicz, W.; Jacobsson, U.; Norin, T. Sesquiterpenoid Constituents of Entandrophragma cylindricum. Phytochemistry 1996, 43, 811–814. [Google Scholar] [CrossRef]
- Milawati, H.; Harneti, D.; Maharani, R.; Hidayat, A.T.; Azmi, M.N.; Shiono, Y.; Supratman, U.; Sciences, N.; Padjadjaran, U.; Science, E. Caryophyllene-Type Sesquiterpenoids from the Stembark of Aglalia harmsiana and Their Cytotoxic Activity Against MCF-7 Breast Cancer Cells. Molekul 2019, 14, 126–132. [Google Scholar] [CrossRef]
- Benosman, A.; Richomme, P.; Sevenet, T.; Hamid, A.; Hadi, A.; Bruneton, J. Secotirucallane Triterpenes from the Stem Bark of Aglaia leucophylla. Phytochemistry 1994, 37, 1143–1145. [Google Scholar] [CrossRef]
- Napagoda, M.; Gerstmeier, J.; Koeberle, A.; Wesely, S.; Popella, S.; Lorenz, S.; Scheubert, K.; Böcker, S.; Svatoš, A.; Werz, O. Munronia pinnata (Wall.) Theob.: Unveiling Phytochemistry and Dual Inhibition of 5-Lipoxygenase and Microsomal Prostaglandin E2 Synthase (MPGES)-1. J. Ethnopharmacol. 2014, 151, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, N.; Pezzuto, J.M.; Kinghorn, D.; Farnsworth, N.R.; Santisuk, T.; Tuchinda, P.; Udchachon, J.; Reutrakul, V. Plant Anticancer Agents, l. Cytotoxic Triterpenes from Sandoricum koetjape Stems. J. Nat. Prod. 1992, 55, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Izdihar, G.; Naini, A.A.; Harneti, D.; Maharani, R.; Nurlelasari, N.; Mayanti, T.; Safari, A.; Farabi, K.; Supratman, U.; Azmi, M.N.; et al. Sesquiterpenoids from the Stem Bark of Aglaia simplicifolia and Their Cytotoxic Activity against B16-F10 Melanoma Skin Cancer Cell. Indones. J. Chem. 2021, 21, 1560–1567. [Google Scholar] [CrossRef]
- Nishizawa, M.; Inoue, A.; Hayashi, Y.; Sastrapradja, S.; Kosela, S.; Iwashita, T. Structure of Aphanamol I and II. J. Org. Chem. 1984, 49, 3660–3662. [Google Scholar] [CrossRef]
- Vieira, I.J.C.; Figueiredo, E.R.; Freitas, V.R.; Mathias, L.; Braz-Filho, R.; Araújo, R.M. A New Sesquiterpene from Trichilia casarettii (Meliaceae). Am. J. Anal. Chem. 2010, 01, 70–72. [Google Scholar] [CrossRef]
- Pointinger, S.; Promdang, S.; Vajrodaya, S.; Pannell, C.M.; Hofer, O.; Mereiter, K.; Greger, H. Silvaglins and Related 2,3-Secodammarane Derivatives—Unusual Types of Triterpenes from Aglaia silvestris. Phytochemistry 2008, 69, 2696–2703. [Google Scholar] [CrossRef] [PubMed]
- Sofian, F.F.; Subarnas, A.; Hakozaki, M.; Uesugi, S.; Koseki, T.; Shiono, Y. Tridysoxyphenols A and B, Two New Trimeric Sesquiterpene Phenols from Dysoxylum parasiticum Leaves. Phytochem. Lett. 2022, 50, 134–140. [Google Scholar] [CrossRef]
- Russell, G.B.; Hunt, M.B.; Bowers, W.S.; Blunt, J.W. A Sesquiterpenoid Ant Repellent from Dysoxylum spectabile. Phytochemistry 1994, 35, 1455–1456. [Google Scholar] [CrossRef]
- Joycharat, N.; Plodpai, P.; Panthong, K.; Yingyongnarongkul, B.E.; Voravuthikunchai, S. Terpenoid Constituents and Antifungal Activity of Aglaia forbesii Seed against Phytopathogens. Can. J. Chem. 2010, 88, 937–944. [Google Scholar] [CrossRef]
- Yuan, H.W.; Zhao, J.P.; Liu, Y.B.; Qiu, Y.X.; Xie, Q.L.; LI, M.J.; Khan, I.A.; Wang, W. Advance in Studies on Chemical Constituents, Pharmacology and Quality Control of Aquilaria sinensis. Digit. Chin. Med. 2018, 1, 316–330. [Google Scholar] [CrossRef]
- Vieira, I.J.C.; De Aquino Azevedo, O.; De Souza, J.J.; Braz-Filho, R.; Dos Santos Gonçalves, M.; De Araújo, M.F. Hirtinone, a Novel Cycloartane-Type Triterpene and Other Compounds from Trichilia hirta L. (Meliaceae). Molecule 2013, 18, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Joycharat, N.; Greger, H.; Hofer, O.; Saifah, E. Flavaglines and Triterpenoids from the Leaves of Aglaia forbesii. Phytochemistry 2008, 69, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Nugroho, A.E.; Sugiura, R.; Momota, T.; Hirasawa, Y.; Wong, C.P.; Kaneda, T.; Hadi, A.H.A.; Morita, H. Dysosesquiflorins A and B, Sesquiterpenoids from Dysoxylum densiflorum. J. Nat. Med. 2015, 69, 411–415. [Google Scholar] [CrossRef]
- Phongmaykin, J.; Kumamoto, T.; Ishikawa, T.; Suttisri, R.; Saifah, E. A New Sesquiterpene and Other Terpenoid Constituents of Chisocheton penduliflorus. Arch. Pharm. Res. 2008, 31, 21–27. [Google Scholar] [CrossRef]
- Inada, A.; Shono, K.; Murata, H.; Inatomi, Y.; Darnaedi, D. Three Putrescine Bisamides from the Leaves of Aglaia grandis. Phytochemistry 2000, 53, 1091–1095. [Google Scholar] [CrossRef]
- Yuan, C.M.; Tang, G.H.; Wang, X.Y.; Zhang, Y.; Cao, M.M.; Li, X.H.; Li, Y.; Li, S.L.; Di, Y.T.; He, H.P.; et al. New Steroids and Sesquiterpene from Turraea pubescens. Fitoterapia 2013, 90, 119–125. [Google Scholar] [CrossRef]
- Stærk, D.; Skole, B.; Jørgensen, F.S.; Budnik, B.A.; Ekpe, P.; Jaroszewski, J.W. Isolation of a Library of Aromadendranes from Landolphia dulcis and Its Characterization Using the Volsurf Approach. J. Nat. Prod. 2004, 67, 799–805. [Google Scholar] [CrossRef]
- Fareza, M.S.; Nurlelasari; Supratman, U.; Katja, D.G.; Husna, M.H.; Awang, K. 1,1,4,7-Tetramethyldecahydro-1H-Cyclopropa[E]Azulen-7-Ol From the Stembark Chisocheton pentandrus. Molbank 2019, 2019, 4–8. [Google Scholar] [CrossRef]
- Doe, M.; Hirai, Y.; Kinoshita, T.; Shibata, K.; Haraguchi, H.; Morimoto, Y. Structure, Synthesis, and Biological Activity of 14-Methoxy-1,2-Dehydrocacalol Methyl Ether, a New Modified Furanoeremophilane Type Sesquiterpene from Trichilia cuneata. Chem. Lett. 2004, 33, 714–715. [Google Scholar] [CrossRef]
- Doe, M.; Shibue, T.; Haraguchi, H.; Morimoto, Y. Structures, Biological Activities, and Total Syntheses of 13-Hydroxy- and 13-Acetoxy-14-Nordehydrocacalohastine, Novel Modified Furanoeremophilane-Type Sesquiterpenes from Trichilia cuneata. Org. Lett. 2005, 7, 1765–1768. [Google Scholar] [CrossRef]
- Hapuarachchi, S.D.; Suresh, T.S.; Hadunnetthi, S.; PriyanthieSandun Kumari Senarath, W.T.; Ranasinghe, C. Anti-Inflammatory Potential of Aqueous Extract and Ethyl Acetate Fractions of Munronia pinnata (Wall) Theob. and the Isolated Compound, Senecrassidiol. Trends Phytochem. Res. 2022, 6, 247–258. [Google Scholar] [CrossRef]
- Ngo, N.T.N.; Lai, N.T.D.D.T.; Le, H.C.; Nguyen, L.T.T.; Trinh, B.T.D.; Nguyen, H.D.; Pham, P.D.; Dang, S.V.; Nguyen, L.H.D. Chemical Constituents of Aglaia elaeagnoidea and Aglaia odorata and Their Cytotoxicity. Nat. Prod. Res. 2022, 36, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Yamada, H.; Sastrapradja, S.; Hayashi, Y. Structure and Synthesis of Bicalamenene. Tetrahedron Lett. 1985, 26, 1535–1536. [Google Scholar] [CrossRef]
- Naini, A.A.; Mayanti, T.; Maharani, R.; Fajriah, S.; Kabayama, K.; Shimoyama, A.; Manabe, Y.; Fukase, K.; Jungsuttiwong, S.; Supratman, U. Dysoticans F-H: Three Unprecedented Dimeric Cadinanes from Dysoxylum parasiticum (Osbeck) Kosterm. Stem Bark. RSC Adv. 2023, 13, 9370–9376. [Google Scholar] [CrossRef] [PubMed]
- Xavier, S.M.; Nair, G.; Sabulal, B.; Mathew, J. Chemical Constituent and Antibacterial Activity of the Root Oil of Naregamia alata. J. Trop. Med. Plants 2011, 12, 163–165. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Wang, J.; Zhang, X.B.; Shang, X.; Gong, S.; Zhang, K. Chemical Composition of the Essential Oils of Swietenia macrophylla from China. Chem. Nat. Compd. 2016, 52, 510–511. [Google Scholar] [CrossRef]
- Dougnon, G.; Ito, M. Essential Oils from Melia azedarach L. (Meliaceae) Leaves: Chemical Variability upon Environmental Factors. J. Nat. Med. 2022, 76, 331–341. [Google Scholar] [CrossRef]
- Geetha, B.S.; Garden, T.B. Essential Oil Composition of Naregamia alata. J. Med. Aromat. Plant Sci. 2008, 32, 123–124. [Google Scholar]
- Wang, C.; Shi, J.-X.; Wu, Y. Chemical and Antimicrobial Analyses of Essential Oil of Toona sinensis from China. Asian J. Chem. 2014, 26, 2557–2560. [Google Scholar] [CrossRef]
- Kharkwal, G.C.; Pande, C.; Tewari, G.; Panwar, A.; Pande, V. Volatile Terpenoid Composition and Antimicrobial Activity of Flowers of Melia azedarach Linn, from North West Himalayas, India. J. Indian Chem. Soc. 2015, 92, 141–145. [Google Scholar] [CrossRef]
- Wu, J.G.; Peng, W.; Yi, J.; Wu, Y.B.; Chen, T.Q.; Wong, K.H.; Wu, J.Z. Chemical Composition, Antimicrobial Activity against Staphylococcus aureus and a pro-Apoptotic Effect in SGC-7901 of the Essential Oil from Toona sinensis (A. Juss.) Roem. Leaves. J. Ethnopharmacol. 2014, 154, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Hardt, I.H.; Rieck, A.; Fricke, C.; Konigt, W.A. Enantiomeric Composition of Sesquiterpene Hydrocarbons of the Essential Oil of Cedrela odorata L. Flavour Fragr. J. 1995, 10, 165–171. [Google Scholar] [CrossRef]
- Hsieh, T.J.; Tsai, Y.H.; Liao, M.C.; Du, Y.C.; Lien, P.J.; Sun, C.C.; Chang, F.R.; Wu, Y.C. Anti-Diabetic Properties of Non-Polar Toona sinensis Roem Extract Prepared by Supercritical-CO2 Fluid. Food Chem. Toxicol. 2012, 50, 779–789. [Google Scholar] [CrossRef]
- Kumar, R.; Verma, G.; Prakash, O.; Pant, A.K. Head Space GC/MS Analysis of Volatile Constituents of Trichilea connaroides Wight and Arn. Extracts and Their in Vitro Anti-Plasmodium Activity against Plasmodium falciparum Isolates. Res. J. Phytochem. 2011, 5, 41–47. [Google Scholar] [CrossRef][Green Version]
- Kavitha, K.; Bopaiah, A.; Kolar, A.B. Chemical Composition Of The Essential Oil From The Leaves Of Cipadessa baccifera (ROTH.) MIQ. Int. J. Pharm. Sci. Res. 2016, 7, 392–396. [Google Scholar] [CrossRef]
- Agarwal, G.; Pant, A.K. Volatile Constituents of Trichilia connaroides (Wight and Arn.) Roots. Asian J. Tradit. Med. 2010, 5, 199–202. [Google Scholar]
- El Souda, S.S.; Aboutabl, E.A.; Maamoun, A.A.; Hashem, F.A. Volatile Constituents and Cytotoxic Activity of Khaya grandifoliola and Khaya senegalensis Flower Extracts. J. Herbs Spices Med. Plants 2016, 22, 183–189. [Google Scholar] [CrossRef]
- Thangadurai, D.; Nagalakshmi, M.A.H.; Pulliah, T.; Ratnam, B.V.V. Essential Oils of the Leaves of Chukrasia Tabularis Collected from the Eastern Ghats of Peninsular India. J. Essent. Oil Res. 2003, 15, 25–27. [Google Scholar] [CrossRef]
- Asekun, O.T.; Ekundayo, O. Constituents of the Leaf Essential Oil of Cedrela odorata L. from Nigeria. Flavour Fragr. J. 1999, 14, 390–392. [Google Scholar] [CrossRef]
- Maia, B.H.L.N.S.; De Paula, J.R.; Sant’Ana, J.; Da Silva, M.F.D.G.F.; Fernandes, J.B.; Vieira, P.C.; Costa, M.D.S.S.; Ohashi, O.S.; Silva, J.N.M. Essential Oils of Toona and Cedrela Species (Meliaceae): Taxonomic and Ecological Implications. J. Braz. Chem. Soc. 2000, 11, 629–639. [Google Scholar] [CrossRef]
- Lago, J.H.G.; de Avila, P.; de Aquino, E.M.; Moreno, P.R.H.; Ohara T., M.T.; Limberger, R.P.; Apel, M.A.; Henriques, A.T. Volatile Oils from Leaves and Stem Barks of Cedrela fissilis (Meliaceae): Chemical Composition and Antibacterial Activities. Flavour Fragr. J. 2004, 19, 448–451. [Google Scholar] [CrossRef]
- Chen, Y.C.; Hsieh, C.L.; Huang, B.M.; Chen, Y.C. Induction of Mitochondrial-Dependent Apoptosis by Essential Oil of Toona sinensis Root through Akt, Mtor and Nf-Kb Signalling Pathways in Human Renal Cell Carcinoma Cells. J. Food Drug Anal. 2021, 29, 433–447. [Google Scholar] [CrossRef]
- Shilaluke, K.C.; Moteetee, A.N. Insecticidal Activities and GC-MS Analysis of the Selected Family Members of Meliaceae Used Traditionally as Insecticides. Plants 2022, 11, 3046. [Google Scholar] [CrossRef]
- Soares, M.G.; Batista-Pereira, L.G.; Fernandes, J.B.; Corrêa, A.G.; Da Silva, M.F.G.F.; Vieira, P.C.; Rodrigues Filho, E.; Ohashi, O.S. Electrophysiological Responses of Female and Male Hypsipyla grandella (Zeller) to Swietenia Macrophylla Essential Oils. J. Chem. Ecol. 2003, 29, 2143–2151. [Google Scholar] [CrossRef]
- Ogunwande, I.A.; Ekundayo, O.; Olawore, N.O.; Adeleke, K.A. Constituents of the Essential Oils of the Leaves and Stem Bark of Cedrela mexicana L. Grown in Nigeria. J. Essent. Oil Res. 2005, 17, 289–291. [Google Scholar] [CrossRef]
- Chen, M.; Wang, C.; Li, L.; Wang, Y.; Jiang, S. Retention of Volatile Constituents in Dried Toona sinensis by GC-MS Analysis. Int. J. Food Eng. 2010, 6, 2. [Google Scholar] [CrossRef]
- Duan, D.; Chen, L.; Yang, X.; Tu, Y.; Jiao, S. Antidepressant-like Effect of Essential Oil Isolated from Toona ciliata Roem. Var. Yunnanensis. J. Nat. Med. 2015, 69, 191–197. [Google Scholar] [CrossRef]
- Joycharat, N.; Thammavong, S.; Voravuthikunchai, S.P.; Plodpai, P.; Mitsuwan, W.; Limsuwan, S.; Subhadhirasakul, S. Chemical Constituents and Antimicrobial Properties of the Essential Oil and Ethanol Extract from the Stem of Aglaia odorata Lour. Nat. Prod. Res. 2014, 28, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Aromdee, C.; Anorach, R.; Sriubolmas, N. Essential Oil of the Flower of Azadirachta indica (Meliaceae). Acta Hortic. 2005, 679, 11–14. [Google Scholar] [CrossRef]
- Siddiqui, B.S.; Ali, S.T.; Rajput, M.T.; Gulzar, T.; Rasheed, M.; Mehmood, R. GC-Based Analysis of Insecticidal Constituents of the Flowers of Azadirachta indica A. Juss. Nat. Prod. Res. 2009, 23, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.G.; Fernandes, J.B.; Lago, J.H.G. Interspecific Variation In The Composition Of Volatile Oils From The Leaves Of Swietenia macrophylla King (Meliaceae). Quim. Nova 2010, 33, 1141–1144. [Google Scholar] [CrossRef]
- Magalhães, L.A.M.I.; Da Paz Lima, M.; Marques, M.O.M.; Facanali, R.; Da Silva Pinto, A.C.; Tadei, W.P. Chemical Composition and Larvicidal Activity against Aedes Aegypti Larvae of Essential Oils from Four Guarea Species. Molecules 2010, 15, 5734–5741. [Google Scholar] [CrossRef]
- Pandini, J.A.; Pinto, F.G.S.; Scur, M.C.; Santana, C.B.; Costa, W.F.; Temponi, L.G. Composição Química, Potencial Antimicrobiano e Antioxidante Do Óleo Essencial de Guarea kunthiana A. Juss. Brazil. J. Biol. 2018, 78, 53–60. [Google Scholar] [CrossRef]
- Oliveira, E.A.; Martins, E.G.A.; Soares, M.G.; Chagas-Paula, D.A.; Passero, L.F.D.; Sartorelli, P.; Baldim, J.L.; Lago, J.H.G. A Comparative Study on Chemical Composition, Antileishmanial and Cytotoxic Activities of the Essential Oils from Leaves of Guarea macrophylla (Meliaceae) from Two Different Regions of São Paulo State, Brazil, Using Multivariate Statistical Analysis. J. Braz. Chem. Soc. 2019, 30, 1395–1405. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Roque, N.F. Analysis of the Essential Oil from Leaves of Three Different Specimens of Guarea guidonia (L.) Sleumer (Meliaceae). J. Essent. Oil Res. 2015, 17, 271–273. [Google Scholar] [CrossRef]
- Menut, C.; Lamaty, G.; Bessière, J.M.; Seuleiman, A.M.; Fendero, P.; Maidou, E.; Dénamganai, J. Aromatic Plants of Tropical Central Africa. XXI. Chemical Composition of Bark Essential Oil of Guarea cedrata (A. Chev.) Pellegr. from Central African Republic. J. Essent. Oil Res. 1995, 7, 207–209. [Google Scholar] [CrossRef]
- Ribeiro, W.H.F.; Arriaga, Â.M.C.; Andrade-Neto, M.; Vasconcelos, J.N.; Santiago, G.M.P.; Nascimento, R.F. Composition of the Essential Oil of Guarea macrophylla Vahl. Ssp. Tuberculata (Meliaceae) from Northeast of Brazil. J. Essent. Oil Res. 2006, 18, 95–96. [Google Scholar] [CrossRef]
- Nogueira, T.S.R.; de Passos, M.S.; Nascimento, L.P.S.; de Arantes, M.B.S.; Monteiro, N.O.; da Boeno, S.I.S.; de Carvalho Junior, A.; de Azevedo, O.A.; da Terra, W.S.; Vieira, M.G.C.; et al. Chemical Compounds and Biologic Activities: A Review of Cedrela Genus. Molecules 2020, 25, 5401. [Google Scholar] [CrossRef]
- Nuñez, C.V.; Lago, J.H.G.; Roque, N.F. Variation on the Chemical Composition of the Oil from Damaged Branches of Guarea guidonia (L.) Sleumer (Meliaceae). J. Essent. Oil Res. 2005, 17, 626–627. [Google Scholar] [CrossRef]
- Wang, G.W.; Jin, H.Z.; Zhang, W.D. Constituents from Aphanamixis Species and Their Biological Activities. Phytochem. Rev. 2013, 12, 915–942. [Google Scholar] [CrossRef]
- Bandi, A.K.R.; Dong-Ung, L. Secondary Metabolites of Plants from the Genus Cipadessa: Chemistry and Biological Activity. Chem. Biodivers. 2012, 9, 1403–1421. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Kernan, M.R.; Jolad, S.D.; Stoddart, C.A.; Bogan, M.; Cooper, R. Dysoxylins A-D, Tetranortriterpenoids with Potent Anti-RSV Activity from Dysoxylum gaudichaudianum. J. Nat. Prod. 2007, 70, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Safriansyah, W.; Sinaga, S.E.; Supratman, U.; Harneti, D. Phytochemistry and Biological Activities of Guarea Genus (Meliaceae). Molecules 2022, 27, 1–34. [Google Scholar] [CrossRef]
- Negi, J.S.; Bisht, V.K.; Bhandari, A.K.; Bharti, M.K.; Sundriyal, R.C. Chemical and Pharmacological Aspects of Toona (Meliaceae). Res. J. Phytochem. 2011, 5, 14–21. [Google Scholar] [CrossRef]
- Curcino Vieira, I.J.; da Silva Terra, W.; dos Santos Gonçalves, M.; Braz-Filho, R. Secondary Metabolites of the Genus Trichilia: Contribution to the Chemistry of Meliaceae Family. Am. J. Anal. Chem. 2014, 05, 91–121. [Google Scholar] [CrossRef]
- Oyedeji-Amusa, M.O.; Sadgrove, N.J.; Van Wyk, B.E. The Ethnobotany and Chemistry of South African Meliaceae: A Review. Plants 2021, 10, 1796. [Google Scholar] [CrossRef]
- Voravuthikunchai, S.P.; Kanchanapoom, T.; Sawangjaroen, N.; Hutadilok-Towatana, N. Antioxidant, Antibacterial and Antigiardial Activities of Walsura robusta Roxb. Nat. Prod. Res. 2010, 24, 813–824. [Google Scholar] [CrossRef]
- Kristianslund, R.; Aursnes, M.; Tungen, J.E.; Görbitz, C.H.; Hansen, T.V. Synthesis, Biological Investigation, and Structural Revision of Sielboldianin A. J. Nat. Prod. 2018, 81, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Ellithey, M.S.; Lall, N.; Hussein, A.A.; Meyer, D. Cytotoxic, Cytostatic and HIV-1 PR Inhibitory Activities of the Soft Coral Litophyton arboreum. Mar. Drugs 2013, 11, 4917–4936. [Google Scholar] [CrossRef]
- Shi, D.; Song, X.; Guo, Y.; Xu, J.; Liu, Y.; Zhang, J.; Cui, C.A.; Jin, D.Q. Alismol, a Sesquiterpenoid Isolated from Vladimiria Souliei, Suppresses Proinflammatory Mediators in Lipopolysaccharide-Stimulated Microglia. J. Mol. Neurosci. 2017, 62, 106–113. [Google Scholar] [CrossRef]
- Zhang, H.J.; Tan, G.T.; Santarsiero, B.D.; Mesecar, A.D.; Van Hung, N.; Cuong, N.M.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.S. New Sesquiterpenes from Litsea verticillata. J. Nat. Prod. 2003, 66, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Pfoh, R.; Rühl, S.; Qin, S.; Laatsch, H. T-Muurolol Sesquiterpenes from the Marine Streptomyces Sp. M491 and Revision of the Configuration of Previously Reported Amorphanes. J. Nat. Prod. 2009, 72, 99–101. [Google Scholar] [CrossRef]
- Akiel, M.A.; Alshehri, O.Y.; Aljihani, S.A.; Almuaysib, A.; Bader, A.; Al-Asmari, A.I.; Alamri, H.S.; Alrfaei, B.M.; Halwani, M.A. Viridiflorol Induces Anti-Neoplastic Effects on Breast, Lung, and Brain Cancer Cells through Apoptosis. Saudi J. Biol. Sci. 2022, 29, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Zhou, L.; Huang, Y.; Wang, Y.; Hao, X.; Wang, J. Antimicrobial Activity of Globulol Isolated from the Fruits of Eucalyptus globulus Labill. Nat. Prod. Res. 2008, 22, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Ayyad, S.E.N.; Alarif, W.M.; Al-Footy, K.O.; Selim, E.A.; Ghandourah, M.A.; Aly, M.M.; Alorfi, H.S. Isolation, Antimicrobial and Antitumor Activities of a New Polyhydroxysteroid and a New Diterpenoid from the Soft Coral Xenia Umbellata. Z. Für Naturforschung C. 2017, 72, 27–34. [Google Scholar] [CrossRef]
- Tseng, Y.J.; Shen, K.P.; Lin, H.L.; Huang, C.Y.; Dai, C.F.; Sheu, J.H. Lochmolins A-G, New Sesquiterpenoids from the Soft Coral Sinularia Lochmodes. Mar. Drugs 2012, 10, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Kurniasih, N.; Supriadin, A.; Fajar, M.; Abdulah, R.; Harneti, D.; Supratman, U.; Taib, M.N.A.B.M. Cytotoxic Sesquterpenoid Compound from the Stembark of Aglaia simplicifolia (Meliaceae). J. Phys. Conf. Ser. 2019, 1402, 3–7. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-Inflammatory Effects of Compounds Alpha-Humulene and (−)-Trans-Caryophyllene Isolated from the Essential Oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, G.; Deng, Q.; Xu, Z.; Cen, J.; Xu, J. Vomifoliol Isolated from Mangrove Plant Ceriops Tagal Inhibits the NFAT Signaling Pathway with CN as the Target Enzyme in Vitro. Bioorganic Med. Chem. Lett. 2021, 48, 128235. [Google Scholar] [CrossRef] [PubMed]
- Sadiqi, S.; Hamza, M.; Ali, F.; Alam, S.; Shakeela, Q.; Ahmed, S.; Ayaz, A.; Ali, S.; Saqib, S.; Ullah, F.; et al. Molecular Characterization of Bacterial Isolates from Soil Samples and Evaluation of Their Antibacterial Potential against MDRS. Molecules 2022, 27, 6281. [Google Scholar] [CrossRef] [PubMed]
- Petrera, E. Antiviral and Immunomodulatory Properties of Meliaceae Family. J. Biol. Act. Prod. Nat. 2015, 5, 241–254. [Google Scholar] [CrossRef]
- Tajuddeen, N.; Van Heerden, F.R. Antiplasmodial Natural Products: An Update. Malar. J. 2019, 18, 1–62. [Google Scholar] [CrossRef]










| Acyclic Sesquiterpenoids | ||||
| Compounds | Species | Parts of Plant | Biological Activity | Ref. |
| 1 | A. polystachya (Wall.) R.Parker | Seeds | Anti-inflammatory activity against RAW 264.7 macrophage IC50 14.2 ± 0.9 μM | [37] |
| 2 | A. grandifolia Blume | Leaves and stems | Cytotoxic activity against HL-60 (IC50 > 50 μM), SMMC-7721 (IC50 > 50 μM), A-549 (IC50 > 50 μM), MCF-7 (IC50 > 50 μM), SW480 (IC50 > 50 μM) Antibacterial activity against Sa, Pa: inactive | [38] |
| Monocyclic Sesquiterpenoids | ||||
| Compounds | Species | Parts of Plant | Biological Activity | Ref. |
| 7 | C. boiviana Baill. | Stembarks | Cytotoxic activity against K562 (IC50 > 50 μM) Antioxidant assay CLPAA (IC50 > 100 μM), CAA (IC50 > 100 μM) | [41,164] |
| 11 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Anti-inflammatory activity BV-2 microglial cell (22.58% at 20 μM) Antidiabetic assay PTP1B (IC50 ≥ 50 μM) | [44] |
| 12 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Anti-inflammatory activity BV-2 microglial cell (10.58% at 20 μM) Antidiabetic assay PTP1B (IC50 16.05 ± 1.09 μM) | [44] |
| 13 | G. guidonia (L.) Sleumer G. macrophylla G.King T. lepidota Mart | Stembarks Stembarks Stems | Anti-inflammatory assay reducing the edema formation induced by carrageenan (300 μg/paw), an effect observed at 30, 60, 120, and 240 min | [46] [45] [47] |
| 17 | T. emetica (Forssk.) Vahl | Roots | Cytotoxic activity against S180 (IC50 7 ± 3 mM); MCF-7 (IC50 78 ±15 mM) | [5] |
| Antiplasmodial activity against Plasmodium falciparum (IC50 76 ± 10 mM) | ||||
| 19 | T. monadelpha (Thonn.) J. De Wild | Leaves and root barks | Cytotoxic activity against L5178Y (IC50 > 50 μM) | [50] |
| 20 | T. monadelpha (Thonn.) J. De Wild | Leaves and root barks | Cytotoxic activity against L5178Y (IC50 > 50 μM) | [50] |
| 25 | L. domesticum Correa | Fruit Peel | Antibacterial activity against Sa (inactive); Ec (inactive); Bs (inactive) | [52] |
| 26 | D. parasiticumi (Osbek). Kosterm | Leaves | Cytotoxic activity HL-60 (IC50 > 50 μM) | [53] |
| 27 | A. grandifolia Blume | Stems | CN activity with inhibition rate of 58.06%, strong immunosuppressive effect on Con A-induced anti-inflammatory activity murine splenocytes and PMA/IO-induced jurkat cells significantly reduced NFAT1 protein expression and downstream gene. Immunosuppressant activity reduced IL-2 expression in the CN/NFAT signaling pathway | [38,54] |
| 28 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B IC50 ≥ 50 μM (inactive) | [44] |
| 31 | L. domesticum Correa | Fruit peel | Cytotoxic activity aginst T-47D (IC50 39.18 ± 1.54 µg/mL), WiDr (IC50 > 50 μg/mL), Hep-G2 (IC50 > 50 μg/mL) | [56] |
| Bicyclic Sesquiterpenoids | ||||
| Compounds | Species | Part of Plant | Biological Activity | Ref. |
| 36 | A. minahassae Koord. | Stembarks | Cytotoxic activity against MCF-7 (IC50 121.65 µg/mL) | [59] |
| C. cinerascens (Pellegr.) Hand.-Mazz. | Branches | [60] | ||
| C. baccifera (Roth) Miq | Stems | [61] | ||
| G. guidonia (L.) Sleumer | Seeds | [62] | ||
| A. tsangii (Merr.) X.M.Chen | Twigs and leaves | [63] | ||
| L. domesticum Correa | Stembarks | [64] | ||
| D. densiflorum (Blume) Miq | Twigs and leaves | [65] | ||
| D. parasiticum Correa | Stembarks | [3] | ||
| T. africanus (Wele. Ex C.DC.) Pellegr. | Stembarks | [66] | ||
| A. grandis (Korth. Ex Miq) Pierre | Stembarks | [6] | ||
| 44 | G. guidonia (L.) Sleumer | Seeds | Cytotoxic activity against MCF-7 (IC50 17.97 µg/mL) | [62] |
| L. domesticum Correa | Stembarks | [64] | ||
| 45 | A. elaeagnoidea (A.Juss.) Benth. | Stembarks | Cytotoxic activity against HeLa (IC50 3544 µg/mL); DU145 (IC50 971.69 µg/mL) | [70] |
| G. guidonia (L.) Sleumer | Seeds | [62] | ||
| 46 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 47 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 48 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | PTP1B inhibitory (IC50 > 50 µM) | [44] |
| A. elaeagnoidea (A.Juss.) Benth. | Stembarks | Cytotoxic activity against HeLa (IC50 9010.62 µg/mL); DU145 (IC50 16,883.7 µg/mL) | [70] | |
| 50 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B inhibitory (IC50 > 50 µM) | [44] |
| 51 | A. foveolata Pannel | Stembarks | Cytotoxic activity against HT-29 (ED50 >10 µM) | [73] |
| Anti-inflammatory activity NF-kB p65 (RelA) IC50 > 20 µM | ||||
| 52 | D. excelsum Blume | Leaves | Cytotoxic activity against KB (IC50 49.4 mg/mL); MCF-7 (IC50 37.8 mg/mL) | [42] |
| 53 | D. excelsum Blume | Leaves | Cytotoxic activity against HL-60 (inactive) | [42] |
| 58 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 27.39 μM) | [77] |
| 68 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 69 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 70 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 71 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 72 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 73 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 74 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 75 | D. densiflorum (Blume) Miq | Fruits | Antiinflammatory activity against RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| 76 | A. rohituka (Roxb.) Wight and Arn. | Stembarks | Antiviral activity HIV-inhibitory (>50 mg/mL) | [82] |
| 77 | A. rohituka (Roxb.) Wight and Arn. | Stembarks | Antiviral activity HIV-inhibitory (>50 mg/mL) | [82] |
| 80 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (SGC-7901 IC50 38.8 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 81 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (SGC-7901 IC50 40.0 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 82 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 83 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 > 100 ± 0.37 µM); HeLa (IC50 >100 ± 0.36 µM) | [3] | |
| 84 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 85 | A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 > 50 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] |
| 87 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 208.06 μM) | [77] |
| A. odorata Lour. | Twigs | Cytotoxic activity against SGC-7901 (IC50 38.0 µM); K-562 (IC50 > 50 µM); BEL-7402 (IC50 > 50 µM) | [71] | |
| A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B (IC50 ≥ 50 μM) | [44] | |
| D. excelsum Blume | Leaves | [42] | ||
| C. baccifera (Roth) Miq | Stems | [61] | ||
| C. lasiocarpus (Miq) Valeton | Stembark | [72] | ||
| 88 | G. kunthiana A.Juss | Leaves | Cytotoxic activity against HeLa (IC50 > 100 μM); Vero (IC50 > 100 μM); U937 (IC50 > 100 μM) | [85,165] |
| T. sinensis (A.Juss.) M. Roem. | Pericarps | [68] | ||
| T. sinensis (A.Juss.) M. Roem. | Pericarps | [69] | ||
| C. cumingianus | Twigs | [76] | ||
| D. densiflorum (Blume) Miq | Twigs and Leaves | [65] | ||
| 90 | G. kunthiana A.Juss | Leaves | Western blot and RT-PCR analyses of anti-inflammatory activity showed that alismol markedly inhibited iNOS and COX-2 expression at both mRNA and protein levels as well as NO and PGE2 production. At 100 μM, alismol almost completely blocked LPS-induced iNOS, COX-2, PGE2, and NO induction. | [85,166] |
| 91 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 183.69 µg/mL) | [77] |
| 92 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 39.61 ± 0.29 µM); HeLa (IC50 26.06 ± 0.21 µM) | [3] |
| G. macrophylla M. Vahl | Stembarks | [90] | ||
| 94 | A. foveolata Pannel | Barks | Cytotoxic activity against HT-29 (ED50 > 20 μM) | [89] |
| A. perviridis Hiern | Fruits, leaves, twigs, and roots | Antidiabetic activity PTP1B inhibitory (IC50 > 50 µM) | [92] | |
| A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antibacterial activity against Sa (inactive); MRSA 92 (inactive); MRSA 98 (inactive); MRSA 111 (inactive) | [91] | |
| W. robusta Roxb. | Leaves | Anti-inflammatory activity Nf-Kb (ED50 0.005 μM) | [93] | |
| 95 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B inhibitory (IC50 > 50 µM) Anti-inflammatory activity BV-2 microglial cell (53.75% at 20 μM) | [44] |
| 96 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B inhibitory (IC50 > 50 µM) | [44] |
| 97 | A. elaeagnoidea (A.Juss.) Benth. | Twigs and Leaves | Cytotoxic activity against HL-60 (inactive); A549 (inactive) | [92] |
| D. excelsum Blume | Leaves | [42] | ||
| 98 | A. elaeagnoidea (A.Juss.) Benth. | Twigs and Leaves | Cytotoxic activity against HL-60 (inactive); A549 (inactive) | [92] |
| D. excelsum Blume | Leaves | [42] | ||
| 101 | D. excelsum Blume | Leaves | Cytotoxic activity against HL-60 (inactive); A549 (inactive) | [42] |
| 102 | W. robusta Roxb. | Leaves | Antibacterial activity Sa (inactive); MRSA 92 (inactive); MRSA 98 (inactive); MRSA 111 (inactive) | [91] |
| 103 | D. densiflorum (Blume) Miq | Twigs and leaves | Cytotoxic activity against HL-60 (inactive); A549 (inactive) | [86] |
| D. excelsum Blume | Leaves | [42] | ||
| 113 | A. tsangii (Merr.) X.M.Chen | Twigs and leaves | Antiviral activity anti-HIV (HOG.R5 IC50 10 μg/mL) | [95] |
| 114 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 33.46 μM) | [77] |
| 115 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 38.79 ± 0.22 µM); HeLa (IC50 39.31 ± 0.14 µM) | [3] |
| 116 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 45.14 ± 0.12 µM); HeLa (IC50 41.82 ± 0.38 µM) | [3] |
| 117 | A. grandis (Korth. Ex Miq) Pierre | Stembarks | Cytotoxic activity against HeLa (IC50 6086.3 µM); B16-F10 (IC50 3375.63 µM) | [6] |
| 121 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 18.25 ± 1.52 µM) | [53] |
| D. densiflorum | Seeds | Antimicrobial activity against Bs (MIC 28 µM); Sa (MIC 57 µM); Ec (MIC 57 µM); Pa (MIC 114 µM); St (MIC 114 µM); Sd (MIC 114 µM); Vc (MIC 57 µM) | [94] | |
| 122 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 119.85 ± 10.03 µM) | [103] |
| 123 | D. densiflorum (Blume) Miq | Seeds | Antimicrobial activity against Bs (MIC 28 µM); Sa (MIC 57 µM); Ec (MIC 57 µM); Pa (MIC 114 µM); St (MIC 114 µM); Sd (MIC 114 µM); Vc (MIC 57 µM) | [94] |
| 124 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 80.66 ± 0.23 µM); HeLa (IC50 82.18 ± 0.35 µM) | [3] |
| 125 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 > 100 ± 0.33 µM); HeLa (IC50 > 100 ± 0.44 µM) | [3] |
| D. shciffneri F. Muell. | Wood | [8] | ||
| 129 | A. harmsiana Perkins | Stembarks | Cytotoxic activity against MCF-7 (IC50 0.62 μM) | [95] |
| A. leucophylla King | Stembarks | [100] | ||
| G. macrophyla M. Vahl | Stembarks | [90] | ||
| M. pinata Harms | Whole plant | [101] | ||
| 133 | A. simplicifolia (Bedd.) Harms | Stembarks | Cytotoxic activity against B16-F10 (IC50 483.2 μg/mL) | [99] |
| 134 | A. grandifolia Blume | Fruit peels | Anti-HIV (HOG.R5 IC50 10 μg/mL) | [100,167] |
| C. baccifera (Roth) Miq | Stems | [61] | ||
| D. excelsum | Leaves | [42] | ||
| 135 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B inhibitory (IC50 >50 µM) | [44] |
| A. grandifolia Blume | Fruit peel | [104] | ||
| D. densiflorum (Blume) Miq | Twigs and leaves | [88] | ||
| D. excelsum Blume | Leaves | [42] | ||
| T. africanus (Wele. Ex C.DC.) Pellegr. | Stembarks | [66] | ||
| 141 | L. domesticum Correa | Stembarks | Cytotoxic activity against MCF-7 (IC50 201.57 µg/mL) | [64] |
| 144 | D. excelsum Blume | Leaves | Cytotoxic activity against HL-60 (inactive); A549 (inactive) | [42] |
| 148 | C. odorata L. | Stems | Cytotoxic activiy against 1218L, T24, 498NL, SF268, HCT116, HT29, 251L, 536L, 1121L, 289L, 526L, 529L, 629L, H460, 401NL, MCF7, DA231, 276L, 394NL, 462NL, 514L, 520L, 1619L, 899L, OVCAR3, 1657L, PANC1, 22RV1, DU145, LNCAP, PC3M, 1752L, 1781L, 393NL, 486L, 944L, and 1138L; Not active (IC50 > 10 μg/mL) | [92,168] |
| C. toona Roxb | Timber | [31] | ||
| 149 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 >150 µM) | [103] |
| C. baccifera (Roth) Miq | Stembarks | [41] | ||
| 150 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Antidiabetic activity PTP1B inhibitory (IC50 >50 µM) | [44] |
| 152 | D. densiflorum (Blume) Miq | Fruits | Antiinflammation activity RAW 264.7 macrophage inactive; IL-1β inactive | [82] |
| Tricyclic Sesquiterpenoids | ||||
| Compounds | Species | Part of Plant | Biological Activity | Ref. |
| 153 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 12.17 μM) | [77] |
| A. minahassae Koord. | Stembarks | [59] | ||
| A. foveolata Pannel | Barks | [88] | ||
| A. forbesii King | Seeds | [105] | ||
| A. forbesii King | Leaves | [108] | ||
| G. macrophylla M. Vahl | Wood | [87] | ||
| G. macrophylla M. Vahl | Leaves | [45] | ||
| S. koetjape Merr. | Stems | [98] | ||
| T. hirta L. | Fruits | [107] | ||
| T. lepidota Mart | Stems | [47] | ||
| 157 | D. densiflorum (Blume) Miq | Barks | Cytotoxic activity against HL-60 (IC50 3.1 µM); MCF-7 (IC50 32.5 µM); A549 (IC50 30.4 µM) | [109] |
| 158 | D. densiflorum (Blume) Miq | Barks | Cytotoxic activity against HL-60 (IC50 14.3 µM); MCF-7 (IC50 39.7 µM); A549 (IC50 31.3 µM) | [109] |
| 159 | A. silvestris (M. Roem.) Merr. | Leaves | Cytotoxic activity against Daoy (IC50 0.1 µM); MCF-7 (IC50 10 µM); A549 (IC50 30 µM | [102,169] |
| D. densiflorum (Blume) Miq | Barks | Cytotoxic activity against Daoy (55.8–72.1%), MCF-7 (36.2–72.7%), and A459 (35–98.9%) | [109,169] | |
| 160 | D. densiflorum (Blume) Miq | Barks | Cytotoxic activity against HL-60 (IC50 >50 µM); MCF-7 (IC50 >50 µM); A549 (IC50 32,5 µM) | [109] |
| Antimicrobial activity against As (IC50 47.1 µg/mL); Fo (IC50 114.3 µg/mL); Fg (IC50 53.4 µg/mL); Rs (IC50 56.9 µg/mL); Vp (IC50 21.8 µg/mL); Xv (IC50 158.0 µg/mL); Bs (IC50 737.2 µg/mL) | [109,170] | |||
| 164 | C. penduliflorus Planch. | Wood and leaves | Antibacterial activity against Mycobacterium tuberculosis H37Ra (MIC 50 µg/mL) | [110] |
| D. densiflorum (Blume) Miq | Twigs and leaves | Cytotoxic activity against NCI-H187 (IC50 > 50 µM); BC (IC50 > 50 µM); KB (IC50 > 50 µM) | [88] | |
| 165 | A. simplicifolia (Bedd.) Harms | Stembarks | Cytotoxic activity against B16-F10 (IC50 44.8 µg/mL) | [99] |
| A. grandis (Korth. Ex Miq) Pierre | Leaves | Cytotoxic activity against HeLa (IC50 10.83 ± 0.17 µM); MCF-7 (IC50 10.37 ± 0.11 µM) | [115] | |
| D. parasiticum (Osbek). Kosterm | Stembarks | [3] | ||
| A. grandis (Korth. Ex Miq) Pierre | Stembarks | [6] | ||
| 166 | A. lawii (Wight) C.J.Saldanha | Twigs and leaves | Cytotoxic activity against MCF-7 (IC50 231.99 ± 5.40 µM) | [44] |
| C. penduliflorus Planch. | Wood and leaves | PTP1B (IC50 ≥ 50 μM) | [111] | |
| C. lasiocarpus (Miq) Valeton | Stembark | [72] | ||
| 167 | C. lasiocarpus (Miq) Valeton | Stembark | Cytotoxic activity against MCF-7 (IC50 258.70 ± 2.49 µM) | [72] |
| C. penduliflorus Planch. | Wood and leaves | Antibacterial activity against Mycobacterium tuberculosis H37Ra (MIC 50 µg/mL) | [110] | |
| 168 | C. penduliflorus Planch. | Wood and leaves | Mycobacterium tuberculosis H37Ra (MIC 50 µg/mL) | [110] |
| Cytotoxic activity against NCI-H187 (IC50 > 50 µM); BC (IC50 > 50 µM); KB (IC50 > 50 µM) | ||||
| 172 | T. pubescens Hell. | Twigs | Cytotoxic activity against HL-60 (IC50 > 20 µM) | [112] |
| Antiinflammatory activity inhibiton of NO production in RAW 264.7 (IC50 > 20 µM) | ||||
| 174 | G. macrophylla M. Vahl | Leaves | Cytotoxic activity Lymphoma (LD50 ≥ 3.60 mM) | [45,171] |
| Toxicity against Artemia Salina (LD50 ≥ 3.60 mM) | ||||
| 175 | T. maynasiana C.DC. | Leaves | Cytotoxic activity against HeLa (inactive); SK-Hep1 (inactive); B-16 (inactive) | [55,172] |
| Did not reduce the accumulation of iNOS protein induced by LPS in concentrations 1, 10, 100 µM | ||||
| 179 | T. cuneata Radlk. | Stembarks | Antioxidant activity Fe(III)–ADP/NADH (IC50 76.8 mM) | [116] |
| 183 | T. cuneata Radlk. | Stembarks | Antioxidant activity Fe(III)–ADP/NADH (inactive) | [116] |
| 186 | T. cuneata Radlk. | Stembarks | NADH-dependent mitochondrial (IC50 of 16.4 µM); NADPH-dependent microsomal lipid peroxidations (IC50 of 41.6 µM) | [117] |
| 187 | T. cuneata Radlk. | Stembarks | NADH-dependent mitochondrial (IC50 of 59.7 µM); NADPH-dependent microsomal lipid peroxidations (IC50 of 54.3 µM) | [117] |
| 188 | T. cuneata Radlk. | Stembarks | NADH-dependent mitochondrial (IC50 of 71.7 µM); NADPH-dependent microsomal lipid peroxidations (IC50 of 74.7 µM) | [117] |
| 194 | A. simplicifolia (Bedd.) Harms | Stembarks | Cytotoxic activity against HeLa (IC50 2.18 µM) | [118] |
| A. harmsiana Perkins | Stembarks | [99] | ||
| 195 | A. elaeagnoidea (A.Juss.) Benth. | Bark | Cytotoxic activity against HepG2 (IC50 > 30 µM) | [118] |
| Others sesquiterpenoids | ||||
| Compounds | Species | Part of Plant | Biological Activity | Ref. |
| 201 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 41.54 ± 0.34 µM); HeLa (IC50 22.15 ± 0.22 µM) | [3] |
| 202 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 > 100 ± 0.27 µM); HeLa (IC50 13.00 ± 0.13 µM) | [3] |
| 203 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 39.04 ± 3.12 µM) | [53] |
| 204 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 >50 µM) | [53] |
| 205 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 >50 µM) | [53] |
| 206 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 40.56 ± 0.24 µM); HeLa (IC50 39.32 ± 0.25 µM) | [3] |
| 207 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 >150 µM) | [103] |
| 208 | D. parasiticum (Osbek). Kosterm | Leaves | Cytotoxic activity against HL-60 (IC50 >150 µM) | [103] |
| 209 | D. parasiticum(Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 12.07 ± 0.17 µM); HeLa (IC50 9.29 ± 0.33 µM) | [120] |
| 210 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 >100 ± 0.27 µM); HeLa (IC50 39.72 ± 0.18 µM) | [120] |
| 211 | D. parasiticum (Osbek). Kosterm | Stembarks | Cytotoxic activity against MCF-7 (IC50 31.59 ± 0.34 µM); HeLa (IC50 27.93 ± 0.25 µM) | [120] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riyadi, S.A.; Naini, A.A.; Supratman, U. Sesquiterpenoids from Meliaceae Family and Their Biological Activities. Molecules 2023, 28, 4874. https://doi.org/10.3390/molecules28124874
Riyadi SA, Naini AA, Supratman U. Sesquiterpenoids from Meliaceae Family and Their Biological Activities. Molecules. 2023; 28(12):4874. https://doi.org/10.3390/molecules28124874
Chicago/Turabian StyleRiyadi, Sandra Amalia, Al Arofatus Naini, and Unang Supratman. 2023. "Sesquiterpenoids from Meliaceae Family and Their Biological Activities" Molecules 28, no. 12: 4874. https://doi.org/10.3390/molecules28124874
APA StyleRiyadi, S. A., Naini, A. A., & Supratman, U. (2023). Sesquiterpenoids from Meliaceae Family and Their Biological Activities. Molecules, 28(12), 4874. https://doi.org/10.3390/molecules28124874