
β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress


Abstract
1. Introduction
2. Results
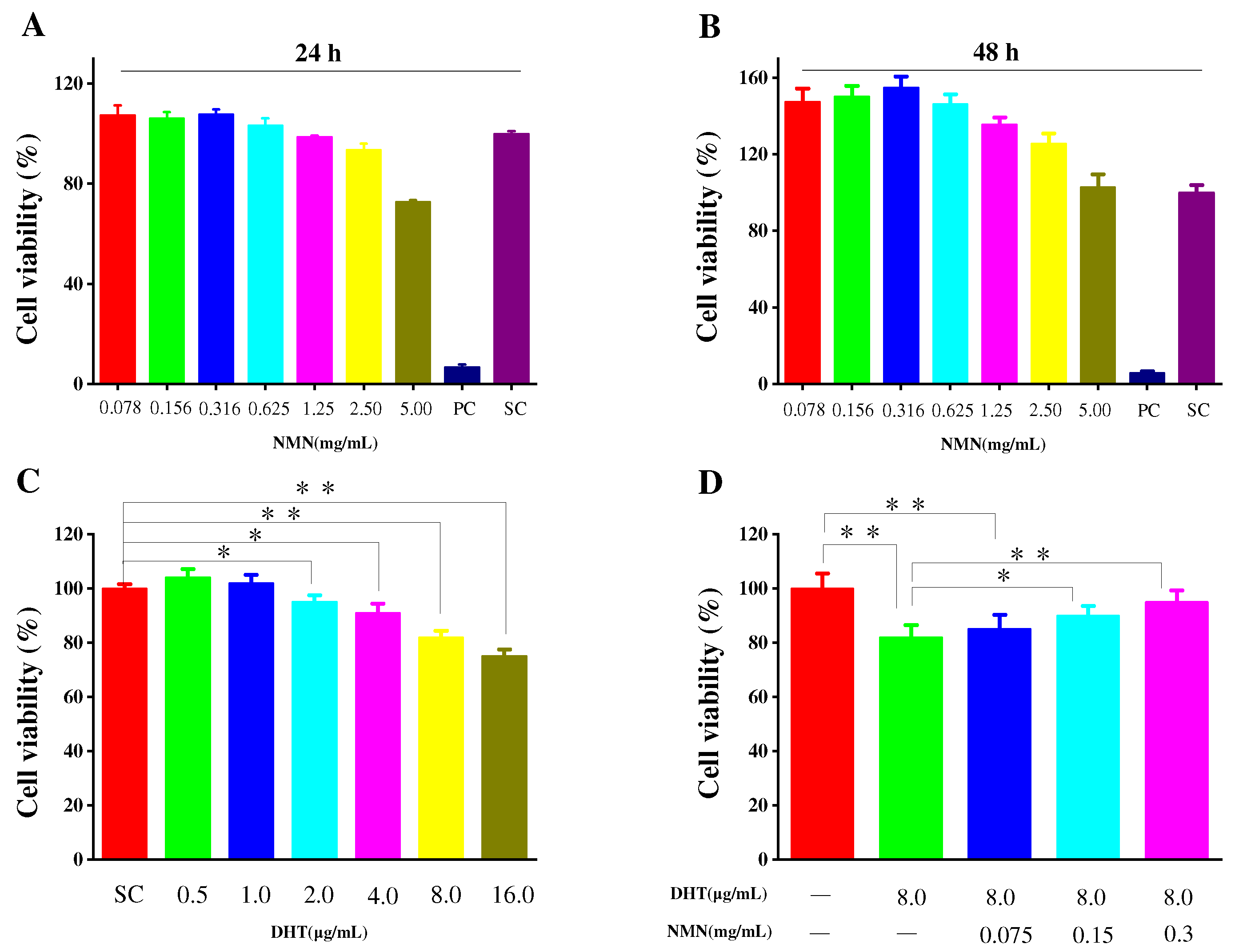
2.1. Effects of NMN on Cell Proliferation
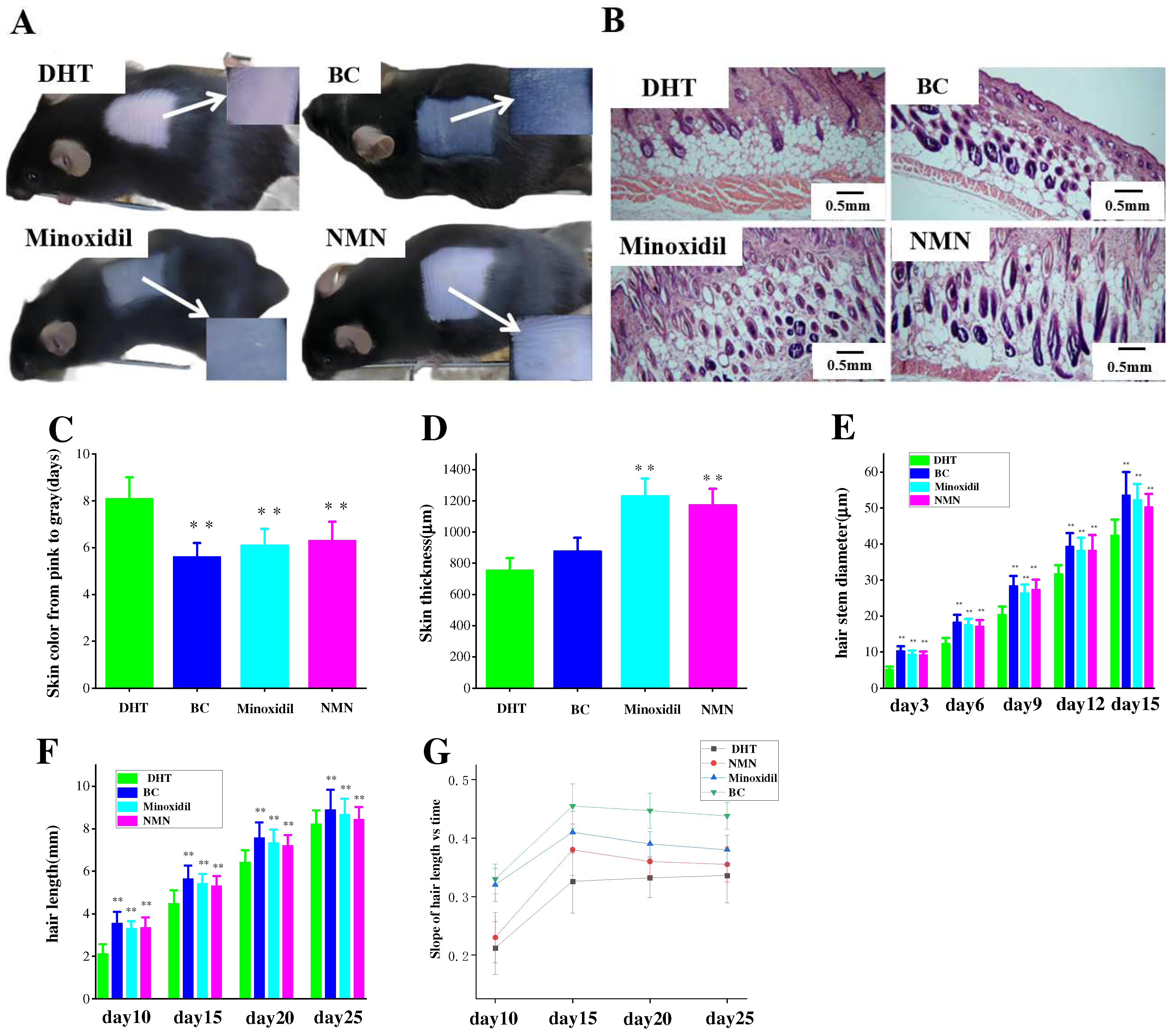
2.2. Induction of Hair Growth in Resting Mice by NMN
2.3. The Effect of NMN on Hair Growth Improvement
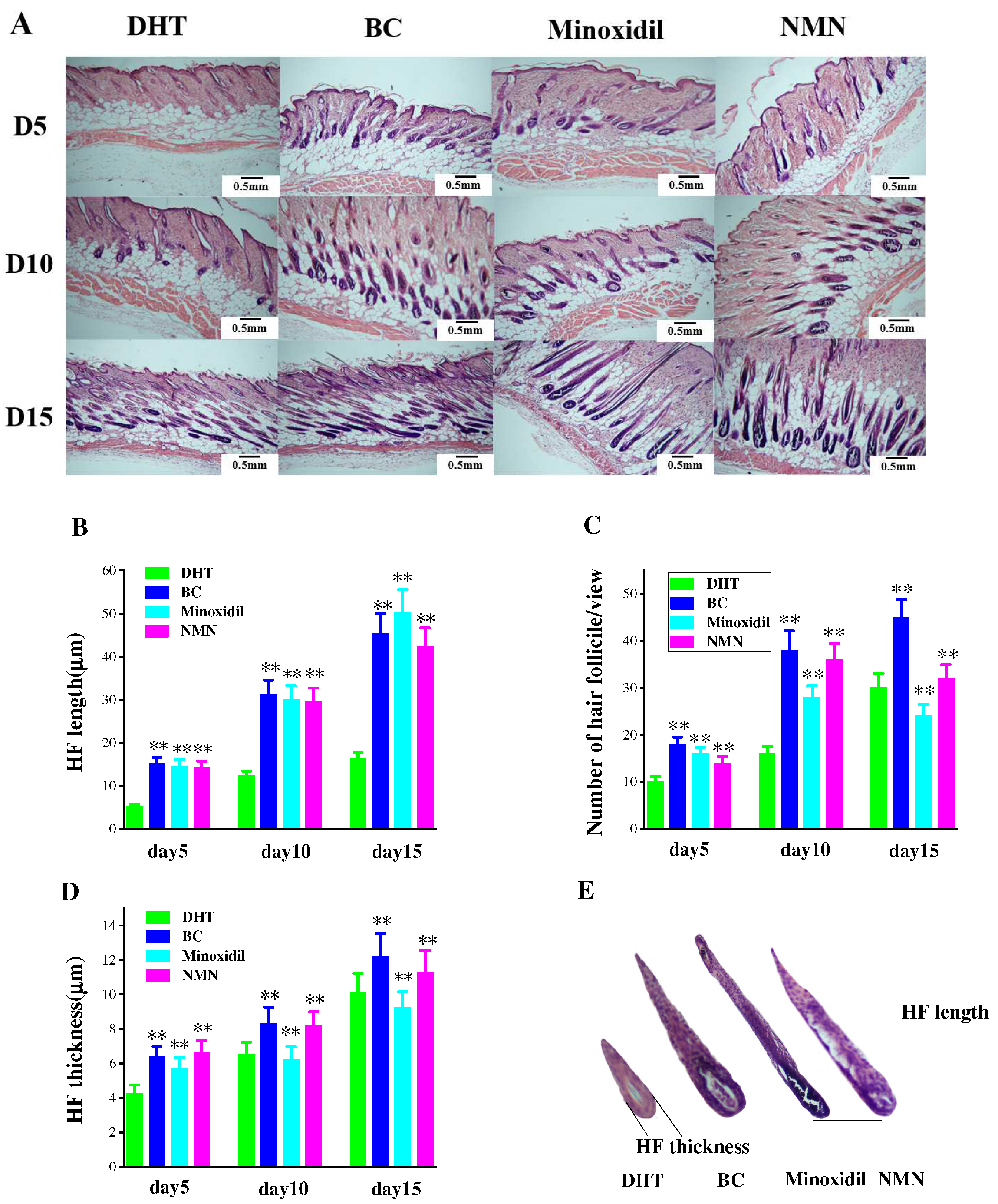
2.4. Enhancement of Hair Follicle Growth by NMN
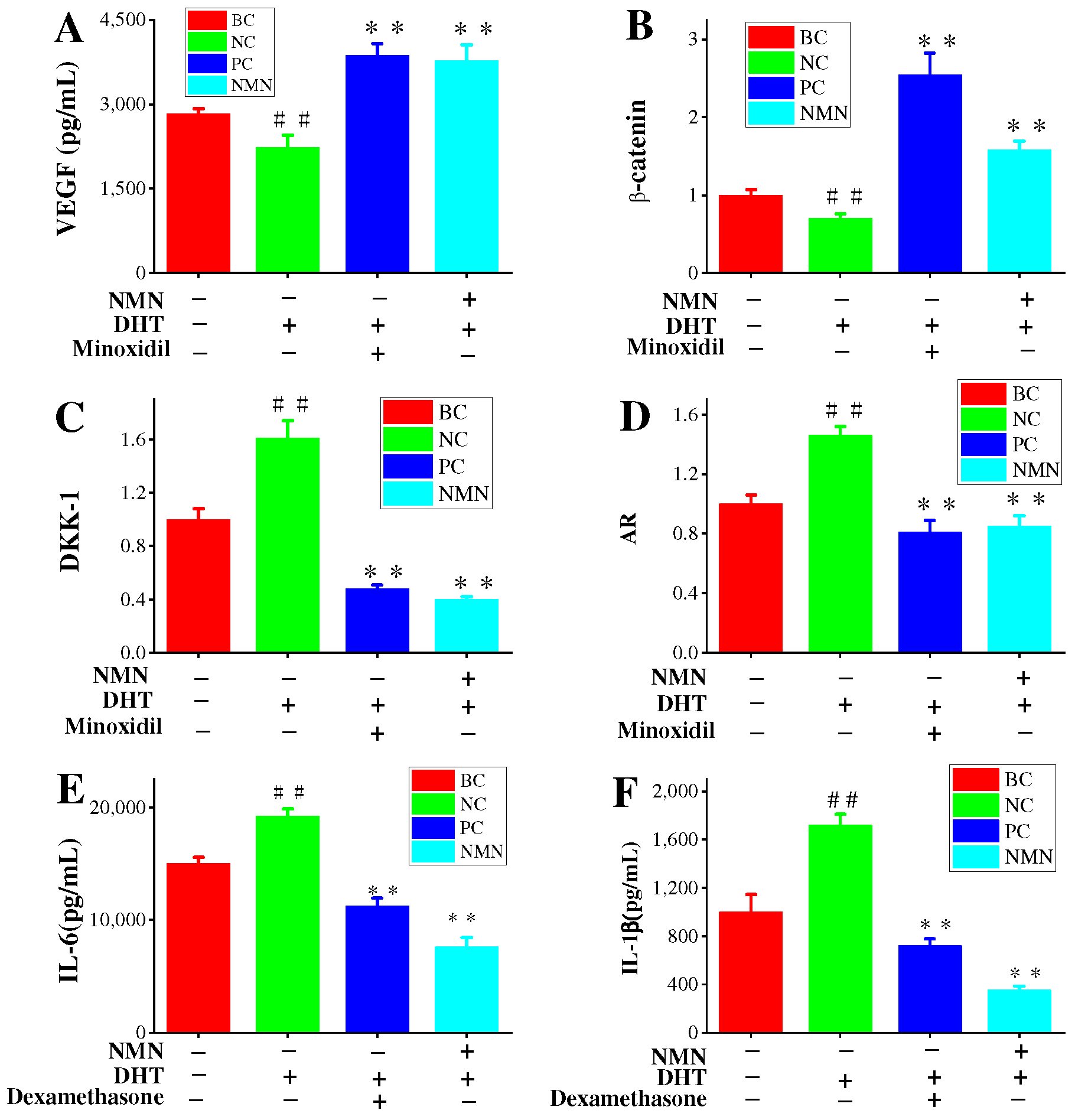
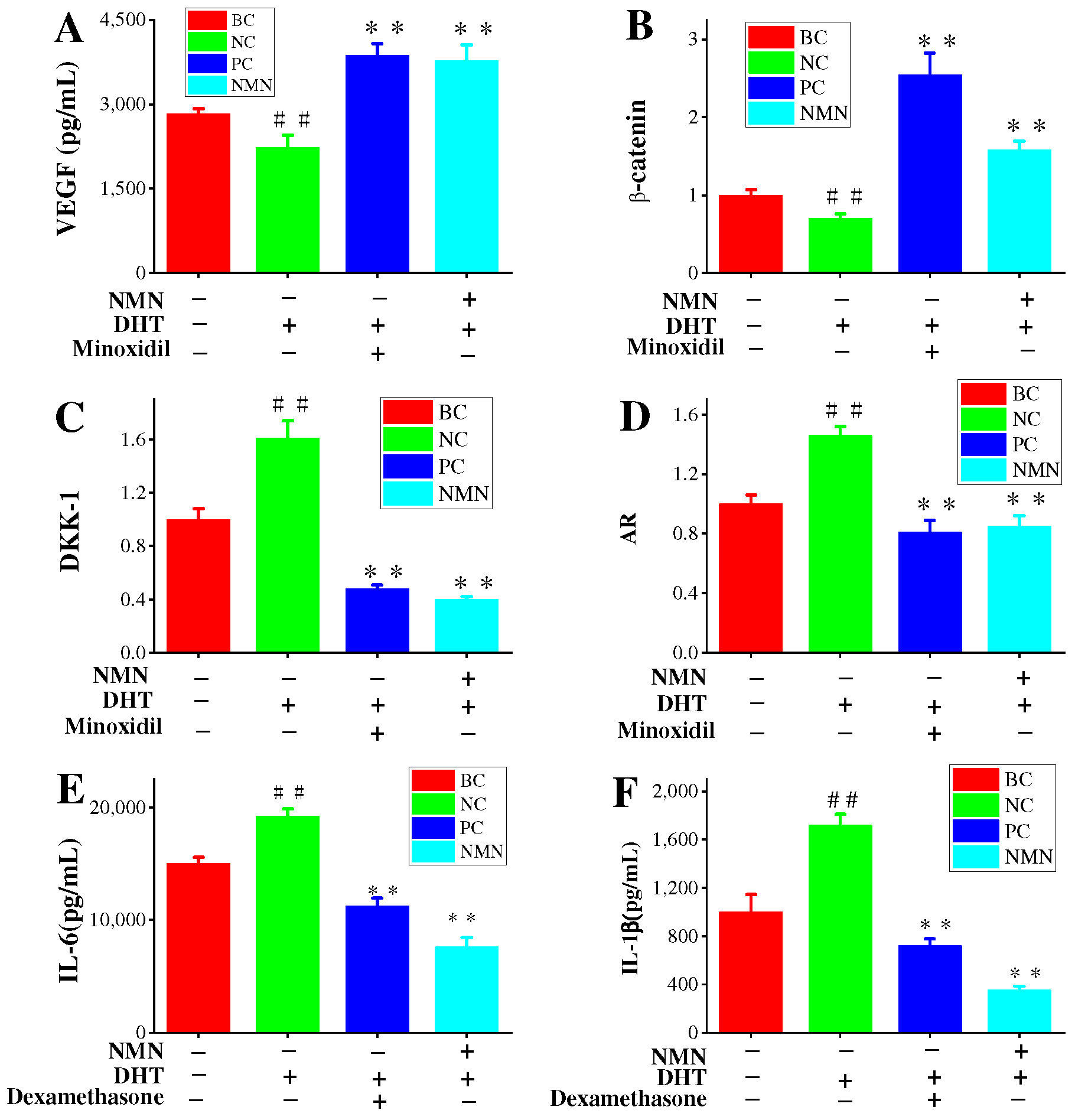
2.5. Effects of NMN on VEGF Expression in HDPCs
2.6. NMN Downregulation AR Expression and Activation Wnt/β-Catenin Pathway
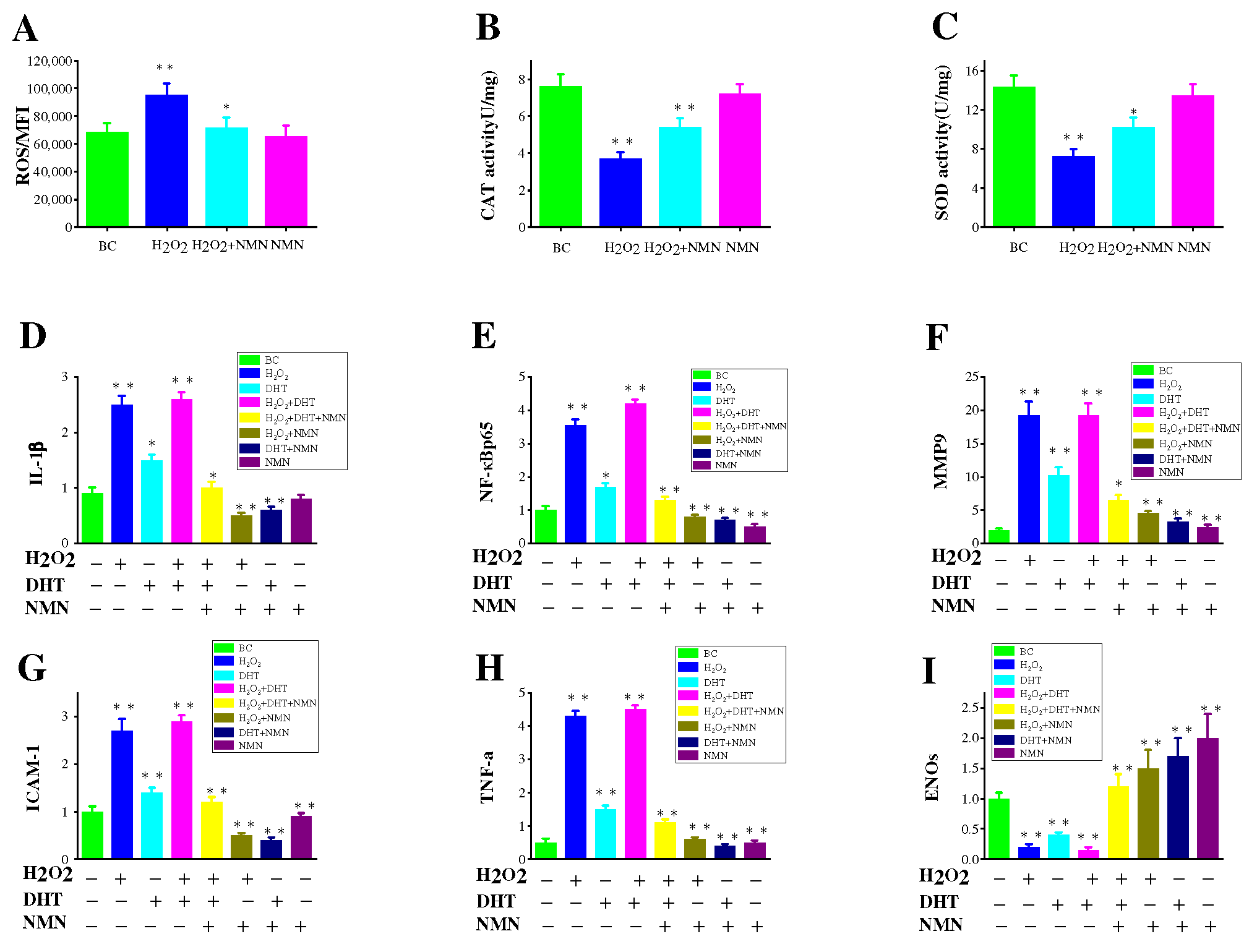
2.7. Effects of NMN on the Capacity to Eliminate ROS
2.8. Inflammatory Pathways Associated with H2O2, DHT and NMN
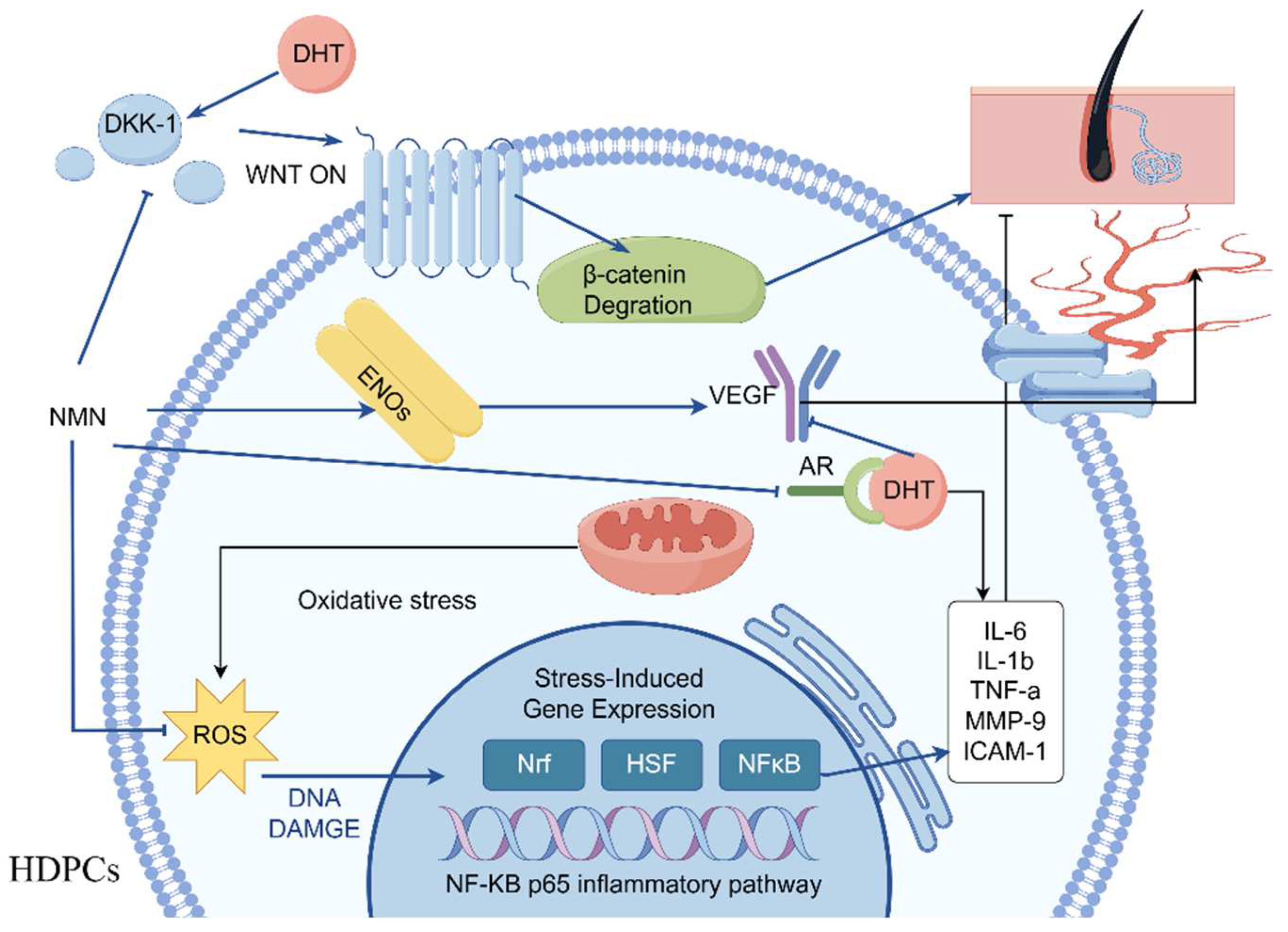
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Animal Experiments
4.3. H & E Staining
4.4. Cell Cultures
4.5. Cytotoxicity Test
4.6. RNA Preparation and qRT-PCR
4.7. VEGF, IL-6 and IL-1β ELISA Detection
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Segal-Engelchin, D.; Shvarts, S. Does Severity of Hair Loss Matter? Factors Associated with Mental Health Outcomes in Women Irradiated for Tinea Capitis in Childhood. Int. J. Environ. Res. Public Health 2020, 17, 7388. [Google Scholar] [CrossRef] [PubMed]
- Dobreva, A. Using Mathematical Tools to Investigate the Autoimmune Hair Loss Disease Alopecia Areata. Ph.D. Thesis, The Florida State University, Tallahassee, FL, USA, 2018. [Google Scholar]
- Chen, S.; Xie, X.; Zhang, G.; Zhang, Y. Comorbidities in Androgenetic Alopecia: A Comprehensive Review. Dermatol. Ther. 2022, 12, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, G.; Caruso, M.R.; Milioto, S.; Fakhrullin, R.; Lazzara, G. Keratin/alginate hybrid hydrogels filled with halloysite clay nanotubes for protective treatment of human hair. Int. J. Biol. Macromol. 2022, 222, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, G.; Milioto, S.; Svetlana, K.; Fakhrullina, G.; Akhatova, F.; Lazzara, G.; Fakhrullin, R.; Lvov, Y. Halloysite/Keratin Nanocomposite for Human Hair Photoprotection Coating. ACS Appl. Mater. Interfaces 2020, 12, 24348–24362. [Google Scholar] [CrossRef] [PubMed]
- Panchal, A.; Fakhrullina, G.; Fakhrullin, R.; Lvov, Y. Self-assembly of clay nanotubes on hair surface for medical and cosmetic formulations. Nanoscale 2018, 10, 18205–18216. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.; Scott, F.H.; Lvov, Y.; Stavitskaya, A.; Akhatova, F.; Konnova, S.; Fakhrullina, G.; Fakhrullin, R. Clay Nanotube Immobilization on Animal Hair for Sustained Anti-Lice Protection. Pharmaceutics 2021, 13, 1477. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Garcovich, S. Systematic Review of Platelet-Rich Plasma Use in Androgenetic Alopecia Compared with Minoxidil®, Finasteride®, and Adult Stem Cell-Based Therapy. Int. J. Mol. Sci. 2020, 21, 2702. [Google Scholar] [CrossRef]
- Gentile, P.; Mun-Sang, K. Hair Growth Booster Effects of Micro-Needling with Low-Level Led Therapy and Growth Factors on Subjects Treated with Finasteride®. Appl. Sci. 2022, 12, 9164. [Google Scholar] [CrossRef]
- Soares Júnior, J.M.; Guimarães, D.Z.; Simões, R.D.S.; Veiga, E.C.d.A.; Roa, C.L.; Sorpreso, I.C.E.; Baracat, M.C.; Baracat, E.C. Systematic review of finasteride effect in women with hirsutism. Rev. Assoc. Medica Bras. 2021, 67, 1043–1049. [Google Scholar] [CrossRef]
- Jung, H.; Da-Min, J.; Sang-Soo, L.; Eun-Mi, K.; Yoon, K.; Kim, K.K. Mangifera Indica leaf extracts promote hair growth via activation of Wnt signaling pathway in human dermal papilla cells. Anim. Cells Syst. 2022, 26, 129–136. [Google Scholar] [CrossRef]
- Ma, L.; Shen, H.; Fang, C.; Chen, T.; Wang, J. Camellia Seed Cake Extract Supports Hair Growth by Abrogating the Effect of Dihydrotestosterone in Cultured Human Dermal Papilla Cells. Molecules 2022, 27, 6443. [Google Scholar] [CrossRef]
- Young-Hyun, B.; Jin-Ho, L.; Sang-Jin, C.; Chae, Y.; Myung-Hun, L.; Sun-Hong, K.; Kwon-Il, H.; Tack-Joong, K. Heat-Killed Enterococcus faecalis EF-2001 Induces Human Dermal Papilla Cell Proliferation and Hair Regrowth in C57BL/6 Mice. Int. J. Mol. Sci. 2022, 23, 5413. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Mechanism of action of herbs and their active constituents used in hair loss treatment. Fitoterapia 2016, 114, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Su Bin, H.; Park, H.J.; Lee, B.-H. Hair-Growth-Promoting Effects of the Fish Collagen Peptide in Human Dermal Papilla Cells and C57BL/6 Mice Modulating Wnt/β-Catenin and BMP Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 11904. [Google Scholar] [CrossRef]
- Kwack, M.H.; Ahn, J.S.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dihydrotestosterone-Inducible IL-6 Inhibits Elongation of Human Hair Shafts by Suppressing Matrix Cell Proliferation and Promotes Regression of Hair Follicles in Mice. J. Investig. Dermatol. 2012, 132, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Bryan Siu-Yin, H.; Vaz, C.; Ramasamy, S.; Elaine Guo Yan, C.; Jameelah Sheik, M.; Jaffar, H.; Hillmer, A.; Tanavde, V.; Bigliardi-Qi, M.; Paul Lorenz, B. Progressive expression of PPARGC1α is associated with hair miniaturization in androgenetic alopecia. Sci. Rep. Nat. Publ. Group 2019, 9, 8771. [Google Scholar] [CrossRef]
- Chew, E.G.Y.; Tan, J.H.J.; Bahta, A.W.; Ho, B.S.Y.; Liu, X.; Lim, T.C.; Sia, Y.Y.; Bigliardi, P.L.; Heilmann, S.; Wan, A.C.A.; et al. Differential Expression between Human Dermal Papilla Cells from Balding and Non-Balding Scalps Reveals New Candidate Genes for Androgenetic Alopecia. J. Investig. Dermatol. 2016, 136, 1559–1567. [Google Scholar] [CrossRef]
- Sick, S.; Reinker, S.; Timmer, J.; Schlake, T. WNT and DKK determine hair follicle spacing through a reaction-diffusion mechanism. Science 2006, 314, 1447–1450. [Google Scholar] [CrossRef] [PubMed]
- Premanand, A.; Rajkumari, B.R. Androgen modulation of Wnt/β-catenin signaling in androgenetic alopecia. Arch. Dermatol. Res. 2018, 310, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.W. The Molecular Mechanism of Natural Products Activating Wnt/β-Catenin Signaling Pathway for Improving Hair Loss. Life 2022, 12, 1856. [Google Scholar] [CrossRef]
- Marinescu, G.C.; Popescu, R.-G.; Stoian, G.; Dinischiotu, A. β-nicotinamide mononucleotide (NMN) production in Escherichia coli. Sci. Rep. Nat. Publ. Group 2018, 8, 12278. [Google Scholar] [CrossRef]
- Sharma, A.; Chabloz, S.; Lapides, R.A.; Roider, E.; Ewald, C.Y. Potential Synergistic Supplementation of NAD+ Promoting Compounds as a Strategy for Increasing Healthspan. Nutrients 2023, 15, 445. [Google Scholar] [CrossRef]
- Kiss, T.; Giles, C.B.; Tarantini, S.; Yabluchanskiy, A.; Balasubramanian, P.; Gautam, T.; Csipo, T.; Nyúl-Tóth, Á.; Lipecz, A.; Szabo, C.; et al. Nicotinamide mononucleotide (NMN) supplementation promotes anti-aging miRNA expression profile in the aorta of aged mice, predicting epigenetic rejuvenation and anti-atherogenic effects. GeroScience 2019, 41, 419–439. [Google Scholar] [CrossRef]
- Deng, X.; Liang, X.; Yang, H.; Huang, Z.; Huang, X.; Liang, C.; Kuang, Y.; Qin, Y.; Lin, F.; Luo, Z. Nicotinamide mononucleotide (NMN) protects bEnd.3 cells against H2O2-induced damage via NAMPT and the NF-κB p65 signalling pathway. FEBS Open Bio 2021, 11, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Katayoshi, T.; Uehata, S.; Nakashima, N.; Nakajo, T.; Kitajima, N.; Kageyama, M.; Tsuji-Naito, K. Nicotinamide adenine dinucleotide metabolism and arterial stiffness after long-term nicotinamide mononucleotide supplementation: A randomized, double-blind, placebo-controlled trial. Sci. Rep. Nat. Publ. Group 2023, 13, 2786. [Google Scholar] [CrossRef] [PubMed]
- Hwa Sun, R.; Jeong, J.; Chun Mong, L.; Lee, K.S.; Lee, J.-N.; Sung-Min, P.; Yong-Moon, L. Activation of Hair Cell Growth Factors by Linoleic Acid in Malva verticillata Seed. Molecules 2021, 26, 2117. [Google Scholar] [CrossRef]
- Meephansan, J.; Thummakriengkrai, J.; Ponnikorn, S.; Yingmema, W.; Deenonpoe, R.; Suchonwanit, P. Efficacy of topical tofacitinib in promoting hair growth in non-scarring alopecia: Possible mechanism via VEGF induction. Arch. Dermatol. Res. 2017, 309, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Khantham, C.; Linsaenkart, P.; Chaitep, T.; Pensak, J.; Chittasupho, C.; Rachtanapun, P.; Jantanasakulwong, K.; Phimolsiripol, Y.; Sarana Rose, S.; Prom-u-thai, C.; et al. Antioxidation, Anti-Inflammation, and Regulation of SRD5A Gene Expression of Oryza sativa cv. Bue Bang 3 CMU Husk and Bran Extracts as Androgenetic Alopecia Molecular Treatment Substances. Plants 2022, 11, 330. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell. Endocrinol. 2018, 465, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Li, S.; Liu, P.; Guo, L.; Gong, S.; Wan, Y. Effects of testosterone on skin structure and factors related to androgen conversion and binding in Hetian sheep. Trop. Anim. Health Prod. 2022, 54, 218. [Google Scholar] [CrossRef] [PubMed]
- Papukashvili, D.; Rcheulishvili, N.; Liu, C.; Xie, F.; Tyagi, D.; He, Y.; Wang, P.G. Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy. Cells 2021, 10, 2957. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, J. Modulation of Hair Growth Promoting Effect by Natural Products. Pharmaceutics 2021, 13, 2163. [Google Scholar] [CrossRef] [PubMed]
- Goldman, C.K.; Tsai, J.C.; Soroceanu, L.; Gillespie, G.Y. Loss of vascular endothelial growth factor in human alopecia hair follicles. J. Investig. Dermatol. 1995, 104, 18S–20S. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, M.; Tabata, Y. In vivo promoted growth of mice hair follicles by the controlled release of growth factors. Biomaterials 2003, 24, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Lachgar, S.; Charveron, M.; Gall, Y.; Bonafe, J.L. Minoxidil upregulates the expression of vascular endothelial growth factor in human hair dermal papilla cells. Br. J. Dermatol. 1998, 138, 407. [Google Scholar] [CrossRef]
- Woo, H.; Lee, S.; Kim, S.; Park, D.; Jung, E. Effect of sinapic acid on hair growth promoting in human hair follicle dermal papilla cells via Akt activation. Arch. Dermatol. Res. 2017, 309, 381–388. [Google Scholar] [CrossRef]
- Choi, M.; Choi, S.-J.; Jang, S.; Choi, H.-I.; Kang, B.-M.; Hwang, S.T.; Kwon, O. Shikimic acid, a mannose bioisostere, promotes hair growth with the induction of anagen hair cycle. Sci. Rep. 2019, 9, 17008. [Google Scholar] [CrossRef]
- Davey, R.A.; Grossmann, M. Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Clin. Biochem. Rev. 2016, 37, 3–15. [Google Scholar] [PubMed]
- Yang, Y.-C.; Fu, H.-C.; Wu, C.-Y.; Wei, K.-T.; Huang, K.-E.; Kang, H.-Y. Androgen Receptor Accelerates Premature Senescence of Human Dermal Papilla Cells in Association with DNA Damage: e79434. PLoS ONE 2013, 8, e79434. [Google Scholar] [CrossRef]
- Kishimoto, J.; Burgeson, R.E.; Morgan, B.A. Wnt signaling maintains the hair-inducing activity of the dermal papilla. Genes Dev. 2000, 14, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Structural and Functional Studies of MESD, a Specialized Chaperone Protein of LRP5/6 in Wnt Signaling. Ph.D. Thesis, Wayne State University, Detroit, MI, USA, 2009. [Google Scholar]
- Yun-Ho, C.; Jae Young, S.; Kim, J.; Kang, N.-G.; Lee, S. Niacinamide Down-Regulates the Expression of DKK-1 and Protects Cells from Oxidative Stress in Cultured Human Dermal Papilla Cells. Clin. Cosmet. Investig. Dermatol. 2021, 14, 1519–1528. [Google Scholar] [CrossRef]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. β-catenin Activity in the Dermal Papilla Regulates Morphogenesis and Regeneration of Hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef]
- Jeong, G.; Shin, S.H.; Kim, S.N.; Na, Y.; Park, B.C.; Cho, J.H.; Park, W.-S.; Kim, H.-J. Ginsenoside Re prevents 3-methyladenine-induced catagen phase acceleration by regulating Wnt/β-catenin signaling in human dermal papilla cells. J. Ginseng Res. 2022, 47, 440–447. [Google Scholar] [CrossRef]
- Li, V.S.W.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt signaling through inhibition of β-catenin degradation in an intact Axin1 complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/β-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Forward Primer | Reverse Primer |
|---|---|---|
| DKK-1 | ATGCGTCACGCTATGTGCTG | TGGAATACCCATCCAAGGTGCTA |
| AR | CTCTCACATGTGGAAGCTGCAAG | TTTCCGAAGACGACAAGATGGAC |
| β-catenin | TTAGCTGGTGGGCTGCAGAA | GGGTCCACCACTAGCCAGTATGA |
| ICAM-1 | CTGAAAGATGAGCTCGAGAGT | AAACGAATACACGGTGATGGTA |
| IL-1β | TCGCAGCAGCACATCAACAAGAG | TGCTCATGTCCTCATCCTGGAAGG |
| MMP-9 | CAAAGACCTGAAAACCTCCAAC | GACTGCTTCTCTCCCATCATC |
| eNOs | CTGAGAGCCTGCAATTACTACC | TTTCCACAGAGAGGATTGTAGC |
| TNF-α | ATGTCTCAGCCTCTTCTCATTC | GCTTGTCACTCGAATTTTGAGA |
| NF-κB p65 | TCGAGTCTCCATGCAGCTACGG | CGGTGGCGATCATCTGTGTCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Dai, J.; Ai, H.; Du, W.; Ji, H. β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress. Molecules 2024, 29, 798. https://doi.org/10.3390/molecules29040798
Xu C, Dai J, Ai H, Du W, Ji H. β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress. Molecules. 2024; 29(4):798. https://doi.org/10.3390/molecules29040798
Chicago/Turabian StyleXu, Chuntao, Jiawei Dai, Hongxia Ai, Weian Du, and Hongbing Ji. 2024. "β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress" Molecules 29, no. 4: 798. https://doi.org/10.3390/molecules29040798
APA StyleXu, C., Dai, J., Ai, H., Du, W., & Ji, H. (2024). β-Nicotinamide Mononucleotide Promotes Cell Proliferation and Hair Growth by Reducing Oxidative Stress. Molecules, 29(4), 798. https://doi.org/10.3390/molecules29040798