
The Potential of Nuclear Pore Complexes in Cancer Therapy
 , , , and
, , , and 
Abstract
1. Introduction
1.1. NPCs’ Structure
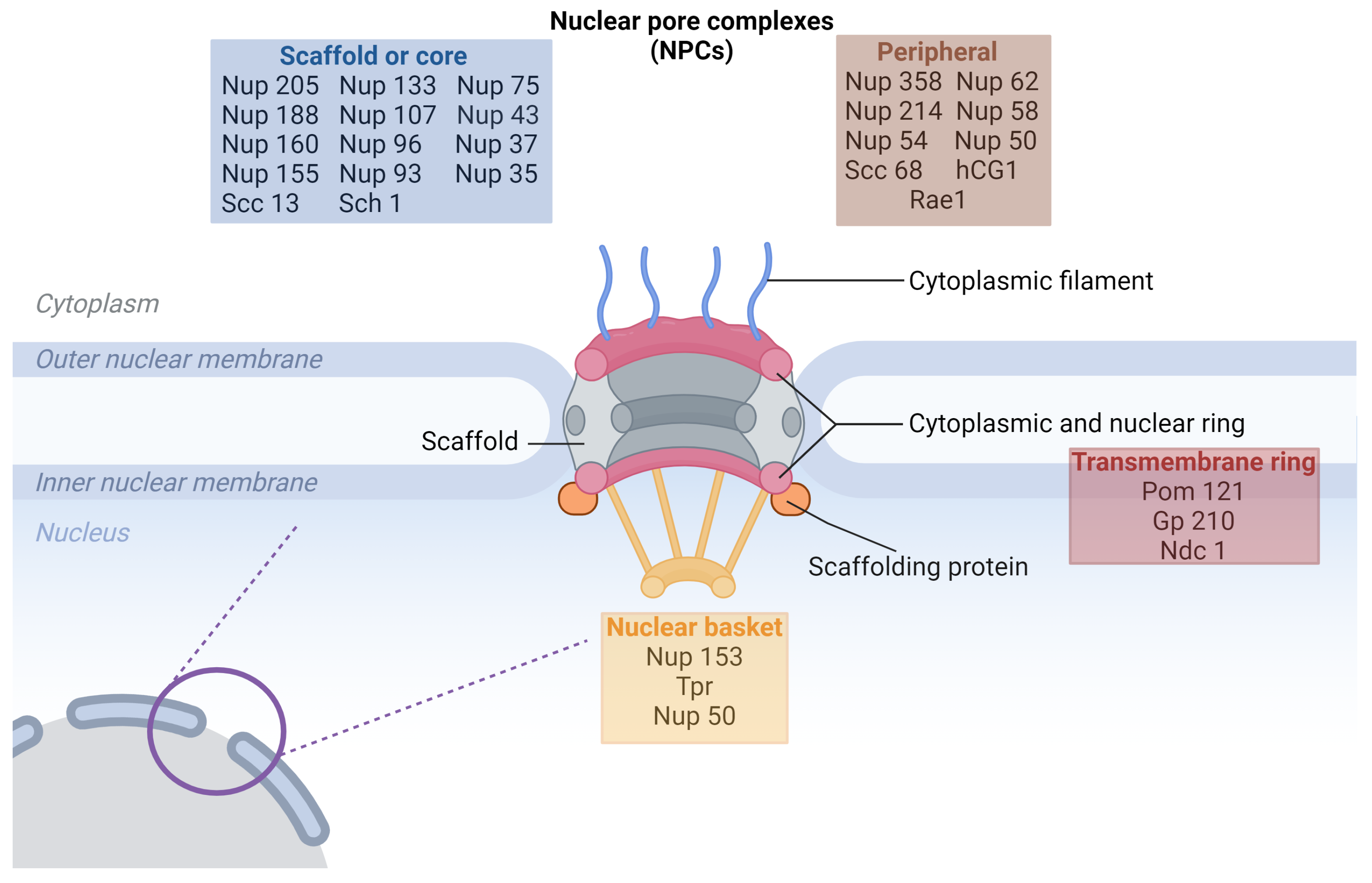

{kind=link}
{kind=link}
{kind=link}
| Protein | Function | Reference |
|---|---|---|
| scaffold | ||
| Nup93/Nup205 complex (Nup93, Nup205, Nup188, Nup155, Nup35), | forming of central pore | [11] |
| Nup107/Nup160 complex (Nup107, Nup160 Nup133, Nup 96, Nup43, Nup37, Nup 85, Sec13, Seh1, Elys) | forming of cytoplasmic and nuclear ring | [11] |
| transmembrane | ||
| Pom121, Gp210, Ndc1 | linking of NPC to the nuclear envelope | [11] |
| peripheral | ||
| Nup358/RanBP2, Nup214, Nup 88 | forming of cytoplasmic filaments | [11] |
| Nup153, Tpr | forming of nuclear basket | [11] |
| Nup98, Rae1 | shuttling transport factors | [24] |
| FG-domain-anchoring Nups | ||
| Nup62 complex (Nup62, Nup58, Nup54, Nup45) | selective permeability barrier regulation of receptor-mediated translocation | [11] |
1.2. NPCs’ Function
2. Targeting Nuclear Pore Complexes
2.1. Selectivity
2.2. Drug Resistance
2.3. Combination Therapies and Nanoparticles
2.4. Potential Mechanisms
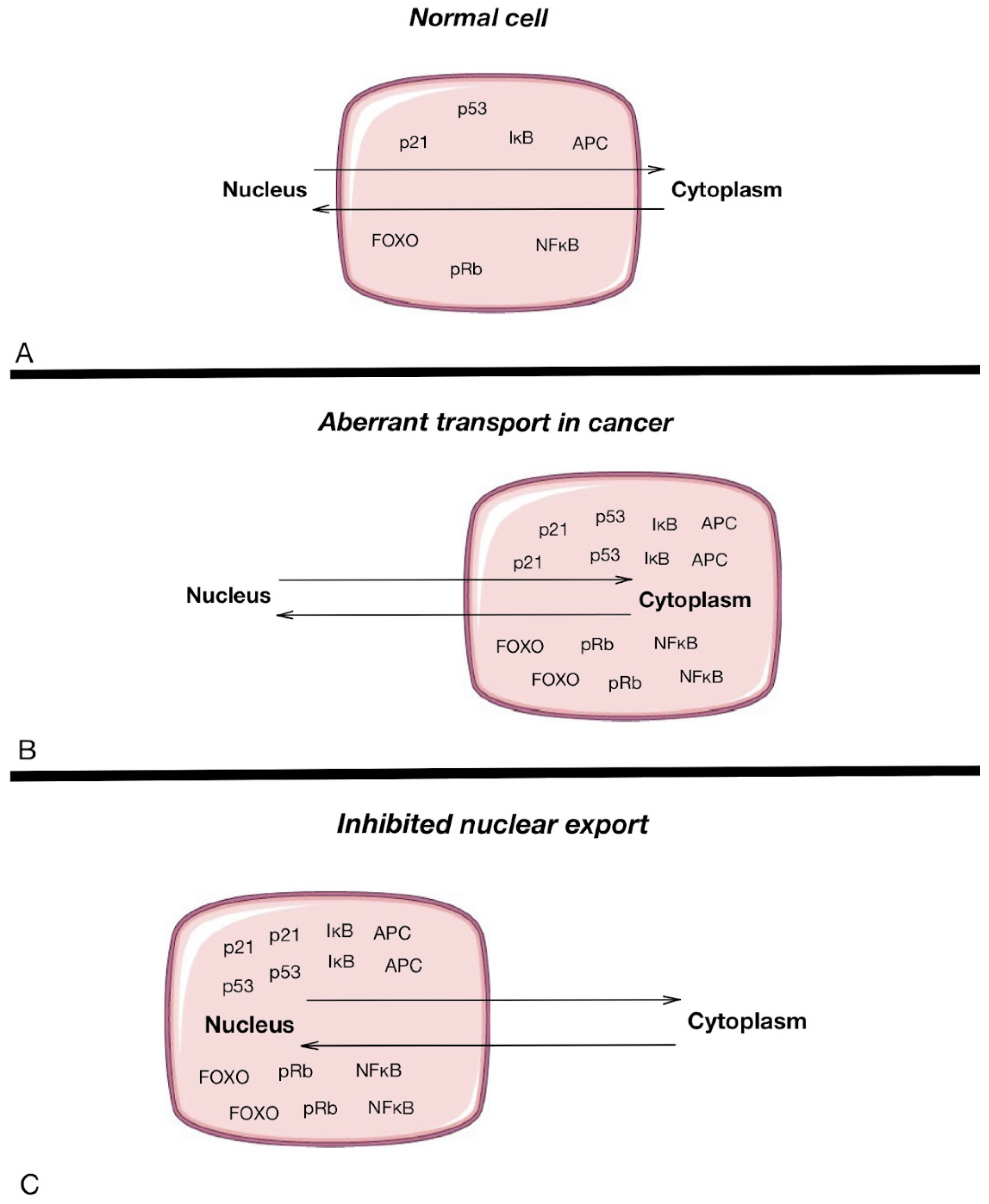
3. Defining NPCs as Potential Targets in Oncotherapy
4. Mislocalization of Transcription Factors Independent of NPC Defects
5. Chemotherapy and NPC Inhibition
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cook, A.; Bono, F.; Jinek, M.; Conti, E. Structural biology of nucleocytoplasmic transport. Annu. Rev. Biochem. 2007, 76, 647–671. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.S.; Wong, R.W. Targeting Nucleoporin POM121-Importin β Axis in Prostate Cancer. Cell Chem. Biol. 2018, 25, 1056–1058. [Google Scholar] [CrossRef] [PubMed]
- Cronshaw, J.M.; Matunis, M.J. The nuclear pore complex: Disease associations and functional correlations. Trends Endocrinol. Metab. 2004, 15, 34–39. [Google Scholar] [CrossRef]
- Gu, Y. The nuclear pore complex: A strategic platform for regulating cell signaling. New Phytol. 2018, 219, 25–30. [Google Scholar] [CrossRef]
- Fallini, C.; Khalil, B.; Smith, C.; Rossoll, W. Traffic jam at the nuclear pore: All roads lead to nucleocytoplasmic transport defects in ALS/FTD. Neurobiol. Dis. 2020, 140, 104835. [Google Scholar] [CrossRef]
- Hill, R.; Cautain, B.; De Pedro, N.; Link, W. Targeting nucleocytoplasmic transport in cancer therapy. Oncotarget 2014, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- D’angelo, M.; Hetzer, M. The role of the nuclear envelope in cellular organization. Cell. Mol. Life Sci. 2006, 63, 316–332. [Google Scholar] [CrossRef]
- Kabachinski, G.; Schwartz, T.U. The nuclear pore complex—Structure and function at a glance. J. Cell Sci. 2015, 128, 423–429. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, M.A.; Hetzer, M.W. Structure, dynamics and function of nuclear pore complexes. Trends Cell Biol. 2008, 18, 456–466. [Google Scholar] [CrossRef]
- Eibauer, M.; Pellanda, M.; Turgay, Y.; Dubrovsky, A.; Wild, A.; Medalia, O. Structure and gating of the nuclear pore complex. Nat. Commun. 2015, 6, 7532. [Google Scholar] [CrossRef]
- Ibarra, A.; Hetzer, M.W. Nuclear pore proteins and the control of genome functions. Genes Dev. 2015, 29, 337–349. [Google Scholar] [CrossRef]
- Lim, R.Y.; Aebi, U.; Fahrenkrog, B. Towards reconciling structure and function in the nuclear pore complex. Histochem. Cell Biol. 2008, 129, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Gravina, G.L.; Senapedis, W.; McCauley, D.; Baloglu, E.; Shacham, S.; Festuccia, C. Nucleo-cytoplasmic transport as a therapeutic target of cancer. J. Hematol. Oncol. 2014, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Tran, E.J.; King, M.C.; Corbett, A.H. Macromolecular transport between the nucleus and the cytoplasm: Advances in mechanism and emerging links to disease. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 2784–2795. [Google Scholar] [CrossRef] [PubMed]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. The molecular architecture of the nuclear pore complex. Nature 2007, 450, 695–701. [Google Scholar] [CrossRef]
- Menon, B.B.; Sarma, N.J.; Pasula, S.; Deminoff, S.J.; Willis, K.A.; Barbara, K.E.; Andrews, B.; Santangelo, G.M. Reverse recruitment: The Nup84 nuclear pore subcomplex mediates Rap1/Gcr1/Gcr2 transcriptional activation. Proc. Natl. Acad. Sci. USA 2005, 102, 5749–5754. [Google Scholar] [CrossRef]
- Hogarth, C.; Itman, C.; Jans, D.A.; Loveland, K.L. Regulated nucleocytoplasmic transport in spermatogenesis: A driver of cellular differentiation? Bioessays 2005, 27, 1011–1025. [Google Scholar] [CrossRef]
- Weis, K. The nuclear pore complex: Oily spaghetti or gummy bear? Cell 2007, 130, 405–407. [Google Scholar] [CrossRef]
- Denning, D.P.; Patel, S.S.; Uversky, V.; Fink, A.L.; Rexach, M. Disorder in the nuclear pore complex: The FG repeat regions of nucleoporins are natively unfolded. Proc. Natl. Acad. Sci. USA 2003, 100, 2450–2455. [Google Scholar] [CrossRef]
- Capelson, M.; Hetzer, M.W. The role of nuclear pores in gene regulation, development and disease. EMBO Rep. 2009, 10, 697–705. [Google Scholar] [CrossRef]
- Fernandez-Martinez, J.; Phillips, J.; Sekedat, M.D.; Diaz-Avalos, R.; Velazquez-Muriel, J.; Franke, J.D.; Williams, R.; Stokes, D.L.; Chait, B.T.; Sali, A.; et al. Structure–function mapping of a heptameric module in the nuclear pore complex. J. Cell Biol. 2012, 196, 419–434. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, M.A.; Raices, M.; Panowski, S.H.; Hetzer, M.W. Age-dependent deterioration of nuclear pore complexes causes a loss of nuclear integrity in postmitotic cells. Cell 2009, 136, 284–295. [Google Scholar] [CrossRef]
- Ben-Efraim, I.; Frosst, P.D.; Gerace, L. Karyopherin binding interactions and nuclear import mechanism of nuclear pore complex protein Tpr. BMC Cell Biol. 2009, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, C.E.; Fornerod, M.; Kasper, L.H.; Van Deursen, J.M. RAE1 is a shuttling mRNA export factor that binds to a GLEBS-like NUP98 motif at the nuclear pore complex through multiple domains. J. Cell Biol. 1999, 145, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Paci, G.; Caria, J.; Lemke, E.A. Cargo transport through the nuclear pore complex at a glance. J. Cell Sci. 2021, 134, jcs247874. [Google Scholar] [CrossRef] [PubMed]
- Cautain, B.; Hill, R.; de Pedro, N.; Link, W. Components and regulation of nuclear transport processes. FEBS J. 2015, 282, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Paine, P.L.; Moore, L.C.; Horowitz, S.B. Nuclear envelope permeability. Nature 1975, 254, 109–114. [Google Scholar] [CrossRef]
- Andersson, J.; Svirelis, J.; Medin, J.; Järlebark, J.; Hailes, R.; Dahlin, A. Pore performance: Artificial nanoscale constructs that mimic the biomolecular transport of the nuclear pore complex. Nanoscale Adv. 2022, 4, 4925–4937. [Google Scholar] [CrossRef]
- Melchior, F.; Paschal, B.; Evans, J.; Gerace, L. Inhibition of nuclear protein import by nonhydrolyzable analogues of GTP and identification of the small GTPase Ran/TC4 as an essential transport factor. J. Cell Biol. 1993, 123, 1649–1659. [Google Scholar] [CrossRef]
- Görlich, D.; Panté, N.; Kutay, U.; Aebi, U.; Bischoff, F.R. Identification of different roles for RanGDP and RanGTP in nuclear protein import. EMBO J. 1996, 15, 5584–5594. [Google Scholar] [CrossRef]
- Tagliazucchi, M.; Peleg, O.; Kröger, M.; Rabin, Y.; Szleifer, I. Effect of charge, hydrophobicity, and sequence of nucleoporins on the translocation of model particles through the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2013, 110, 3363–3368. [Google Scholar] [CrossRef]
- Jovanovic-Talisman, T.; Zilman, A. Protein transport by the nuclear pore complex: Simple biophysics of a complex biomachine. Biophys. J. 2017, 113, 6–14. [Google Scholar] [CrossRef]
- Zanella, F.; Dos Santos, N.R.; Link, W. Moving to the Core: Spatiotemporal Analysis of Forkhead Box O (FOXO) and Nuclear Factor-κB (NF-κB) Nuclear Translocation. Traffic 2013, 14, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.Y.J.; Chook, Y.M. Atomic basis of CRM1-cargo recognition, release and inhibition. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 27, pp. 52–61. [Google Scholar]
- Matsuda, A.; Mofrad, M.R. On the nuclear pore complex and its emerging role in cellular mechanotransduction. APL Bioeng. 2022, 6, 011504. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, C.E.; Allegretti, M.; Rantos, V.; Goetz, S.K.; Obarska-Kosinska, A.; Zagoriy, I.; Halavatyi, A.; Hummer, G.; Mahamid, J.; Kosinski, J.; et al. Nuclear pores dilate and constrict in cellulo. Science 2021, 374, eabd9776. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, S.; Tempkin, J.O.; Zhang, W.; Politi, A.Z.; Rybina, A.; Hossain, M.J.; Kueblbeck, M.; Callegari, A.; Koch, B.; Morero, N.R.; et al. A quantitative map of nuclear pore assembly reveals two distinct mechanisms. Nature 2023, 613, 575–581. [Google Scholar] [CrossRef]
- McCloskey, A.; Ibarra, A.; Hetzer, M.W. Tpr regulates the total number of nuclear pore complexes per cell nucleus. Genes Dev. 2018, 32, 1321–1331. [Google Scholar] [CrossRef]
- Li, Y.; Bertozzi, A.; Mann, M.R.; Kühn, B. Interdependent changes of nuclear lamins, nuclear pore complexes, and ploidy regulate cellular regeneration and stress response in the heart. Nucleus 2023, 14, 2246310. [Google Scholar] [CrossRef]
- Han, L.; Choudhury, S.; Mich-Basso, J.D.; Ammanamanchi, N.; Ganapathy, B.; Suresh, S.; Khaladkar, M.; Singh, J.; Maehr, R.; Zuppo, D.A.; et al. Lamin B2 levels regulate polyploidization of cardiomyocyte nuclei and myocardial regeneration. Dev. Cell 2020, 53, 42–59. [Google Scholar] [CrossRef]
- Sakuma, S.; Raices, M.; Borlido, J.; Guglielmi, V.; Zhu, E.Y.; D’Angelo, M.A. Inhibition of nuclear pore complex formation selectively induces cancer cell death. Cancer Discov. 2021, 11, 176–193. [Google Scholar] [CrossRef]
- Klughammer, N.; Barth, A.; Dekker, M.; Fragasso, A.; Onck, P.R.; Dekker, C. Diameter dependence of transport through nuclear pore complex mimics studied using optical nanopores. Elife 2024, 12, RP87174. [Google Scholar] [CrossRef] [PubMed]
- Peters, R. Functionalization of a nanopore: The nuclear pore complex paradigm. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- El-Tanani, M.; Dakir, E.H.; Raynor, B.; Morgan, R. Mechanisms of nuclear export in cancer and resistance to chemotherapy. Cancers 2016, 8, 35. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Kalir, T.; Rahaman, J.; Dottino, P.; Kohtz, D.S. Alterations in nuclear pore architecture allow cancer cell entry into or exit from drug-resistant dormancy. Am. J. Pathol. 2012, 180, 375–389. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022. [Google Scholar] [CrossRef]
- Delbaldo, C.; Michiels, S.; Syz, N.; Soria, J.C.; Le Chevalier, T.; Pignon, J.P. Benefits of adding a drug to a single-agent or a 2-agent chemotherapy regimen in advanced non–small-cell lung cancer: A meta-analysis. Jama 2004, 292, 470–484. [Google Scholar] [CrossRef]
- Higby, K.J.; Bischak, M.M.; Campbell, C.A.; Anderson, R.G.; Broskin, S.A.; Foltz, L.E.; Koper, J.A.; Nickle, A.C.; Resendes, K.K. 5-Flurouracil disrupts nuclear export and nuclear pore permeability in a calcium dependent manner. Apoptosis 2017, 22, 393–405. [Google Scholar] [CrossRef]
- Can, G.; Akpinar, B.; Baran, Y.; Zhivotovsky, B.; Olsson, M. 5-Fluorouracil signaling through a calcium–calmodulin-dependent pathway is required for p53 activation and apoptosis in colon carcinoma cells. Oncogene 2013, 32, 4529–4538. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Zhang, X.; Chen, Q.; Liu, M.; Xiang, Y.; Yang, Y.; Xiao, Z.; Huang, J.; Wang, X.; Liu, C.; et al. Nucleus-targeting phototherapy nanodrugs for high-effective anti-cancer treatment. Front. Pharmacol. 2022, 13, 905375. [Google Scholar] [CrossRef]
- Nie, Y.; Fu, G.; Leng, Y. Nuclear Delivery of Nanoparticle-Based Drug Delivery Systems by Nuclear Localization Signals. Cells 2023, 12, 1637. [Google Scholar] [CrossRef]
- Tkachenko, A.; Xie, H.; Coleman, D.; Glomm, W.; Ryan, J.; Anderson, M.; Franzen, S.; Feldheim, D. Multifunctional Gold Nanoparticle-Peptide Complexes for Nuclear Targeting. J. Am. Chem. Soc. 2003, 125, 4700–4701. [Google Scholar] [CrossRef] [PubMed]
- Ozcicek, I.; Aysit, N.; Cakici, C.; Aydeger, A. The effects of surface functionality and size of gold nanoparticles on neuronal toxicity, apoptosis, ROS production and cellular/suborgan biodistribution. Mater. Sci. Eng. C 2021, 128, 4700–4701. [Google Scholar] [CrossRef] [PubMed]
- Skowicki, M.; Tarvirdipour, S.; Kraus, M.; Schoenenberger, C.; Palivan, C. Nanoassemblies designed for efficient nuclear targeting. Adv. Drug Deliv. Rev. 2024, 211, 115354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yue, P.; Peng, Y.; Sun, Y.; Chen, X.; Zhao, Z.; Han, B. Recent advances in drug delivery systems for targeting brain tumors. Drug Deliv. 2023, 30, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, H.; Ye, Y.; Lei, Y.; Islam, R.; Tan, S.; Tong, R.; Miao, Y.B.; Cai, L. Smart nanoparticles for cancer therapy. Signal Transduct. Target. Ther. 2023, 8, 418. [Google Scholar] [CrossRef]
- Rodriguez-Bravo, V.; Pippa, R.; Song, W.M.; Carceles-Cordon, M.; Dominguez-Andres, A.; Fujiwara, N.; Woo, J.; Koh, A.P.; Ertel, A.; Lokareddy, R.K.; et al. Nuclear pores promote lethal prostate cancer by increasing POM121-driven E2F1, MYC, and AR nuclear import. Cell 2018, 174, 1200–1215. [Google Scholar] [CrossRef]
- Chen, S.L.; Qin, Z.Y.; Hu, F.; Wang, Y.; Dai, Y.J.; Liang, Y. The role of the HOXA gene family in acute myeloid leukemia. Genes 2019, 10, 621. [Google Scholar] [CrossRef]
- Fahrenkrog, B. Nucleoporin gene fusions and hematopoietic malignancies. New J. Sci. 2014, 2014, 468306. [Google Scholar] [CrossRef]
- Xu, F.; Xu, H.; Li, Z.; Huang, Y.; Huang, X.; Li, Y.; Zheng, X.; Chen, Y.; Lin, L. Glycolysis-based genes are potential biomarkers in thyroid cancer. Front. Oncol. 2021, 11, 534838. [Google Scholar] [CrossRef]
- Yu, Y.; Farooq, M.S.; Eberhart Meessen, S.; Jiang, Y.; Kato, D.; Zhan, T.; Weiss, C.; Seger, R.; Kang, W.; Zhang, X.; et al. Nuclear pore protein POM121 regulates subcellular localization and transcriptional activity of PPARγ. Cell Death Dis. 2024, 15, 7. [Google Scholar] [CrossRef]
- Amin, R.; Shukla, A.; Zhu, J.J.; Kim, S.; Wang, P.; Tian, S.Z.; Tran, A.D.; Paul, D.; Cappell, S.D.; Burkett, S.; et al. Nuclear pore protein NUP210 depletion suppresses metastasis through heterochromatin-mediated disruption of tumor cell mechanical response. Nat. Commun. 2021, 12, 7216. [Google Scholar] [CrossRef] [PubMed]
- Ernst, S.; Müller-Newen, G. Nucleocytoplasmic shuttling of STATs. A target for intervention? Cancers 2019, 11, 1815. [Google Scholar] [CrossRef] [PubMed]
- Ohkoshi, S.; Yano, M.; Matsuda, Y. Oncogenic role of p21 in hepatocarcinogenesis suggests a new treatment strategy. World J. Gastroenterol. 2015, 21, 12150. [Google Scholar] [CrossRef]
- Ikliptikawati, D.K.; Hirai, N.; Makiyama, K.; Sabit, H.; Kinoshita, M.; Matsumoto, K.; Lim, K.; Meguro-Horike, M.; Horike, S.i.; Hazawa, M.; et al. Nuclear transport surveillance of p53 by nuclear pores in glioblastoma. Cell Rep. 2023, 42, 112882. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.; Wong, C.; Huang, Y.; Li, H. Survivin withdrawal by nuclear export failure as a physiological switch to commit cells to apoptosis. Cell Death Dis. 2010, 1, e57. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.G.; Dawson, J.; Cubitt, C.L.; Baz, R.; Sullivan, D.M. Inhibition of CRM1-dependent nuclear export sensitizes malignant cells to cytotoxic and targeted agents. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 27, pp. 62–73. [Google Scholar]
- Sun, Q.; Chen, X.; Zhou, Q.; Burstein, E.; Yang, S.; Jia, D. Inhibiting cancer cell hallmark features through nuclear export inhibition. Signal Transduct. Target. Ther. 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Nguyen, K.T.; Holloway, M.P.; Altura, R.A. The CRM1 nuclear export protein in normal development and disease. Int. J. Biochem. Mol. Biol. 2012, 3, 137. [Google Scholar]
- Gao, J.; Azmi, A.S.; Aboukameel, A.; Kauffman, M.; Shacham, S.; Abou-Samra, A.B.; Mohammad, R.M. Nuclear retention of Fbw7 by specific inhibitors of nuclear export leads to Notch1 degradation in pancreatic cancer. Oncotarget 2014, 5, 3444. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Lin, M.; Cheng, X.; Zhang, Z.; Deng, S.; Lang, K.; Yang, Z.; Sun, X. KPNB1-mediated nuclear import in cancer. Eur. J. Pharmacol. 2023, 955, 175925. [Google Scholar] [CrossRef]
- Wagstaff, K.M.; Sivakumaran, H.; Heaton, S.M.; Harrich, D.; Jans, D.A. Ivermectin is a specific inhibitor of importin α/β-mediated nuclear import able to inhibit replication of HIV-1 and dengue virus. Biochem. J. 2012, 443, 851–856. [Google Scholar] [CrossRef]
- Hussain, M.K.; Ali, R.; Ahamad, S.; Khan, M.F.; Saquib, M. Ivermectin: A Potential Repurposed Anti-Cancer Therapeutic. In Chemistry and Biological Activities of Ivermectin; Scrivener Publishing LLC: Beverly, MA, USA, 2023; pp. 121–139. [Google Scholar]
- Lombardi, Z.; Gardini, L.; Kashchuk, A.V.; Menconi, A.; Lulli, M.; Tusa, I.; Tubita, A.; Maresca, L.; Stecca, B.; Capitanio, M.; et al. Importin subunit beta-1 mediates ERK5 nuclear translocation, and its inhibition synergizes with ERK5 kinase inhibitors in reducing cancer cell proliferation. Mol. Oncol. 2024. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Peng, Z.; Chen, Y.; Li, H.; Du, M.; Tan, Y.; Zhang, X.; Lu, Z.; Cui, C.P.; Liu, C.H.; et al. Neddylation of PTEN regulates its nuclear import and promotes tumor development. Cell Res. 2021, 31, 291–311. [Google Scholar] [CrossRef] [PubMed]

| Name of the Drug | Action in the Cell | Usage Indications | Reference |
|---|---|---|---|
| Melphalan | Increases DNA damage | Multiple myeloma | [67] |
| Cisplatin | Interferes with DNA replication | Advanced or relapsed cancers, cancers that cannot be treated with more traditional oncological treatments (bladder cancer, cervical cancer, malignant mesothelioma, non-small-cell lung cancer, ovarian cancer, squamous-cell carcinoma of the head and neck, neuroblastoma, and testicular cancer) | [67] |
| Melphalan | Increases DNA damage | Multiple myeloma | [67] |
| Bortezomib | Increases the expression of p53 and IkB levels | Multiple myeloma | [67] |
| Carfilzomib | Increases the expression of p53 and IkB levels | Multiple myeloma | [67] |
| Doxorubicin | Inhibits the progression of topoisomerase II, stops the process of replication | Multiple myeloma | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaitsava, H.; Gachowska, M.; Bartoszewska, E.; Kmiecik, A.; Kulbacka, J. The Potential of Nuclear Pore Complexes in Cancer Therapy. Molecules 2024, 29, 4832. https://doi.org/10.3390/molecules29204832
Zaitsava H, Gachowska M, Bartoszewska E, Kmiecik A, Kulbacka J. The Potential of Nuclear Pore Complexes in Cancer Therapy. Molecules. 2024; 29(20):4832. https://doi.org/10.3390/molecules29204832
Chicago/Turabian StyleZaitsava, Hanna, Martyna Gachowska, Elżbieta Bartoszewska, Alicja Kmiecik, and Julita Kulbacka. 2024. "The Potential of Nuclear Pore Complexes in Cancer Therapy" Molecules 29, no. 20: 4832. https://doi.org/10.3390/molecules29204832
APA StyleZaitsava, H., Gachowska, M., Bartoszewska, E., Kmiecik, A., & Kulbacka, J. (2024). The Potential of Nuclear Pore Complexes in Cancer Therapy. Molecules, 29(20), 4832. https://doi.org/10.3390/molecules29204832