

Teadenol B as a Component of Microorganism-Fermented Tea Extract Inhibited Breast Cancers by Promoting Autophagy
 ,
, 
Abstract
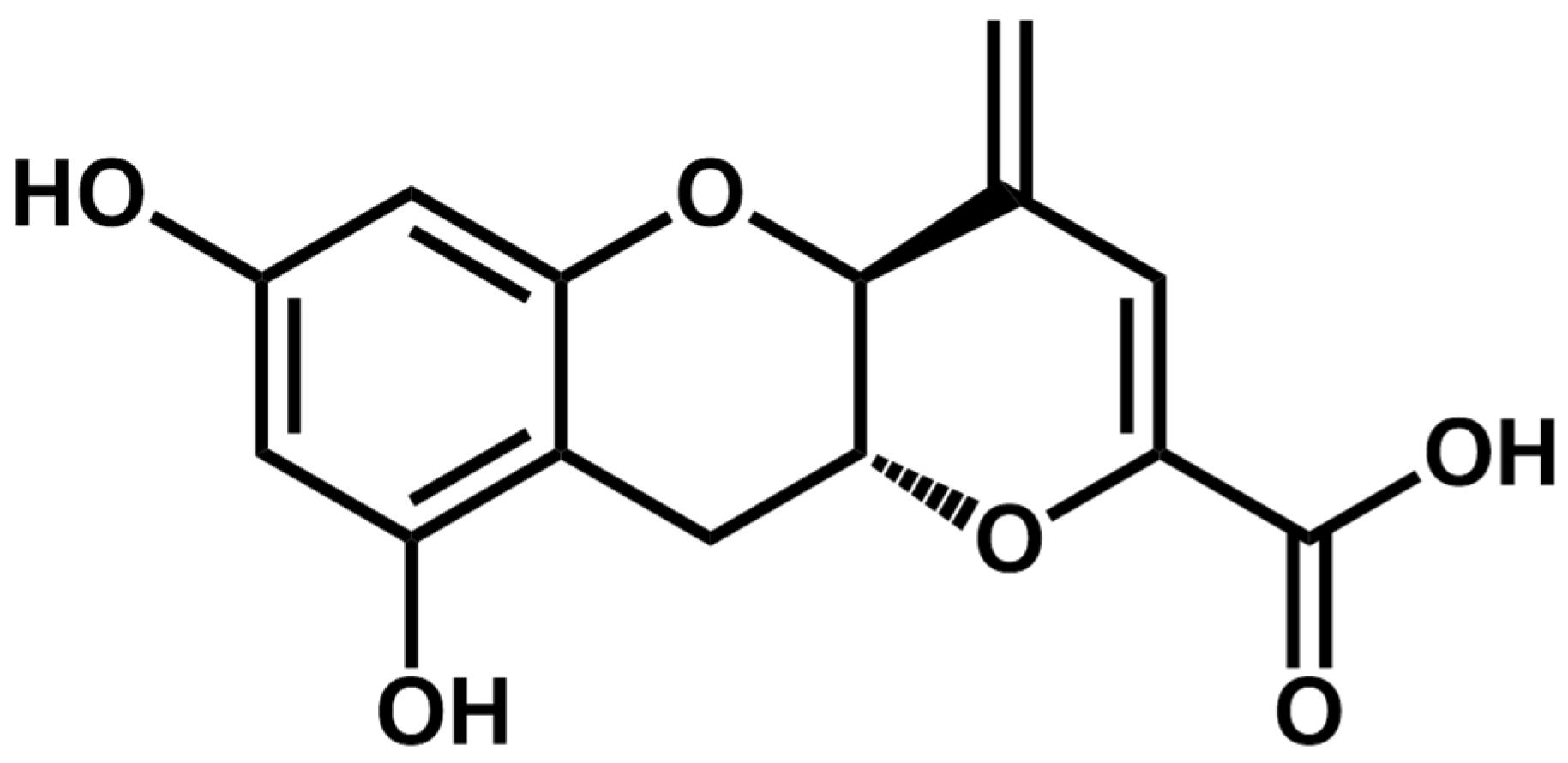
1. Introduction
2. Results and Discussion
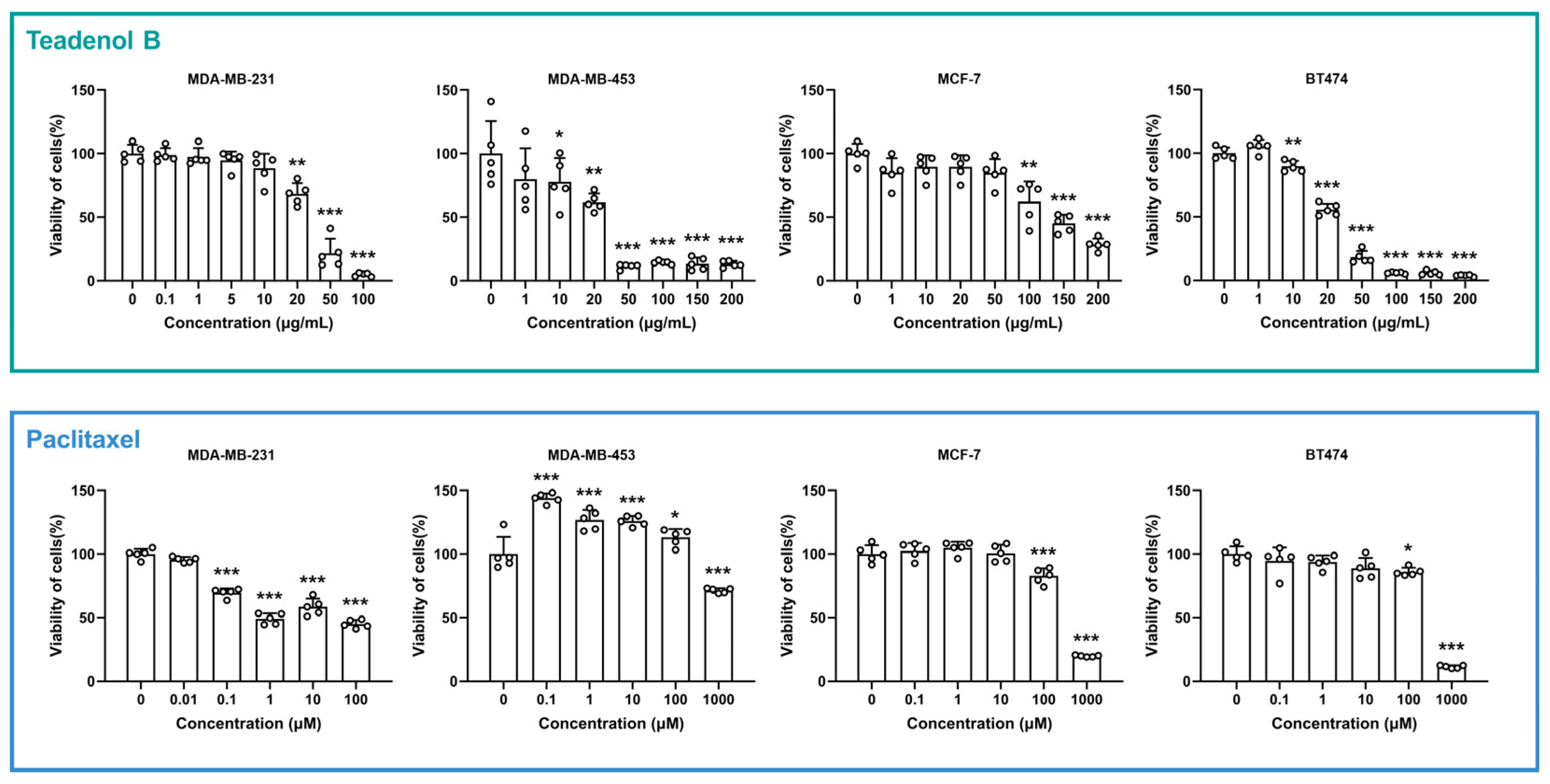
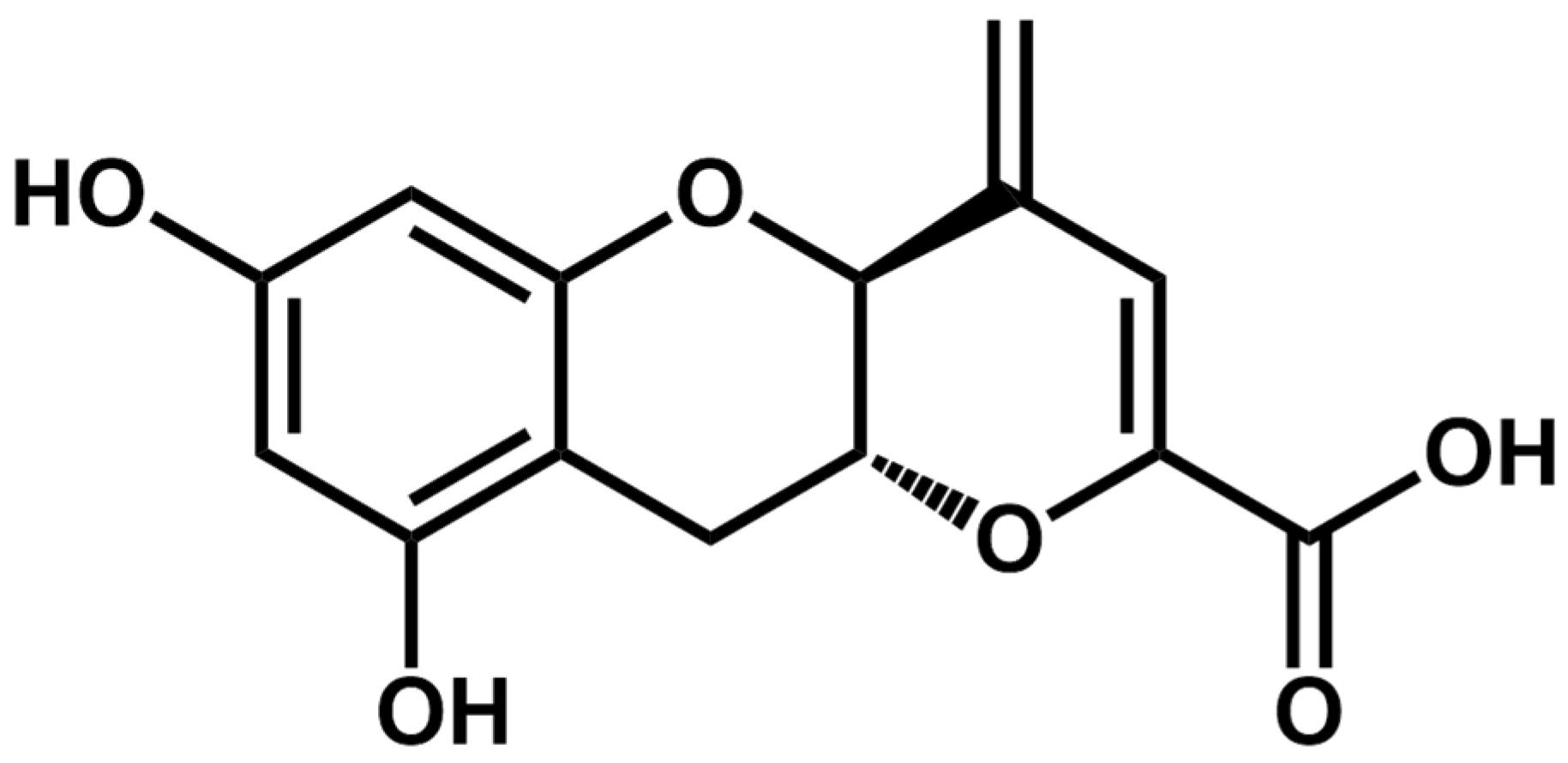
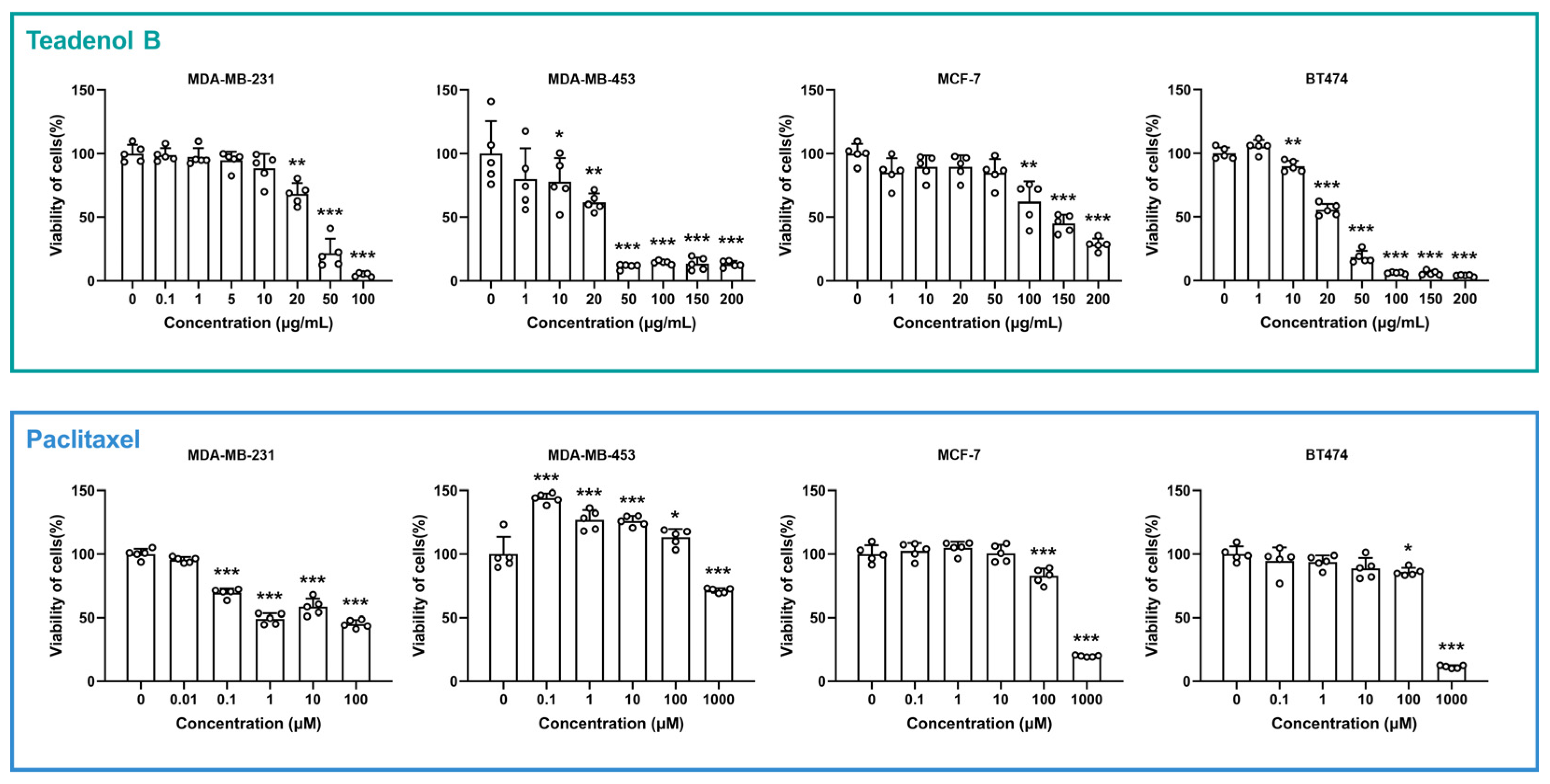
2.1. Teadenol B Has Inhibitory Activity on Breast Cancer Cells
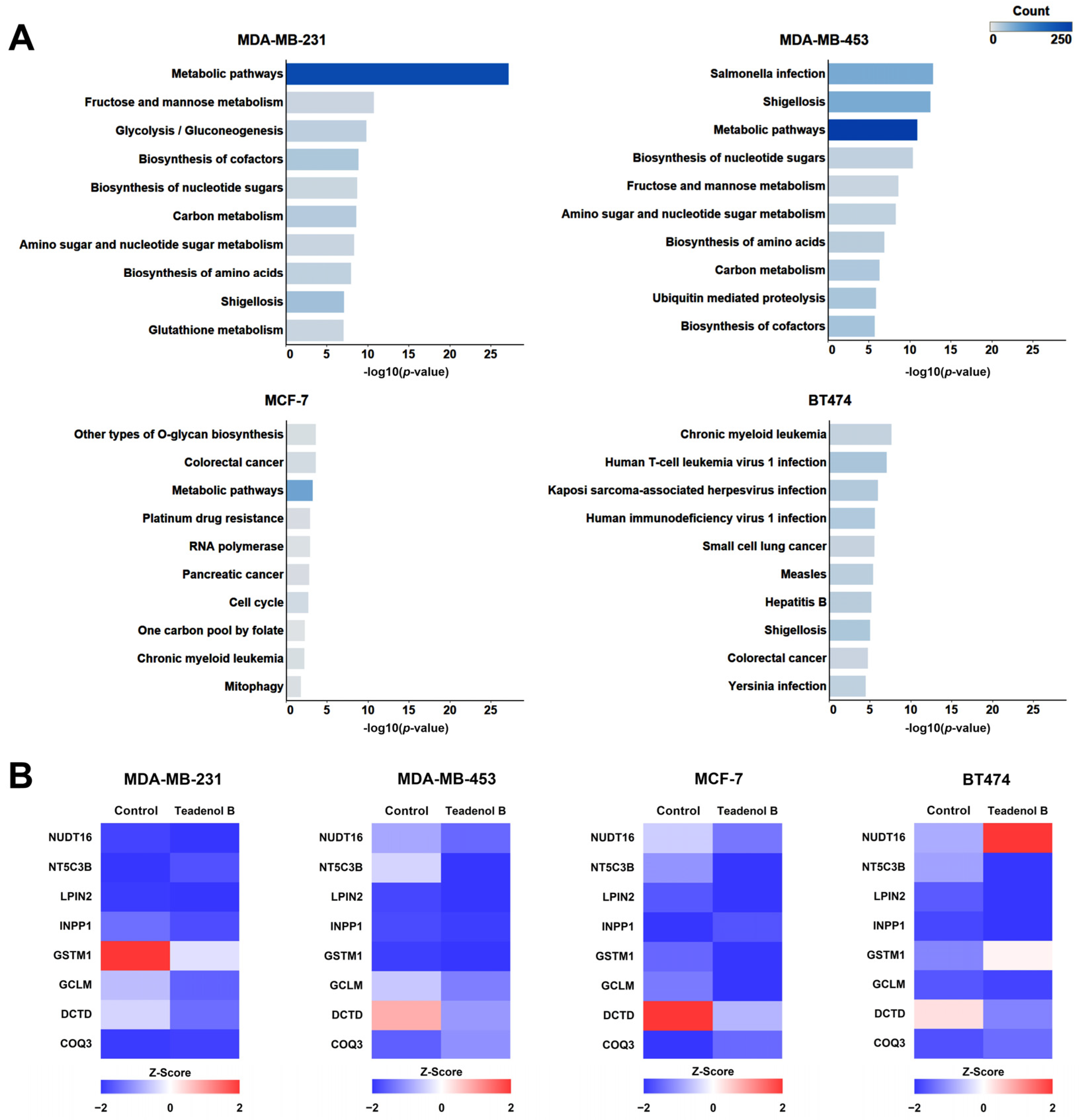
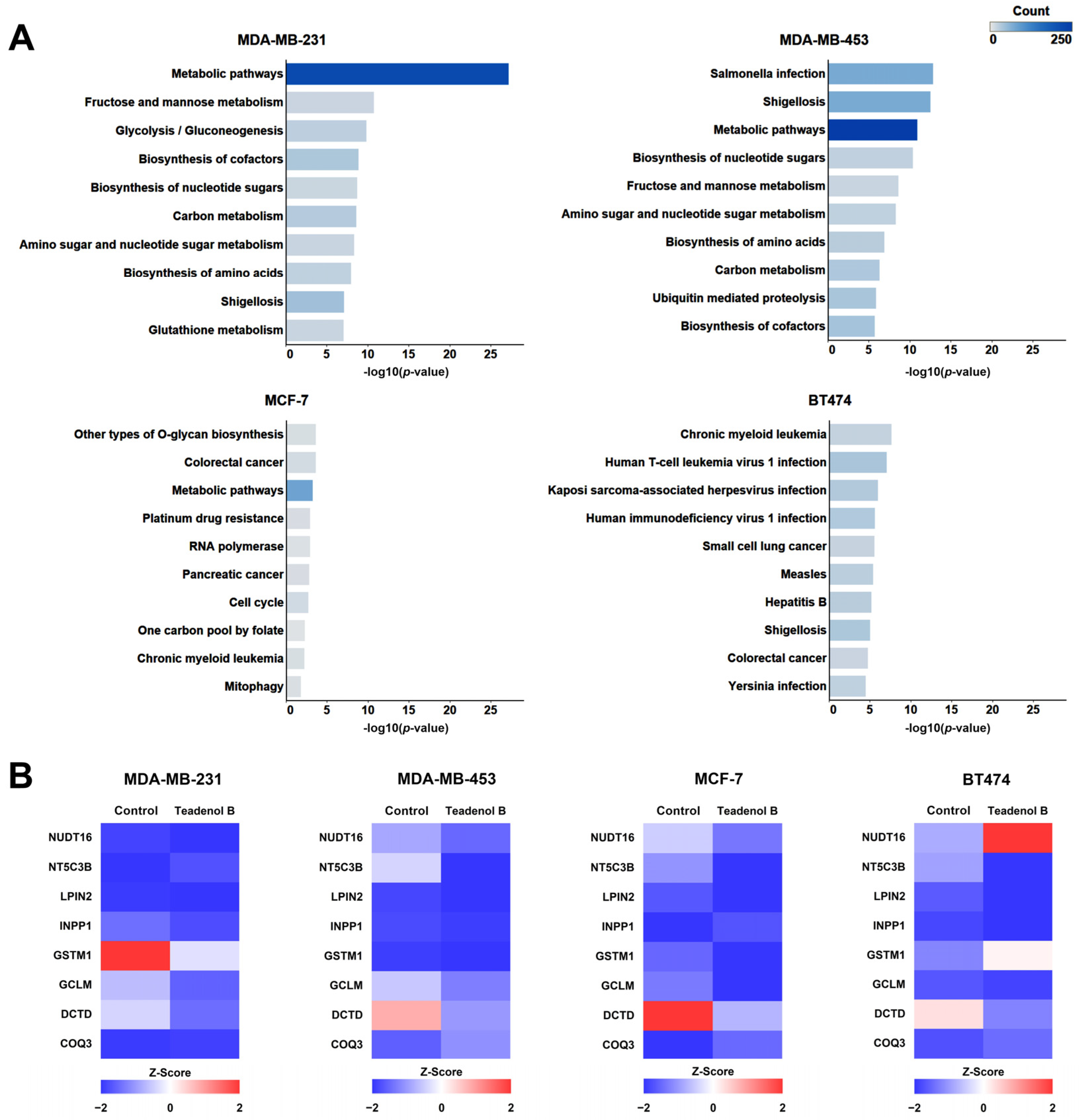
2.2. Proteomic Study of the Potential Mechanism of Action of Teadenol B against Breast Cancer
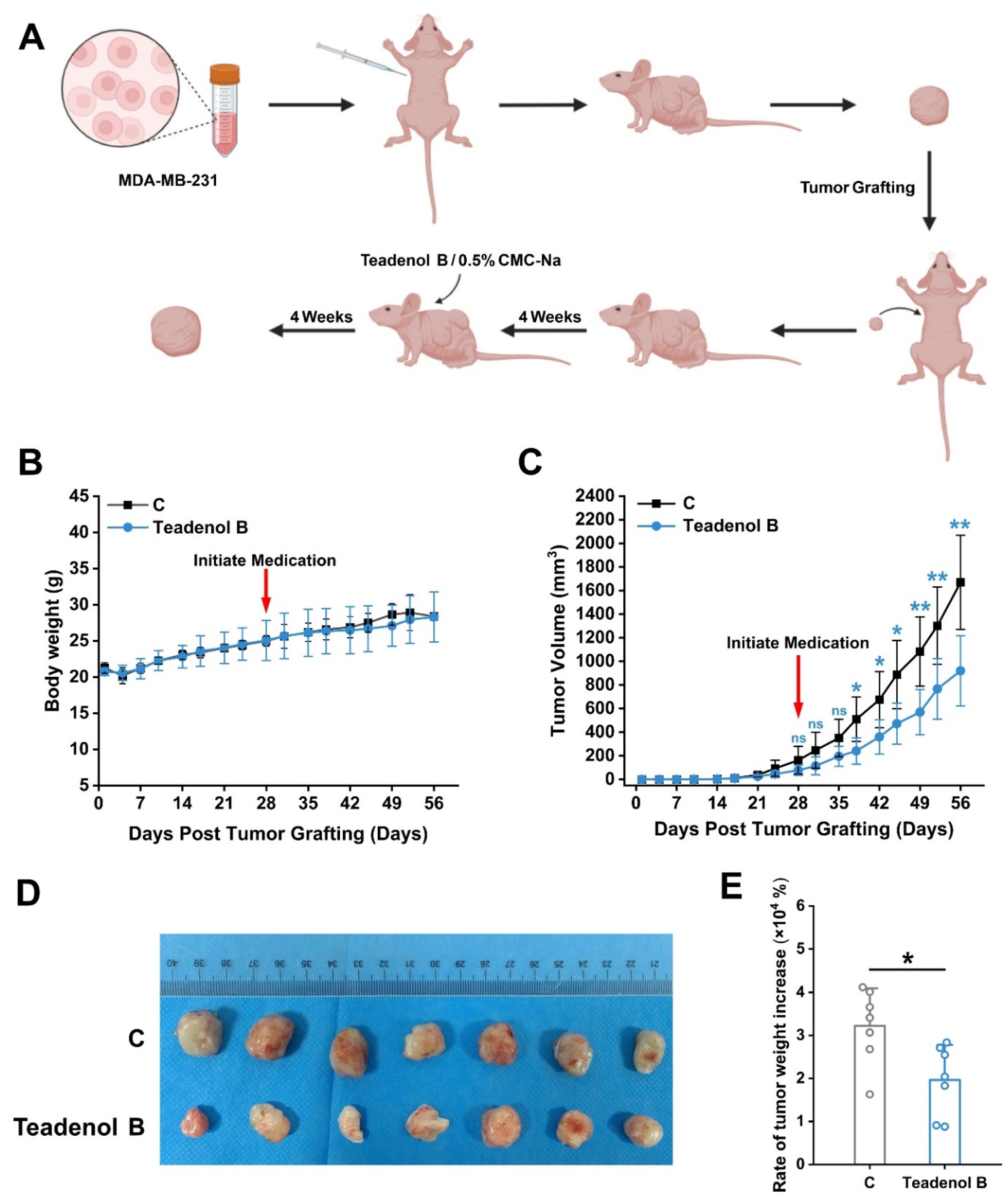
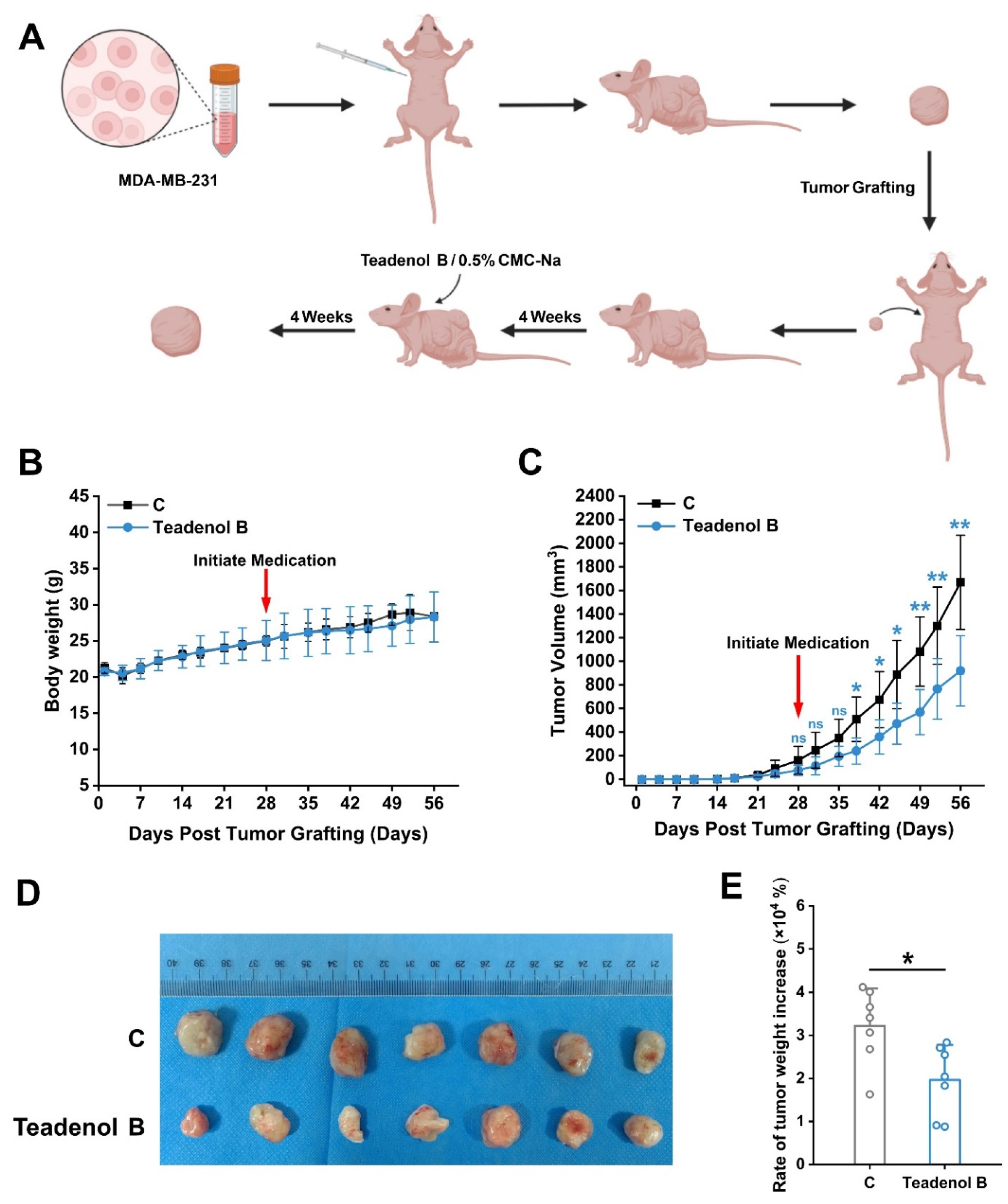
2.3. Teadenol B Inhibits the Growth of Transplanted Tumors in Nude Mice
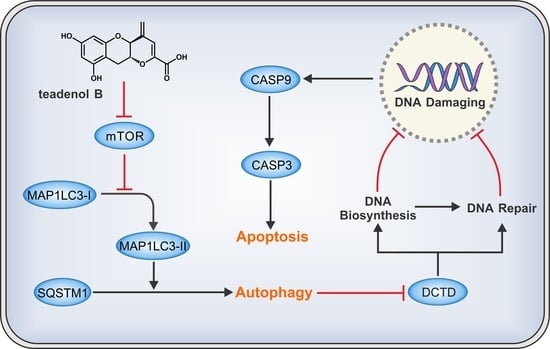
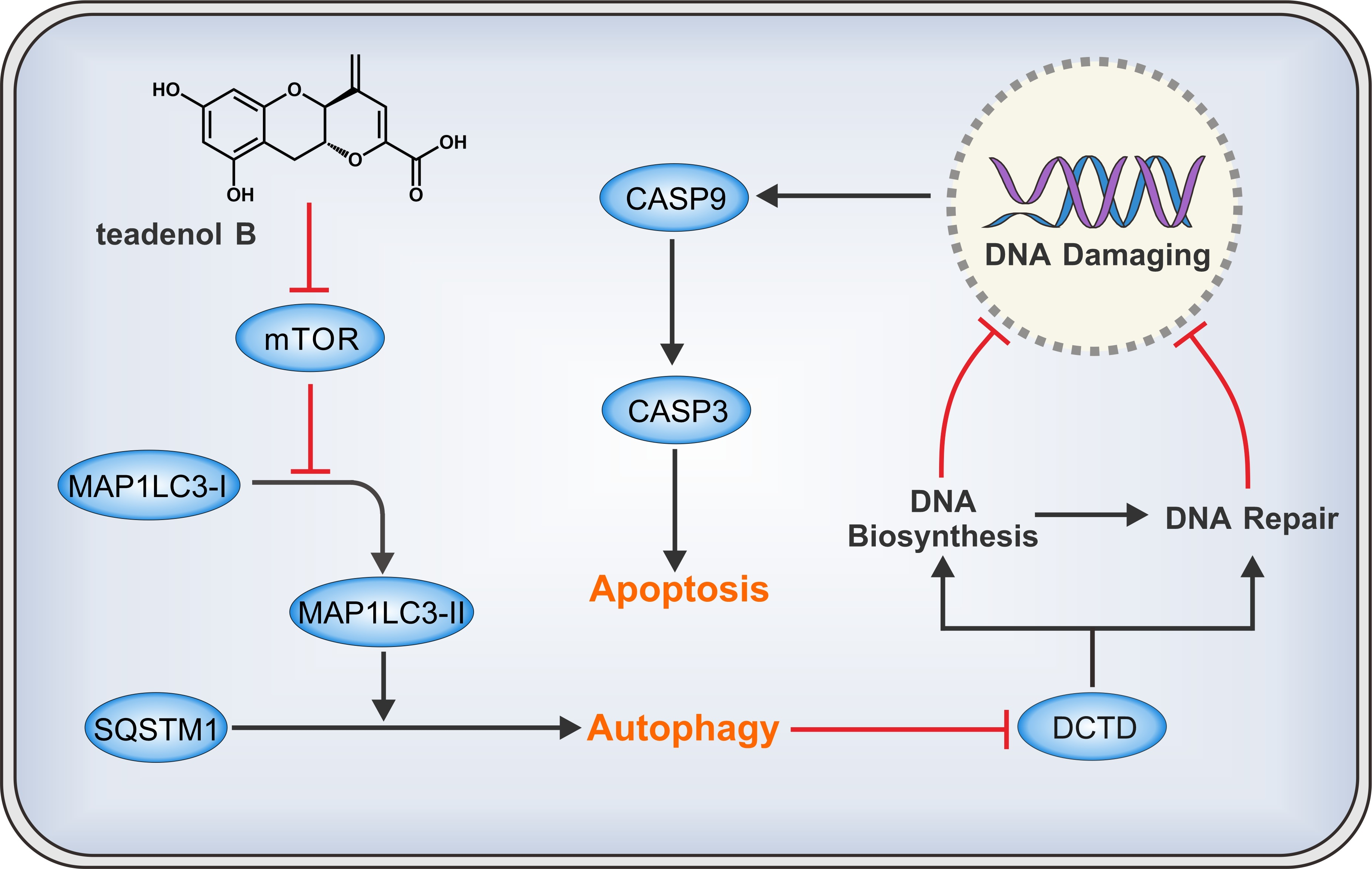
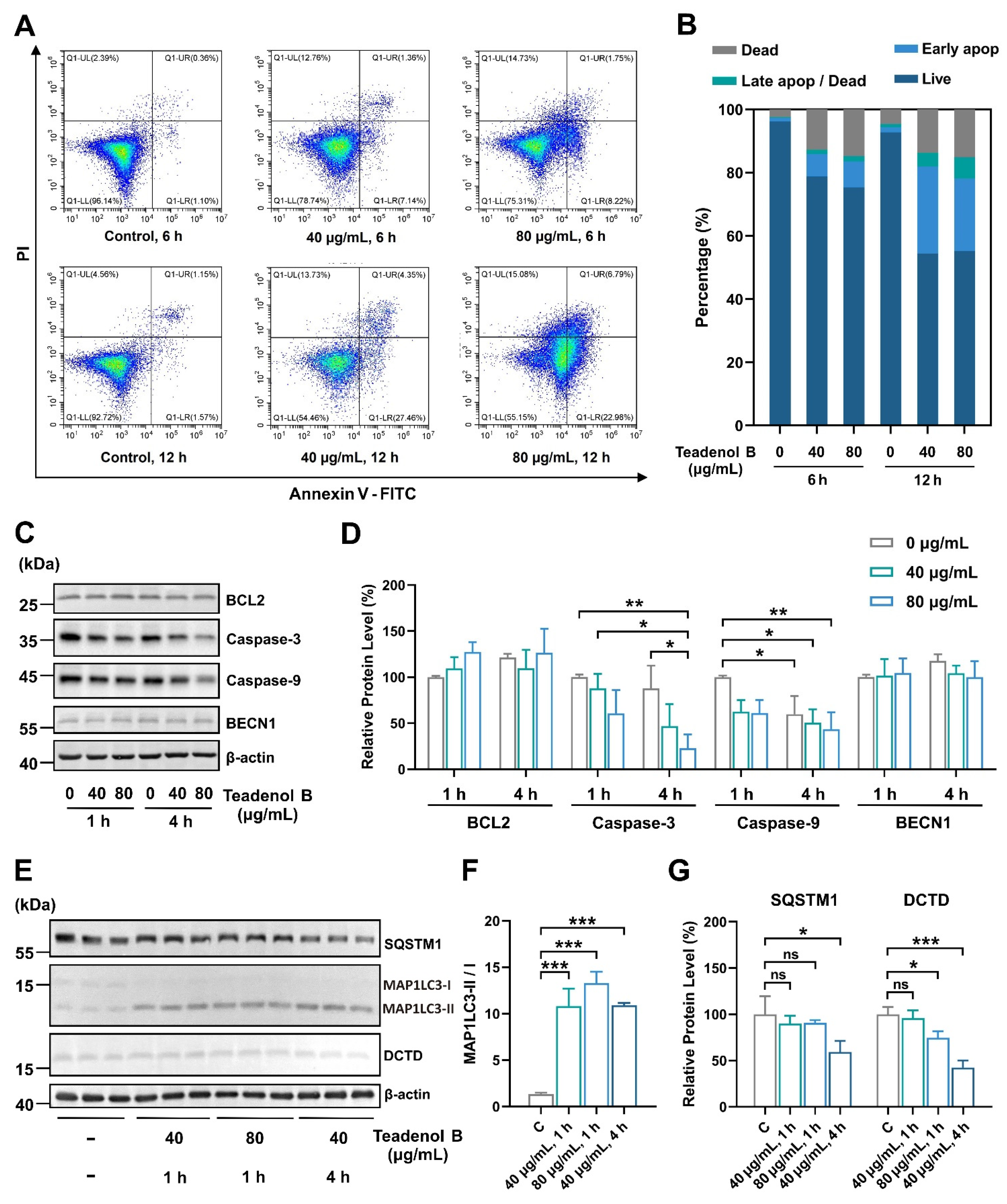
2.4. Teadenol B Inhibits MDA-MB-231 Cells by Inhibiting DCTD as an Autophagy Promotor
2.5. Discussion
3. Materials and Methods
3.1. Cell Culture
3.2. MTT Assays
3.3. Proteomic Sample Preparation
3.4. Label-Free MS Analysis
3.5. Human Breast Cancer Xenograft Nude Mice Model
3.6. Western Blot
3.7. Apoptosis Assay
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kolak, A.; Kaminska, M.; Sygit, K.; Budny, A.; Surdyka, D.; Kukielka-Budny, B.; Burdan, F. Primary and secondary prevention of breast cancer. Ann. Agric. Environ. Med. 2017, 24, 549–553. [Google Scholar] [CrossRef]
- McGuire, A.; Brown, J.A.; Malone, C.; McLaughlin, R.; Kerin, M.J. Effects of age on the detection and management of breast cancer. Cancers 2015, 7, 908–929. [Google Scholar] [CrossRef] [PubMed]
- Sviderskiy, V.O.; Blumenberg, L.; Gorodetsky, E.; Karakousi, T.R.; Hirsh, N.; Alvarez, S.W.; Terzi, E.M.; Kaparos, E.; Whiten, G.C.; Ssebyala, S.; et al. Hyperactive CDK2 Activity in Basal-like Breast Cancer Imposes a Genome Integrity Liability that Can Be Exploited by Targeting DNA Polymerase ε. Mol. Cell 2020, 80, 682–698.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, J.; Qu, C.; Peng, Y.; Lei, J.; Sun, L.; Zong, B.; Liu, S. A look into the link between centrosome amplification and breast cancer. Biomed. Pharmacother. 2020, 132, 110924. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Han, X.; Lu, J.; McEachern, D.; Wang, S. A highly potent PROTAC androgen receptor (AR) degrader ARD-61 effectively inhibits AR-positive breast cancer cell growth in vitro and tumor growth in vivo. Neoplasia 2020, 22, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Liu, X.; Zhou, Y.; Wang, H.; Zhao, Y.; Ren, J.; Xiang, J. Berberine Inhibits MDA-MB-231 Cells as an Agonist of G Protein-Coupled Estrogen Receptor 1. Int. J. Mol. Sci. 2021, 22, 11466. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Wang, Y.; Su, K.; Liu, M.; Hu, P.C.; Ma, T.; Li, J.X.; Wei, L.; Zheng, Z.; Yang, F. Ritonavir binds to and downregulates estrogen receptors: Molecular mechanism of promoting early atherosclerosis. Exp. Cell Res. 2014, 327, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Gnant, M. Breast cancer. Lancet 2017, 389, 1134–1150. [Google Scholar] [CrossRef]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast cancer: Biology, biomarkers, and treatments. Int. Immunopharmacol. 2020, 84, 106535. [Google Scholar] [CrossRef]
- Maughan, K.L.; Lutterbie, M.A.; Ham, P.S. Treatment of breast cancer. Am. Fam. Physician 2010, 81, 1339–1346. [Google Scholar]
- Brody, H. Tea. Nature 2019, 566, S1. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, Z.Y. Tea and cancer. J. Natl. Cancer Inst. 1993, 85, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Trevisanato, S.I.; Kim, Y.I. Tea and health. Nutr. Rev. 2000, 58, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Camfield, D.A.; Stough, C.; Farrimond, J.; Scholey, A.B. Acute effects of tea constituents L-theanine, caffeine, and epigallocatechin gallate on cognitive function and mood: A systematic review and meta-analysis. Nutr. Rev. 2014, 72, 507–522. [Google Scholar] [CrossRef] [PubMed]
- van Dam, R.M.; Hu, F.B.; Willett, W.C. Coffee, Caffeine, and Health. N. Engl. J. Med. 2020, 383, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Kolesárová, A. The anti-obesity and health-promoting effects of tea and coffee. Physiol. Res. 2021, 70, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Randell, R. Fat burners: Nutrition supplements that increase fat metabolism. Obes. Rev. 2011, 12, 841–851. [Google Scholar] [CrossRef]
- Vázquez Cisneros, L.C.; López-Uriarte, P.; López-Espinoza, A.; Navarro Meza, M.; Espinoza-Gallardo, A.C.; Guzmán Aburto, M.B. Effects of green tea and its epigallocatechin (EGCG) content on body weight and fat mass in humans: A systematic review. Nutr. Hosp. 2017, 34, 731–737. [Google Scholar] [CrossRef]
- Sinha, D.; Biswas, J.; Nabavi, S.M.; Bishayee, A. Tea phytochemicals for breast cancer prevention and intervention: From bench to bedside and beyond. Semin. Cancer Biol. 2017, 46, 33–54. [Google Scholar] [CrossRef]
- Mao, X.; Xiao, X.; Chen, D.; Yu, B.; He, J. Tea and Its Components Prevent Cancer: A Review of the Redox-Related Mechanism. Int. J. Mol. Sci. 2019, 20, 5249. [Google Scholar] [CrossRef]
- Zhao, L.G.; Li, Z.Y.; Feng, G.S.; Ji, X.W.; Tan, Y.T.; Li, H.L.; Gunter, M.J.; Xiang, Y.B. Tea Drinking and Risk of Cancer Incidence: A Meta-Analysis of Prospective Cohort Studies and Evidence Evaluation. Adv. Nutr. 2021, 12, 402–412. [Google Scholar] [CrossRef]
- Zhu, M.Z.; Li, N.; Zhou, F.; Ouyang, J.; Lu, D.M.; Xu, W.; Li, J.; Lin, H.Y.; Zhang, Z.; Xiao, J.B.; et al. Microbial bioconversion of the chemical components in dark tea. Food Chem. 2020, 312, 126043. [Google Scholar] [CrossRef]
- Lv, S.; Wu, Y.; Zhou, J.; Lian, M.; Li, C.; Xu, Y.; Liu, S.; Wang, C.; Meng, Q. The study of fingerprint characteristics of Dayi Pu-Erh tea using a fully automatic HS-SPME/GC-MS and combined chemometrics method. PLoS ONE 2014, 9, e116428. [Google Scholar] [CrossRef]
- Wulandari, R.A.; Amano, M.; Yanagita, T.; Tanaka, T.; Kouno, I.; Kawamura, D.; Ishimaru, K. New phenolic compounds from Camellia sinensis L. leaves fermented with Aspergillus sp. J. Nat. Med. 2011, 65, 594–597. [Google Scholar] [CrossRef]
- Song, J.-H.; Miyazaki, H.; Yoshida, S. Simple Method for the Preparation of Teadenols A and B by a Combined Process of Submerged Culture with Aspergillus sp. and Chromatographic Separation. Food Sci. Technol. Res. 2017, 23, 91–100. [Google Scholar] [CrossRef]
- Yanagita, T.; Ishimaru, K.; Tanaka, T.; Koba, K.; Miyazaki, H.; Aoki, N.; Kawamura, D. Functional Microbially Fermented Tea Extract Containing Polyphenol Derivative and Method for Producing the Same. U.S. Patent US9029110B2, 12 May 2015. [Google Scholar]
- Yanagita, T.; Ishimaru, K.; Tanaka, T.; Koba, K.; Miyazaki, H.; Aoki, N.; Kawamura, D. Polyphenol Derivative and Method for Producing the Same. U.S. Patent US8558016B2, 15 October 2013. [Google Scholar]
- Liu, J.; Ding, Z.; Gao, L.; Chen, D.; Liu, M.; Tang, S. Methods for Separating Teadenol A and Teadenol B from Fermented Tea. CN Patent No. CN108117558B, 30 October 2020. [Google Scholar]
- Nagasawa, T.; Ishimaru, K.; Higashiyama, S.; Hama, Y.; Mitsutake, S. Teadenol A in microbial fermented tea acts as a novel ligand on GPR120 to increase GLP-1 secretion. Food Funct. 2020, 11, 10534–10541. [Google Scholar] [CrossRef]
- Weiner, K.X.B.; Ciesla, J.; Jaffe, A.B.; Ketring, R.; Maley, F.; Maley, G.F. Chromosomal Location and Structural Organization of the Human Deoxycytidylate Deaminase Gene. J. Biol. Chem. 1995, 270, 18727–18729. [Google Scholar] [CrossRef]
- Radha, G.; Raghavan, S.C. BCL2: A promising cancer therapeutic target. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Liu, X.; Ren, J.; Chen, K.; Wang, H.L.; Miao, Y.Y.; Qi, M.M. How does estrogen work on autophagy? Autophagy 2019, 15, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yan, Y.; Lv, H.; Li, J.L.; Wang, Z.Y.; Wang, K.; Wang, L.; Li, Y.X.; Jiang, H.; Zhang, Y.Y. Rapamycin targets STAT3 and impacts c-Myc to suppress tumor growth. Cell Chem. Biol. 2022, 29, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, G.A.; Al-Abd, A.M.; Tadros, M.G.; Al-Abbasi, F.A.; Khalifa, A.E.; Abdel-Naim, A.B. The chemomodulatory effects of resveratrol and didox on herceptin cytotoxicity in breast cancer cell lines. Sci. Rep. 2015, 5, 12054. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.J.; Zhou, Y.L.; Xie, S.D.; Wang, J.; Li, Z.Q.; Chen, L.N.; Mao, M.S.; Chen, C.; Huang, A.H.; Chen, Y.X.; et al. Metformin induces Ferroptosis by inhibiting UFMylation of SLC7A11 in breast cancer. J. Exp. Clin. Cancer Res. 2021, 40, 206. [Google Scholar] [CrossRef] [PubMed]
- Yesil-Celiktas, O.; Sevimli, C.; Bedir, E.; Vardar-Sukan, F. Inhibitory Effects of Rosemary Extracts, Carnosic Acid and Rosma rinic Acid on the Growth of Various Human Cancer Cell Lines. Plant Food Hum. Nutr. 2010, 65, 158–163. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Receptor Phenotypic Characteristics | Teadenol B | Paclitaxel | ||
|---|---|---|---|---|---|
| (µg/mL) | (µM) | (µg/mL) | (µM) | ||
| MDA-MB-231 | ER−, PR−, HER2− | 35 | 126 | 64 | 75 |
| MDA-MB-453 | ER−, PR−, HER2+ | 30 | 108 | >850 | >1000 |
| MCF-7 | ER+, PR+/−, HER2− | 145 | 525 | 425 | 500 |
| BT474 | ER+, PR+/−, HER2+ | 25 | 90 | 425 | 500 |
| Cell Line | Up Regulation (fc ≥ 2) | Down Regulation (fc ≤ 0.5) | Differentially Expressed Proteins |
|---|---|---|---|
| MDA-MB-231 | 473 | 872 | 1345 |
| MDA-MB-453 | 472 | 1604 | 2076 |
| MCF-7 | 272 | 416 | 688 |
| BT474 | 331 | 932 | 1263 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Ding, Z.-G.; Yan, Y.-J.; Yang, R.; Qi, M.-M.; Pan, S.-K.; Xie, J.-L.; Sun, Y.-H.; Xiang, J. Teadenol B as a Component of Microorganism-Fermented Tea Extract Inhibited Breast Cancers by Promoting Autophagy. Molecules 2024, 29, 872. https://doi.org/10.3390/molecules29040872
Zhao Y, Ding Z-G, Yan Y-J, Yang R, Qi M-M, Pan S-K, Xie J-L, Sun Y-H, Xiang J. Teadenol B as a Component of Microorganism-Fermented Tea Extract Inhibited Breast Cancers by Promoting Autophagy. Molecules. 2024; 29(4):872. https://doi.org/10.3390/molecules29040872
Chicago/Turabian StyleZhao, Ying, Zhang-Gui Ding, Yu-Jie Yan, Rui Yang, Miao-Miao Qi, Shu-Kang Pan, Ji-Ling Xie, Yu-Hui Sun, and Jin Xiang. 2024. "Teadenol B as a Component of Microorganism-Fermented Tea Extract Inhibited Breast Cancers by Promoting Autophagy" Molecules 29, no. 4: 872. https://doi.org/10.3390/molecules29040872
APA StyleZhao, Y., Ding, Z.-G., Yan, Y.-J., Yang, R., Qi, M.-M., Pan, S.-K., Xie, J.-L., Sun, Y.-H., & Xiang, J. (2024). Teadenol B as a Component of Microorganism-Fermented Tea Extract Inhibited Breast Cancers by Promoting Autophagy. Molecules, 29(4), 872. https://doi.org/10.3390/molecules29040872