





Integrating High-Resolution Mass Spectral Data, Bioassays and Computational Models to Annotate Bioactives in Botanical Extracts: Case Study Analysis of C. asiatica Extract Associates Dicaffeoylquinic Acids with Protection against Amyloid-β Toxicity

 ,
,  , ,
, ,  ,
,  , and
, and
Abstract
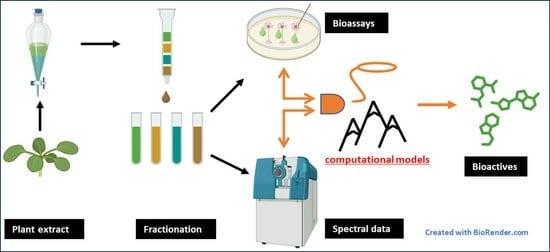
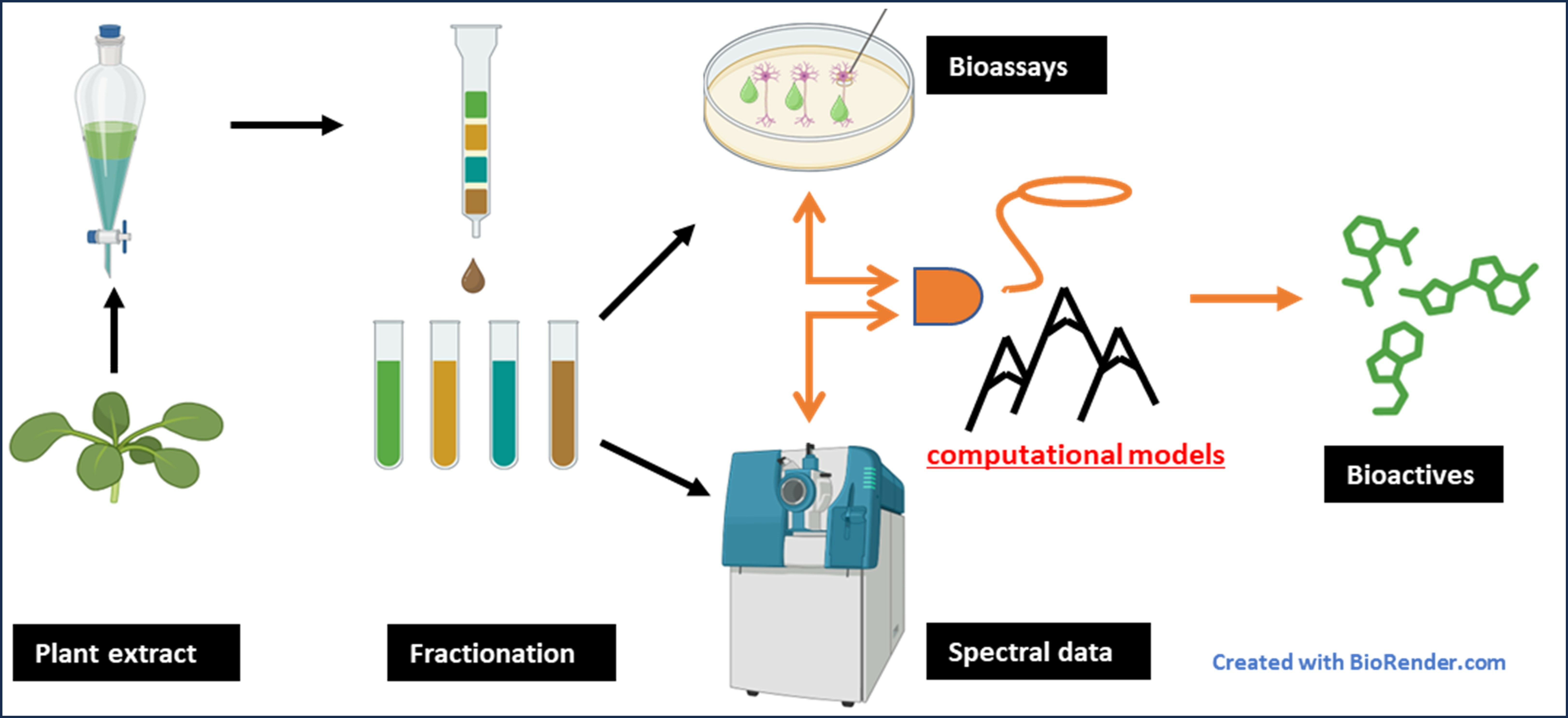
1. Introduction
2. Results and Discussion
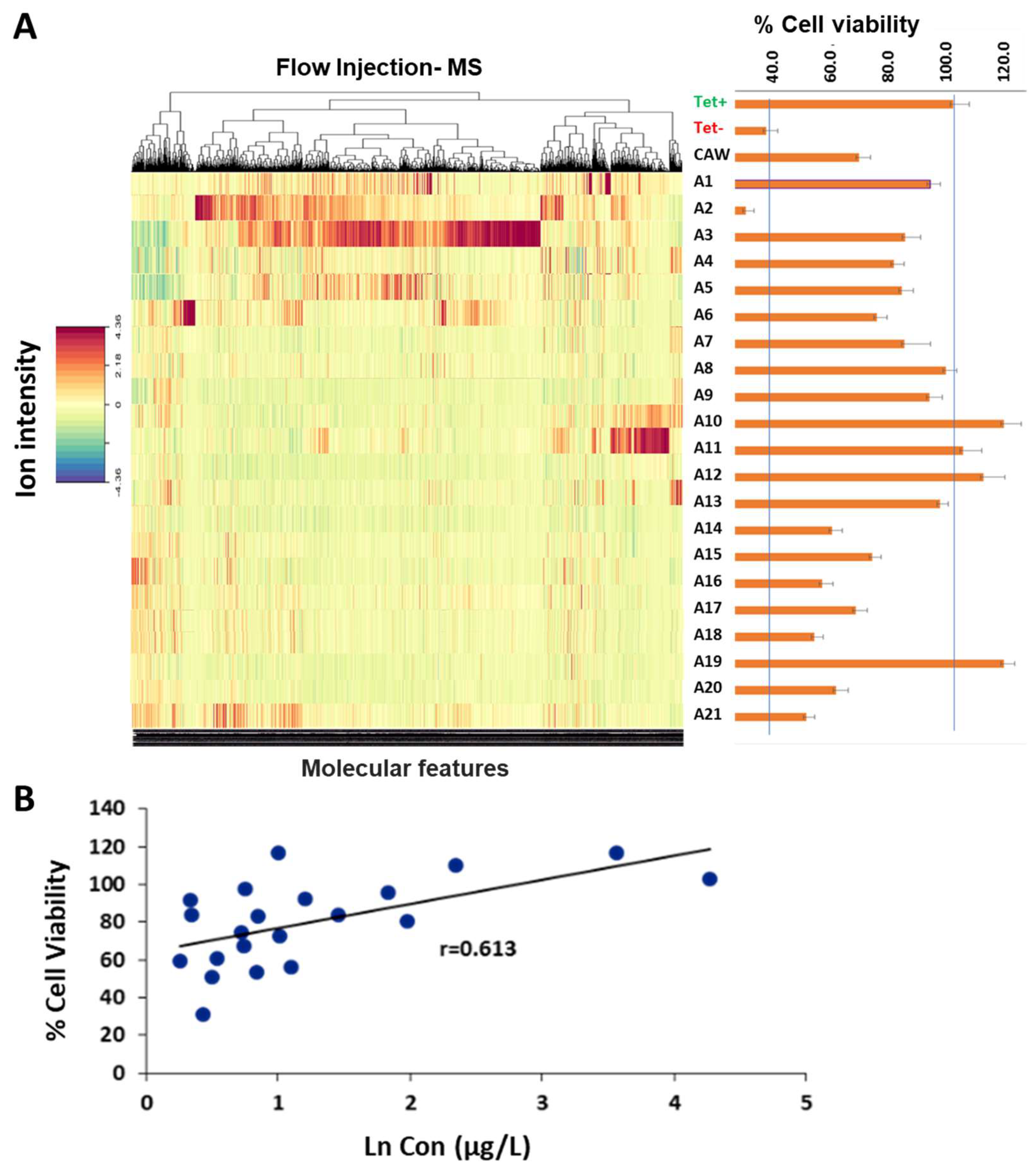
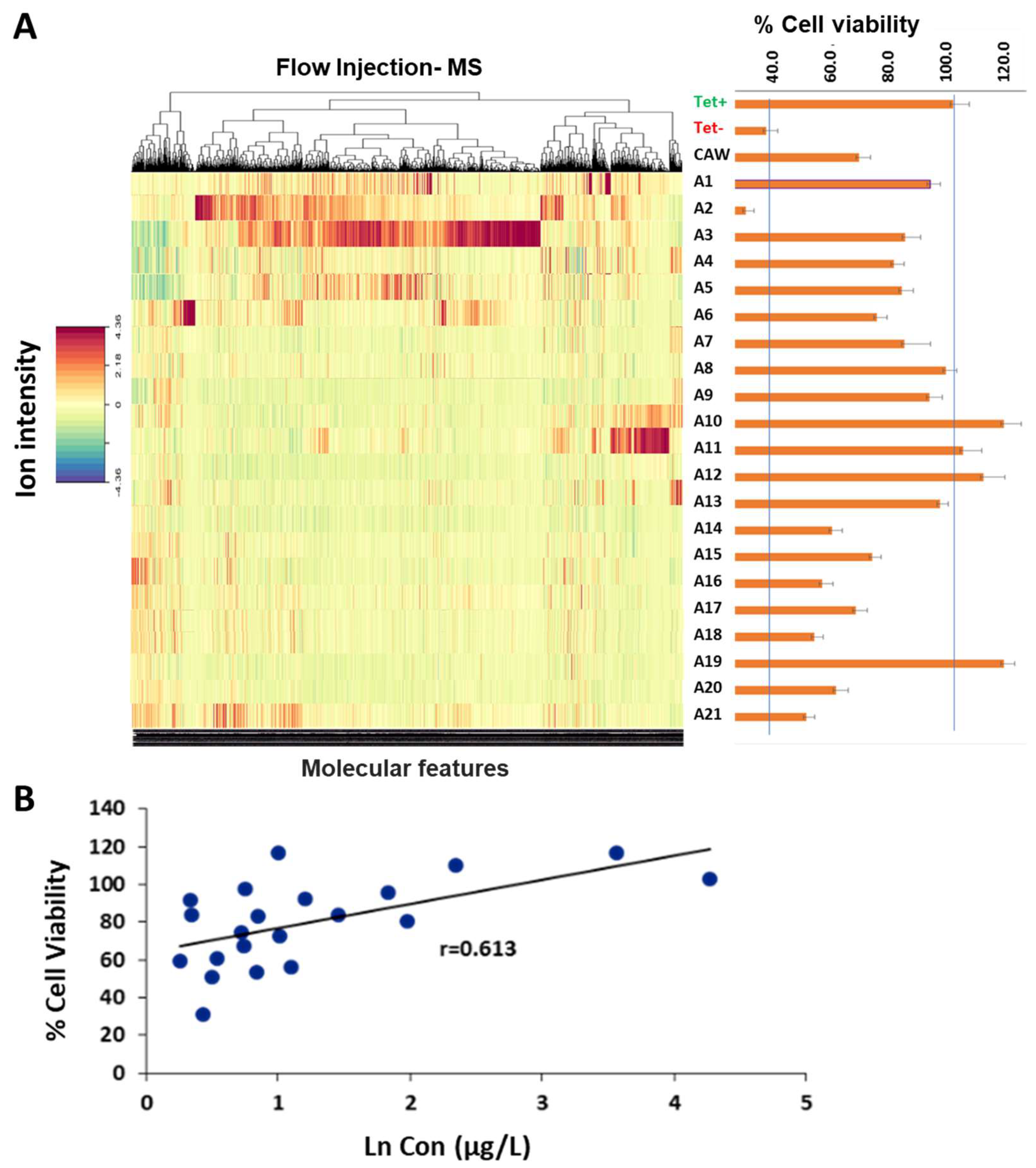
2.1. Chemical Diversity and Viability in C. asiatica Fractions
2.2. Correlation between Phytochemical Profiles and Neuroprotective Effect
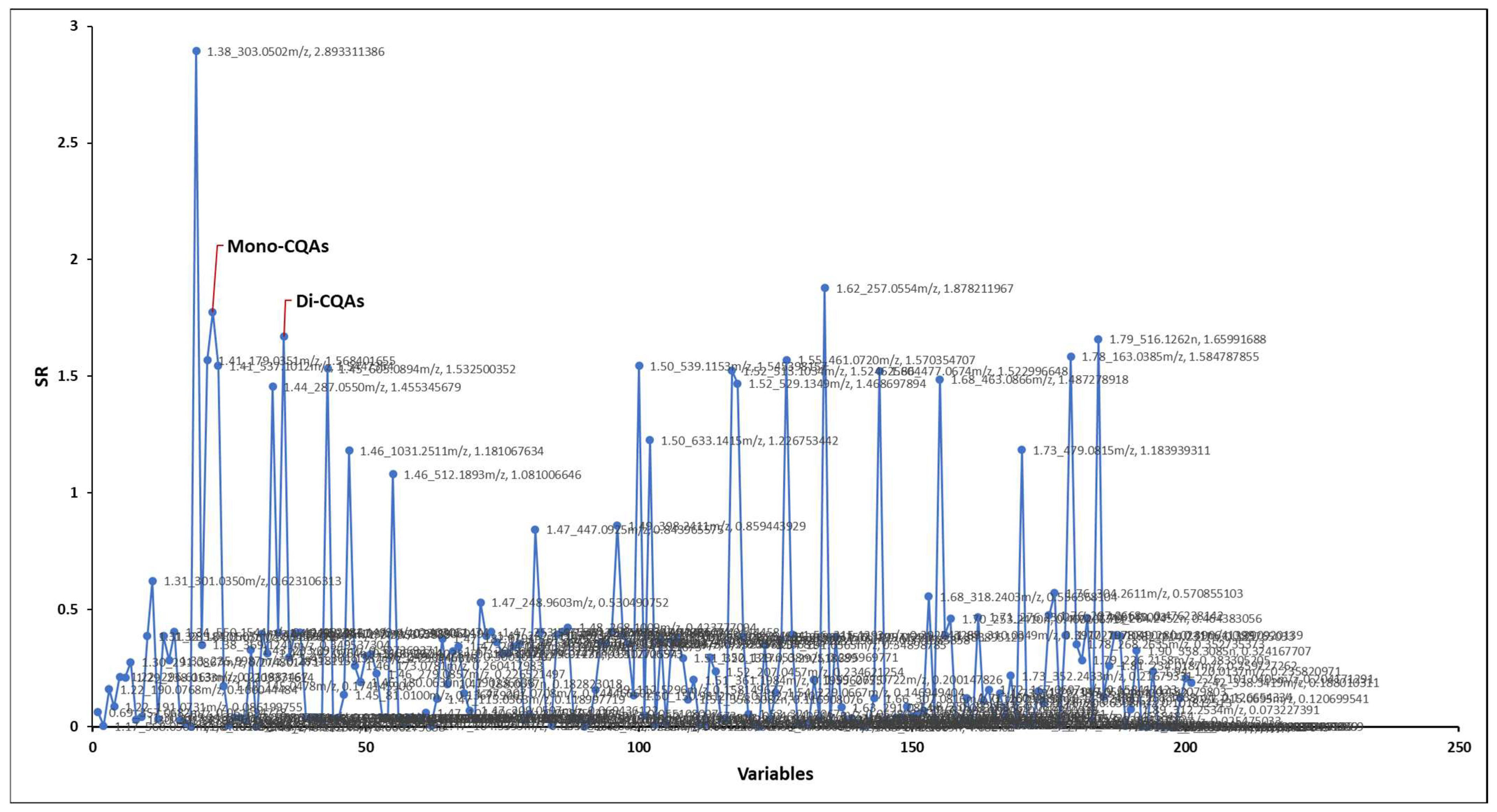
2.3. Identification of Neuroprotective Phytochemicals
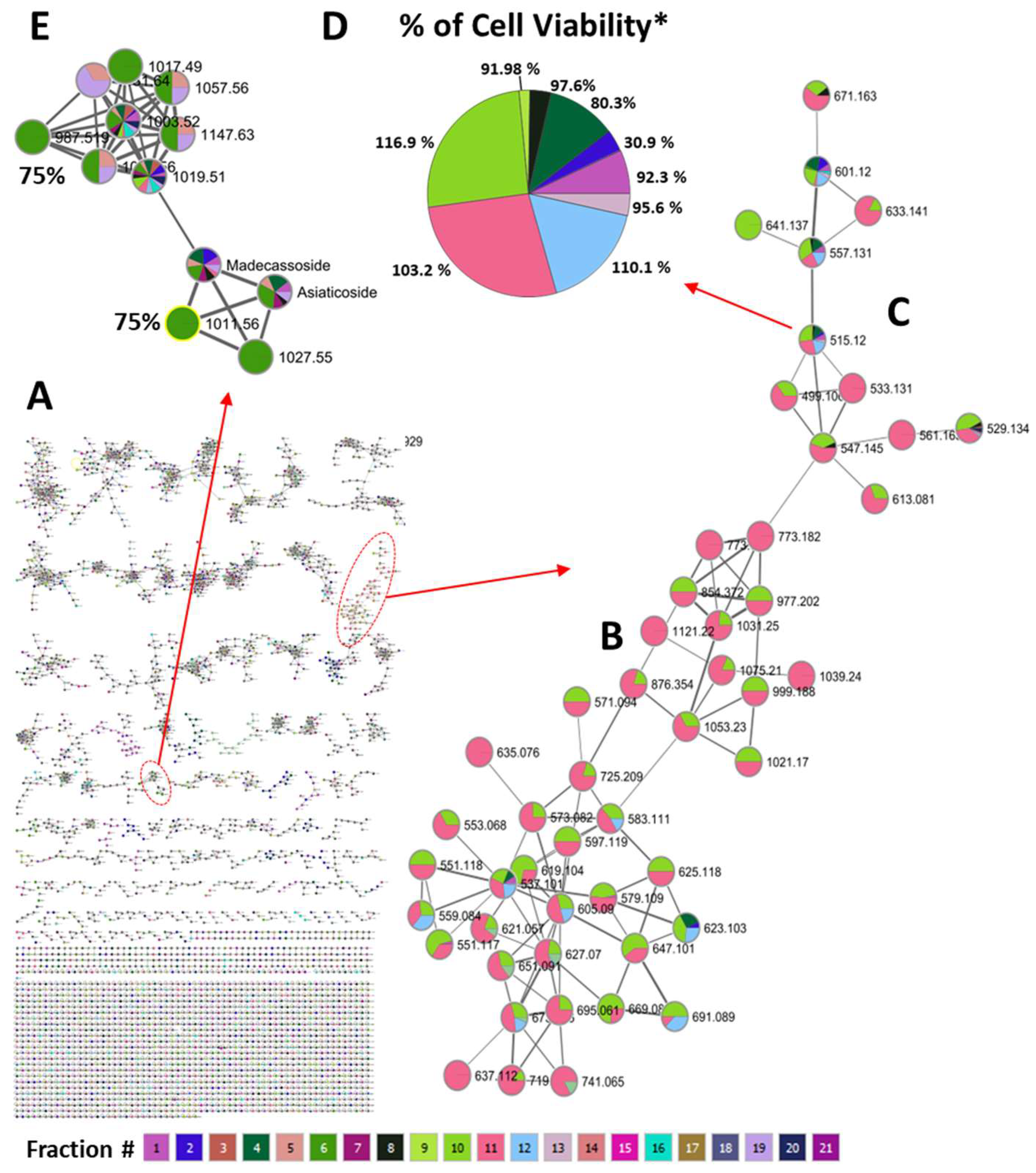
2.4. Molecular Networking for Analyzing Chemical Diversity
3. Materials and Methods
3.1. Associating Chemical Diversity of C. asiatica with % Viability from MC65 Bioassay
3.2. Biological Activity in MC65 Cellular Line
3.3. Profiling of Fractions Using Flow-Injection-HRMS
3.4. Predicting Protective Biological Activity with Mass Spectral Data
3.5. Compound Identification
3.6. Molecular Networking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyne, M.E.; Narcross, L.; Martin, V.J.J. Engineering Plant Secondary Metabolism in Microbial Systems. Plant Physiol. 2019, 179, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Mahady, G.B. Global harmonization of herbal health claims. J. Nutr. 2001, 131, 1120s–1123s. [Google Scholar] [CrossRef] [PubMed]
- Commisso, M.; Strazzer, P.; Toffali, K.; Stocchero, M.; Guzzo, F. Untargeted metabolomics: An emerging approach to determine the composition of herbal products. Comput. Struct. Biotechnol. J. 2013, 4, e201301007. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.G. A Unifying Review of Bioassay-Guided Fractionation, Effect-Directed Analysis and Related Techniques. Sensors 2012, 12, 9181–9209. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.F.; Nothias-Esposito, M.; da Silva, R.; Wang, M.; Protsyuk, I.; Zhang, Z.; Sarvepalli, A.; Leyssen, P.; Touboul, D.; Costa, J.; et al. Bioactivity-Based Molecular Networking for the Discovery of Drug Leads in Natural Product Bioassay-Guided Fractionation. J. Nat. Prod. 2018, 81, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Stagliano, M.C.; DeKeyser, J.G.; Omiecinski, C.J.; Jones, A.D. Bioassay-directed fractionation for discovery of bioactive neutral lipids guided by relative mass defect filtering and multiplexed collision-induced dissociation. Rapid Commun. Mass Spectrom. 2010, 24, 3578–3584. [Google Scholar] [CrossRef]
- Abbas-Mohammadi, M.; Farimani, M.M.; Salehi, P.; Ebrahimi, S.N.; Sonboli, A.; Kelso, C.; Skropeta, D. Acetylcholinesterase-inhibitory activity of Iranian plants: Combined HPLC/bioassay-guided fractionation, molecular networking and docking strategies for the dereplication of active compounds. J. Pharm. Biomed. Anal. 2018, 158, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Shine, V.J.; Anuja, G.I.; Suja, S.R.; Raj, G.; Latha, P.G. Bioassay guided fractionation of Cyclea peltata using in vitro RAW 264.7 cell culture, antioxidant assays and isolation of bioactive compound tetrandrine. J. Ayurveda Integr. Med. 2018, 11, 281–286. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Sidebottom, A.M.; Johnson, A.R.; Karty, J.A.; Trader, D.J.; Carlson, E.E. Integrated metabolomics approach facilitates discovery of an unpredicted natural product suite from Streptomyces coelicolor M145. ACS Chem. Biol. 2013, 8, 2009–2016. [Google Scholar] [CrossRef]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.T.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular Networking as a Dereplication Strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Ali, K.; Iqbal, M.; Yuliana, N.D.; Lee, Y.J.; Park, S.; Han, S.; Lee, J.W.; Lee, H.S.; Verpoorte, R.; Choi, Y.H. Identification of bioactive metabolites against adenosine A1 receptor using NMR-based metabolomics. Metabolomics 2013, 9, 778–785. [Google Scholar] [CrossRef]
- Rajalahti, T.; Arneberg, R.; Berven, F.S.; Myhr, K.M.; Ulvik, R.J.; Kvalheim, O.M. Biomarker discovery in mass spectral profiles by means of selectivity ratio plot. Chemom. Intell. Lab. Syst. 2009, 95, 35–48. [Google Scholar] [CrossRef]
- Rajalahti, T.; Arneberg, R.; Kroksveen, A.C.; Berle, M.; Myhr, K.-M.; Kvalheim, O.M. Discriminating Variable Test and Selectivity Ratio Plot: Quantitative Tools for Interpretation and Variable (Biomarker) Selection in Complex Spectral or Chromatographic Profiles. Anal. Chem. 2009, 81, 2581–2590. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, J.J.; Todd, D.A.; Egan, J.M.; Raja, H.A.; Oberlies, N.H.; Kvalheim, O.M.; Cech, N.B. Biochemometrics for Natural Products Research: Comparison of Data Analysis Approaches and Application to Identification of Bioactive Compounds. J. Nat. Prod. 2016, 79, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Hastie, T. Regularization and variable selection via the elastic net. J. R. Stat. Soc. B 2005, 67, 768. [Google Scholar] [CrossRef]
- Brown, K.S.; Jamieson, P.; Wu, W.; Vaswani, A.; Alcazar Magana, A.; Choi, J.; Mattio, L.M.; Cheong, P.H.; Nelson, D.; Reardon, P.N.; et al. Computation-Assisted Identification of Bioactive Compounds in Botanical Extracts: A Case Study of Anti-Inflammatory Natural Products from Hops. Antioxidants 2022, 11, 1400. [Google Scholar] [CrossRef] [PubMed]
- Brinkhaus, B.; Lindner, M.; Schuppan, D.; Hahn, E.G. Chemical, pharmacological and clinical profile of the East Asian medical plant Centella asiatica. Phytomedicine 2000, 7, 427–448. [Google Scholar] [CrossRef]
- Shinomol, G.K.; Muralidhara, M.S.; Bharath, M. Exploring the Role of “Brahmi” (Bacopa monnieri and Centella asiatica) in Brain Function and Therapy. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 33–49. [Google Scholar]
- Gray, N.E.; Alcazar Magana, A.; Lak, P.; Wright, K.M.; Quinn, J.; Stevens, J.F.; Maier, C.S.; Soumyanath, A. Centella asiatica—Phytochemistry and mechanisms of neuroprotection and cognitive enhancement. Phytochem. Rev. 2018, 17, 161–194. [Google Scholar] [CrossRef]
- Kapoor, L.D. CRC Handbook of Ayurvedic Medicinal Plants, 1st ed.; CRC Press: Boca Raton, FL, USA, 1990; p. 416. [Google Scholar]
- Kumar, A.; Dogra, S.; Prakash, A. Neuroprotective Effects of Centella asiatica against Intracerebroventricular Colchicine-Induced Cognitive Impairment and Oxidative Stress. Int. J. Alzheimers Dis. 2009, 2009, 972178. [Google Scholar] [PubMed]
- Sabaragamuwa, R.; Perera, C.O.; Fedrizzi, B. Centella asiatica (Gotu kola) as a neuroprotectant and its potential role in healthy ageing. Trends Food Sci. Technol. 2018, 79, 88–97. [Google Scholar] [CrossRef]
- Lokanathan, Y.; Omar, N.; Ahmad Puzi, N.N.; Saim, A.; Hj Idrus, R. Recent Updates in Neuroprotective and Neuroregenerative Potential of Centella asiatica. Malays. J. Med. Sci. 2016, 23, 4–14. [Google Scholar] [PubMed]
- Kirkpatrick, C.L.; Broberg, C.A.; McCool, E.N.; Lee, W.J.; Chao, A.; McConnell, E.W.; Pritchard, D.A.; Hebert, M.; Fleeman, R.; Adams, J.; et al. The “PepSAVI-MS” Pipeline for Natural Product Bioactive Peptide Discovery. Anal. Chem. 2017, 89, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Clark, T.N.; Houriet, J.; Vidar, W.S.; Kellogg, J.J.; Todd, D.A.; Cech, N.B.; Linington, R.G. Interlaboratory Comparison of Untargeted Mass Spectrometry Data Uncovers Underlying Causes for Variability. J. Nat. Prod. 2021, 84, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Gray, N.E.; Morré, J.; Kelley, J.; Maier, C.S.; Stevens, J.F.; Quinn, J.F.; Soumyanath, A. Caffeoylquinic acids in Centella asiatica protect against amyloid-β toxicity. J. Alzheimers Dis. 2014, 40, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef] [PubMed]
- Magana, A.A.; Wright, K.; Vaswani, A.; Caruso, M.; Reed, R.L.; Bailey, C.F.; Nguyen, T.; Gray, N.E.; Soumyanath, A.; Quinn, J.; et al. Integration of mass spectral fingerprinting analysis with precursor ion (MS1) quantification for the characterisation of botanical extracts: Application to extracts of Centella asiatica (L.) Urban. Phytochem. Anal. 2020, 31, 722–738. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134 (Suppl. 12), 3479s–3485s. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Liang, N.; Dupuis, J.H.; Yada, R.Y.; Kitts, D.D. Chlorogenic acid isomers directly interact with Keap 1-Nrf2 signaling in Caco-2 cells. Mol. Cell. Biochem. 2019, 457, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Miyamae, Y.; Shigemori, H.; Isoda, H. Neuroprotective effect of 3,5-di-O-caffeoylquinic acid on SH-SY5Y cells and senescence-accelerated-prone mice 8 through the up-regulation of phosphoglycerate kinase-1. Neuroscience 2010, 169, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jiang, X.-W.; Yang, Y.; Liu, W.-W.; Xu, Z.-H.; Zhao, Q.-C. Isolation, structure elucidation and neuroprotective effects of caffeoylquinic acid derivatives from the roots of Arctium lappa L. Phytochemistry 2020, 177, 112432. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Lee, H.; Nam, T.G.; Eom, S.H.; Heo, H.J.; Lee, C.Y.; Kim, D.O. Neuroprotective effect of caffeoylquinic acids from Artemisia princeps Pampanini against oxidative stress-induced toxicity in PC-12 cells. J. Food Sci. 2011, 76, C250–C256. [Google Scholar] [CrossRef] [PubMed]
- Liberato, J.L.; Rosa, M.N.; Miranda, M.C.R.; Lopes, J.L.C.; Lopes, N.P.; Gobbo-Neto, L.; Fontana, A.C.K.; Dos Santos, W.F. Neuroprotective Properties of Chlorogenic Acid and 4,5-Caffeoylquinic Acid from Brazilian arnica (Lychnophora ericoides) after Acute Retinal Ischemia. Planta Med. 2023, 89, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.Y.D.; Dobrachinski, F.; Silva, H.B.; Lopes, J.P.; Gonçalves, F.Q.; Soares, F.A.A.; Porciúncula, L.O.; Andrade, G.M.; Cunha, R.A.; Tomé, A.R. Neuromodulation and neuroprotective effects of chlorogenic acids in excitatory synapses of mouse hippocampal slices. Sci. Rep. 2021, 11, 10488. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.G.; Caruso, M.; Alcazar Magana, A.; Wright, K.M.; Maier, C.S.; Stevens, J.F.; Gray, N.E.; Quinn, J.F.; Soumyanath, A. Caffeoylquinic Acids in Centella asiatica Reverse Cognitive Deficits in Male 5XFAD Alzheimer’s Disease Model Mice. Nutrients 2020, 12, 3488. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.-C.; Tsai, T.-Y.; Wang, C.-J. The Potential Benefits of Quercetin for Brain Health: A Review of Anti-Inflammatory and Neuroprotective Mechanisms. Int. J. Mol. Sci. 2023, 24, 6328. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Aschner, M.; Cheang, W.S.; Akkol, E.K. Neuroprotective Effects of Quercetin in Alzheimer’s Disease. Biomolecules 2019, 10, 59. [Google Scholar] [CrossRef]
- Ma, Z.-X.; Zhang, R.-Y.; Rui, W.-J.; Wang, Z.-Q.; Feng, X. Quercetin alleviates chronic unpredictable mild stress-induced depressive-like behaviors by promoting adult hippocampal neurogenesis via FoxG1/CREB/ BDNF signaling pathway. Behav. Brain Res. 2021, 406, 113245. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Sopher, B.L.; Fukuchi, K.; Kavanagh, T.J.; Furlong, C.E.; Martin, G.M. Neurodegenerative mechanisms in Alzheimer disease—A role for oxidative damage in amyloid beta protein precursor-mediated cell death. Mol. Chem. Neuropathol. 1996, 29, 153–168. [Google Scholar] [CrossRef]
- Woltjer, R.L.; McMahan, W.; Milatovic, D.; Kjerulf, J.D.; Shie, F.S.; Rung, L.G.; Montine, K.S.; Montine, T.J. Effects of chemical chaperones on oxidative stress and detergent-insoluble species formation following conditional expression of amyloid precursor protein carboxy-terminal fragment. Neurobiol. Dis. 2007, 25, 427–437. [Google Scholar] [CrossRef]
- Woltjer, R.L.; Maezawa, I.; Ou, J.J.; Montine, K.S.; Montine, T.J. Advanced glycation endproduct precursor alters intracellular amyloid- beta/A beta PP carboxy-terminal fragment aggregation and cytotoxicity. J. Alzheimers Dis. 2003, 5, 467–476. [Google Scholar] [CrossRef]
- Kucheryayskiy, S. mdatools—R package for chemometrics. Chemom. Intell. Lab. Syst. 2020, 198, 103937. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
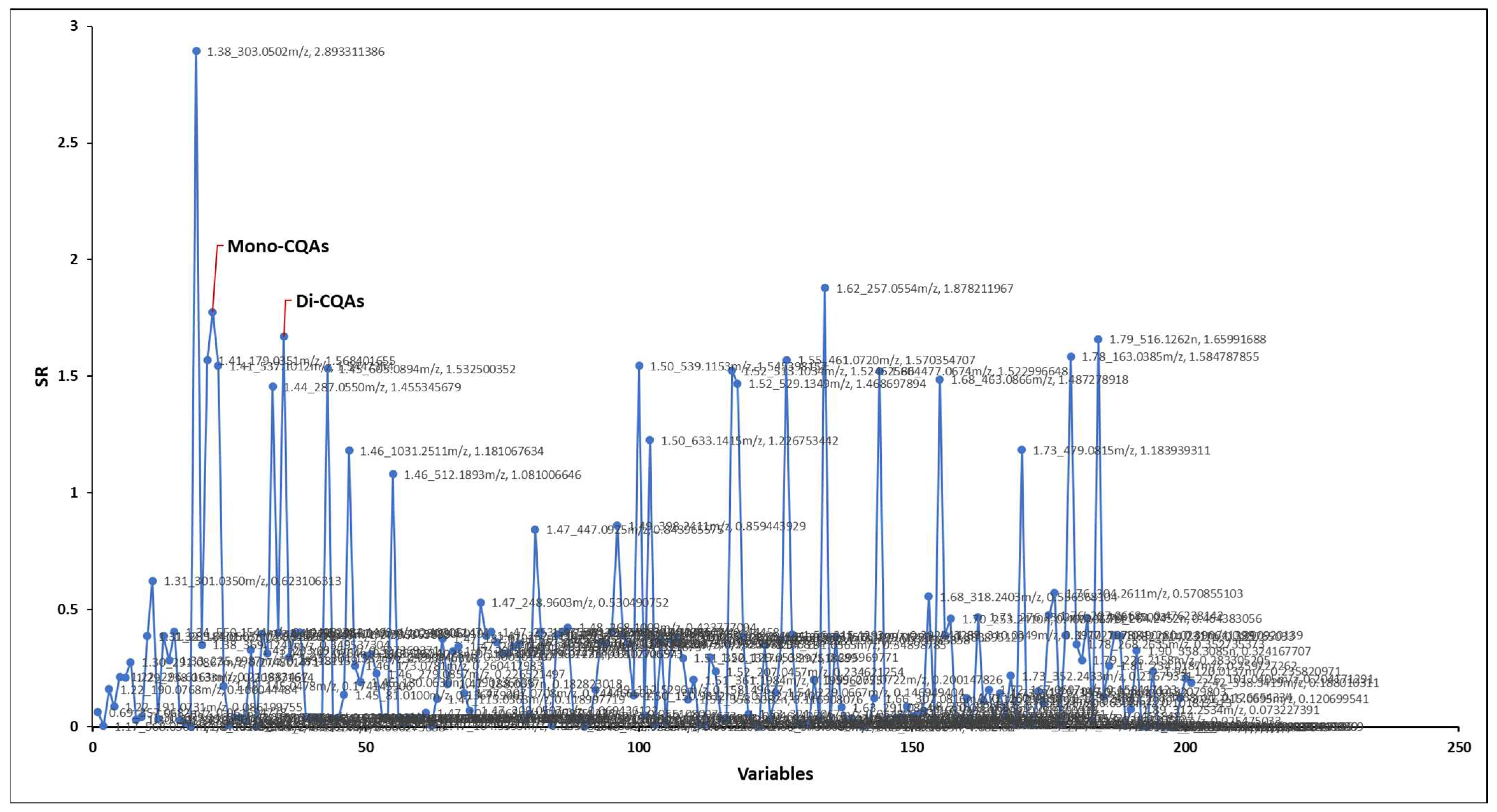
| Feature 1 | SR 2 | Variable Rank in Elastic Net Pipeline 3 | Annotation 4 | Ion Mode |
|---|---|---|---|---|
| 1.38_303.0502 m/z | 2.89 | 47 (of 119) | Quercetin | POS |
| 1.62_257.0554 m/z | 1.88 | 9 (of 85) | N/A | NEG |
| 1.41_353.0874 m/z | 1.78 | 10 (of 85) | Mono-CQAs | NEG |
| 1.79_515.1191 m/z | 1.66 | 1 (of 85) | Di-CQA’s | NEG |
| 1.78_163.0385 m/z | 1.58 | 23 (of 119) | Hydroxycoumarin | POS |
| 1.55_461.0720 m/z | 1.57 | 47 (of 85) | Myricetin 3-glucoside | NEG |
| 1.41_179.0351 m/z | 1.57 | 29 (of 85) | Caffeic Acid | NEG |
| 1.50_539.1153 m/z | 1.55 | 18 (of 119) | N/A | POS |
| 1.41_537.1012 m/z | 1.54 | 44 (of 85) | N/A | NEG |
| 1.45_605.0894 m/z | 1.53 | 45 (of 85) | N/A | NEG |
| 1.52_513.1034 m/z | 1.52 | 34 (of 85) | N/A | NEG |
| 1.66_477.0674 m/z | 1.52 | 50 (of 85) | Quercetin 7-glucuronide | NEG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcázar Magaña, A.; Vaswani, A.; Brown, K.S.; Jiang, Y.; Alam, M.N.; Caruso, M.; Lak, P.; Cheong, P.; Gray, N.E.; Quinn, J.F.; et al. Integrating High-Resolution Mass Spectral Data, Bioassays and Computational Models to Annotate Bioactives in Botanical Extracts: Case Study Analysis of C. asiatica Extract Associates Dicaffeoylquinic Acids with Protection against Amyloid-β Toxicity. Molecules 2024, 29, 838. https://doi.org/10.3390/molecules29040838
Alcázar Magaña A, Vaswani A, Brown KS, Jiang Y, Alam MN, Caruso M, Lak P, Cheong P, Gray NE, Quinn JF, et al. Integrating High-Resolution Mass Spectral Data, Bioassays and Computational Models to Annotate Bioactives in Botanical Extracts: Case Study Analysis of C. asiatica Extract Associates Dicaffeoylquinic Acids with Protection against Amyloid-β Toxicity. Molecules. 2024; 29(4):838. https://doi.org/10.3390/molecules29040838
Chicago/Turabian StyleAlcázar Magaña, Armando, Ashish Vaswani, Kevin S. Brown, Yuan Jiang, Md Nure Alam, Maya Caruso, Parnian Lak, Paul Cheong, Nora E. Gray, Joseph F. Quinn, and et al. 2024. "Integrating High-Resolution Mass Spectral Data, Bioassays and Computational Models to Annotate Bioactives in Botanical Extracts: Case Study Analysis of C. asiatica Extract Associates Dicaffeoylquinic Acids with Protection against Amyloid-β Toxicity" Molecules 29, no. 4: 838. https://doi.org/10.3390/molecules29040838
APA StyleAlcázar Magaña, A., Vaswani, A., Brown, K. S., Jiang, Y., Alam, M. N., Caruso, M., Lak, P., Cheong, P., Gray, N. E., Quinn, J. F., Soumyanath, A., Stevens, J. F., & Maier, C. S. (2024). Integrating High-Resolution Mass Spectral Data, Bioassays and Computational Models to Annotate Bioactives in Botanical Extracts: Case Study Analysis of C. asiatica Extract Associates Dicaffeoylquinic Acids with Protection against Amyloid-β Toxicity. Molecules, 29(4), 838. https://doi.org/10.3390/molecules29040838