
GC-MS and PCA Analysis of Fatty Acid Profile in Various Ilex Species

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results
2.1. Chemical Composition of Samples
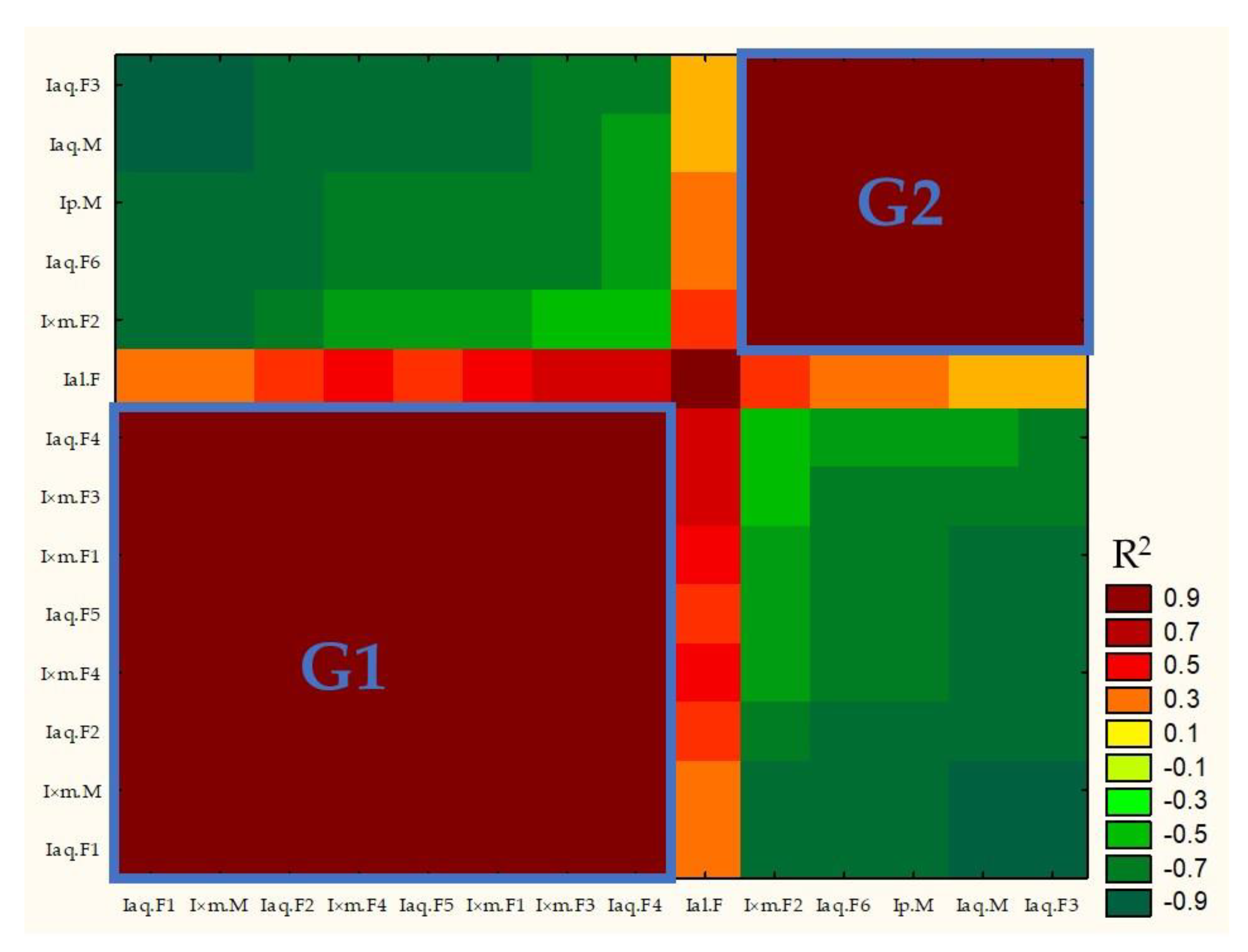
2.2. Statistical Analysis
2.2.1. Principal Components Analysis (PCA)
2.2.2. Analyses of Tendencies in Fatty Acid Profile of Selected Ilex sp.
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Total Lipid Extraction and Fatty Acids Analysis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Czekalski, M. Liściaste Krzewy Ozdobne II; Państwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 2015; pp. 95–98. [Google Scholar]
- Czekalski, M. Krzewy ozdobne mało znane: Ostrokrzew kolczasty. Zieleń Miejska 2015, 3, 28. [Google Scholar]
- Tumiłowicz, J.; Banaszczak, P. Drzewa i krzewy z rodziny Aquifoliaceae w arboretach w Rogowie i Glinnej. Rocz. Dendr 2007, 55, 41–56. [Google Scholar]
- Seneta, W.; Dolatowski, J. Dendrologia; PWN: Warszawa, Poland, 2009. [Google Scholar]
- Galle, F.C. Hollies–The genus Ilex; Timber Press: Portland, OR, USA, 1998. [Google Scholar]
- Czekalski, M. Ostrokrzew Meservy. Zieleń Miejska 2012, 12, 27. [Google Scholar]
- Yao, X.; Zhang, F.; Corlett, R.T. Utilization of the Hollies (Ilex L. spp.): A Review. Forests 2022, 13, 94. [Google Scholar] [CrossRef]
- Cansian, R.L.; Mossi, A.J.; Mosele, S.H.; Toniazzo, G.; Treichel, H.; Paroul, N.; Oliveira, J.V.; Mazutti, M.; Echeverrigaray, S. Genetic conservation and medicinal properties of mate (Ilex paraguariensis St Hil.). Phcog Rev. 2008, 2, 326–338. [Google Scholar]
- Zwyrzykowska, A.; Kupczyński, R.; Jarosz, B.; Szumny, A.; Kucharska, A.Z. Qualitative and quantitative analysis of polyphenolic compounds in Ilex sp. Open Chem. 2015, 13, 1303–1312. [Google Scholar] [CrossRef]
- Paluch, E.; Okińczyc, P.; Zwyrzykowska-Wodzińska, A.; Szperlik, J.; Żarowska, B.; Duda-Madej, A.; Bąbelewski, P.; Włodarczyk, M.; Wojtasik, W.; Kupczyński, R.; et al. Composition and Antimicrobial Activity of Ilex Leaves Water Extracts. Molecules 2021, 26, 7442. [Google Scholar] [CrossRef]
- Balzan, S.; Hernandes, A.; Reichert, C.L.; Donaduzzi, C.; Pires, V.A.; Gasparotto, A.; Cardozo, E.L. Lipid-lowering effects of standardized extracts of Ilex paraguariensis in high-fat-diet rats. Fitoterapia 2013, 86, 115–122. [Google Scholar] [CrossRef]
- Bracesco, N.; Sanchez, A.G.; Contreras, V.; Menini, T.; Gugliucci, A. Recent advances on Ilex paraguariensis research: Minireview. J. Ethnopharmacol. 2011, 136, 378–384. [Google Scholar] [CrossRef]
- Heck, C.I.; de Mejia, E.G. Yerba Mate Tea (Ilex paraguariensis): A comprehensive review on chemistry, health implications, and technological considerations. J. Food Sci. 2007, 72, 138–151. [Google Scholar] [CrossRef]
- Rustan, A.C.; Drevon, C.A. Fatty Acids: Structures and Properties. In Encyclopedia of Life Sciences; Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2005. [Google Scholar]
- Avato, P.; Tava, A. Rare fatty acids and lipids in plant oilseeds: Occurrence and bioactivity. Phytochem. Rev. 2022, 21, 401–428. [Google Scholar] [CrossRef]
- Yao, X.; Song, Y.; Yang, J.B.; Tan, Y.H.; Corlett, R.T. Phylogeny and biogeography of the hollies (Ilex L., Aquifoliaceae). J. Syst. Evol. 2021, 59, 73–82. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; Liu, H.; Peng, D.; Zhang, J.; Chen, M. Linum usitatissimum FAD2A and FAD3A enhance seed polyunsaturated fatty acid accumulation and seedling cold tolerance in Arabidopsis thaliana. Plant Sci. 2021, 311, 111014. [Google Scholar] [CrossRef] [PubMed]
- Saffaryazdi, A.; Ganjeali, A.; Farhoosh, R.; Cheniany, M. Variation in phenolic compounds, α-linolenic acid and linoleic acid contents and antioxidant activity of purslane (Portulaca oleracea L.) during phenological growth stages. Physiol. Mol. Biol. Plants 2020, 26, 1519–1529. [Google Scholar] [CrossRef]
- Zhukov, A.V. Palmitic acid and its role in the structure and functions of plant cell membranes. Russ. J. Plant Physiol. 2015, 62, 706–713. [Google Scholar] [CrossRef]
- Kunst, L.; Browse, J.; Somerville, C. Enhanced thermal tolerance in a mutant of Arabidopsis deficient in palmitic acid unsaturation. Plant Physiol. 1989, 91, 401–408. [Google Scholar] [CrossRef]
- Ma, K.; Kou, J.; Rahman, M.K.U.; Du, W.; Liang, X.; Wu, F.; Li, W.; Pan, K. Palmitic acid mediated change of rhizosphere and alleviation of Fusarium wilt disease in watermelon. Saudi J. Biol. Sci. 2021, 28, 3616–3623. [Google Scholar] [CrossRef]
- Kurima, K.; Jimbo, H.; Fujihara, T.; Saito, M.; Ishikawa, T.; Wada, H. High myristic acid in glycerolipids enhances the repair of photodamaged photosystem II under strong light. Plant Cell Physiol. 2024, 65, 790–797. [Google Scholar] [CrossRef]
- Li, S.; Xu, C.; Wang, J.; Guo, B.; Yang, L.; Chen, J.; Ding, W. Cinnamic, myristic and fumaric acids in tobacco root exudates induce the infection of plants by Ralstonia solanacearum. Plant Soil 2017, 412, 381–395. [Google Scholar] [CrossRef]
- Jimbo, H.; Takagi, K.; Hirashima, T.; Nishiyama, Y.; Wada, H. Long-chain saturated fatty acids, palmitic and stearic acids, enhance the repair of photosystem II. Int. J. Mol. Sci. 2020, 21, 7509. [Google Scholar] [CrossRef]
- Ullah, S.; Khan, M.N.; Lodhi, S.S.; Ahmed, I.; Tayyab, M.; Mehmood, T.; Din, I.U.; Khan, M.; Sohail, Q.; Akram, M. Targeted metabolomics reveals fatty acid abundance adjustments as playing a crucial role in drought-stress response and post-drought recovery in wheat. Front. Genet. 2022, 13, 972696. [Google Scholar] [CrossRef] [PubMed]
- Lakhssassi, N.; Colantonio, V.; Flowers, N.D.; Zhou, Z.; Henry, J.; Liu, S.; Meksem, K. Stearoyl-acyl carrier protein desaturase mutations uncover an impact of stearic acid in leaf and nodule structure. Plant Physiol. 2017, 174, 1531–1543. [Google Scholar] [CrossRef]
- Bach, L.; Faure, J.D. Role of very-long-chain fatty acids in plant development, when chain length does matter. C R. Biol. 2010, 333, 361–370. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.; Greenhalgh, J.F.D.; Morgan, C.; Edwards, R.; Sinclair, L.; Wilkinson, R. Animal Nutrition, 8th ed.; Pearson Education: Harlow, UK, 2022. [Google Scholar]
- Grela, E.R. Optymalizacja żywienia świń z wykorzystaniem nowej generacji dodatków paszowych. Pr. Mat. Zootech. 2004, 15, 53–63. [Google Scholar]
- Available online: https://www.consilium.europa.eu/en/policies/from-farm-to-fork/ (accessed on 9 June 2024).
- Available online: https://commission.europa.eu/strategy-and-policy/priorities-2019-2024/european-green-deal_en (accessed on 9 June 2024).
- Prost, E.K. Zwierzęta Rzeźne i Mięso-Ocena i Higiena; LTN: Lublin, Poland, 2006. [Google Scholar]
- Sears, A.; Gonzalez, O.; Alberto, A.; Young, A.; de Souza, J.; Relling, A.; Batistel, F. Effect of feeding a palmitic acid-enriched supplement on production responses and nitrogen metabolism of mid-lactating Holstein and Jersey cows. J. Dairy Sci. 2020, 103, 8898–8909. [Google Scholar] [CrossRef] [PubMed]
- Odongo, N.E.; Or-Rashid, M.M.; Kebreab, E.; France, J.; McBride, B.W. Effect of supplementing myristic acid in dairy cow rations on ruminal methanogenesis and fatty acid profile in milk. J. Dairy Sci. 2007, 90, 1851–1858. [Google Scholar] [CrossRef]
- Zwyrzykowska-Wodzińska, A.; Bielas, W.; Niżański, W.; Jankowska-Mąkosa, A.; Knecht, D. Dietary Supplementation with Linseed Oil Ethyl Esters Improves Sexual Behavior and Chosen Seminal Parameters in Porcine Species. Animals 2023, 13, 1347. [Google Scholar] [CrossRef]
- Sierżant, K.; Korzeniowska, M.; Półbrat, T.; Rybarczyk, A.; Smoliński, J. The use of an optimised concentration of quercetin limits peroxidation of lipids in the meat of broiler chickens fed a diet containing flaxseed oil rich in omega-3. Animal 2022, 16, 100603. [Google Scholar] [CrossRef]
- Po, E.; Horsburgh, K.; Raadsma, H.W.; Celi, P. Yerba mate (Ilex paraguarensis) as a novel feed supplement or growing lambs. Small Rumint Res. 2012, 106, 131–136. [Google Scholar] [CrossRef]
- Po, E.; Xu, Z.; Celi, P. The effect of yerba mate (Ilex paraguarensis) supplementation on the productive performance of Dorper ewes and their progeny. Asian-Australas. J. Anim. Sci. 2012, 25, 945–949. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [PubMed]
- Celi, P.; Robinson, A. Effects of Yerba Mate (Ilex paraguariensis) supplementation on the performance of dairy calves. Anim. Prod. Sci. 2010, 50, 376–381. [Google Scholar] [CrossRef]
- Celi, P. Yerba mate (Ilex paraguariensis) as strategic supplement for dairy cows. In Enhancing Animal Welfare and Farmer Income through Strategic Animal Feeding—Some Case Studies; Makkar, H.P.S., Ed.; FAO Animal Production and Health: Rome, Italy, 2013. [Google Scholar]
- Hartemink, E.; Giorgio, D.; Kaur, R.; Di Trana, A.; Celi, P. The Effect of Yerba Mate (Ilex Paraguariensis) Supplementation on Nutrient Degradability in Dairy Cows: An In sacco and In vitro Study. Asian-Australas. J. Anim. Sci. 2015, 28, 1606–1613. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAS | KI (Found) | KI (NIST) | Rt | FAME | PCA G1 | PCA G2 | PCA G3 | PCA G4 | Outside Main PCA Groups | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Iaq.F1 | I×m.M | I×m.F1 | I×m.F3 | I×m.F4 | Iaq.F2 | Iaq.F4 | Iaq.F5 | I×m.F2 | Iaq.M | Ip.M | Iaq.F3 | Ial.F | Iaq.F6 | |||||
| 124-10-7 | 2024 | 2037 | 9.27 | 14:00 | 0.74 | 0.74 | 2.48 | 2.31 | 1.94 | 3.22 | 3.27 | 1.59 | 2.4 | 3.34 | 3.99 | 2.8 | 3.14 | 3.06 |
| 14101-90-7 | 2109 | 2237 1 | 11.03 | 15:01 | 0 | 0 | 0 | 0.19 | 0 | 1.13 | 0.39 | 0.26 | 0.2 | 0 | 0 | 0 | 0.34 | 0 |
| 7132-64-1 | 2125 | 2124 | 11.37 | 15:00 | 0 | 0 | 0.7 | 0.28 | 0.41 | 0 | 0.59 | 0.31 | 0.61 | 0 | 0.63 | 0.08 | 0.84 | 0.82 |
| 1120-25-8 | 2201 | 2237 1 | 13.01 | 16:1Δ 9 | 0 | 0 | 0 | 0.3 | 0 | 0 | 0 | 0.35 | 0.29 | 0 | 0 | 0 | 0 | 0 |
| 56875-67-3 | 2205 | n.r. | 13.1 | 16:1Δ 7 | 0 | 0 | 0 | 0.26 | 0 | 0 | 0 | 0.17 | 0.18 | 0 | 0 | 0 | 0 | 0 |
| 112-39-0 | 2226 | 2225 | 13.54 | 16:00 | 21.94 | 21.53 | 31.08 | 32.71 | 28.92 | 25.04 | 31.64 | 26.25 | 50.64 | 59.47 | 55.18 | 92.48 | 41.23 | 62.78 |
| 112-63-0 | 2395 | 2472 1 | 17.24 | 18:2 Δ 9,12 | 15.68 | 14.93 | 11.8 | 18.14 | 17.08 | 17.86 | 16.89 | 20.01 | 8.35 | 3.42 | 4.58 | 0.89 | 11 | 4.39 |
| 301-00-8 | 2402 | 2583 2 | 17.39 | 18:3 Δ 9,12,15 | 57.11 | 58.93 | 41.06 | 36.82 | 39.9 | 38.39 | 32.31 | 37.9 | 16.55 | 5.61 | 11.24 | 1.41 | 28.44 | 12.67 |
| 112-62-9 | 2407 | 2433 1 | 17.5 | 18:1 Δ 9 | 0 | 0 | 0.9 | 0 | 0 | 0 | 0.57 | 0.18 | 0.5 | 0.27 | 0 | 0 | 0 | 0 |
| 112-61-8 | 2427 | 2434 3 | 17.93 | 18:00 | 1.86 | 1.38 | 4.76 | 3.28 | 3.59 | 4.15 | 4.64 | 4.7 | 5.96 | 6.89 | 6.57 | 1.46 | 4.77 | 5.35 |
| 18287-20-2 | 2489 | n.r. | 19.26 | 18:2 Δ 6,9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.33 | 0 | 1.12 | 0 | 0 | 0 |
| 1731-94-8 | 2528 | 2534 4 | 20.07 | 19:00 | 0.45 | 0.43 | 0.58 | 0.17 | 0.19 | 1.31 | 0.99 | 0.86 | 1.66 | 4.34 | 0.74 | 0.06 | 2.65 | 2.26 |
| 67810-35-9 | 2602 | n.r. | 21.62 | 20:01 | 0 | 0 | 0 | 0.41 | 0 | 0 | 0 | 0.00 | 0 | 0 | 0.32 | 0 | 0 | 0 |
| 1120-28-1 | 2629 | n.r. | 22.15 | 20:00 | 0.47 | 0 | 1.22 | 0.72 | 0.83 | 1.33 | 1.42 | 1.37 | 2.18 | 2.34 | 2.67 | 0.42 | 1.97 | 1.91 |
| 2566-89-4 | 2666 | n.r. | 22.9 | 18:04 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.3 | 0 | 0 | 0 | 0 | 0 |
| 6064-90-0 | 2729 | n.r. | 24.17 | 21:00 | 0 | 0 | 0.37 | 0.37 | 0.42 | 0.53 | 0.43 | 0.39 | 0.57 | 0.82 | 0.76 | 0 | 0.56 | 0 |
| 1120-34-9 | 2805 | n.r. | 25.65 | 22:01 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.13 | 0 | 0 | 0 | 0 | 0 | 0 |
| 929-77-1 | 2830 | n.r. | 26.13 | 22:00 | 0.75 | 0.72 | 1.37 | 1.3 | 1.4 | 1.49 | 1.94 | 1.37 | 2.06 | 2.73 | 1.93 | 0.26 | 1.59 | 1.28 |
| 1120-34-9 | 2837 | 2878 5 | 26.26 | 22:01 | 0 | 0 | 0.22 | 0 | 0 | 0 | 0 | 0 | 0 | 0.21 | 0 | 0 | 0 | 0 |
| 2433-97-8 | 2931 | 2951 5 | 28.02 | 23:00 | 0.25 | 0.42 | 0.73 | 0.67 | 0.64 | 0.96 | 1.03 | 0.78 | 0.58 | 1.02 | 0.84 | 0 | 0.73 | 0.54 |
| 2733-88-2 | 3007 | 3079 5 | 29.42 | 24:01:00 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2442-49-1 | 3032 | 3053 5 | 29.85 | 24:00:00 | 0.38 | 0.36 | 1 | 0.83 | 0.75 | 1.43 | 1.32 | 0.95 | 1.16 | 1.99 | 1.60 | 0.08 | 1.22 | 0.99 |
| 55373-89-2 | 3133 | n.r. | 31.62 | 25:00:00 | 0 | 0 | 0.27 | 0.19 | 0.16 | 0.49 | 0.39 | 0.27 | 0 | 0.22 | 0.56 | 0 | 0.2 | 0 |
| 5802-82-4 | 3228 | n.r. | 33.24 | 26:00:00 | 0.28 | 0.18 | 0.92 | 0.52 | 0.77 | 1.68 | 0.95 | 0.97 | 2.2 | 2.74 | 0.23 | 0.05 | 0.95 | 0 |
| 55682-92-3 | 3435 | n.r. | 36.59 | 28:00:00 | 0.11 | 0.37 | 0.55 | 0.53 | 0.73 | 0.99 | 0.46 | 0.86 | 1.7 | 1.72 | 1.71 | 0 | 0.36 | 0 |
| 4082-55-7 | 3535 | n.r. | 38.15 | 29:00:00 | 0 | 0 | 0 | 0 | 0.12 | 0 | 0 | 0 | 0 | 0.3 | 0.79 | 0 | 0 | 0 |
| 629-83-4 | 3635 | n.r. | 39.76 | 30:00:00 | 0 | 0 | 0 | 0 | 2.14 | 0 | 0.78 | 0 | 1.57 | 2.58 | 4.54 | 0 | 0.24 | 3.94 |
| Sum of saturated FA | 27.12 | 25.76 | 45.48 | 71.59 | 42.28 | 41.63 | 49.39 | 39.81 | 71.59 | 88.78 | 81.03 | 97.69 | 60.09 | 82.93 | ||||
| Sum of unsaturated FA | 72.79 | 73.86 | 53.98 | 26.7 | 56.98 | 57.38 | 50.16 | 59.1 | 26.7 | 9.51 | 17.26 | 2.3 | 39.78 | 17.06 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zwyrzykowska-Wodzińska, A.; Jarosz, B.; Okińczyc, P.; Szperlik, J.; Bąbelewski, P.; Zadák, Z.; Jankowska-Mąkosa, A.; Knecht, D. GC-MS and PCA Analysis of Fatty Acid Profile in Various Ilex Species. Molecules 2024, 29, 4833. https://doi.org/10.3390/molecules29204833
Zwyrzykowska-Wodzińska A, Jarosz B, Okińczyc P, Szperlik J, Bąbelewski P, Zadák Z, Jankowska-Mąkosa A, Knecht D. GC-MS and PCA Analysis of Fatty Acid Profile in Various Ilex Species. Molecules. 2024; 29(20):4833. https://doi.org/10.3390/molecules29204833
Chicago/Turabian StyleZwyrzykowska-Wodzińska, Anna, Bogdan Jarosz, Piotr Okińczyc, Jakub Szperlik, Przemysław Bąbelewski, Zdeněk Zadák, Anna Jankowska-Mąkosa, and Damian Knecht. 2024. "GC-MS and PCA Analysis of Fatty Acid Profile in Various Ilex Species" Molecules 29, no. 20: 4833. https://doi.org/10.3390/molecules29204833
APA StyleZwyrzykowska-Wodzińska, A., Jarosz, B., Okińczyc, P., Szperlik, J., Bąbelewski, P., Zadák, Z., Jankowska-Mąkosa, A., & Knecht, D. (2024). GC-MS and PCA Analysis of Fatty Acid Profile in Various Ilex Species. Molecules, 29(20), 4833. https://doi.org/10.3390/molecules29204833