
GlcNAc6ST2/CHST4 Is Essential for the Synthesis of R-10G-Reactive Keratan Sulfate/Sulfated N-Acetyllactosamine Oligosaccharides in Mouse Pleural Mesothelium

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
2.1. R10G-Reactive Sulfated Glycans Are Abundant in the Mouse Pleural Mesothelium
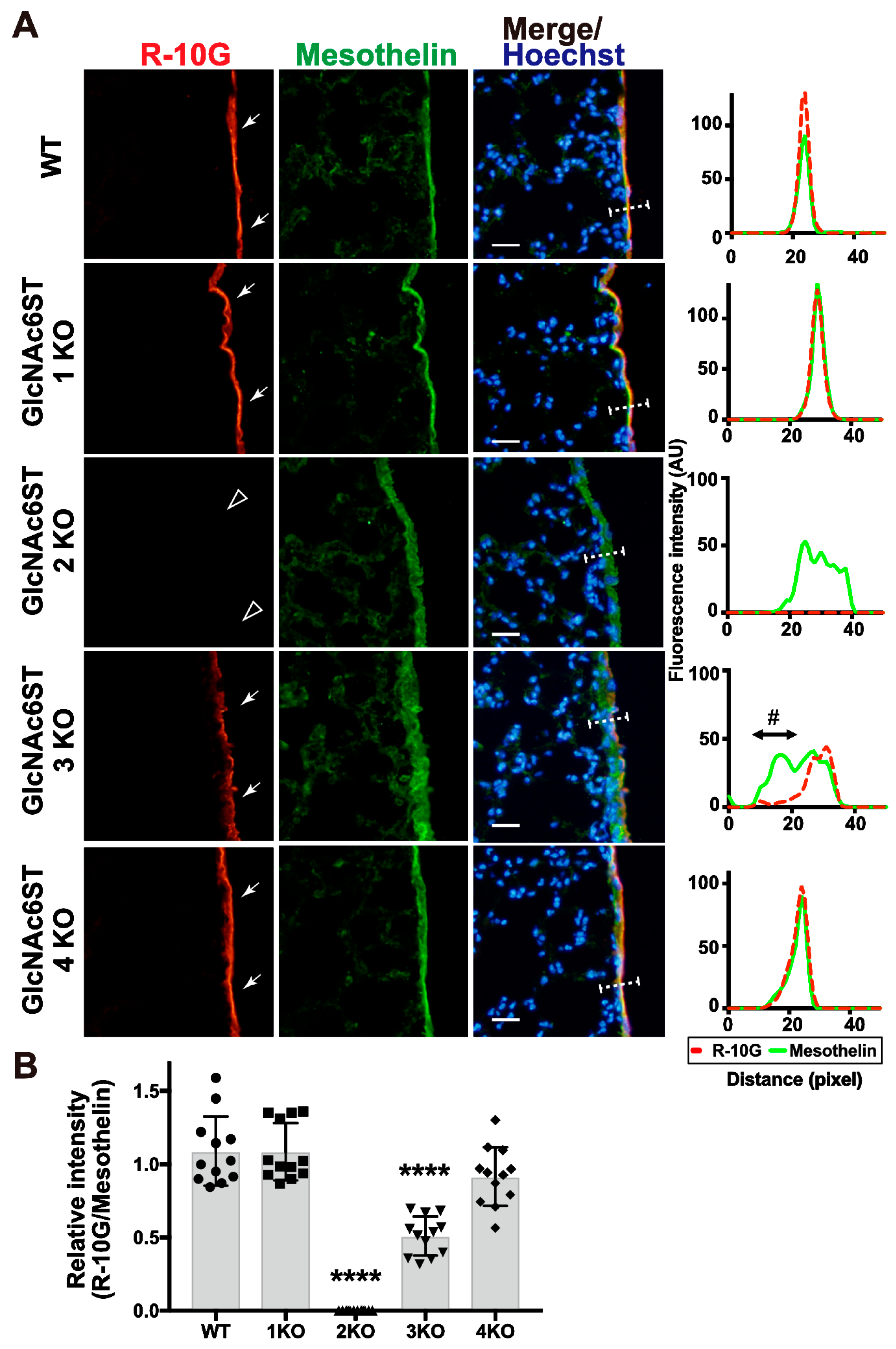
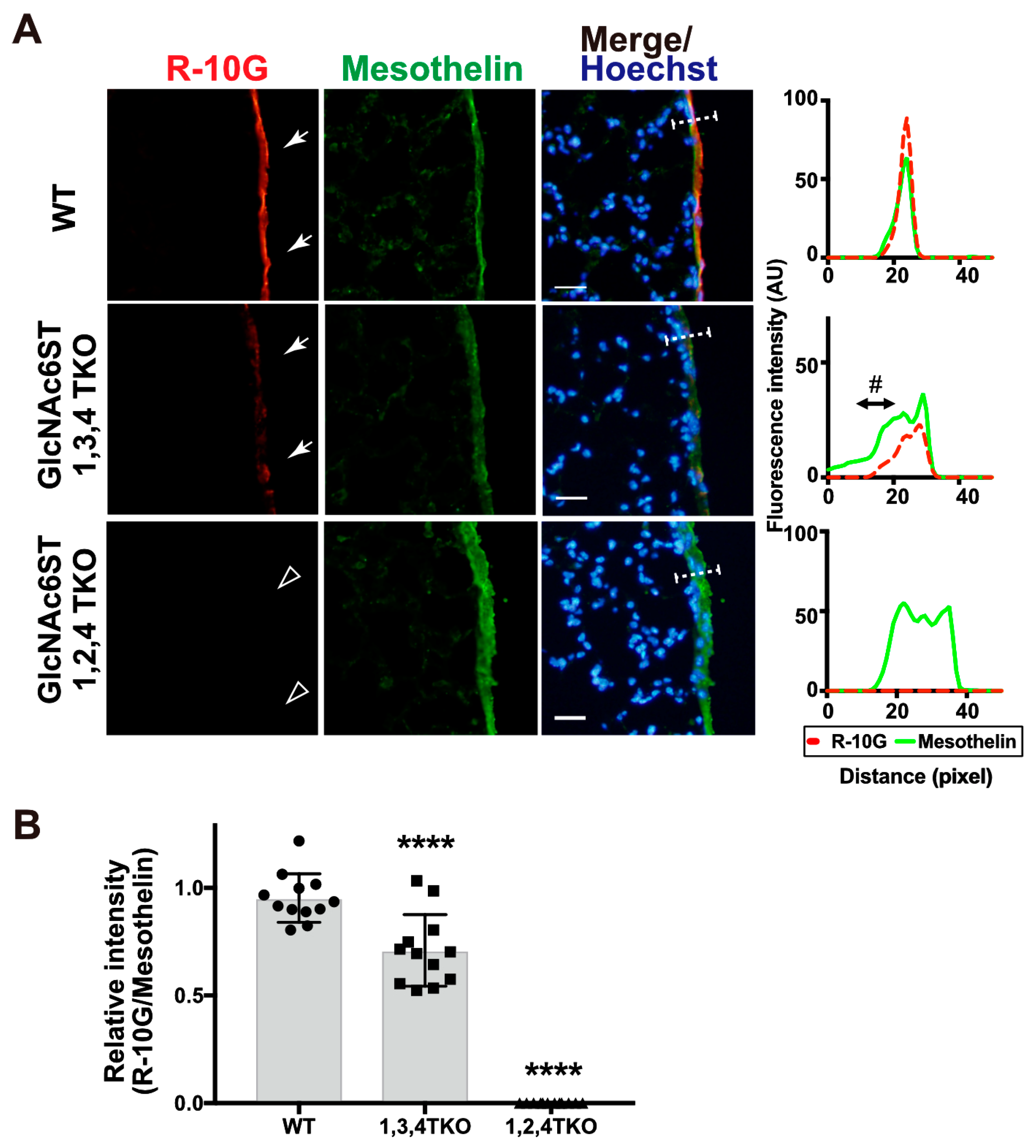
2.2. GlcNAc6ST2 Is Required and Sufficient for the Synthesis of R10G-Reactive KS Glycans in the Mouse Pleural Mesothelium
3. Discussion
4. Materials and Methods
4.1. Antibodies and Enzymes
4.2. Mice
4.3. Mouse Tissues
4.4. Immunohistochemistry and Fluorescence Microscopy
4.5. Immunoprecipitation
4.6. WGA-Bead-Bound Precipitation
4.7. Immunoblots
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uchimura, K.; Rosen, S.D. Sulfated L-selectin ligands as a therapeutic target in chronic inflammation. Trends Immunol. 2006, 27, 559–565. [Google Scholar] [CrossRef]
- Arata-Kawai, H.; Singer, M.S.; Bistrup, A.; Zante, A.; Wang, Y.Q.; Ito, Y.; Bao, X.; Hemmerich, S.; Fukuda, M.; Rosen, S.D. Functional contributions of N- and O-glycans to L-selectin ligands in murine and human lymphoid organs. Am. J. Pathol. 2011, 178, 423–433. [Google Scholar] [CrossRef]
- Takeda-Uchimura, Y.; Ikezaki, M.; Akama, T.O.; Nishioka, K.; Ihara, Y.; Allain, F.; Nishitsuji, K.; Uchimura, K. Complementary Role of GlcNAc6ST2 and GlcNAc6ST3 in Synthesis of CL40-Reactive Sialylated and Sulfated Glycans in the Mouse Pleural Mesothelium. Molecules 2022, 27, 4543. [Google Scholar] [CrossRef]
- Kawabe, K.; Tateyama, D.; Toyoda, H.; Kawasaki, N.; Hashii, N.; Nakao, H.; Matsumoto, S.; Nonaka, M.; Matsumura, H.; Hirose, Y.; et al. A novel antibody for human induced pluripotent stem cells and embryonic stem cells recognizes a type of keratan sulfate lacking oversulfated structures. Glycobiology 2013, 23, 322–336. [Google Scholar] [CrossRef]
- Nakao, H.; Nagai, Y.; Kojima, A.; Toyoda, H.; Kawasaki, N.; Kawasaki, T. Binding specificity of R-10G and TRA-1-60/81, and substrate specificity of keratanase II studied with chemically synthesized oligosaccharides. Glycoconj. J. 2017, 34, 789–795. [Google Scholar] [CrossRef]
- Wu, N.; Silva, L.M.; Liu, Y.; Zhang, Y.; Gao, C.; Zhang, F.; Fu, L.; Peng, Y.; Linhardt, R.; Kawasaki, T.; et al. Glycan Markers of Human Stem Cells Assigned with Beam Search Arrays. Mol. Cell Proteom. 2019, 18, 1981–2002. [Google Scholar] [CrossRef]
- Nakao, H.; Yamaguchi, T.; Kawabata, K.; Higashi, K.; Nonaka, M.; Tuiji, M.; Nagai, Y.; Toyoda, H.; Yamaguchi, Y.; Kawasaki, N.; et al. Characterization of novel antibodies that recognize sialylated keratan sulfate and lacto-N-fucopentaose I on human induced pluripotent cells: Comparison with existing antibodies. Glycobiology 2023, 33, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Takeda-Uchimura, Y.; Uchimura, K.; Sugimura, T.; Yanagawa, Y.; Kawasaki, T.; Komatsu, Y.; Kadomatsu, K. Requirement of keratan sulfate proteoglycan phosphacan with a specific sulfation pattern for critical period plasticity in the visual cortex. Exp. Neurol. 2015, 274, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Narentuya; Takeda-Uchimura, Y.; Foyez, T.; Zhang, Z.; Akama, T.O.; Yagi, H.; Kato, K.; Komatsu, Y.; Kadomatsu, K.; Uchimura, K. GlcNAc6ST3 is a keratan sulfate sulfotransferase for the protein-tyrosine phosphatase PTPRZ in the adult brain. Sci. Rep. 2019, 9, 4387. [Google Scholar] [CrossRef] [PubMed]
- Takeda-Uchimura, Y.; Nishitsuji, K.; Ikezaki, M.; Akama, T.O.; Ihara, Y.; Allain, F.; Uchimura, K. Beta3Gn-T7 Is a Keratan Sulfate beta1,3 N-Acetylglucosaminyltransferase in the Adult Brain. Front. Neuroanat. 2022, 16, 813841. [Google Scholar] [CrossRef] [PubMed]
- Patnode, M.L.; Cheng, C.W.; Chou, C.C.; Singer, M.S.; Elin, M.S.; Uchimura, K.; Crocker, P.R.; Khoo, K.H.; Rosen, S.D. Galactose 6-O-sulfotransferases are not required for the generation of Siglec-F ligands in leukocytes or lung tissue. J. Biol. Chem. 2013, 288, 26533–26545. [Google Scholar] [CrossRef]
- Uchimura, K.; Muramatsu, H.; Kadomatsu, K.; Fan, Q.W.; Kurosawa, N.; Mitsuoka, C.; Kannagi, R.; Habuchi, O.; Muramatsu, T. Molecular cloning and characterization of an N-acetylglucosamine-6-O-sulfotransferase. J. Biol. Chem. 1998, 273, 22577–22583. [Google Scholar] [CrossRef]
- Li, X.; Tedder, T.F. CHST1 and CHST2 sulfotransferases expressed by human vascular endothelial cells: cDNA cloning, expression, and chromosomal localization. Genomics 1999, 55, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Kadomatsu, K.; El-Fasakhany, F.M.; Singer, M.S.; Izawa, M.; Kannagi, R.; Takeda, N.; Rosen, S.D.; Muramatsu, T. N-acetylglucosamine 6-O-sulfotransferase-1 regulates expression of L-selectin ligands and lymphocyte homing. J. Biol. Chem. 2004, 279, 35001–35008. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Gauguet, J.M.; Singer, M.S.; Tsay, D.; Kannagi, R.; Muramatsu, T.; von Andrian, U.H.; Rosen, S.D. A major class of L-selectin ligands is eliminated in mice deficient in two sulfotransferases expressed in high endothelial venules. Nat. Immunol. 2005, 6, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, H.; Petryniak, B.; Hiraoka, N.; Mitoma, J.; Huckaby, V.; Nakayama, J.; Uchimura, K.; Kadomatsu, K.; Muramatsu, T.; Lowe, J.B.; et al. N-acetylglucosamine-6-O-sulfotransferases 1 and 2 cooperatively control lymphocyte homing through L-selectin ligand biosynthesis in high endothelial venules. Nat. Immunol. 2005, 6, 1096–1104. [Google Scholar] [CrossRef]
- Zhang, H.; Muramatsu, T.; Murase, A.; Yuasa, S.; Uchimura, K.; Kadomatsu, K. N-Acetylglucosamine 6-O-sulfotransferase-1 is required for brain keratan sulfate biosynthesis and glial scar formation after brain injury. Glycobiology 2006, 16, 702–710. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, Z.; Takeda-Uchimura, Y.; Foyez, T.; Ohtake-Niimi, S.; Narentuya; Akatsu, H.; Nishitsuji, K.; Michikawa, M.; Wyss-Coray, T.; Kadomatsu, K.; et al. Deficiency of a sulfotransferase for sialic acid-modified glycans mitigates Alzheimer’s pathology. Proc. Natl. Acad. Sci. USA 2017, 114, E2947–E2954. [Google Scholar] [CrossRef]
- Bistrup, A.; Bhakta, S.; Lee, J.K.; Belov, Y.Y.; Gunn, M.D.; Zuo, F.R.; Huang, C.C.; Kannagi, R.; Rosen, S.D.; Hemmerich, S. Sulfotransferases of two specificities function in the reconstitution of high endothelial cell ligands for L-selectin. J. Cell Biol. 1999, 145, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, N.; Petryniak, B.; Nakayama, J.; Tsuboi, S.; Suzuki, M.; Yeh, J.C.; Izawa, D.; Tanaka, T.; Miyasaka, M.; Lowe, J.B.; et al. A novel, high endothelial venule-specific sulfotransferase expresses 6-sulfo sialyl Lewis(x), an L-selectin ligand displayed by CD34. Immunity 1999, 11, 79–89. [Google Scholar] [CrossRef]
- Hemmerich, S.; Bistrup, A.; Singer, M.S.; van Zante, A.; Lee, J.K.; Tsay, D.; Peters, M.; Carminati, J.L.; Brennan, T.J.; Carver-Moore, K.; et al. Sulfation of L-selectin ligands by an HEV-restricted sulfotransferase regulates lymphocyte homing to lymph nodes. Immunity 2001, 15, 237–247. [Google Scholar] [CrossRef]
- Lee, J.K.; Bhakta, S.; Rosen, S.D.; Hemmerich, S. Cloning and characterization of a mammalian N-acetylglucosamine-6-sulfotransferase that is highly restricted to intestinal tissue. Biochem. Biophys. Res. Commun. 1999, 263, 543–549. [Google Scholar] [CrossRef]
- Akama, T.O.; Nakayama, J.; Nishida, K.; Hiraoka, N.; Suzuki, M.; McAuliffe, J.; Hindsgaul, O.; Fukuda, M.; Fukuda, M.N. Human corneal GlcNac 6-O-sulfotransferase and mouse intestinal GlcNac 6-O-sulfotransferase both produce keratan sulfate. J. Biol. Chem. 2001, 276, 16271–16278. [Google Scholar] [CrossRef]
- Hayashida, Y.; Akama, T.O.; Beecher, N.; Lewis, P.; Young, R.D.; Meek, K.M.; Kerr, B.; Hughes, C.E.; Caterson, B.; Tanigami, A.; et al. Matrix morphogenesis in cornea is mediated by the modification of keratan sulfate by GlcNAc 6-O-sulfotransferase. Proc. Natl. Acad. Sci. USA 2006, 103, 13333–13338. [Google Scholar] [CrossRef]
- Kitagawa, H.; Fujita, M.; Ito, N.; Sugahara, K. Molecular cloning and expression of a novel chondroitin 6-O-sulfotransferase. J. Biol. Chem. 2000, 275, 21075–21080. [Google Scholar] [CrossRef]
- Bhakta, S.; Bartes, A.; Bowman, K.G.; Kao, W.M.; Polsky, I.; Lee, J.K.; Cook, B.N.; Bruehl, R.E.; Rosen, S.D.; Bertozzi, C.R.; et al. Sulfation of N-acetylglucosamine by chondroitin 6-sulfotransferase 2 (GST-5). J. Biol. Chem. 2000, 275, 40226–40234. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Fasakhany, F.; Kadomatsu, K.; Matsukawa, T.; Yamakawa, T.; Kurosawa, N.; Muramatsu, T. Diversity of N-acetylglucosamine-6-O-sulfotransferases: Molecular cloning of a novel enzyme with different distribution and specificities. Biochem. Biophys. Res. Commun. 2000, 274, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Jung, S.; Zhao, J.; Kasinath, V.; Ichimura, T.; Joseph, J.; Fiorina, P.; Liss, A.S.; Shah, K.; Annabi, N.; et al. Simultaneous targeting of primary tumor, draining lymph node, and distant metastases through high endothelial venule-targeted delivery. Nano Today 2021, 36, 101045. [Google Scholar] [CrossRef]
- Akama, T.O.; Nishida, K.; Nakayama, J.; Watanabe, H.; Ozaki, K.; Nakamura, T.; Dota, A.; Kawasaki, S.; Inoue, Y.; Maeda, N.; et al. Macular corneal dystrophy type I and type II are caused by distinct mutations in a new sulphotransferase gene. Nat. Genet. 2000, 26, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Hemmerich, S.; Lee, J.K.; Bhakta, S.; Bistrup, A.; Ruddle, N.R.; Rosen, S.D. Chromosomal localization and genomic organization for the galactose/ N-acetylgalactosamine/N-acetylglucosamine 6-O-sulfotransferase gene family. Glycobiology 2001, 11, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, K.; Hayashida, Y.; Nishida, K.; Akama, T.O. Enzymes responsible for synthesis of corneal keratan sulfate glycosaminoglycans. J. Biol. Chem. 2007, 282, 30085–30096. [Google Scholar] [CrossRef]
- Fukuta, M.; Inazawa, J.; Torii, T.; Tsuzuki, K.; Shimada, E.; Habuchi, O. Molecular cloning and characterization of human keratan sulfate Gal-6-sulfotransferase. J. Biol. Chem. 1997, 272, 32321–32328. [Google Scholar] [CrossRef]
- Torii, T.; Fukuta, M.; Habuchi, O. Sulfation of sialyl N-acetyllactosamine oligosaccharides and fetuin oligosaccharides by keratan sulfate Gal-6-sulfotransferase. Glycobiology 2000, 10, 203–211. [Google Scholar] [CrossRef]
- Akama, T.O.; Misra, A.K.; Hindsgaul, O.; Fukuda, M.N. Enzymatic synthesis in vitro of the disulfated disaccharide unit of corneal keratan sulfate. J. Biol. Chem. 2002, 277, 42505–42513. [Google Scholar] [CrossRef]
- Patnode, M.L.; Yu, S.Y.; Cheng, C.W.; Ho, M.Y.; Tegesjo, L.; Sakuma, K.; Uchimura, K.; Khoo, K.H.; Kannagi, R.; Rosen, S.D. KSGal6ST generates galactose-6-O-sulfate in high endothelial venules but does not contribute to L-selectin-dependent lymphocyte homing. Glycobiology 2013, 23, 381–394. [Google Scholar] [CrossRef]
- Hoshino, H.; Foyez, T.; Ohtake-Niimi, S.; Takeda-Uchimura, Y.; Michikawa, M.; Kadomatsu, K.; Uchimura, K. KSGal6ST is essential for the 6-sulfation of galactose within keratan sulfate in early postnatal brain. J. Histochem. Cytochem. 2014, 62, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Foyez, T.; Takeda-Uchimura, Y.; Ishigaki, S.; Narentuya; Zhang, Z.; Sobue, G.; Kadomatsu, K.; Uchimura, K. Microglial keratan sulfate epitope elicits in central nervous tissues of transgenic model mice and patients with amyotrophic lateral sclerosis. Am. J. Pathol. 2015, 185, 3053–3065. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Vos, G.M.; Huang, C.; Chapla, D.; Kimpel, A.L.M.; Moremen, K.W.; de Vries, R.P.; Boons, G.-J. Exploiting Substrate Specificities of 6-O-Sulfotransferases to Enzymatically Synthesize Keratan Sulfate Oligosaccharides. JACS Au 2023, 3, 3155–3164. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, M.; Uchimura, K.; Nakashima, K.; Kato, M.; Kimata, K.; Shinomura, T.; Habuchi, O. Molecular cloning and expression of chick chondrocyte chondroitin 6-sulfotransferase. J. Biol. Chem. 1995, 270, 18575–18580. [Google Scholar] [CrossRef] [PubMed]
- Habuchi, O.; Hirahara, Y.; Uchimura, K.; Fukuta, M. Enzymatic sulfation of galactose residue of keratan sulfate by chondroitin 6-sulfotransferase. Glycobiology 1996, 6, 51–57. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Uchimura, K.; Kadomatsu, K.; Nishimura, H.; Muramatsu, H.; Nakamura, E.; Kurosawa, N.; Habuchi, O.; El-Fasakhany, F.M.; Yoshikai, Y.; Muramatsu, T. Functional analysis of the chondroitin 6-sulfotransferase gene in relation to lymphocyte subpopulations, brain development, and oversulfated chondroitin sulfates. J. Biol. Chem. 2002, 277, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.N.; Matsumura, G. Endo-beta-galactosidase of Escherichia freundii. Purification and endoglycosidic action on keratan sulfates, oligosaccharides, and blood group active glycoprotein. J. Biol. Chem. 1976, 251, 6218–6225. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.N. Endo-β-Galactosidases and Keratanase. Curr. Protoc. Mol. Biol. 1995, 32, 17. [Google Scholar] [CrossRef]
- Caterson, B.; Christner, J.E.; Baker, J.R. Identification of a monoclonal antibody that specifically recognizes corneal and skeletal keratan sulfate. Monoclonal antibodies to cartilage proteoglycan. J. Biol. Chem. 1983, 258, 8848–8854. [Google Scholar] [CrossRef]
- Angelidis, I.; Simon, L.M.; Fernandez, I.E.; Strunz, M.; Mayr, C.H.; Greiffo, F.R.; Tsitsiridis, G.; Ansari, M.; Graf, E.; Strom, T.M.; et al. An atlas of the aging lung mapped by single cell transcriptomics and deep tissue proteomics. Nat. Commun. 2019, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, N.; Ikezaki, M.; Nishikawa, T.; Namba, N.; Ohgita, T.; Saito, H.; Ihara, Y.; Shimanouchi, T.; Ino, K.; Uchimura, K.; et al. Sulfated glycosaminoglycans mediate prion-like behavior of p53 aggregates. Proc. Natl. Acad. Sci. USA 2020, 117, 33225–33234. [Google Scholar] [CrossRef] [PubMed]
- Pedram, K.; Laqtom, N.N.; Shon, D.J.; Di Spiezio, A.; Riley, N.M.; Saftig, P.; Abu-Remaileh, M.; Bertozzi, C.R. Lysosomal cathepsin D mediates endogenous mucin glycodomain catabolism in mammals. Proc. Natl. Acad. Sci. USA 2022, 119, e2117105119. [Google Scholar] [CrossRef]
- Uchimura, K.; El-Fasakhany, F.M.; Hori, M.; Hemmerich, S.; Blink, S.E.; Kansas, G.S.; Kanamori, A.; Kumamoto, K.; Kannagi, R.; Muramatsu, T. Specificities of N-acetylglucosamine-6-O-sulfotransferases in relation to L-selectin ligand synthesis and tumor-associated enzyme expression. J. Biol. Chem. 2002, 277, 3979–3984. [Google Scholar] [CrossRef]
- Werlang, C.A.; Chen, W.G.; Aoki, K.; Wheeler, K.M.; Tymm, C.; Mileti, C.J.; Burgos, A.C.; Kim, K.; Tiemeyer, M.; Ribbeck, K. Mucin O-glycans suppress quorum-sensing pathways and genetic transformation in Streptococcus mutans. Nat. Microbiol. 2021, 6, 574–583. [Google Scholar] [CrossRef]
- Brown, G.M.; Huckerby, T.N.; Nieduszynski, I.A. Oligosaccharides derived by keratanase II digestion of bovine articular cartilage keratan sulphates. Eur. J. Biochem. 1994, 224, 281–308. [Google Scholar] [CrossRef]
- Bochner, B.S.; Alvarez, R.A.; Mehta, P.; Bovin, N.V.; Blixt, O.; White, J.R.; Schnaar, R.L. Glycan array screening reveals a candidate ligand for Siglec-8. J. Biol. Chem. 2005, 280, 4307–4312. [Google Scholar] [CrossRef] [PubMed]
- Tateno, H.; Crocker, P.R.; Paulson, J.C. Mouse Siglec-F and human Siglec-8 are functionally convergent paralogs that are selectively expressed on eosinophils and recognize 6′-sulfo-sialyl Lewis X as a preferred glycan ligand. Glycobiology 2005, 15, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Propster, J.M.; Yang, F.; Rabbani, S.; Ernst, B.; Allain, F.H.; Schubert, M. Structural basis for sulfation-dependent self-glycan recognition by the human immune-inhibitory receptor Siglec-8. Proc. Natl. Acad. Sci. USA 2016, 113, E4170–E4179. [Google Scholar] [CrossRef] [PubMed]
- Bull, C.; Nason, R.; Sun, L.; Van Coillie, J.; Madriz Sorensen, D.; Moons, S.J.; Yang, Z.; Arbitman, S.; Fernandes, S.M.; Furukawa, S.; et al. Probing the binding specificities of human Siglecs by cell-based glycan arrays. Proc. Natl. Acad. Sci. USA 2021, 118, e2026102118. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Enterina, J.R.; Bui, D.T.; Mozaneh, F.; Lin, P.H.; Nitin; Kuo, C.W.; Rodrigues, E.; Bhattacherjee, A.; Raeisimakiani, P.; et al. Carbohydrate Sulfation As a Mechanism for Fine-Tuning Siglec Ligands. ACS Chem. Biol. 2021, 16, 2673–2689. [Google Scholar] [CrossRef] [PubMed]
- Kiwamoto, T.; Katoh, T.; Evans, C.M.; Janssen, W.J.; Brummet, M.E.; Hudson, S.A.; Zhu, Z.; Tiemeyer, M.; Bochner, B.S. Endogenous airway mucins carry glycans that bind Siglec-F and induce eosinophil apoptosis. J. Allergy Clin. Immunol. 2015, 135, 1329–1340.e1329. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.; Porell, R.N.; Fernandes, S.M.; Wei, Y.; Yu, H.; Carroll, D.J.; McBride, R.; Paulson, J.C.; Tiemeyer, M.; Aoki, K.; et al. Sialylated keratan sulfate proteoglycans are Siglec-8 ligands in human airways. Glycobiology 2018, 28, 786–801. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.; Li, T.A.; Porell, R.N.; Fernandes, S.M.; Tarbox, H.E.; Lee, H.S.; Aoki, K.; Tiemeyer, M.; Kim, J.; Schnaar, R.L. Isolation, identification, and characterization of the human airway ligand for the eosinophil and mast cell immunoinhibitory receptor Siglec-8. J. Allergy Clin. Immunol. 2021, 147, 1442–1452. [Google Scholar] [CrossRef]
- Kaneko, O.; Gong, L.; Zhang, J.; Hansen, J.K.; Hassan, R.; Lee, B.; Ho, M. A binding domain on mesothelin for CA125/MUC16. J. Biol. Chem. 2009, 284, 3739–3749. [Google Scholar] [CrossRef]
- Rump, A.; Morikawa, Y.; Tanaka, M.; Minami, S.; Umesaki, N.; Takeuchi, M.; Miyajima, A. Binding of ovarian cancer antigen CA125/MUC16 to mesothelin mediates cell adhesion. J. Biol. Chem. 2004, 279, 9190–9198. [Google Scholar] [CrossRef]
- Gubbels, J.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-MUC16 binding is a high affinity, N-glycan dependent interaction that facilitates peritoneal metastasis of ovarian tumors. Mol. Cancer 2006, 5, 50. [Google Scholar] [CrossRef]
- Belisle, J.A.; Horibata, S.; Jennifer, G.A.; Petrie, S.; Kapur, A.; Andre, S.; Gabius, H.J.; Rancourt, C.; Connor, J.; Paulson, J.C.; et al. Identification of Siglec-9 as the receptor for MUC16 on human NK cells, B cells, and monocytes. Mol. Cancer 2010, 9, 118. [Google Scholar] [CrossRef]
- Das, S.; Batra, S.K. Understanding the Unique Attributes of MUC16 (CA125): Potential Implications in Targeted Therapy. Cancer Res. 2015, 75, 4669–4674. [Google Scholar] [CrossRef]
- Hanson, R.L.; Hollingsworth, M.A. Functional Consequences of Differential O-glycosylation of MUC1, MUC4, and MUC16 (Downstream Effects on Signaling). Biomolecules 2016, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Saldova, R.; Struwe, W.B.; Wynne, K.; Elia, G.; Duffy, M.J.; Rudd, P.M. Exploring the glycosylation of serum CA125. Int. J. Mol. Sci. 2013, 14, 15636–15654. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Sakai, Y.; Hoshino, H.; Umeda, Y.; Kawashima, H.; Sekido, Y.; Ishizuka, T.; Kobayashi, M. Sulfated Glycans Recognized by S1 Monoclonal Antibody can Serve as a Diagnostic Marker for Malignant Pleural Mesothelioma. Lung 2022, 200, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hosono-Fukao, T.; Ohtake-Niimi, S.; Hoshino, H.; Britschgi, M.; Akatsu, H.; Hossain, M.M.; Nishitsuji, K.; van Kuppevelt, T.H.; Kimata, K.; Michikawa, M.; et al. Heparan sulfate subdomains that are degraded by Sulf accumulate in cerebral amyloid ss plaques of Alzheimer’s disease: Evidence from mouse models and patients. Am. J. Pathol. 2012, 180, 2056–2067. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Other Nomenclature | Gene | Substrate | KS/Sulfated LacNAc Oligosaccharide Synthesis | Refs. |
|---|---|---|---|---|---|
| GlcNAc6ST1 * | N-acetylglucosamine 6-O-sulfotransferase (GlcNAc6ST); Carbohydrate sulfotransferase 2 (CHST2) | CHST2 | GlcNAc | Yes | [8,12,13,14,15,16,17,18] |
| GlcNAc6ST2 * | High endothelial cell N-acetylglucosamine 6-O-sulfotransferase (HEC-GlcNAc6ST); L-Selectin ligand sulfotransferase (LSST); Carbohydrate sulfotransferase 4 (CHST4) | CHST4 | GlcNAc | Yes (shown in this article) | [15,16,19,20,21] |
| GlcNAc6ST3 | Intestinal N-acetylglucosamine 6-O-sulfotransferase (I-GlcNAc6ST); Carbohydrate sulfotransferase 5 (CHST5) | CHST5 ** | GlcNAc | Yes | [9,22,23,24] |
| GlcNAc6ST4 | N-acetylglucosamine 6-O-sulfotransferase-4; Chondroitin 6-O-sulfotransferase-2 (C6ST-2); Carbohydrate sulfotransferase 7 (CHST7) | CHST7 | GlcNAc, GalNAc | N.d. # | [9,25,26,27,28] |
| GlcNAc6ST5 | Cornial N-acetylglucosamine 6-O-sulfotransferase (C-GlcNAc6ST); Carbohydrate sulfotransferase 6 (CHST6) | CHST6 ** | GlcNAc | Yes | [23,29,30,31] |
| KSGal6ST | Keratan sulfate galactose-6-O-sulfotransferase; Carbohydrate sulfotransferase 1 (CHST1) | CHST1 | Gal | Yes | [19,32,33,34,35,36,37,38] |
| C6ST1 | Chondroitin-6-O-sulfotransferase (C6ST); Carbohydrate sulfotransferase 3 (CHST3) | CHST3 | GalNAc, Gal | Yes | [11,39,40,41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takeda-Uchimura, Y.; Ikezaki, M.; Akama, T.O.; Ihara, Y.; Allain, F.; Nishitsuji, K.; Uchimura, K. GlcNAc6ST2/CHST4 Is Essential for the Synthesis of R-10G-Reactive Keratan Sulfate/Sulfated N-Acetyllactosamine Oligosaccharides in Mouse Pleural Mesothelium. Molecules 2024, 29, 764. https://doi.org/10.3390/molecules29040764
Takeda-Uchimura Y, Ikezaki M, Akama TO, Ihara Y, Allain F, Nishitsuji K, Uchimura K. GlcNAc6ST2/CHST4 Is Essential for the Synthesis of R-10G-Reactive Keratan Sulfate/Sulfated N-Acetyllactosamine Oligosaccharides in Mouse Pleural Mesothelium. Molecules. 2024; 29(4):764. https://doi.org/10.3390/molecules29040764
Chicago/Turabian StyleTakeda-Uchimura, Yoshiko, Midori Ikezaki, Tomoya O. Akama, Yoshito Ihara, Fabrice Allain, Kazuchika Nishitsuji, and Kenji Uchimura. 2024. "GlcNAc6ST2/CHST4 Is Essential for the Synthesis of R-10G-Reactive Keratan Sulfate/Sulfated N-Acetyllactosamine Oligosaccharides in Mouse Pleural Mesothelium" Molecules 29, no. 4: 764. https://doi.org/10.3390/molecules29040764
APA StyleTakeda-Uchimura, Y., Ikezaki, M., Akama, T. O., Ihara, Y., Allain, F., Nishitsuji, K., & Uchimura, K. (2024). GlcNAc6ST2/CHST4 Is Essential for the Synthesis of R-10G-Reactive Keratan Sulfate/Sulfated N-Acetyllactosamine Oligosaccharides in Mouse Pleural Mesothelium. Molecules, 29(4), 764. https://doi.org/10.3390/molecules29040764