
Variation in the Phenolic Profile and Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity in Leaves of Cotoneaster zabelii during Growing Season


Abstract

1. Introduction
2. Results
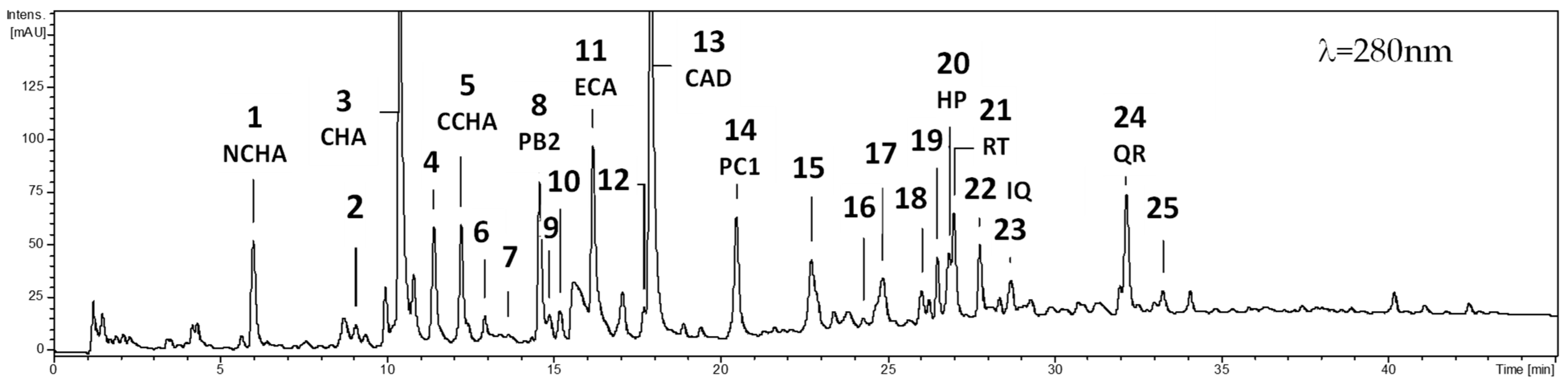
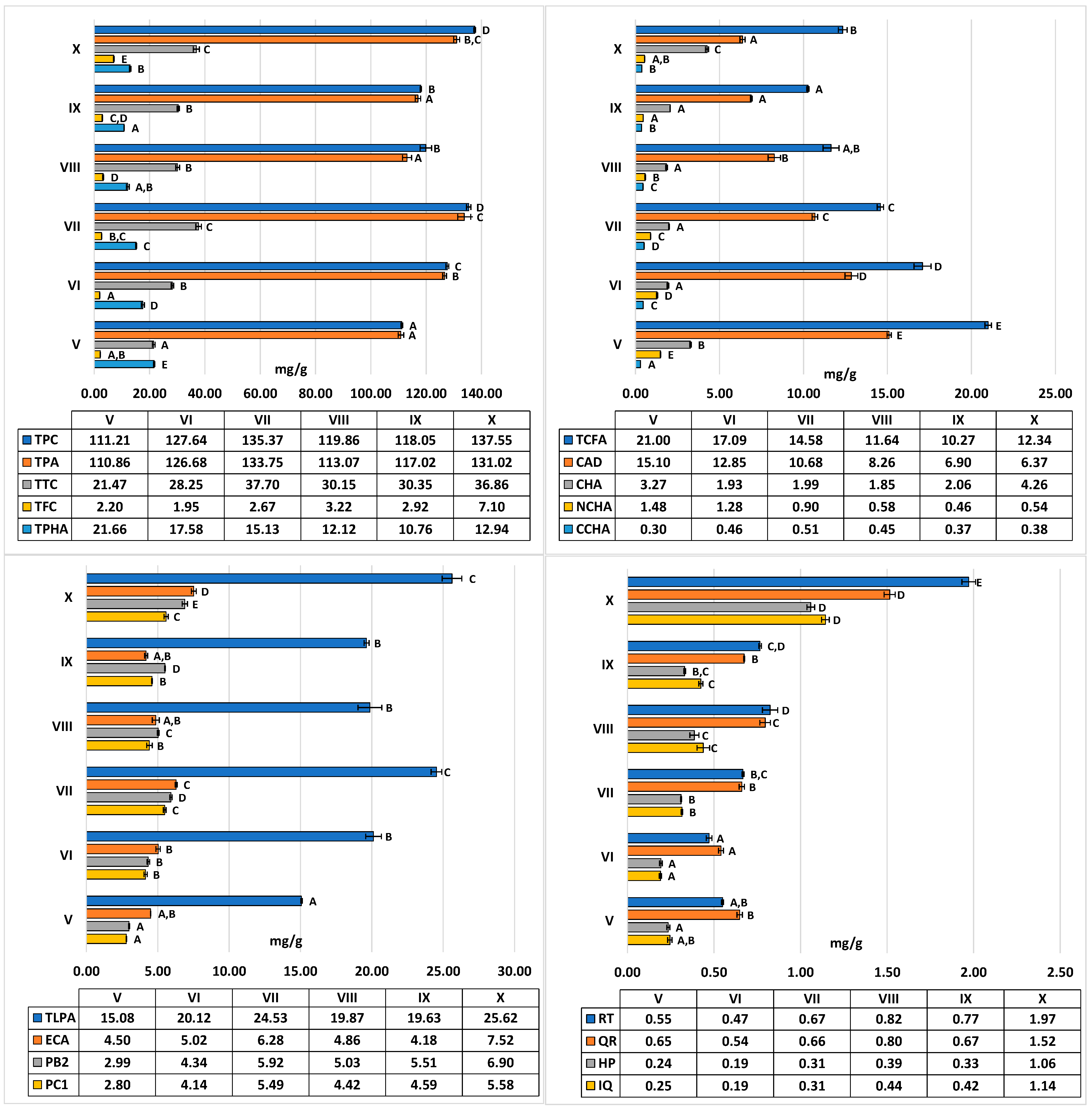
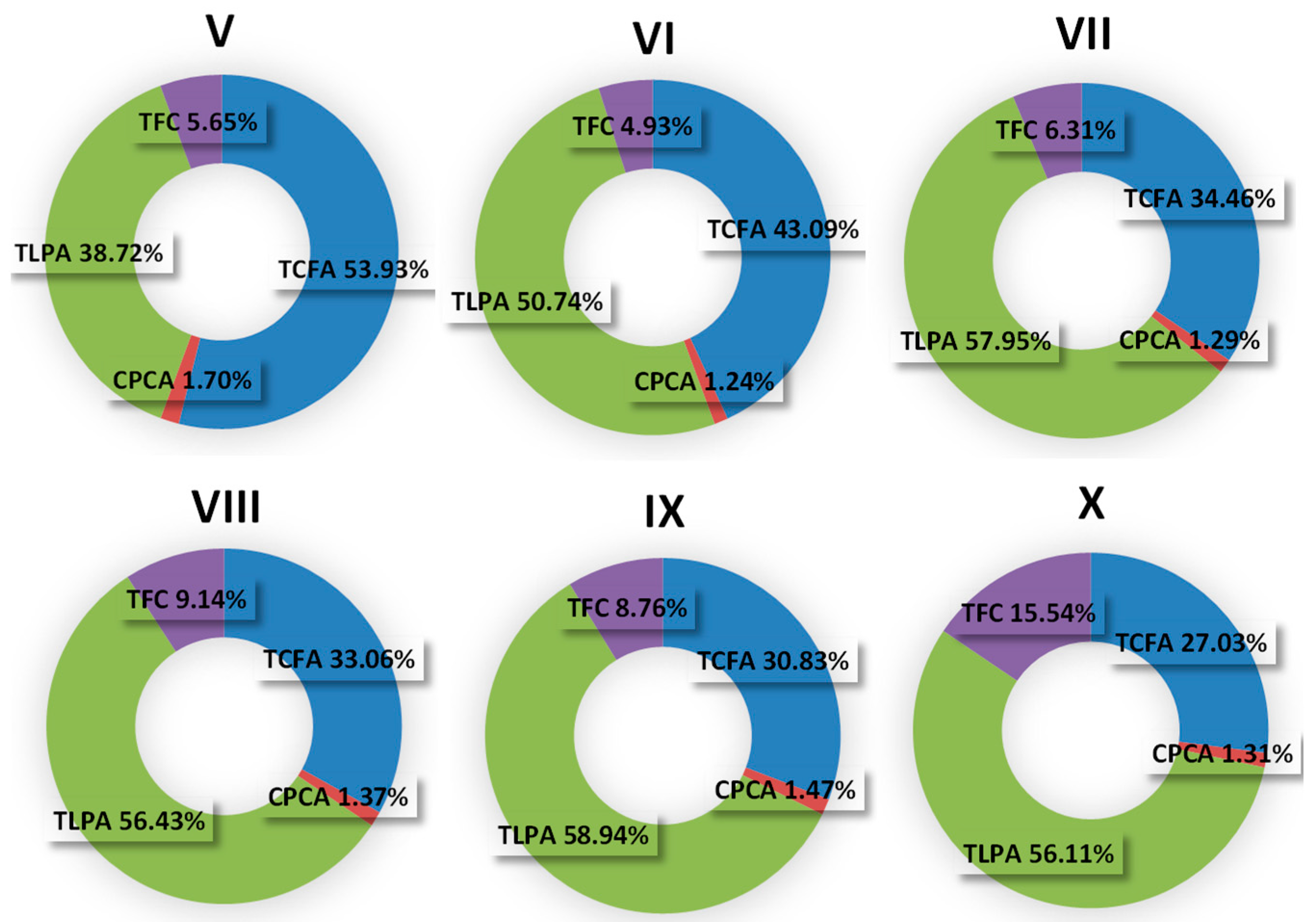
2.1. Seasonal Variability in the Levels of Polyphenolic Components
2.2. Seasonal Variability in the Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Plant Material and Sample Preparation
4.3. Phytochemical Profiling
4.4. Antioxidant Activity
4.5. Inhibition of Hyaluronidase and α-Glucosidase
4.6. Statistical and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ferreira, I.C.F.R.; Martins, N.; Barros, L. Phenolic compounds and its bioavailability: In vitro bioactive compounds or health promoters? Adv. Food Nutr. Res. 2017, 82, 1–44. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros Gomes, J.; Cahino Terto, M.V.; Golzio do Santos, S.; Sobral da Silva, M.; Fechine Tavares, J. Seasonal variations of polyphenols content, sun protection factor and antioxidant activity of two Lamiaceae species. Pharmaceutics 2021, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Krzemińska, B.; Dybowski, M.P.; Klimek, K.; Typek, R.; Miazga-Karska, M.; Ginalska, G.; Dos Santos Szewczyk, K. Can extracts from the leaves and fruits of the Cotoneaster species be considered promising anti-acne agents? Molecules 2022, 27, 2907. [Google Scholar] [CrossRef] [PubMed]
- Krzemińska, B.; Dybowski, M.P.; Klimek, K.; Typek, R.; Miazga-Karska, M.; Dos Santos Szewczyk, K. The anti-acne potential and chemical composition of two cultivated Cotoneaster species. Cells 2022, 11, 367. [Google Scholar] [CrossRef]
- Kicel, A.; Michel, P.; Owczarek, A.; Marchelak, A.; Żyżelewicz, D.; Budryn, G.; Oracz, J.; Olszewska, M.A. Phenolic profile and antioxidant potential of leaves from selected Cotoneaster Medik. species. Molecules 2016, 21, 688. [Google Scholar] [CrossRef]
- Kicel, A.; Kolodziejczyk-Czepas, J.; Owczarek, A.; Rutkowska, M.; Wajs-Bonikowska, A.; Granica, S.; Nowak, P.; Olszewska, M.A. Multifunctional phytocompounds in Cotoneaster fruits: Phytochemical profiling, cellular safety, anti-inflammatory and antioxidant effects in chemical and human plasma models in vitro. Oxid. Med. Cell. Longev. 2018, 2018, 3482521. [Google Scholar] [CrossRef]
- Kicel, A. An Overview of the genus Cotoneaster (Rosaceae): Phytochemistry, biological activity, and toxicology. Antioxidants 2020, 9, 1002. [Google Scholar] [CrossRef]
- Kicel, A.; Kolodziejczyk-Czepas, J.; Owczarek, A.; Marchelak, A.; Sopinska, M.; Ciszewski, P.; Nowak, P.; Olszewska, M.A. Polyphenol-rich extracts from Cotoneaster leaves inhibit pro-inflammatory enzymes and protect human plasma components against oxidative stress In Vitro. Molecules 2018, 23, 2472. [Google Scholar] [CrossRef]
- Kicel, A.; Owczarek, A.; Kapusta, P.; Kolodziejczyk-Czepas, J.; Olszewska, M.A. Contribution of individual polyphenols to antioxidant activity of Cotoneaster bullatus and Cotoneaster zabelii leaves—structural relationships, synergy effects and application for quality control. Antioxidants 2020, 9, 69. [Google Scholar] [CrossRef]
- Kicel, A.; Magiera, A.; Skrzywanek, M.; Malczuk, M.; Olszewska, M.A. The Inhibition of α-glucosidase, α-amylase and protein glycation by phenolic extracts of Cotoneaster bullatus, Cotoneaster zabelii, and Cotoneaster integerrimus leaves and fruits: Focus on anti-hyperglycemic activity and kinetic parameters. Molecules 2022, 27, 7081. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, J.; Mitić, S.; Mitić, M.; Kocić, G.; Pavlović, A.; Tošić, S. Variation in the phenolic compounds profile and antioxidant activity in different parts of hawthorn (Crataegus pentagyna Willd.) during harvest periods. Pol. J. Food Nutr. Sci. 2019, 69, 367–378. [Google Scholar] [CrossRef]
- Zheng, W.; Zhou, M.; Chai, R.; Liang, R.; Zhang, J.; Zhao, Y.; Zheng, X.; Jin, Y.; Guo, B.; Ma, B. Quality analysis of hawthorn leaves (the leaves of Crataegus pinnatifida Bge. var major N.E.Br) in different harvest time. Phytochem. Anal. 2022, 33, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Yang, X.; Hu, J.Y.; Jiao, J.; Mu, F.S.; Song, Z.Y.; Gai, Q.Y.; Qiao, Q.; Ruan, X.; Fu, Y.J. Antioxidant properties of phenolic compounds in renewable parts of Crataegus pinnatifida inferred from seasonal variations. J. Food Sci. 2016, 81, C1102–C1109. [Google Scholar] [CrossRef]
- Gębalski, J.; Graczyk, F.; Załuski, D. Paving the way towards effective plant-based inhibitors of hyaluronidase and tyrosinase: A critical review on a structure–activity relationship. J. Enzyme Inhib. Med. Chem. 2022, 37, 1120–1195. [Google Scholar] [CrossRef]
- Koga, Y.; Setoguchi, Y.; Sugamoto, K.; Goto, Y.; Hirano, T.; Kunitake, H. Seasonal variation and mean degree of polymerization of proanthocyanidin in leaves and branches of rabbiteye blueberry (Vaccinium virgatum Aiton). Plants 2024, 13, 1864. [Google Scholar] [CrossRef]
- Mandim, F.; Petropoulos, S.A.; Giannoulis, K.D.; Dias, M.I.; Fernandes, Â.; Pinela, J.; Kostic, M.; Soković, M.; Barros, L.; Santos-Buelga, C.; et al. Seasonal variation of bioactive properties and phenolic composition of Cynara cardunculus var. altilis. Food Res. Int. 2020, 134, 109281. [Google Scholar] [CrossRef]
- Wang, T.K.; Xu, S.; Li, S.; Zhang, Y. Proanthocyanidins should be a candidate in the treatment of cancer, cardiovascular diseases and lipid metabolic disorder. Molecules 2020, 25, 5971. [Google Scholar] [CrossRef]
- Chen, H.; Wang, W.; Yu, S.; Wang, H.; Tian, Z.; Zhu, S. Procyanidins and their therapeutic potential against oral diseases. Molecules 2022, 27, 2932. [Google Scholar] [CrossRef]
- Yu, D.; Huang, T.; Tian, B.; Zhan, J. Advances in biosynthesis and biological functions of proanthocyanidins in horticultural plants. Foods 2020, 9, 1774. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, L.; Han, Z.; Jiang, Y.; Zhao, L.; Liu, H.; Yang, L.; Luo, K. Molecular cloning and characterization of PtrLAR3, a gene encoding leucoanthocyanidin reductase from Populus trichocarpa, and its constitutive expression enhances fungal resistance in transgenic plants. J. Exp. Bot. 2012, 63, 2513–2524. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Unsicker, S.B.; Fellenberg, C.; Constabel, C.P.; Schmidt, A.; Gershenzon, J.; Hammerbacher, A. Flavan-3-ols are an effective chemical defense against rust infection. Plant Physiol. 2017, 175, 1560–1578. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, X.; Jiang, L.; Chu, Y.; Gao, S.; Jiang, X.; Zhang, Y.; Chen, Y.; Luo, S.; Peng, C. The biological activity mechanism of chlorogenic acid and its applications in food industry: A Review. Front. Nutr. 2022, 9, 943911. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; Taine, E.G.; Meng, D.; Cui, T.; Tan, W. Chlorogenic Acid: A systematic review on the biological functions, mechanistic actions, and therapeutic potentials. Nutrients 2024, 16, 924. [Google Scholar] [CrossRef]
- Mei, Y.; Sun, H.; Du, G.; Wang, X.; Lyu, D. Exogenous chlorogenic acid alleviates oxidative stress in apple leaves by enhancing antioxidant capacity. Sci. Hortic. 2020, 274, 109676. [Google Scholar] [CrossRef]
- Moreira, E.A.; Pilon, A.C.; Andrade, L.E.; Lopes, N.P. New perspectives on chlorogenic acid accumulation in harvested leaf tissue: Impact on traditional medicine preparations. ACS Omega 2018, 3, 18380–18386. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a bioactive compound from medicinal plants and its therapeutic applications. Biomed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Khalid, M.; Saeed-ur-Rahman; Bilal, M.; Huang, D. Role of flavonoids in plant interactions with the environment and against human pathogens—A review. J. Integr. Agric. 2019, 18, 211–230. [Google Scholar] [CrossRef]
- Mattila, H.; Valev, D.; Havurinne, V.; Khorobrykh, S.; Virtanen, O.; Antinluoma, M.; Mishra, K.B.; Tyystjärvi, E. Degradation of chlorophyll and synthesis of flavonols during autumn senescence—The story told by individual leaves. AoB Plants 2018, 10, ply028. [Google Scholar] [CrossRef] [PubMed]
- European Pharmacopoeia Commission. Tannins in Herbal Drugs. In European Pharmacopoeia, 11th ed.; Council of Europe: Strasbourg, France, 2022; p. 326. [Google Scholar]
- Granica, S.; Czerwińska, M.E.; Piwowarski, J.P.; Ziaja, M.; Kiss, A.K. Chemical composition, antioxidative and anti-inflammatory activity of extracts prepared from aerial parts of Oenothera biennis L. and Oenothera paradoxa Hudziok obtained after seeds cultivation. J. Agric. Food Chem. 2013, 61, 801–810. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Group | CV (%) | Compound Group | CV (%) |
|---|---|---|---|
| TPC (mg/g) | 8.29 | TFC (mg/g) HPLC | 56.76 |
| TPA (mg/g) | 7.94 | TCFA (mg/g) HPLC | 27.56 |
| TTC (mg/g) | 19.40 | CPCA (mg/g) HPLC | 13.41 |
| TLPA (mg/g) HPLC | 18.31 | TPHA (mg/g) HPLC | 26.87 |
| Sample/Variability | DPPH Scavenging | FRAP | O2•− Scavenging | α-Glucosidase Inhibition | Hyaluronidase Inhibition |
|---|---|---|---|---|---|
| EC50 (µg/mL) a | mmol Fe2+/g b | EC50 (µg/mL) a | IC50 (µg/mL) c | IC50 µg/mL c | |
| TX | 4.70 ± 0.09 A | 13.54 ± 0.26 D | 136.97 ± 1.84 E | - | - |
| AA | 4.13 ± 0.02 A | 26.25 ± 0.10 E | 3.24 ± 0.11 A | - | - |
| AR | - | - | - | 201.69 ± 2.76 D | - |
| HP | - | - | - | - | 58.93 ± 1.63 A |
| V | 21.59 ± 0.44 D | 2.78 ± 0.03 A,B | 39.77 ± 1.25 D | 44.19 ± 1.39 C | 135.87 ± 1.00 D |
| VI | 19.04 ± 0.06 B,C | 2.97 ± 0.02 B,C | 21.23 ± 0.17 B | 36.39 ± 1.46 A,B | 120.87 ± 0.48 C |
| VII | 17.97 ± 0.33 B | 3.09 ± 0.02 C | 19.78 ± 0.48 B | 32.62 ± 0.79 A | 99.82 ± 0.39 B |
| VIII | 19.06 ± 0.59 B,C | 2.73 ± 0.08 A | 28.72 ± 1.91 C | 44.36 ± 1.07 C | 119.88 ± 2.22 C |
| IX | 20.54 ± 0.68 C,D | 2.73 ± 0.04 A | 32.54 ± 0.21 C | 41.10 ± 0.17 B,C | 99.88 ± 1.32 B |
| X | 17.27 ± 0.14 B | 3.05 ± 0.03 C | 30.96 ± 0.51 C | 39.29 ± 0.39 B | 100.29 ± 1.52 B |
| CV (%) d | 8.31 | 5.69 | 25.84 | 11.56 | 13.39 |
| r (p) for: | DPPH-Scavenging EC50 (µg/mL) | FRAP mmol Fe2+/g | O2•−-Scavenging EC50 (µg/mL) | α-Glucosidase Inhibition IC50 (µg/mL) | Hyaluronidase Inhibition IC50 (µg/mL) |
|---|---|---|---|---|---|
| TPC (mg/g) | −0.9530 (0.003) * | 0.9088 (0.012) * | −0.6819 (0.136) | −0.7701 (0.073) | −0.6840 (0.134) |
| TPA (mg/g) | −0.8346 (0.039) * | 0.9553 (0.003) * | −0.7256 (0.103) | −0.9044 (0.013) * | −0.6609 (0.153) |
| TTC (mg/g) | −0.89016 (0.017) * | 0.6813 (0.136) | −0.6011 (0.207) | −0.6458 (0.166) | −0.8869(0.019) * |
| TLPA (mg/g) | −0.9488 (0.004) * | 0.7749 (0.070) | −0.5998 (0.208) | −0.6618 (0.152) | −0.8265 (0.043) * |
| TFC (mg/g) | −0.6250 (0.185) | 0.3640 (0.478) | 0.1502 (0.776) | 0.0354 (0.947) | −0.4976 (0.315) |
| TCFA (mg/g) | 0.4364 (0.387) | 0.0905 (0.865) | 0.2065 (0.695) | 0.0183 (0.973) | 0.7674 (0.045) |
| CPCA (mg/g) | 0.0421 (0.697) | 0.1664 (0.753) | 0.6048 (0.203) | 0.1982 (0.707) | 0.3339 (0.518) |
| TPHA (mg/g) | 0.4352 (0.389) | 0.0925 (0.862) | 0.2151 (0.682) | 0.0217 (0.967) | 0.7647 (0.047) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kicel, A.; Magiera, A.; Olszewska, M.A. Variation in the Phenolic Profile and Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity in Leaves of Cotoneaster zabelii during Growing Season. Molecules 2024, 29, 4745. https://doi.org/10.3390/molecules29194745
Kicel A, Magiera A, Olszewska MA. Variation in the Phenolic Profile and Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity in Leaves of Cotoneaster zabelii during Growing Season. Molecules. 2024; 29(19):4745. https://doi.org/10.3390/molecules29194745
Chicago/Turabian StyleKicel, Agnieszka, Anna Magiera, and Monika Anna Olszewska. 2024. "Variation in the Phenolic Profile and Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity in Leaves of Cotoneaster zabelii during Growing Season" Molecules 29, no. 19: 4745. https://doi.org/10.3390/molecules29194745
APA StyleKicel, A., Magiera, A., & Olszewska, M. A. (2024). Variation in the Phenolic Profile and Antioxidant, Antihyperglycemic, and Anti-Inflammatory Activity in Leaves of Cotoneaster zabelii during Growing Season. Molecules, 29(19), 4745. https://doi.org/10.3390/molecules29194745