
A Marine Natural Product, Harzianopyridone, as an Anti-ZIKV Agent by Targeting RNA-Dependent RNA Polymerase


 and
and
Abstract
1. Introduction
2. Results
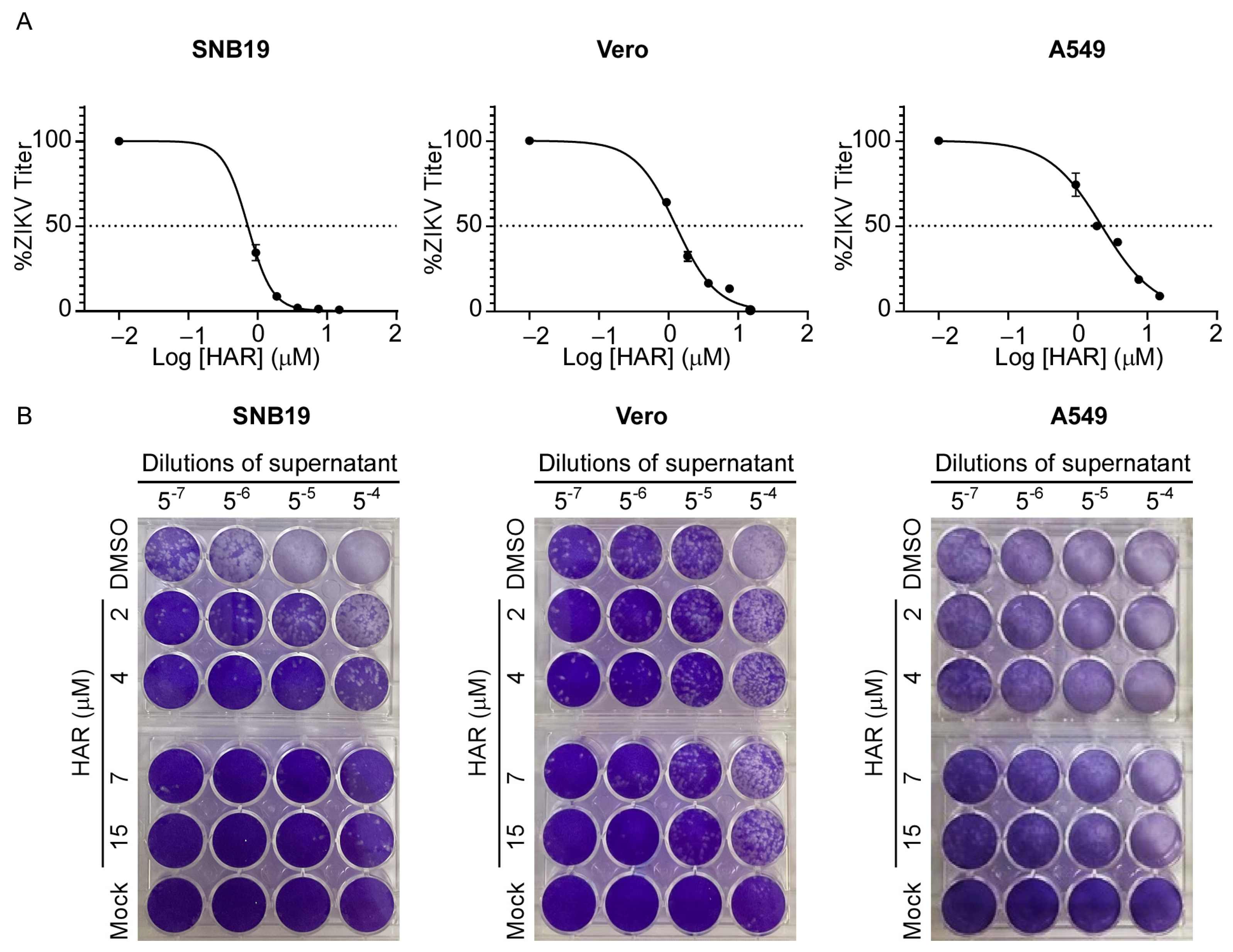
2.1. HAR Exerts Antiviral Potential without Obvious Cytotoxicity
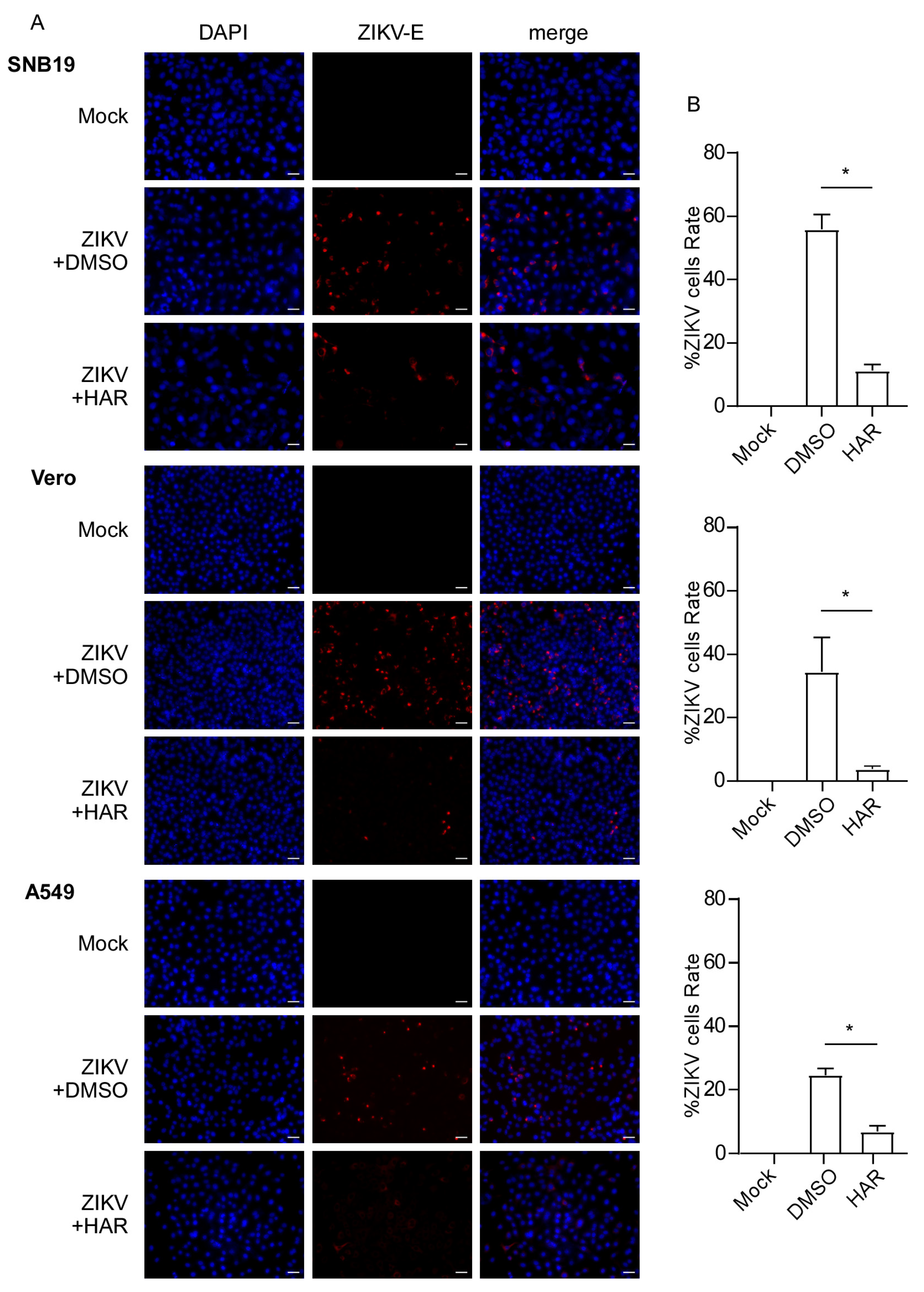
2.2. HAR Inhibits the Expression of ZIKV Proteins
2.3. HAR Protects Cells against ZIKV Infection
2.4. HAR Directly Targets the ZIKV RdRp
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Virus
4.2. Compound Preparation
4.3. Cell Viability Assay
4.4. RNA Extraction, Reverse Transcription, and Real-Time PCR
4.5. Plaque-Forming Assay
4.6. Western Blotting Analysis
4.7. Immunofluorescence
4.8. ZIKV NS5 RdRp Activity Assay
4.9. BIAcore Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, A.; Zhang, X.; Dejnirattisai, W.; Dai, X.; Gong, D.; Wongwiwat, W.; Duquerroy, S.; Rouvinski, A.; Vaney, M.C.; Guardado-Calvo, P.; et al. The epitope arrangement on flavivirus particles contributes to Mab C10’s extraordinary neutralization breadth across Zika and dengue viruses. Cell 2021, 184, 6052–6066.e18. [Google Scholar] [CrossRef]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Ko, A.I.; Baud, D. Zika Virus Infection—After the Pandemic. N. Engl. J. Med. 2019, 381, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013-15: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef]
- Triunfol, M. Microcephaly in Brazil: Confidence builds in Zika connection. Lancet Infect. Dis. 2016, 16, 527–528. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Zika Virus. Available online: https://www.who.int/news-room/fact-sheets/detail/zika-virus (accessed on 13 February 2024).
- Zika virus: A new global threat for 2016. Lancet 2016, 387, 96. [CrossRef] [PubMed]
- Lucey, D.R. Time for global action on Zika virus epidemic. BMJ 2016, 352, i781. [Google Scholar] [CrossRef][Green Version]
- Lessler, J.; Chaisson, L.H.; Kucirka, L.M.; Bi, Q.; Grantz, K.; Salje, H.; Carcelen, A.C.; Ott, C.T.; Sheffield, J.S.; Ferguson, N.M.; et al. Assessing the global threat from Zika virus. Science 2016, 353, aaf8160. [Google Scholar] [CrossRef]
- Driggers, R.W.; Ho, C.Y.; Korhonen, E.M.; Kuivanen, S.; Jääskeläinen, A.J.; Smura, T.; Rosenberg, A.; Hill, D.A.; DeBiasi, R.L.; Vezina, G.; et al. Zika Virus Infection with Prolonged Maternal Viremia and Fetal Brain Abnormalities. N. Engl. J. Med. 2016, 374, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Brasil, P.; Sequeira, P.C.; Freitas, A.D.; Zogbi, H.E.; Calvet, G.A.; de Souza, R.V.; Siqueira, A.M.; de Mendonca, M.C.; Nogueira, R.M.; de Filippis, A.M.; et al. Guillain-Barré syndrome associated with Zika virus infection. Lancet 2016, 387, 1482. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Hammack, C.; Ogden, S.C.; Wen, Z.; Qian, X.; Li, Y.; Yao, B.; Shin, J.; Zhang, F.; Lee, E.M.; et al. Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth. Cell Stem Cell 2016, 18, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.J. Defeating Zika. Sci. Transl. Med. 2017, 9, eaan2785. [Google Scholar] [CrossRef] [PubMed]
- Plourde, A.R.; Bloch, E.M. A Literature Review of Zika Virus. Emerg. Infect. Dis. 2016, 22, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Marris, E. Marine natural products: Drugs from the deep. Nature 2006, 443, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P. How might we increase success in marine-based drug discovery? Expert. Opin. Drug Discov. 2014, 9, 985–990. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Bayer, K.; Hentschel, U. Diversity, abundance and natural products of marine sponge-associated actinomycetes. Nat. Prod. Rep. 2014, 31, 381–399. [Google Scholar] [CrossRef]
- Villa, F.A.; Gerwick, L. Marine natural product drug discovery: Leads for treatment of inflammation, cancer, infections, and neurological disorders. Immunopharmacol. Immunotoxicol. 2010, 32, 228–237. [Google Scholar] [CrossRef]
- Gogineni, V.; Schinazi, R.F.; Hamann, M.T. Role of Marine Natural Products in the Genesis of Antiviral Agents. Chem. Rev. 2015, 115, 9655–9706. [Google Scholar] [CrossRef]
- Yang, C.; Li, D.; Wang, S.; Xu, M.; Wang, D.; Li, X.; Xu, X.; Li, C. Inhibitory activities of alginate phosphate and sulfate derivatives against SARS-CoV-2 in vitro. Int. J. Biol. Macromol. 2023, 227, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Tempone, A.G.; Pieper, P.; Borborema, S.E.T.; Thevenard, F.; Lago, J.H.G.; Croft, S.L.; Anderson, E.A. Marine alkaloids as bioactive agents against protozoal neglected tropical diseases and malaria. Nat. Prod. Rep. 2021, 38, 2214–2235. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Luis, S.; Gómez, J.F.; Spadafora, C.; Guzmán, H.M.; Gutiérrez, M. Antitrypanosomal alkaloids from the marine bacterium Bacillus pumilus. Molecules 2012, 17, 11146–11155. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue and Severe Dengue. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 13 February 2024).
- Teixeira, R.R.; Pereira, W.L.; Oliveira, A.F.; da Silva, A.M.; de Oliveira, A.S.; da Silva, M.L.; da Silva, C.C.; de Paula, S.O. Natural products as source of potential dengue antivirals. Molecules 2014, 19, 8151–8176. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Muñoz-Soto, R.B.; Oliva-Ramírez, J.; Vázquez-Villegas, P.; Aghamohammadi, N.; Rodriguez-Garcia, A.; Martinez-Chapa, S.O. Latest Updates in Dengue Fever Therapeutics: Natural, Marine and Synthetic Drugs. Curr. Med. Chem. 2020, 27, 719–744. [Google Scholar] [CrossRef] [PubMed]
- Koishi, A.C.; Zanello, P.R.; Bianco, É.M.; Bordignon, J.; Nunes Duarte dos Santos, C. Screening of Dengue virus antiviral activity of marine seaweeds by an in situ enzyme-linked immunosorbent assay. PLoS ONE 2012, 7, e51089. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Wu, Z.; Ji, W.; Liu, D.; Guo, X.; Yang, D.; Fan, A.; Jia, H.; Ma, M.; Lin, W. Discovery of cyclohexadepsipeptides with anti-Zika virus activities and biosynthesis of the nonproteinogenic building block (3S)-methyl-l-proline. J. Biol. Chem. 2021, 297, 100822. [Google Scholar] [CrossRef]
- Cirne-Santos, C.C.; Barros, C.D.S.; Rabelo, V.W.; Esteves, P.O.; Gomes, M.W.L.; Teixeira, V.L.; Paixão, I.C.D.P. In vitro antiviral activity against Zika virus from a natural product of the Brazilian red seaweed Bryothamnion triquetrum. Acta Virol. 2021, 65, 402–410. [Google Scholar] [CrossRef]
- Khan, R.A.A.; Najeeb, S.; Hussain, S.; Xie, B.; Li, Y. Bioactive Secondary Metabolites from Trichoderma spp. against Phytopathogenic Fungi. Microorganisms 2020, 8, 817. [Google Scholar] [CrossRef]
- Shah, A.A.; Shah, A.N.; Bilal Tahir, M.; Abbas, A.; Javad, S.; Ali, S.; Rizwan, M.; Alotaibi, S.S.; Kalaji, H.M.; Telesinski, A.; et al. Harzianopyridone Supplementation Reduced Chromium Uptake and Enhanced Activity of Antioxidant Enzymes in Vigna radiata Seedlings Exposed to Chromium Toxicity. Front. Plant Sci. 2022, 13, 881561. [Google Scholar] [CrossRef]
- Bat-Erdene, U.; Kanayama, D.; Tan, D.; Turner, W.C.; Houk, K.N.; Ohashi, M.; Tang, Y. Iterative Catalysis in the Biosynthesis of Mitochondrial Complex II Inhibitors Harzianopyridone and Atpenin B. J. Am. Chem. Soc. 2020, 142, 8550–8554. [Google Scholar] [CrossRef] [PubMed]
- Selby, T.P.; Hughes, K.A.; Rauh, J.J.; Hanna, W.S. Synthetic atpenin analogs: Potent mitochondrial inhibitors of mammalian and fungal succinate-ubiquinone oxidoreductase. Bioorg. Med. Chem. Lett. 2010, 20, 1665–1668. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.S.; Hutchins, D.C.; Mainali, R.; Goslen, K.H.; Quinn, M.A. Itaconate and Its Derivatives Repress Early Myogenesis In Vitro and In Vivo. Front. Immunol. 2022, 13, 748375. [Google Scholar] [CrossRef]
- Grant, A.; Ponia, S.S.; Tripathi, S.; Balasubramaniam, V.; Miorin, L.; Sourisseau, M.; Schwarz, M.C.; Sánchez-Seco, M.P.; Evans, M.J.; Best, S.M.; et al. Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell. Host Microbe 2016, 19, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Larocca, R.A.; Abbink, P.; Peron, J.P.; Zanotto, P.M.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’ang’a, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Coloma, J.; Jain, R.; Rajashankar, K.R.; García-Sastre, A.; Aggarwal, A.K. Structures of NS5 Methyltransferase from Zika Virus. Cell Rep. 2016, 16, 3097–3102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Tan, Z.; Wei, J.; Zhang, S.; Liu, Y.; Miao, Y.; Ding, Q.; Yi, W.; Gan, M.; Li, C.; et al. Identification of novel anti-ZIKV drugs from viral-infection temporal gene expression profiles. Emerg. Microbes Infect. 2023, 12, 2174777. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yu, J.; Chen, T.; Liu, W.; Huang, Y.; Li, J.; Zhang, B.; Zhu, G.; He, Z.; Long, Y.; et al. Design, synthesis, and biological evaluation of a series of new anthraquinone derivatives as anti-ZIKV agents. Eur. J. Med. Chem. 2023, 258, 115620. [Google Scholar] [CrossRef]
- Zhang, B.; Yu, J.; Zhu, G.; Huang, Y.; Zhang, K.; Xiao, X.; He, W.; Yuan, J.; Gao, X. Dapoxetine, a Selective Serotonin Reuptake Inhibitor, Suppresses Zika Virus Infection In Vitro. Molecules 2023, 28, 8142. [Google Scholar] [CrossRef]
- Yuan, J.; Yu, J.; Huang, Y.; He, Z.; Luo, J.; Wu, Y.; Zheng, Y.; Wu, J.; Zhu, X.; Wang, H.; et al. Antibiotic fidaxomicin is an RdRp inhibitor as a potential new therapeutic agent against Zika virus. BMC Med. 2020, 18, 204. [Google Scholar] [CrossRef]
- Han, Y.; Mesplède, T.; Xu, H.; Quan, Y.; Wainberg, M.A. The antimalarial drug amodiaquine possesses anti-ZIKA virus activities. J. Med. Virol. 2018, 90, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Baz, M.; Goyette, N.; Griffin, B.D.; Kobinger, G.P.; Boivin, G. In vitro susceptibility of geographically and temporally distinct Zika viruses to favipiravir and ribavirin. Antivir. Ther. 2017, 22, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.S.; Santos, F.C.P.; Campana, P.R.V.; Costa, V.V.; de Pádua, R.M.; Souza, D.G.; Teixeira, M.M.; Braga, F.C. Natural Products and Derivatives as Potential Zika virus Inhibitors: A Comprehensive Review. Viruses 2023, 15, 1211. [Google Scholar] [CrossRef] [PubMed]
- Barrows, N.J.; Campos, R.K.; Liao, K.C.; Prasanth, K.R.; Soto-Acosta, R.; Yeh, S.C.; Schott-Lerner, G.; Pompon, J.; Sessions, O.M.; Bradrick, S.S.; et al. Biochemistry and Molecular Biology of Flaviviruses. Chem. Rev. 2018, 118, 4448–4482. [Google Scholar] [CrossRef] [PubMed]
- Godoy, A.S.; Lima, G.M.; Oliveira, K.I.; Torres, N.U.; Maluf, F.V.; Guido, R.V.; Oliva, G. Crystal structure of Zika virus NS5 RNA-dependent RNA polymerase. Nat. Commun. 2017, 8, 14764. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, I.; Santos-Júnior, P.; Aquino, T.M.; Araújo-Júnior, J.X.; Silva-Júnior, E.F.D. Insights on Dengue and Zika NS5 RNA-dependent RNA polymerase (RdRp) inhibitors. Eur. J. Med. Chem. 2021, 224, 113698. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, H.; Song, W.; Si, S.; Han, Y.; Jiang, J. Identification and characterization of Zika virus NS5 RNA-dependent RNA polymerase inhibitors. Int. J. Antimicrob. Agents 2019, 54, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Franco, E.J.; Cella, E.; Tao, X.; Hanrahan, K.C.; Azarian, T.; Brown, A.N. Favipiravir Suppresses Zika Virus (ZIKV) through Activity as a Mutagen. Microorganisms 2023, 11, 1342. [Google Scholar] [CrossRef]
- Sariyer, I.K.; Gordon, J.; Burdo, T.H.; Wollebo, H.S.; Gianti, E.; Donadoni, M.; Bellizzi, A.; Cicalese, S.; Loomis, R.; Robinson, J.A.; et al. Suppression of Zika Virus Infection in the Brain by the Antiretroviral Drug Rilpivirine. Mol. Ther. 2019, 27, 2067–2079. [Google Scholar] [CrossRef]
- Zhu, Y.; Liang, M.; Yu, J.; Zhang, B.; Zhu, G.; Huang, Y.; He, Z.; Yuan, J. Repurposing of Doramectin as a New Anti-Zika Virus Agent. Viruses 2023, 15, 1068. [Google Scholar] [CrossRef]
- Mbaoji, F.N.; Nweze, J.A.; Yang, L.; Huang, Y.; Huang, S.; Onwuka, A.M.; Peter, I.E.; Mbaoji, C.C.; Jiang, M.; Zhang, Y.; et al. Novel Marine Secondary Metabolites Worthy of Development as Anticancer Agents: A Review. Molecules 2021, 26, 5769. [Google Scholar] [CrossRef] [PubMed]
- Schwartsmann, G. Marine organisms and other novel natural sources of new cancer drugs. Ann. Oncol. 2000, 11 (Suppl. S3), 235–243. [Google Scholar] [CrossRef] [PubMed]
- Vinothkumar, S.; Parameswaran, P.S. Recent advances in marine drug research. Biotechnol. Adv. 2013, 31, 1826–1845. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Zhang, L.; Liu, J.Z.; Chen, S.G.; Xiao, T.W.; Liu, G.Z.; Wang, J.X.; Wang, L.X.; Hou, M. Marine drug Haishengsu increases chemosensitivity to conventional chemotherapy and improves quality of life in patients with acute leukemia. Biomed. Pharmacother. 2016, 81, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Kim, H.J.; Lee, S.H. Therapeutic Application of Diverse Marine-derived Natural Products in Cancer Therapy. Anticancer. Res. 2019, 39, 5261–5284. [Google Scholar] [CrossRef]
- Sawadogo, W.R.; Boly, R.; Cerella, C.; Teiten, M.H.; Dicato, M.; Diederich, M. A Survey of Marine Natural Compounds and Their Derivatives with Anti-cancer Activity Reported in 2012. Molecules 2015, 20, 7097–7142. [Google Scholar] [CrossRef] [PubMed]
- Wender, P.A.; Hardman, C.T.; Ho, S.; Jeffreys, M.S.; Maclaren, J.K.; Quiroz, R.V.; Ryckbosch, S.M.; Shimizu, A.J.; Sloane, J.L.; Stevens, M.C. Scalable synthesis of bryostatin 1 and analogs, adjuvant leads against latent HIV. Science 2017, 358, 218–223. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, W.; Zhao, X.; Xian, Y.; Wu, W.; Zhang, X.; Zhao, N.; Xu, F.J.; Wang, C. Natural Melanin/Alginate Hydrogels Achieve Cardiac Repair through ROS Scavenging and Macrophage Polarization. Adv. Sci. 2021, 8, e2100505. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef]
- Vinale, F.; Marra, R.; Scala, F.; Ghisalberti, E.L.; Lorito, M.; Sivasithamparam, K. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Lett. Appl. Microbiol. 2006, 43, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Imperatore, C.; Gimmelli, R.; Persico, M.; Casertano, M.; Guidi, A.; Saccoccia, F.; Ruberti, G.; Luciano, P.; Aiello, A.; Parapini, S.; et al. Investigating the Antiparasitic Potential of the Marine Sesquiterpene Avarone, Its Reduced form Avarol, and the Novel Semisynthetic Thiazinoquinone Analogue Thiazoavarone. Mar. Drugs 2020, 18, 112. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.C.; Lu, C.H.; Chakraborty, R.; Kuo, Y.H. Isolation of sesquiterpenoids from sponge Dysidea avara and chemical modification of avarol as potential antitumor agents. Nat. Prod. Res. 2003, 17, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sarin, P.S.; Sun, D.; Thornton, A.; Müller, W.E. Inhibition of replication of the etiologic agent of acquired immune deficiency syndrome (human T-lymphotropic retrovirus/lymphadenopathy-associated virus) by avarol and avarone. J. Natl. Cancer Inst. 1987, 78, 663–666. [Google Scholar] [PubMed]
- Casertano, M.; Genovese, M.; Santi, A.; Pranzini, E.; Balestri, F.; Piazza, L.; Del Corso, A.; Avunduk, S.; Imperatore, C.; Menna, M.; et al. Evidence of Insulin-Sensitizing and Mimetic Activity of the Sesquiterpene Quinone Avarone, a Protein Tyrosine Phosphatase 1B and Aldose Reductase Dual Targeting Agent from the Marine Sponge Dysidea avara. Pharmaceutics 2023, 15, 528. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.C.; Deng, Q.X.; Dai, S.X. Remdesivir for severe acute respiratory syndrome coronavirus 2 causing COVID-19: An evaluation of the evidence. Travel. Med. Infect. Dis. 2020, 35, 101647. [Google Scholar] [CrossRef] [PubMed]
- Ferner, R.E.; Aronson, J.K. Chloroquine and hydroxychloroquine in covid-19. BMJ 2020, 369, m1432. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Suzuki, T.; Hashimoto, K. Mechanisms of action of fluvoxamine for COVID-19: A historical review. Mol. Psychiatry 2022, 27, 1898–1907. [Google Scholar] [CrossRef]
- Kamiyama, N.; Soma, R.; Hidano, S.; Watanabe, K.; Umekita, H.; Fukuda, C.; Noguchi, K.; Gendo, Y.; Ozaki, T.; Sonoda, A.; et al. Ribavirin inhibits Zika virus (ZIKV) replication in vitro and suppresses viremia in ZIKV-infected STAT1-deficient mice. Antivir. Res. 2017, 146, 1–11. [Google Scholar] [CrossRef]
- Snyder, B.; Goebel, S.; Koide, F.; Ptak, R.; Kalkeri, R. Synergistic antiviral activity of Sofosbuvir and type-I interferons (α and β) against Zika virus. J. Med. Virol. 2018, 90, 8–12. [Google Scholar] [CrossRef]
- Pan, T.; Peng, Z.; Tan, L.; Zou, F.; Zhou, N.; Liu, B.; Liang, L.; Chen, C.; Liu, J.; Wu, L.; et al. Nonsteroidal Anti-inflammatory Drugs Potently Inhibit the Replication of Zika Viruses by Inducing the Degradation of AXL. J. Virol. 2018, 92, 10–128. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Barbetti, M.J.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Liang, M.; Li, Y.; Zhang, K.; Zhu, Y.; Liang, J.; Liu, M.; Zhang, S.; Chen, D.; Liang, H.; Liang, L.; et al. Host factor DUSP5 potently inhibits dengue virus infection by modulating cytoskeleton rearrangement. Antivir. Res. 2023, 215, 105622. [Google Scholar] [CrossRef]
- Zhou, R.; Li, Q.; Yang, B.; Quan, Y.; Liu, Y.; Liu, M.; Zhang, Y.; Shan, G.; Li, Z.; Wang, J.; et al. Repurposing of the antihistamine mebhydrolin napadisylate for treatment of Zika virus infection. Bioorg. Chem. 2022, 128, 106024. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | EC50 (µM) 1 | EC90 (µM) 2 | CC50 (µM) 3 | SI 4 |
|---|---|---|---|---|
| SNB19 | 0.46 ± 0.13 | 1.82 ± 0.01 | 45.17 ± 2.16 | 98.20 |
| Vero | 1.40 ± 0.37 | 5.27 ± 0.26 | 92.31 ± 6.66 | 65.94 |
| A549 | 2.63 ± 0.68 | 11.19 ± 0.22 | 82.87 ± 4.94 | 34.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Liang, J.; Zhang, B.; Huang, L.; Yu, J.; Xiao, X.; He, Z.; Tao, H.; Yuan, J. A Marine Natural Product, Harzianopyridone, as an Anti-ZIKV Agent by Targeting RNA-Dependent RNA Polymerase. Molecules 2024, 29, 978. https://doi.org/10.3390/molecules29050978
Zhang K, Liang J, Zhang B, Huang L, Yu J, Xiao X, He Z, Tao H, Yuan J. A Marine Natural Product, Harzianopyridone, as an Anti-ZIKV Agent by Targeting RNA-Dependent RNA Polymerase. Molecules. 2024; 29(5):978. https://doi.org/10.3390/molecules29050978
Chicago/Turabian StyleZhang, Kexin, Jingyao Liang, Bingzhi Zhang, Lishan Huang, Jianchen Yu, Xuhan Xiao, Zhenjian He, Huaming Tao, and Jie Yuan. 2024. "A Marine Natural Product, Harzianopyridone, as an Anti-ZIKV Agent by Targeting RNA-Dependent RNA Polymerase" Molecules 29, no. 5: 978. https://doi.org/10.3390/molecules29050978
APA StyleZhang, K., Liang, J., Zhang, B., Huang, L., Yu, J., Xiao, X., He, Z., Tao, H., & Yuan, J. (2024). A Marine Natural Product, Harzianopyridone, as an Anti-ZIKV Agent by Targeting RNA-Dependent RNA Polymerase. Molecules, 29(5), 978. https://doi.org/10.3390/molecules29050978