
Phytochemical Profile and In Vitro Bioactivities of Wild Asparagus stipularis

 , ,
, ,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Analysis of Simple Phenolics
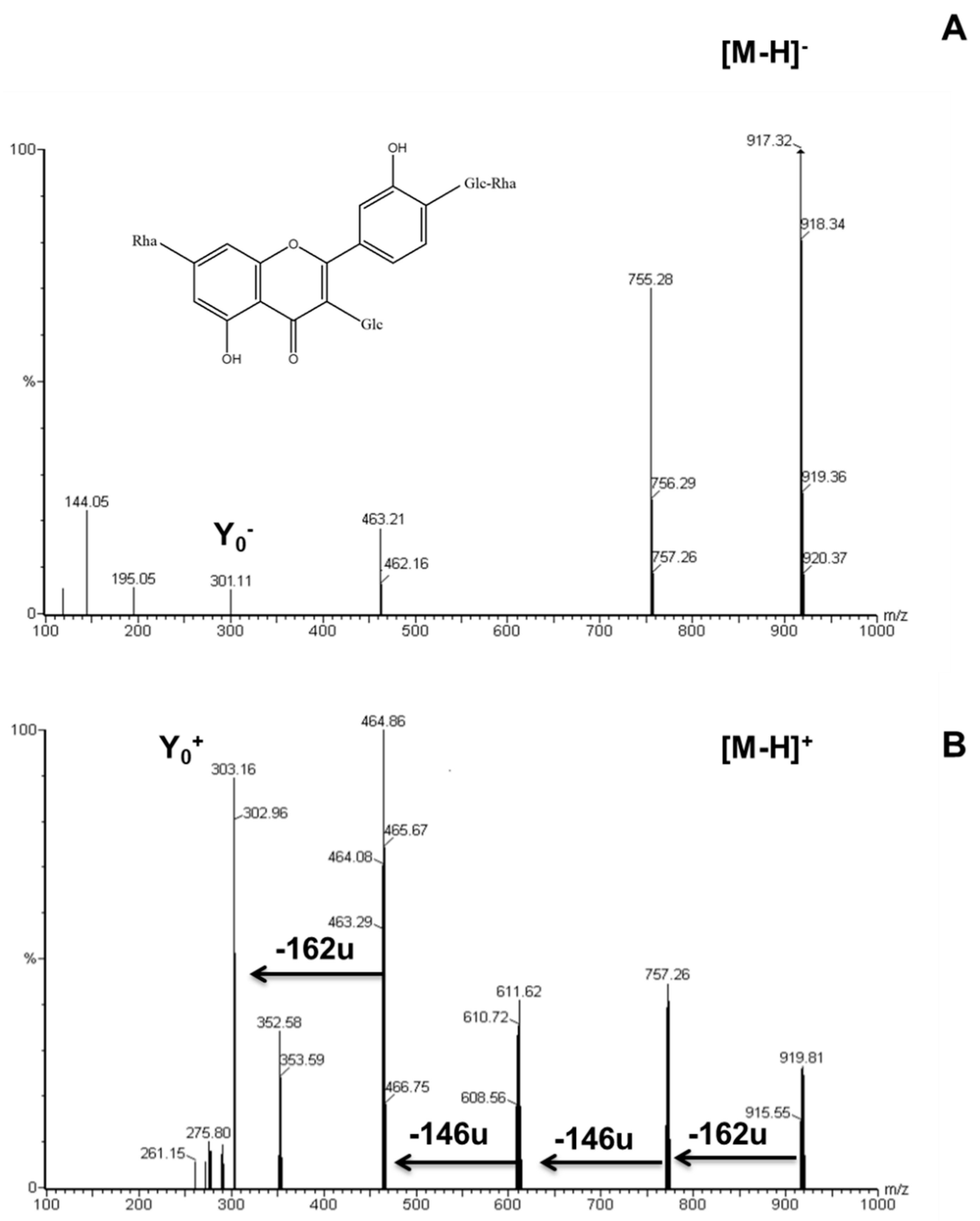
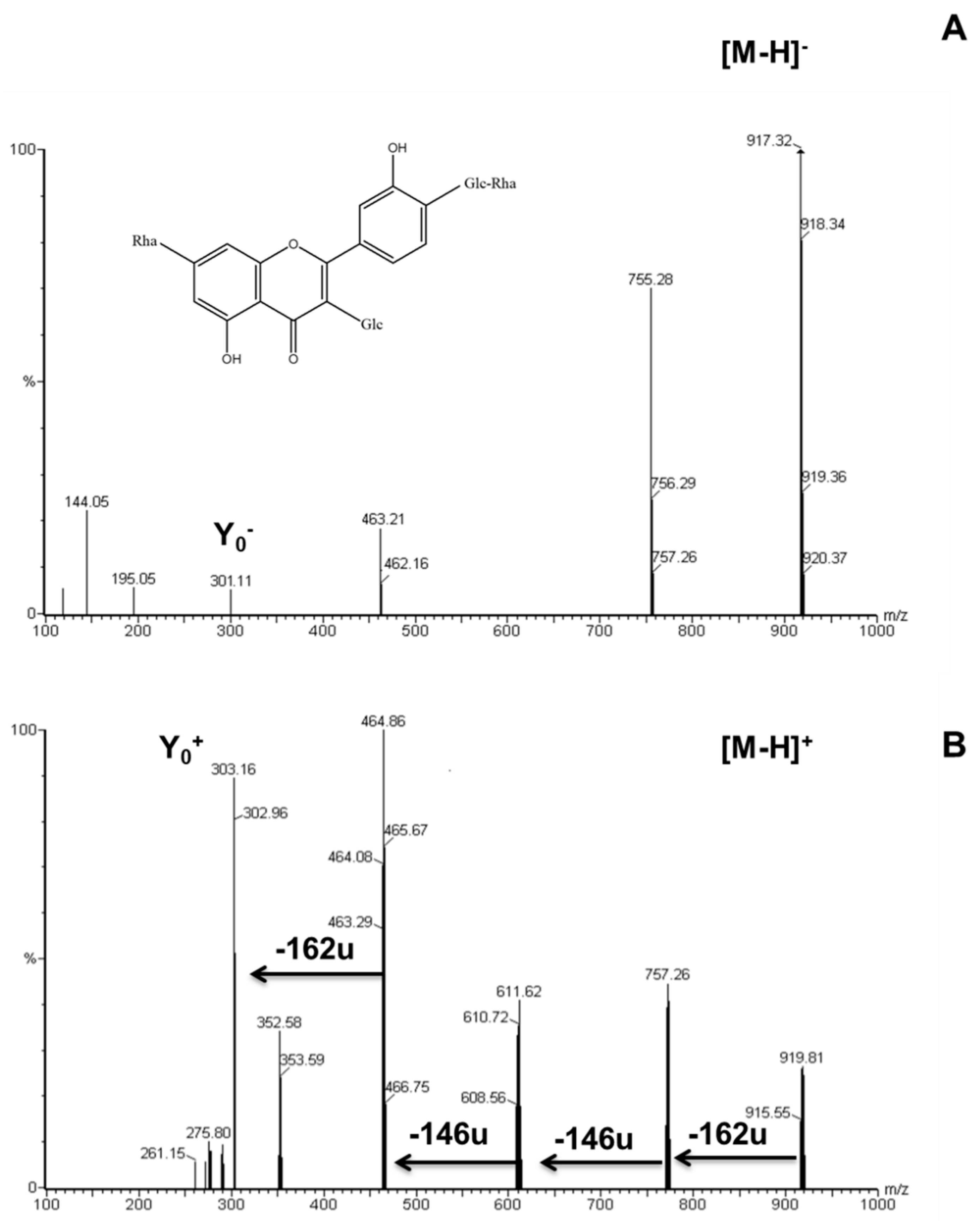
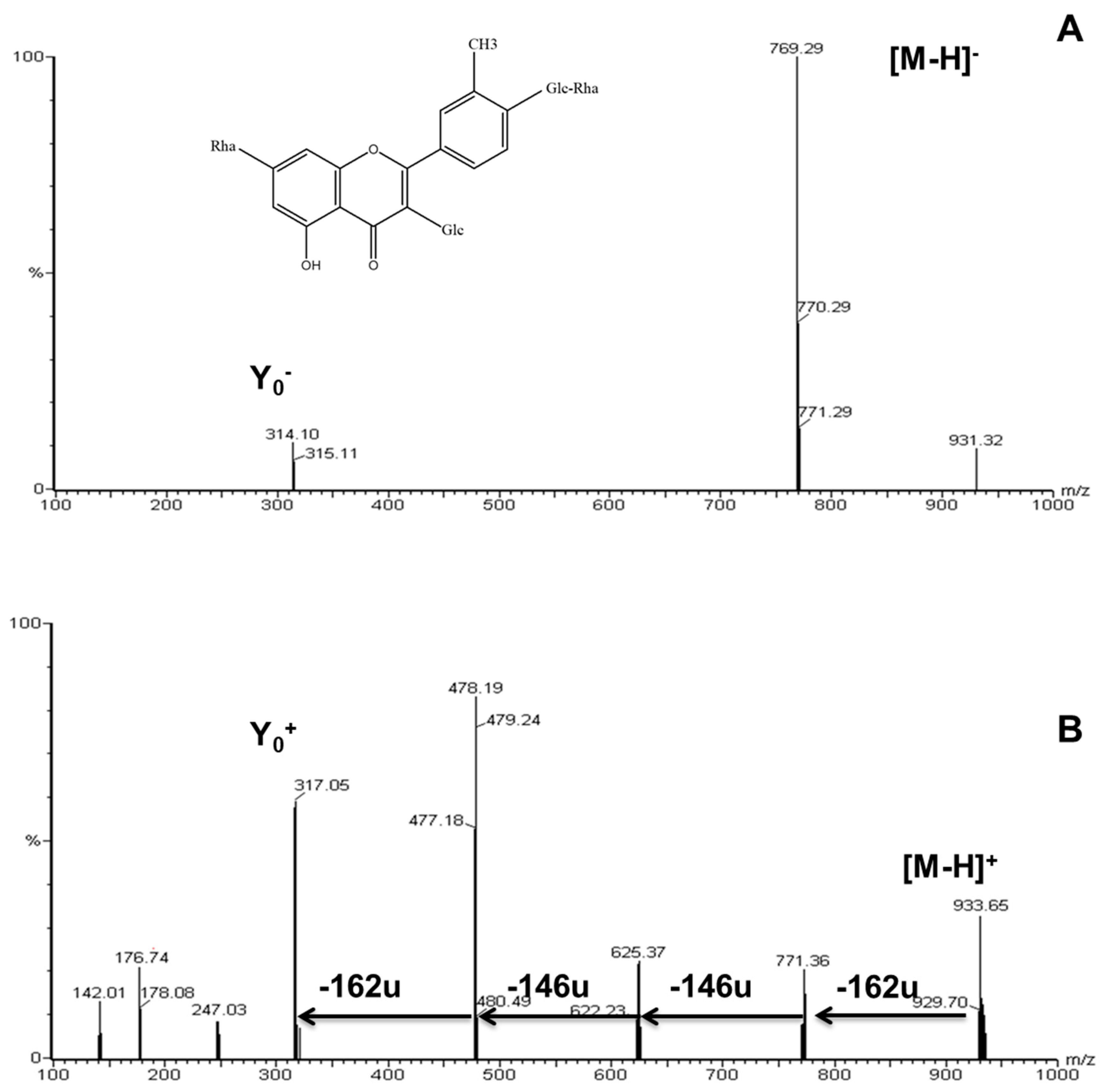
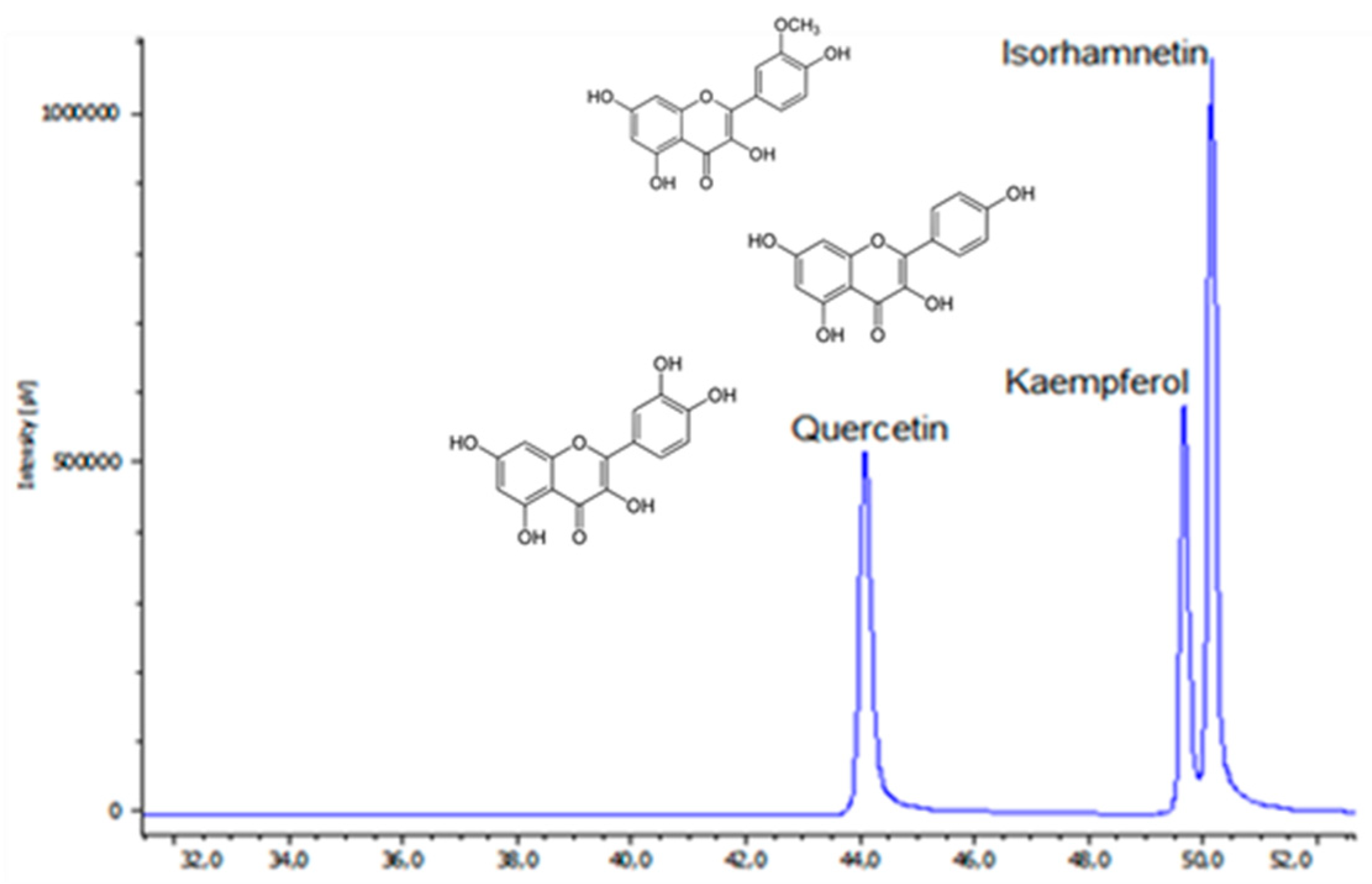
2.2. Identification of Flavonoid Glycosides in Different Asparagus Parts
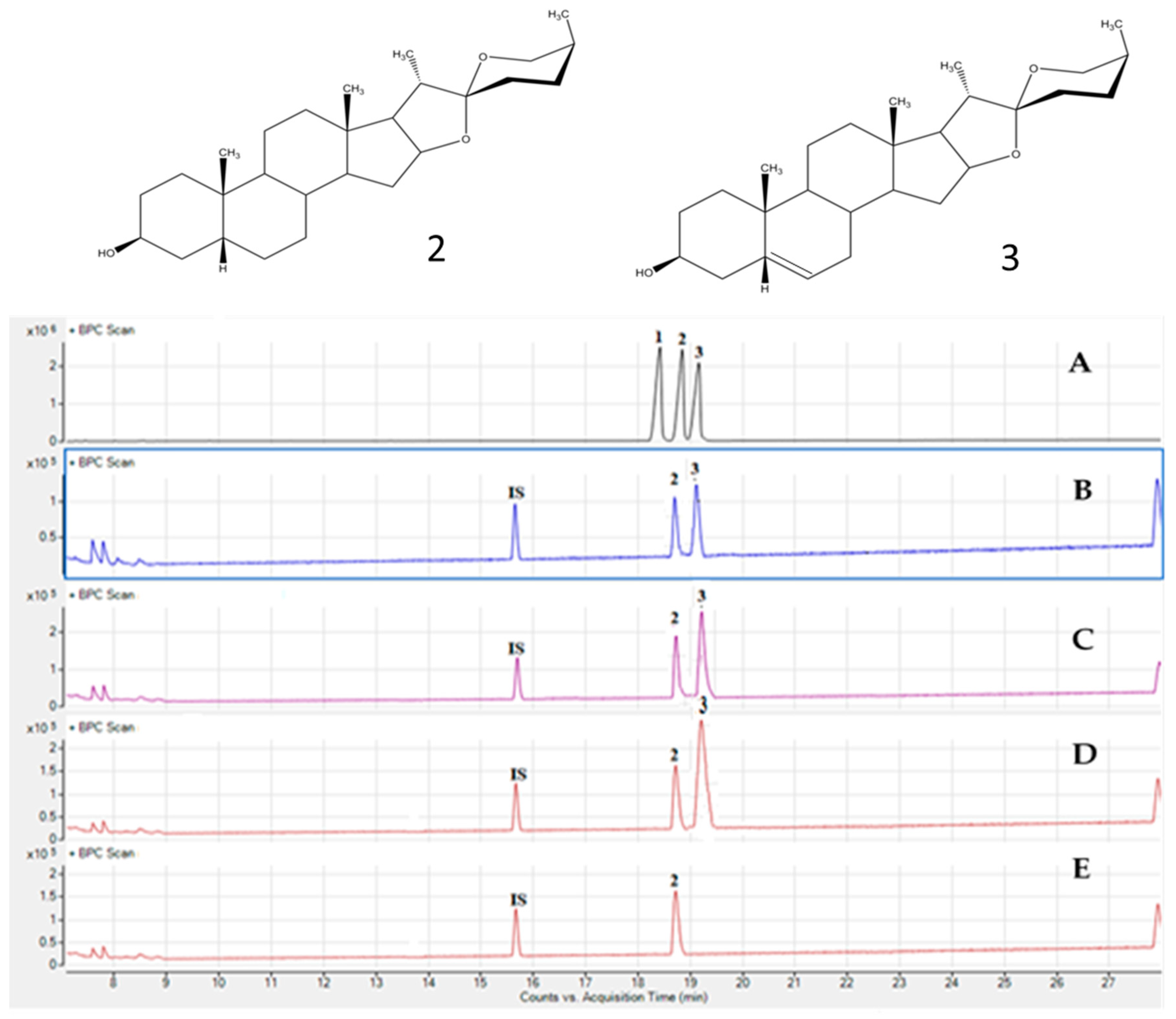
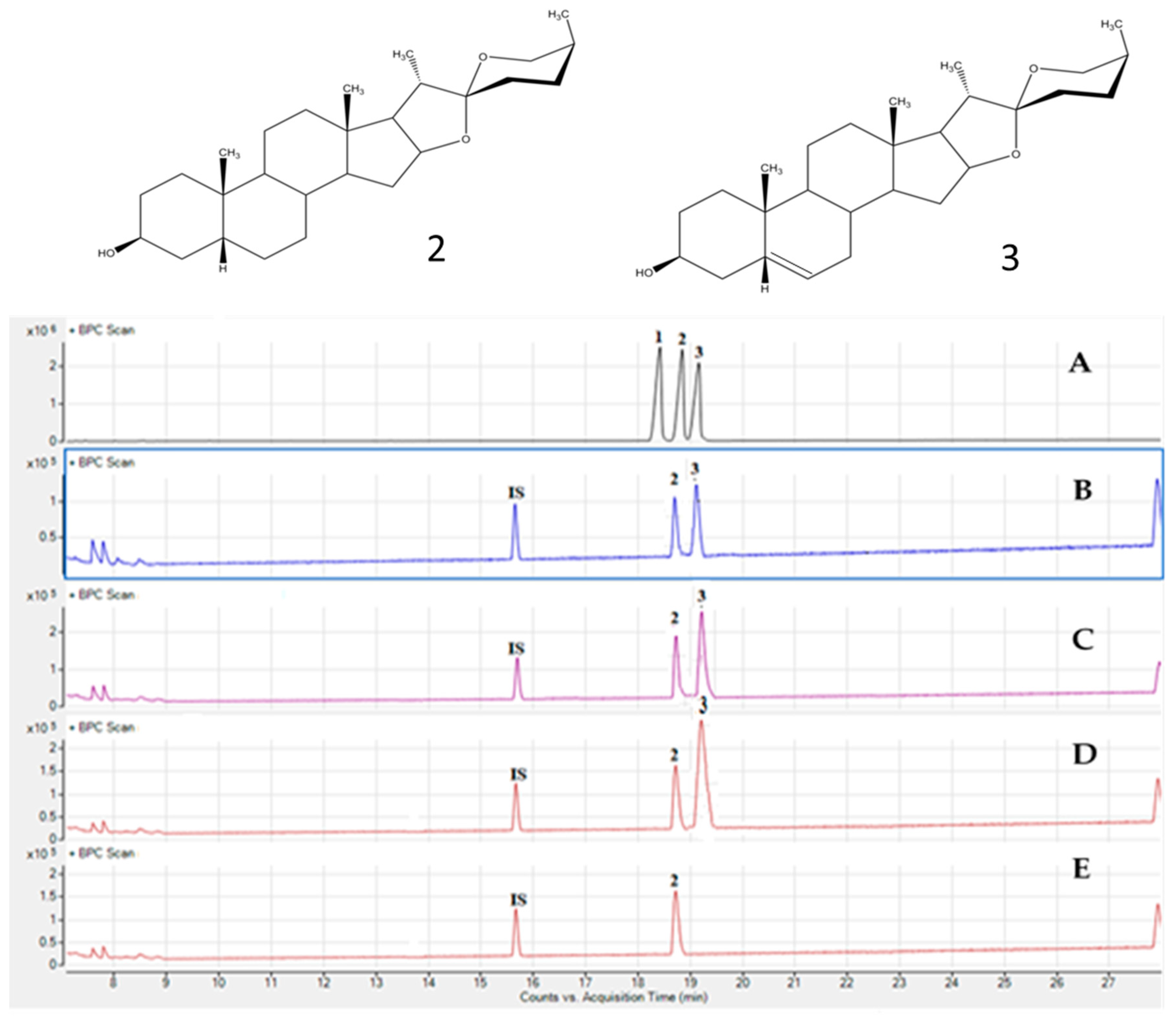
2.3. Analysis of Saponins in Asparagus stipularis Samples
2.4. Quantification of Saponins in Asparagus stipularis Samples
2.5. Determination of Antioxidant Activities
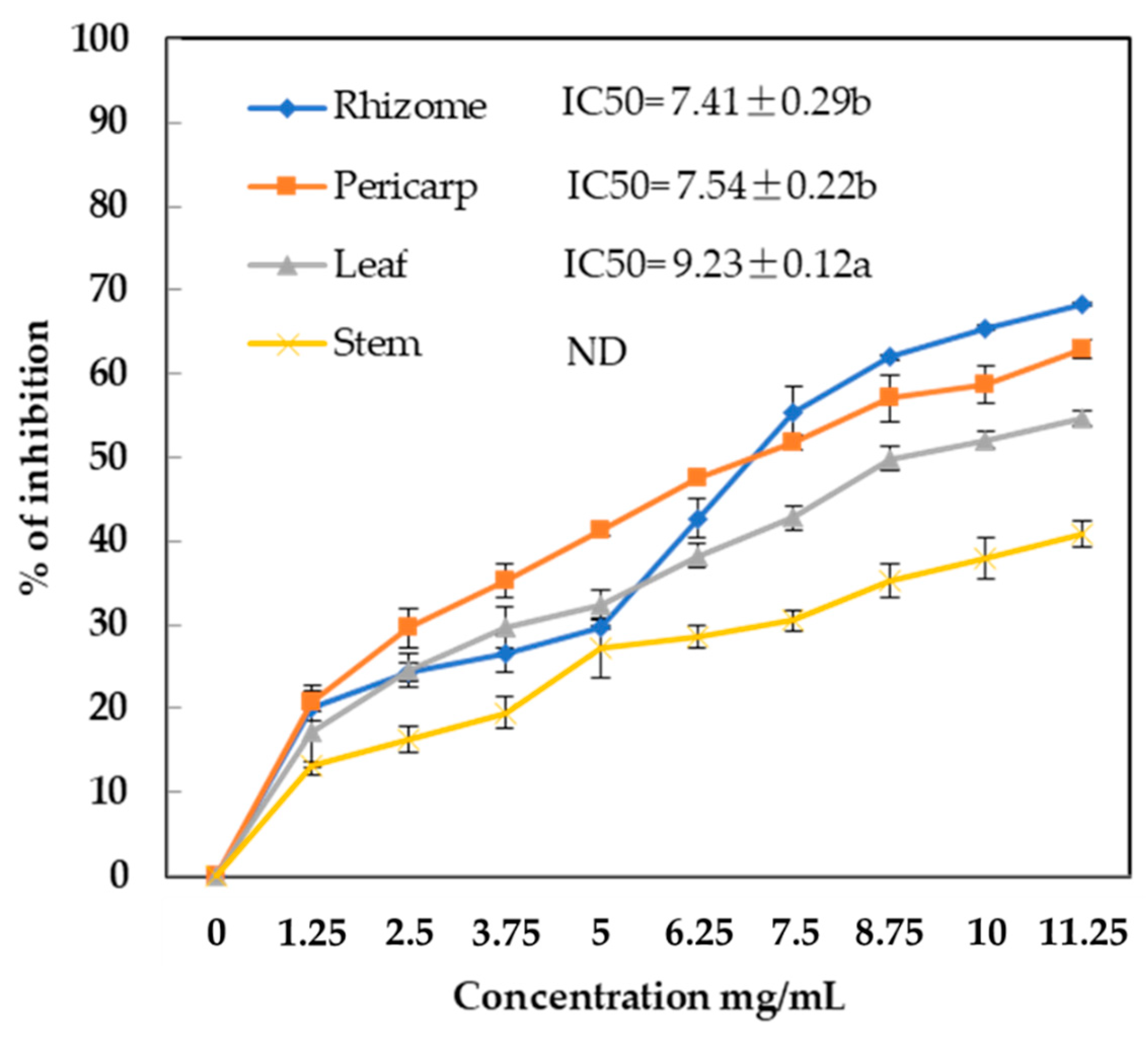
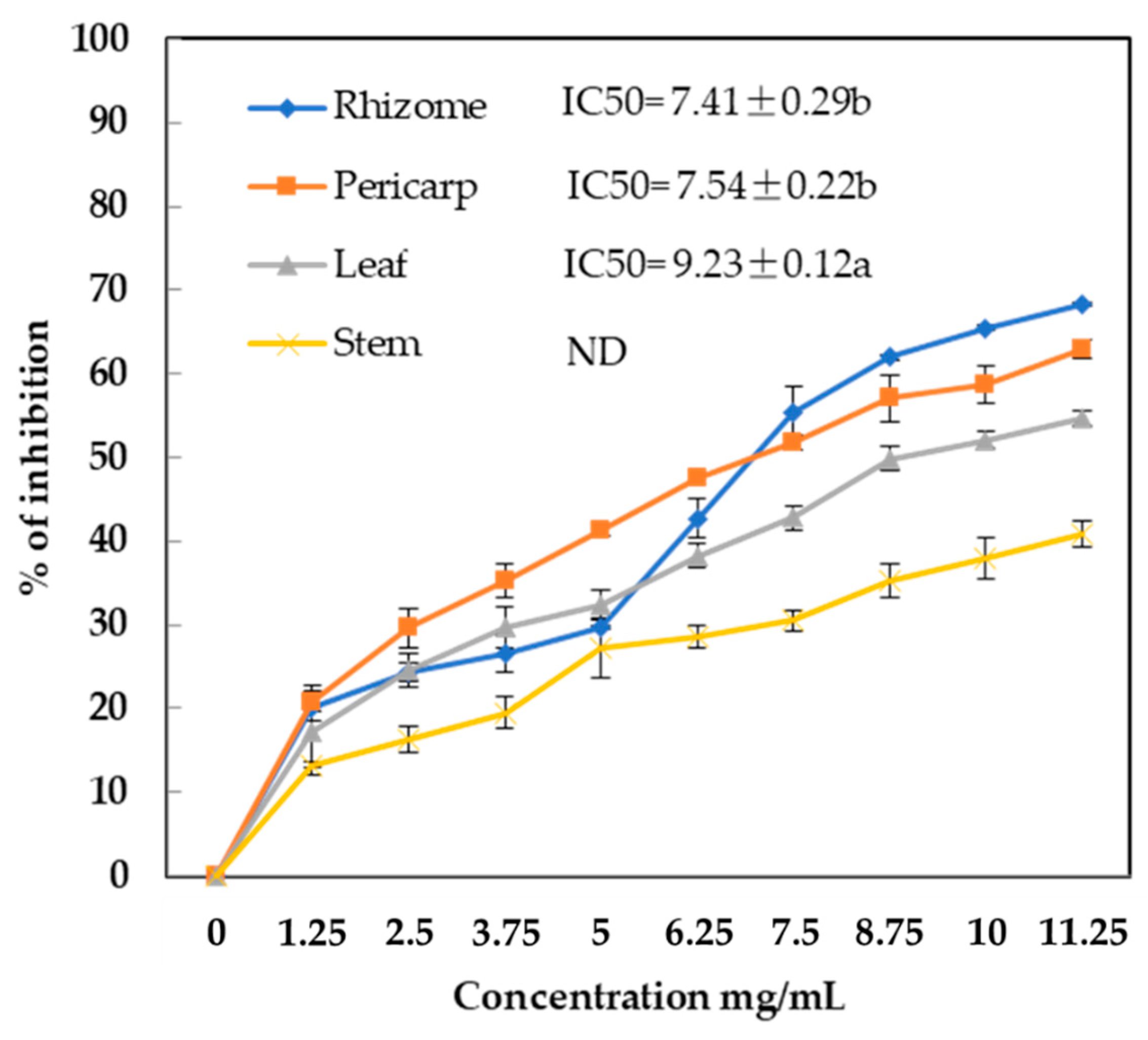
2.6. Lipase Inhibitory Activity
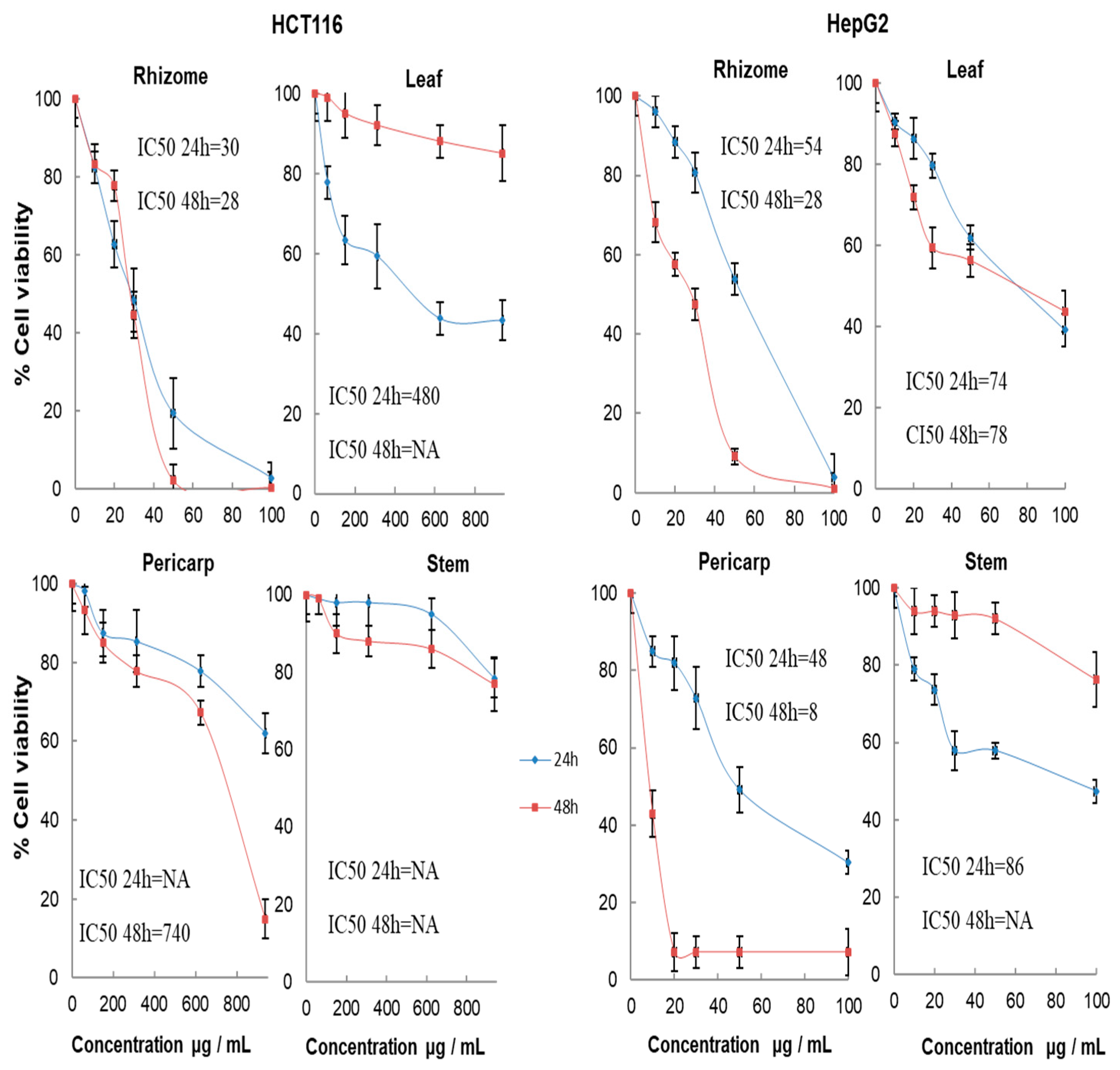
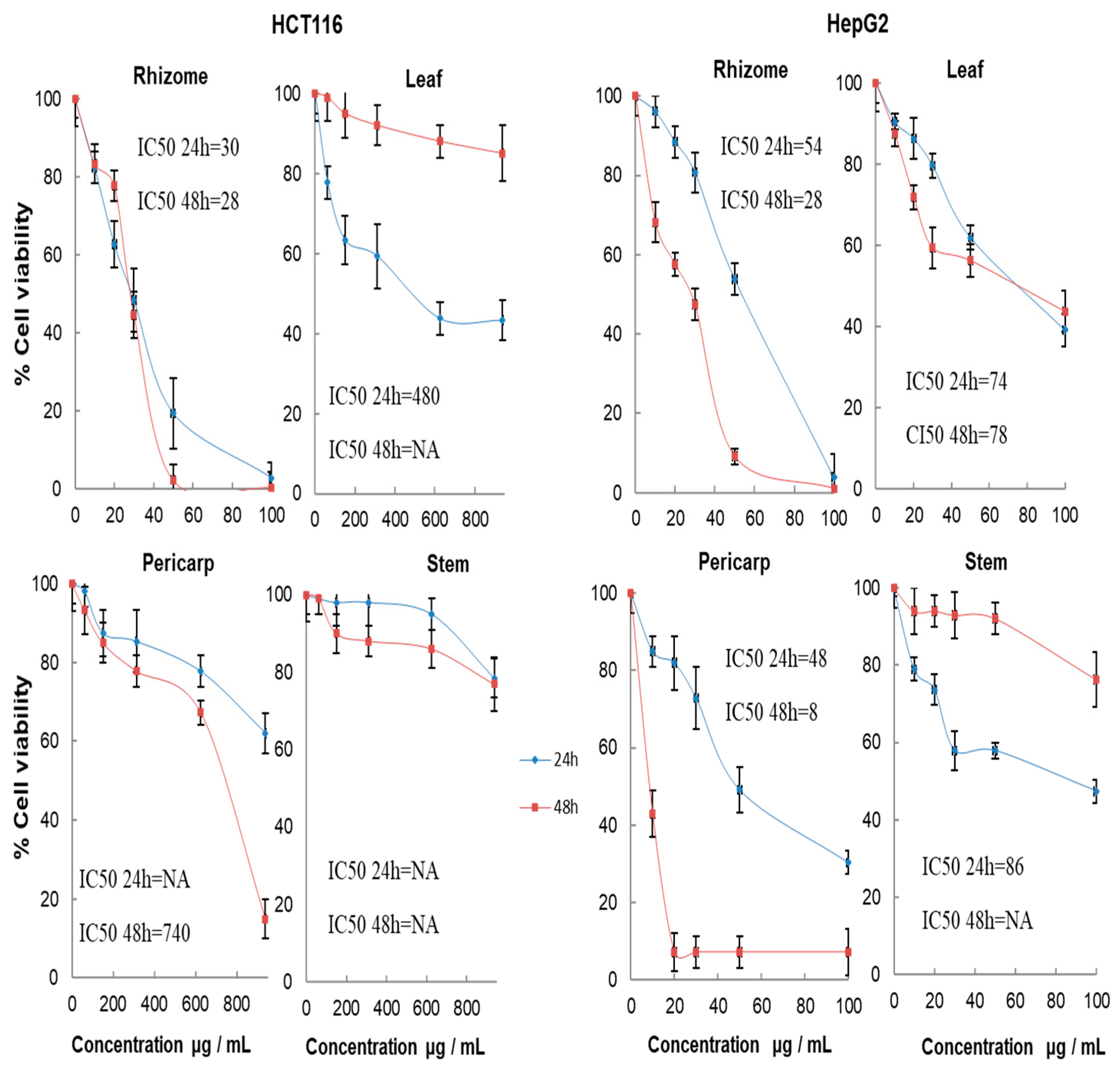
2.7. Cytotoxic Activity
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Preparation of Ethanolic Extract
3.4. Extraction of Phenolic Compounds
3.5. Analysis of Free Phenolics Composition by HPLC/MS
3.6. Analysis and Quantification of Flavonoids by HPLC–DAD
3.7. Acid Hydrolysis of Flavonoid Glycosides
3.8. Analysis of Saponins
3.8.1. Characterization and Quantification of Saponins by LC-MS
3.8.2. Determination of Exact Mass
3.8.3. Preparation of Saponin Extract and Purification of Individual Saponins
3.8.4. Hydrolysis of Saponins
3.8.5. Analysis of Saponin Glycosidic Moiety
3.9. Determination of the Antioxidant Capacity
3.10. Pancreatic Lipase Inhibitory Activity Assay In Vitro
3.11. Cytotoxicity against Cancer Cell Lines
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manandhar, N.P. Medicinal Plant-lore of Tamang Tribe of Kabhrepalanchok District, Nepal. Econ. Bot. 1991, 45, 58–71. [Google Scholar] [CrossRef]
- Kamboj, V.P. Herbal Medicine. Curr. Sci. 2000, 78, 35–39. [Google Scholar]
- He, C.Y.; Hsiang, T.; Wolyn, D.J. Induction of systemic disease resistance and pathogen defence responses in Asparagus officinalis by nonpathogenic strains of Fusarium oxysporum. Plant Pathol. 2002, 51, 225–230. [Google Scholar] [CrossRef]
- Ferrara, L.; Dosi, R.; Di Maro, A.; Guida, V.; Cafarelli, G.; Pacifico, S.; Mastellone, C.; Fiorentino, A.; Rosati, A.; Parente, A. Nutritional values, metabolic profile and radical scavenging capacities of wild asparagus (A. acutifolius L.). J. Food Compos. Anal. 2011, 24, 326–333. [Google Scholar] [CrossRef]
- Hamdi, A.; Jaramillo-Carmona, S.; Srairi Beji, R.; Tej, R.; Zaoui, S.; Rodríguez-Arco, R.; Jiménez-Araujo, A.; Kasri, M.; Lachaal, M.; Karray Bouraoui, N.; et al. The phytochemical and bioactivity profiles of wild Asparagus albus L. plant. Food Res. Int. 2017, 99, 720–729. [Google Scholar] [CrossRef]
- Adouni, K.; Chahdoura, H.; Mosbah, H.; Santos-Buelga, C.; González-Paramás, A.M.; Ciudad-Mulero, M.; Achour, L. Revalorization of wild Asparagus stipularis Forssk. as a traditional vegetable with nutritional and functional properties. Food Funct. 2018, 9, 1578–1586. [Google Scholar] [CrossRef]
- Sautour, M.; Miyamoto, T.; Lacaille-Dubois, M.A. Steroidal saponins from Asparagus acutifolius. Phytochemistry 2007, 68, 2554–2562. [Google Scholar] [CrossRef]
- Fuentes-Alventosa, J.M.; Jaramillo-Carmona, S.; Rodríguez-Gutiérrez, G.; Rodríguez-Arcos, R.; Fernández-Bolaños, J.; Guillén-Bejarano, R.; Jiménez-Araujo, A. Effect of the extraction method on phytochemical composition and antioxidant activity of high dietary fibre powders obtained from asparagus by-products. Food Chem. 2009, 116, 484–490. [Google Scholar] [CrossRef]
- Hossain, M.I.; Sharmin, F.A.; Akhter, S.; Bhuiyan, M.A.; Shahriar, M. Investigation of cytotoxicity and in-vitro antioxidant activity of Asparagus racemosus root extract. Int. Curr. Pharm. J. 2012, 1, 250–257. [Google Scholar] [CrossRef]
- Agrawal, A.; Sharma, M.; Rai, S.K.; Singh, B.; Tiwari, M.; Chandra, R. The effect of the aqueous extract of the roots of Asparagus racemosus on hepatocarcinogenesis initiated by diethylnitrosamine. Phytother. Res. 2008, 22, 1175–1182. [Google Scholar] [CrossRef]
- Dahanukar, S.; Thatte, U.; Pai, N.; Mose, P.; Karandikar, S. Protective effect of Asparagus racemosus against induced abdominal sepsis. Indian Drugs 1986, 24, 125–128. [Google Scholar]
- Ulukapi, K.; Nasircilar, A.G.; Onus, A.N.; Baktir, I. In vitro propagation and determination of the nutrient content of naturally grown Asparagus stipularis, Forssk. Arch. Biol. Sci. 2014, 66, 1333–1338. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Shabasy, R.; Sakr, H.; Zayed, M.; El-Said, A.M.A.; Helmy, K.M.H.; Göransson, U. Anti-schistosomiasis triterpene glycoside from the Egyptian medicinal plant Asparagus stipularis. Rev. Bras. Farmacogn. 2012, 22, 314–318. [Google Scholar] [CrossRef]
- Boulos, L. Medicinal Plants of North Africa. Chin. Med. 1983, 6, 286. [Google Scholar]
- Galala, A.A.; Ahmad, K.F.; Zaghloul, M.G.; Mansour, E.S. Two new alkaloids from Asparagus stipularis Forssk roots. Phytochem. Lett. 2015, 12, 220–223. [Google Scholar] [CrossRef]
- Truong, V.D.; McFeeters, R.F.; Thompson, R.T.; Dean, L.L.; Shofran, B. Phenolic acid content and composition in leaves and roots of common commercial sweetpotato (Ipomea batatas L.) cultivars in the United States. J. Food Sci. 2007, 72, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Mamphiswana, N.D.; Mashela, P.W.; Mdee, L.K. Distribution of total phenolics and antioxidant activity in fruit, leaf, stem and root of Monsonia burkeana. Afr. J. Agric. Res. 2010, 5, 2570–2575. [Google Scholar]
- Daniels, C.W.; Rautenbach, F.; Mabusela, W.; Valentine, A.J.; Marnewick, J.L. Comparative antioxidant-capacity and content of leaves, bulbs, roots, flowers and fruit of Gethyllis multifolia L. Bolus and G. villosa Thunb species. S. Afr. J. Bot. 2011, 77, 711–717. [Google Scholar] [CrossRef]
- Chung, I.M.; Lim, J.J.; Ahn, M.S.; Jeong, H.N.; An, T.J.; Kim, S.H. Comparative phenolic compound profiles and antioxidative activity of the fruit, leaves, and roots of Korean ginseng (Panax ginseng Meyer) according to cultivation years. J. Ginseng Res. 2016, 40, 68–75. [Google Scholar] [CrossRef]
- Ayan, A.K.C.; Irak, C.; Kevseroğlu, K.; Mzen, T. Hypericin in some Hypericum species from Turkey. Asian J. Plant. Sci. 2004, 3, 200–202. [Google Scholar] [CrossRef]
- Achakzai, A.K.K.; Achakzai, P.; Masood, A.; Kayan, I.S.A.; Tareen, R.B. Response of plant parts and age on the distribution of secondary metabolites on plants found in Quetta. Pak. J. Bot. 2009, 41, 2129–2135. [Google Scholar]
- Fuentes-Alventosa, J.M.; Rodríguez, G.; Cermeño-Sacristán, P.; Jiménez, A.; Guillén, R.; Fernández-Bolaños, J.; Rodríguez-Arcos, R. Identification of flavonoid diglycosides in several genotypes of asparagus from the Huétor Tájar population variety. J. Agric. Food Chem. 2007, 55, 10028–10035. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Alventosa, J.M.; Jaramillo, S.; Rodríguez-Gutiérrez, G.; Cermeño, P.; Espejo, J.A.; Jiménez-Araujo, A.; Rodríguez-Arcos, R. Flavonoid profile of green asparagus genotypes. J. Agric. Food Chem. 2008, 56, 6977–6984. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, S.; Lopez, S.; Varela, L.M.; Rodriguez-Arcos, R.; Jimenez, A.; Abia, R.; Guillen, R.G.; Muriana, F.J. The flavonol isorhamnetin exhibits cytotoxic effects on human colon cancer cells. J. Agric. Food Chem. 2010, 58, 10869–10875. [Google Scholar] [CrossRef] [PubMed]
- Manguro, L.O.A.; Ogweno Midiwo, J.; Kraus, W. A New Flavonol tetraglycoside from Myrsine africana leaves. Nat. Prod. Lett. 1996, 9, 121–126. [Google Scholar] [CrossRef]
- Krasteva, I.; Bratkov, V.; Bucar, F.; Kunert, O.; Kollroser, M.; Kondeva-Burdina, M.; Ionkova, I. Flavoalkaloids and flavonoids from Astragalus monspessulanus. J. Nat. Prod. 2015, 78, 2565–2571. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.; Mackenzie, S. Why are all colour combinations not equally represented as flower-colour polymorphisms? New Phytol. 2001, 151, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Streisfeld, M.A.; Joshua, R.; Kohn, J.R. Contrasting patterns of floral and molecular variation across a cline in Mimulus aurantiacus. Evolution 2005, 59, 2548–2559. [Google Scholar]
- Dick, C.A.; Buenrostro, J.; Butler, T.; Carlson, M.L.; Kliebenstein, D.J.; Whittall, J.B. Arctic mustard flower color polymorphism controlled by petal-specific downregulation at the threshold of the anthocyanin biosynthetic pathway. PLoS ONE 2011, 6, e18230. [Google Scholar] [CrossRef]
- Jaramillo-Carmona, S.; Rodriguez-Arcos, R.; Jiménez-Araujo, A.; López, S.; Gil, J.; Moreno, R.; Guillén-Bejarano, R. Saponin profile of wild Asparagus species. J. Food Sci. 2017, 82, 638–646. [Google Scholar] [CrossRef]
- Vázquez-Castilla, S.; Jaramillo-Carmona, S.; Fuentes-Alventosa, J.M.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; Cermeño-Sacristán, P.; Espejo-Calvo, J.A.; Guillén-Bejarano, R. Optimization of a method for the profiling and quantification of saponins in different green asparagus genotypes. J. Agric. Food Chem. 2013, 61, 6250–6258. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Yuan, X.; Zhao, B. Simultaneous analysis of diosgenin and sarsasapogenin in Asparagus officinalis byproduct by thin-layer chromatography. Phytochem. Anal. 2011, 22, 14–17. [Google Scholar] [CrossRef]
- Huang, X.; Kong, L. Steroidal saponins from roots of Asparagus officinalis. Steroids 2006, 71, 171–176. [Google Scholar] [CrossRef]
- Zhou, L.B.; Chen, D.F. Steroidal saponins from the roots of Asparagus filicinus. Steroids 2008, 73, 83–87. [Google Scholar] [CrossRef]
- Karuna, D.S.; Dey, P.; Das, S.; Kundu, A.; Bhakta, T. In vitro antioxidant activities of root extract of Asparagus racemosus Linn. J. Tradit. Complement. Med. 2018, 8, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Symes, A.; Shavandi, A.; Zhang, H.; Mohamed Ahmed, I.; Al-Juhaimi, F.; Bekhit, A. Antioxidant activities and caffeic acid content in New Zealand Asparagus (Asparagus officinalis) roots extracts. Antioxidants 2018, 7, 52. [Google Scholar] [CrossRef]
- Acharyaa, S.J.; Acharyaa, N.S.; Bhangalea, J.O.; Shahb, S.K.; Pandya, S.S. Antioxidant and hepatoprotective action of Asparagus racemosus Willd. root extracts. Indian J. Exp. Biol. 2012, 50, 795–801. [Google Scholar]
- Nile, S.H.; Park, S.W. HPTLC Analysis, Antioxidant and Antigout Activity of Indian Plants. Iran. J. Pharm. Sci. 2014, 13, 531–539. [Google Scholar]
- Vu, O.H.; Hang, T.T.M.; Yaguchi, S.; Ono, Y.; Pham, T.M.P.; Yamauchi, N.; Shigyo, M. Assessment of biochemical and antioxidant diversities in a shallot germplasm collection from Vietnam and its surrounding countries. Gen. Res. Crop. Evol. 2013, 60, 297–1312. [Google Scholar] [CrossRef]
- Hamouz, K.; Lachman, J.; Pazderů, K.; Tomášek, J.; Hejtmánková, K.; Pivec, V. Differences in anthocyanin content and antioxidant activity of potato tubers with different flesh color. Plant Soil Environ. 2011, 57, 478–485. [Google Scholar] [CrossRef]
- Basar, M.H.; Hossain, S.J.; Sadhu, S.K.; Rahman, M.H. A comparative study of antioxidant potential of commonly used antidiabetic plants in Bangladesh. Orient. Pharm. Exp. Med. 2013, 13, 21–28. [Google Scholar] [CrossRef]
- Terpinc, P.; Čeh, B.; Poklar Ulrih, N.; Abramovič, H. Studies of the correlation between antioxidant properties and the total phenolic content of different oil cake extracts. Ind. Crops Prod. 2012, 39, 2010–2017. [Google Scholar] [CrossRef]
- Piluzza, G.; Bullitta, S. Correlations between phenolic content and antioxidant properties in twenty-four plant species of traditional ethnoveterinary use in the Mediterranean area. Pharm. Biol. 2011, 49, 240–247. [Google Scholar] [CrossRef]
- Liu, W.; Huang, X.F.; Qi, Q.; Dai, Q.S.; Yang, L.; Nie, F.F.; Lu, N.; Gong, D.D.; Kong, L.Y.; Guo, Q.L. Asparanin A induces G(2)/M cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells. Biochem. Biophys. Res. Commun. 2009, 381, 700–705. [Google Scholar] [CrossRef]
- Xi, M.; Hai, C.; Tang, H.; Chen, M.; Fang, K.; Liang, X. Antioxidant and antiglycation properties of total saponins extracted from traditional Chinese medicine used to treat diabetes Mellitus. Phytother. Res. 2008, 22, 228–237. [Google Scholar] [CrossRef]
- Lee, J.H.; Jeon, J.K.; Kim, S.G.; Kim, S.H.; Chun, T.; Imm, J.-Y. Comparative analyses of total phenols, flavonoids, saponins and antioxidant activity in yellow soy beans and mung beans. Int. J. Food Sci. Technol. 2011, 46, 2513–2519. [Google Scholar] [CrossRef]
- Quiroga, P.R.; Grosso, N.R.; Lante, A.; Lomolino, G.; Zygadlo, J.A.; Nepote, V. Chemical composition, antioxidant activity and anti-lipase activityof Origanum vulgare and Lippia turbinata essential oils. Int. J. Food Sci. Technol. 2013, 48, 642–649. [Google Scholar] [CrossRef]
- Guillen, R.; Rodrıguez, R.; Jaramillo, S.; Rodrıguez, G.; Espejo, J.A.; Fernandez-Bolanos, J.; Heredia, A.; Jiménez, A. Antioxidants from asparagus spears: Phenolics. Acta Hortic. 2008, 776, 247–253. [Google Scholar] [CrossRef]
- Birari, R.B.; Bhutani, K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Perri, V.; Menichini, F.; Marrelli, M.; Uzunov, D.; Statti, G.A.; Menichini, F. Wild mediterranean dietary plants as inhibitors of pancreatic lipase. Phytother. Res. 2012, 26, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Harding, S.V.; Marinangeli, C.P.F.; Kim, Y.S.; Jones, P.J.H. Hypocholesterolemic and anti-obesity effects of saponins from Platycodon grandiflorum in hamsters fed atherogenic diets. J. Food Sci. 2008, 73, H195–H200. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Sim, J.S.; Shim, S.H.; Ha, Y.W.; Kang, S.S.; Kim, Y.S. Antiobese and hypolipidemic effects of platycodin saponins in diet-induced obese rats: Evidences for lipase inhibition and calorie intake restriction. Int. J. Obes. 2005, 29, 983–990. [Google Scholar] [CrossRef]
- Xu, B.J.; Han, L.K.; Zheng, Y.N.; Lee, J.H.; Sung, C.K. In vitro inhibitory effect of triterpenoidal saponins from Platycodi radix on pancreatic lipase. Arch. Pharm. Res. 2005, 28, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Biswas, D.; Mathur, M.; Bhargava, S.; Malhotra, H.; Malhotra, B. Anticancer activity of root extracts in non-small cell lung cancer Asparagus racemosus A549 cells. Asian J. Pharm. Pharmacol. 2018, 4, 764–770. [Google Scholar] [CrossRef]
- Shao, Y.; Poobrasert, O.; Kennelly, E.J.; Chin, C.K.; Ho, C.T.; Huang, M.T.; Garrison, S.A.; Cordell, G.A. Cytotoxic activity of steroidal saponins from Asparagus officinalis. In IX International Asparagus Symposium; Benson, B., Ed.; International Society for Horticultural Science: Leuven, Belgium, 1999; pp. 277–282. [Google Scholar]
- Verma, S.; Tripathi, V.; Das, P. Asparagus racemosus leaf extract inhibits growth of UOK renal cell carcinoma cell line: Simultaneous oncogenic PRCCTFE3 fusion transcript inhibition and apoptosis independent cell death. Asian Pac. J. Cancer Prev. 2014, 15, 1937–1941. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, V.; Tadinada, S.M.; Jain, A.; Sehdev, V.; Daniels, C.K.; Lai, J.C.; Bhushan, A. Biochanin A reduces pancreatic cancer survival and progression. Anticancer Drugs 2014, 25, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y.; Li, J.P. Saikosaponin-d inhibits proliferation of human undifferentiated thyroid carcinoma cells through induction of apoptosis and cell cycle arrest. Riv. Eur. Sci. Med. Farmacol. 2014, 18, 2435–2443. [Google Scholar]
- Zhang, W.; Zhang, D.; Ma, X.; Liu, Z.; Li, F.; Wu, D. Paris saponin VII suppressed the growth of human cervical cancer Hela cells. Eur. J. Med. Res. 2014, 19, 41. [Google Scholar] [CrossRef]
- Weidner, S.; Paproka, J.; Lukaszewicz, D. Change in free, sterified and glycosidic phenolic acids in cereal grains during the after-ripening. Seed Sci. Technol. 1996, 24, 107–114. [Google Scholar]
- Soluski, F.; Krygier, K.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids 3. Composition of phenolic acids in cereal and potatoe flours. J. Agric. Food Chem. 1982, 30, 337–340. [Google Scholar]
- De Ruiter, J.; Burns, J. Characterization of trifluoroacetic acid hydrolyzed subtropical forage grass cell walls. J. Agric. Food Chem. 1987, 4, 308–316. [Google Scholar] [CrossRef]
- Englyst, H.N.; Cummings, J.H. Simplified method for the measurement of total non-starch polysaccharides by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1984, 109, 937. [Google Scholar] [CrossRef]
- Rodríguez, R.; Jaramillo, S.; Rodríguez, G.; Espejo, J.A.; Guillén, R.; Fernández-Bolaños, J.; Jiménez, A. Antioxidant activity of ethanolic extracts from several asparagus cultivars. J. Agric. Food Chem. 2005, 53, 5212–5217. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Volatile composition, phenolic content, and in vitro antioxidant activity of red wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar] [CrossRef]
- Rubio-Senent, F.; Rodríguez-Gutiérrez, G.; Lama-Muñoz, A.; Fernández-Bolaños, J. New phenolic compounds hydrothermally extracted from the olive oil by-product alperujo and their antioxidative activities. J. Agric. Food Chem. 2012, 60, 1175–1186. [Google Scholar] [CrossRef]
- Psarra, E.; Makris, D.P.; Kallithraka, S.; Kefalas, P. Evaluation of the antiradical and reducing properties of selected Greek whitewines: Correlation with polyphenolic composition. J. Sci. Food Agric. 2002, 82, 1014–1020. [Google Scholar] [CrossRef]
- Kim, J.; Jang, D.S.; Kim, H.; Kim, J.S. Anti-lipase and lipolytic activities of ursolic acid isolated from the roots of Actinidia arguta. Arch. Pharm. Res. 2009, 32, 983–987. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolics | Leaf | Stem | Pericarp | Rhizome |
|---|---|---|---|---|
| Gallic acid | n.d. | 35.48 ± 0.12 a | 36.85 ± 0.79 a | 61.72 ± 0.71 b |
| Protocatechuic acid | 14.12 ± 0.64 a | 13.70 ± 0.27 a | 173.66 ± 0.80 c | 105.97 ± 0.11 b |
| p-Hydroxybenzoic acid | 23.30 ± 0.34 c | 13.75 ± 0.37 a | 20.83 ± 0.41 b | 160.85 ± 0.54 d |
| Vanillic acid | 6.72 ± 0.37 a | n.d. | n.d. | 84.75 ± 0.65 b |
| Caffeic acid | 32.70 ± 1.13 b | 93.06 ± 0.22 d | 2.89 ± 0.17 a | 78.09 ± 0.20 c |
| OH-benzaldehyde | 12.15 ± 1.03 b | 10.25 ± 0.18 a | 10.10 ± 0.37 a | 39.76 ± 0.16 c |
| Syringic acid | n.d. | n.d. | n.d. | 81.20 ± 0.46 |
| Vanillin | n.d. | 20.86 ± 0.34 b | n.d. | 15.00 ± 0.34 a |
| p-Coumaric acid | 8.27 ± 0.21 c | 2.79 ± 0.13 a | 7.66 ± 0.01 b | 133.25 ± 0.01 d |
| t-ferulic acid | 25.00 ± 0.27 a | 28.82 ± 0.38 b | 24.60 ± 0.76 a | 156.99 ± 0.34 c |
| t-Sinapic acid | n.d. | n.d. | 12.94 ± 0.28 b | 12.66 ± 0.76 a |
| Total | 122.27 ± 0.97 a | 218.70 ± 0.90 b | 289.53 ± 1.99 c | 930.23 ± 4.27 d |
| Peak # | Rt (min) | λmax (nm) | Molecular Ion [M + H]+ (m/z) | Ions (ESI+) (m/z) | Assigned Structures |
|---|---|---|---|---|---|
| 1 | 19.10 | 256, 308sh, 352 | 919 | 757,611,465,303 | Quercetin-3-glucosyl-(1->4)-rhamnoside-7-rutinoside |
| 2 | 20.40 | 252, 288sh, 352 | 933 | 771,625,479,317 | isorhamnetin-3-glucosyl-(1->4)-rhamnoside-7-rutinoside |
| 3 | 25.30 | 255, 267sh, 352 | 757 | 611,465,303 | Quercetin-3-O-rhamnosylrutinoside |
| 4 | 27.10 | 256, 267sh, 354 | 773 | 627,481,303 | Quercetin-3-O-glucosyl-rutinoside |
| 5 | 27.50 | 254, 268sh, 355 | 771 | 625,479,317 | Isorhamnetin-3-O-rhamnosyl-rutinoside |
| 6 | 29.70 | 255, 267sh, 355 | 611 | 465,303 | Quercetin-3-O-rhamnoglucoside (rutin) |
| 7 | 41.45 | 252, 267sh, 372 | 303 | - | Quercetin |
| 8 | 42.36 | 257, 288 | 273 | - | Naringenin |
| 9 | 43.58 | 264, 296sh, 364 | 287 | - | Kaempferol |
| Peak | Flavonoids Compound | Quantification (mg/kg, dw) | |||
|---|---|---|---|---|---|
| Leaf | Stem | Pericarp | Rhizome | ||
| 1 | Q-3-glucosyl-rhamnosyl-rutinoside | 519 ± 48.12 b | 236 ± 5 a | n.d. | n.d. |
| 2 | IR-3-glucosyl-rhamnosyl-rutinoside | 1343 ± 92.88 b | 570 ± 38 a | n.d. | n.d. |
| 3 | Q-3-rhamnosylrutinoside | 621 ± 60 c | 250 ± 3 a | 370 ± 25 b | n.d. |
| 4 | Q-3-glucosyl-rutinoside | 296 ± 18 a | 1095 ± 41 b | n.d. | n.d. |
| 5 | IR-3-rhamnosyl-rutinoside | 619 ± 40 | n.d. | n.d. | n.d. |
| 6 | Rutin | 337 ± 27 a | n.d. | 925 ± 72 b | n.d. |
| 7 | Quercetin | n.d. | n.d. | 43.98 ± 0.33 c | n.d |
| 8 | Naringenin | 5.06 ± 0.21 a | 6.18 ± 0.21 b | 6.24 ± 0.44 b | n.d. |
| 9 | Kaempferol | 47.78 ± 0.65 b | 33.80 ± 1.75 a | 69.93 ± 0.07 c | n.d. |
| Free flavonoids | 52.84 ± 0.86 c | 39.98 ± 1.96 b | 120.15 ± 0.70 d | n.d | |
| Total flavonoids glycoside | 3734 ± 287 c | 2151 ± 88 b | 1598 ± 94 a | n.d. | |
| Molecular Ion (m/z) | Ion Fragmentation | ||||||
|---|---|---|---|---|---|---|---|
| saponins | Rt a (min) | Formula | [M − H]− Experimental | [M − H]− Theoretical | Error (ppm) | Neg mode | Pos mode |
| HTSAP-1 | 24.57 | C50H83O23 | 1051.5331 | 1051.5311 | −1.8 | Pen919-Hex757-Hex595-Hex433 Hex889-Pen757-Hex595-Hex433 | [-Na-H2O]1035-Hex873-Hex711-Pen579-Hex417 [-Na-H2O]1035-Hex873-Pen741-Hex579-Hex417 |
| STIPSAP-1 | 28.29 | C49H79O22 | 1019.5068 | 1019.5050 | 1.8 | b Pen887-Pen755-c Hex593-Hex431 | [-Na-H2O]1003-Pen871-Hex709-Pen577-Hex415 |
| HTSAP-2 | 29.37 | C49H81O22 | 1021.5225 | 1021.5210 | −1.5 | Pen889-Pen757-Hex595-Hex433 | [-Na-H2O]1005-Pen873-Hex711-Pen579-Hex417 |
| STIPSAP-2 | 37.12 | C35H63O22 | 835.3816 | 835.3816 | −0.1 | nd | [-Na-H2O]819-Hex657-d DoHex511-e Unk415 |
| WSAP-4 | 37.59 | C35H65O22 | 837.3997 | 837.3973 | 1.4 | nd | [-Na-H2O]821-Hex659-DoHex513-Unk417 |
| STIPSAP-3 | 43.01 | C35H63O22 | 835.3792 | 835.3816 | 3.0 | nd | [-Na-H2O]819-Hex657-DoHex511-Unk415 |
| STIPSAP-4 | 48.75 | C43H69O16 | 841.4591 | 841.4614 | 2.7 | Pen709-Pen577-Unk433 | [-Na-H2O]825-Pen693-Pen561-Unk 417 |
| Saponin | Leaf | Stem | Pericarp | Rhizome |
|---|---|---|---|---|
| HTSAP-1 | - | - | 411.8 ± 10.3 1% | - |
| STIPSAP-1 | - | 179.8 ± 3.5 1% | 3844.2 ± 340.6 7% | - |
| HTSAP-2 | - | - | 5640.3 ± 439.6 10% | 10,772.3 ± 31.9 19% |
| STIPSAP-2 | 12,656.0 ± 964.6 60% | 16,367.8 ± 1048.6 60% | 19,730.2 ± 948.1 37% | - |
| WSAP-4 | 6937.7 ± 470.3 33% | 7727.0 ± 664.0 28% | 16,374.2 ± 997.2 30% | 32,245.9 ± 142.3 58% |
| STIPSAP-3 | 1994.9 ± 135.6 9% | 3171.6 ± 275.2 12% | 6383.2 ± 195.5 12% | 4540.8 ± 483.5 8% |
| STIPSAP-4 | - | - | 1638.3 ± 94.8 3% | 8128.6 ± 582.1 15% |
| Total saponins | 21,588.6 ± 1570.5 d | 27,446.3 ± 1984.3 c | 54,022.2 ± 1129.9 b | 55,687.6 ± 209.0 a |
| DPPH | FRAP | ABTS | |
|---|---|---|---|
| TPs | ** 0.586826 | *** 0.868038 | −0.253 |
| TFs | *** 0.861817 | *** −0.966198 | −0.055 |
| TSs | *** 0.910861 | *** 0.916607 | 0.488 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdi, A.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Karray Bouraoui, N.; Guillén-Bejarano, R. Phytochemical Profile and In Vitro Bioactivities of Wild Asparagus stipularis. Molecules 2024, 29, 817. https://doi.org/10.3390/molecules29040817
Hamdi A, Jaramillo-Carmona S, Rodríguez-Arcos R, Jiménez-Araujo A, Karray Bouraoui N, Guillén-Bejarano R. Phytochemical Profile and In Vitro Bioactivities of Wild Asparagus stipularis. Molecules. 2024; 29(4):817. https://doi.org/10.3390/molecules29040817
Chicago/Turabian StyleHamdi, Amel, Sara Jaramillo-Carmona, Rocío Rodríguez-Arcos, Ana Jiménez-Araujo, Najoua Karray Bouraoui, and Rafael Guillén-Bejarano. 2024. "Phytochemical Profile and In Vitro Bioactivities of Wild Asparagus stipularis" Molecules 29, no. 4: 817. https://doi.org/10.3390/molecules29040817
APA StyleHamdi, A., Jaramillo-Carmona, S., Rodríguez-Arcos, R., Jiménez-Araujo, A., Karray Bouraoui, N., & Guillén-Bejarano, R. (2024). Phytochemical Profile and In Vitro Bioactivities of Wild Asparagus stipularis. Molecules, 29(4), 817. https://doi.org/10.3390/molecules29040817