

Composition and Activities of Carpesium macrocephalum Franch. & Sav. Essential Oils

 ,
,  ,
,  and
and
Abstract

1. Introduction
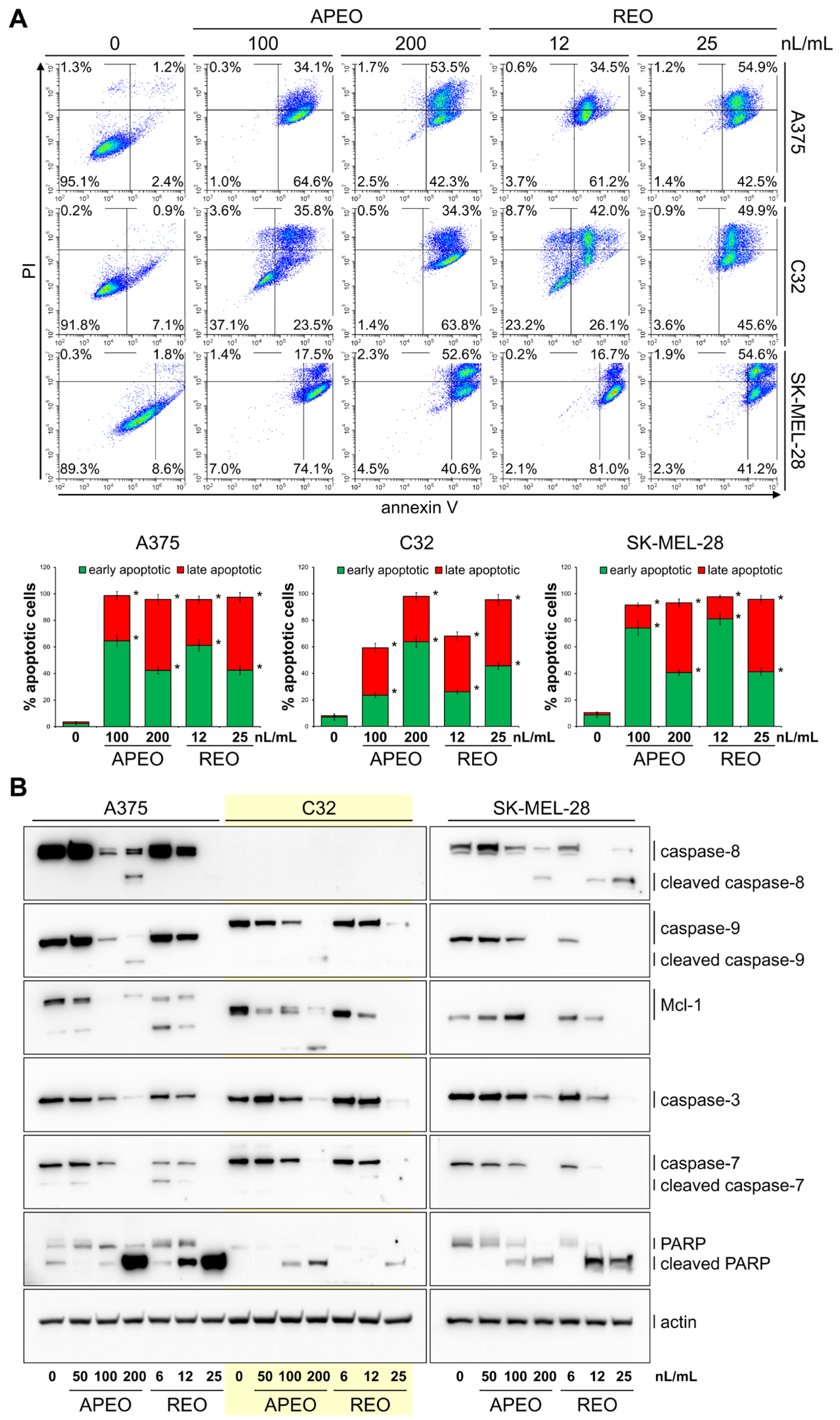
2. Results
2.1. Chemical Composition of C. macrocephalum Essential Oils
2.2. Antibacterial and Cytotoxic Activities of C. macrocephalum Essential Oils
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Isolation of Essential Oil
4.4. Identification of Essential Oil Constituents
4.5. Antibacterial Activity of Carpesium macrocephalum Essential Oils
4.5.1. Bacterial Strains and Culture Conditions
4.5.2. Determination of the Minimum Inhibitory Concentrations (MICs) and the Effectiveness of the Tested C. macrocephalum EOs against the Reference Bacterial Strains
4.6. Cytotoxic Activity of Essential Oils from Carpesium macrocephalum
4.6.1. Cell Lines and Culture Conditions
4.6.2. Cell-Viability Assay
4.6.3. Apoptosis Assay
4.6.4. Western Immunoblot
4.6.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- eFloras. Missouri Botanical Garden; St. Louis, MO & Harvard University Herbaria: Cambridge, MA, USA, 2008; Available online: http://www.efloras.org (accessed on 30 July 2024).
- Zhang, J.-P.; Wang, G.-W.; Tian, X.-H.; Yang, Y.-X.; Liu, Q.-X.; Chen, L.-P.; Li, H.-L.; Zhang, W.-D. The genus Carpesium: A review of its ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharm. 2015, 163, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Koppula, S.; Kim, W.-J.; Jiang, J.; Shim, D.-W.; Oh, N.-H.; Kim, T.-J.; Kang, T.-B.; Lee, K.-H. Carpesium macrocephalum attenuates lipopolysaccharide-induced inflammation in macrophages by regulating the NF-κB/IκB-α, Akt, and STAT signaling pathways. Am. J. Chin. Med. 2013, 41, 927–943. [Google Scholar] [CrossRef] [PubMed]
- Butala, S.; Suvarna, V.; Mallya, R.; Khan, T. An insight into cytotoxic activity of flavonoids and sesquiterpenoids from selected plants of Asteraceae species. Chem. Biol. Drug Des. 2021, 98, 1116–1130. [Google Scholar] [CrossRef]
- Shi, N.-N.; Hou, C.-C.; Liu, Y.; Li, K.-Y.; Mi, S.-D.; Tong, B.-L.; Zhang, M.-L. Chemical constituents of plants from the genus Carpesium. Heterocycl. Comm. 2022, 28, 95–123. [Google Scholar] [CrossRef]
- Konovalova, O.A.; Rybalko, K.S.; Kabanov, V.S. Sesquiterpene lactones from Carpesium eximium. Chem. Nat. Compd. 1972, 8, 705–707. [Google Scholar] [CrossRef]
- Yang, C.; Yuan, C.S.; Han, Y.F.; Jia, Z.-J. A new eudesmanolide from Carpesium macrocephalum. Chin. Chem. Lett. 2002, 13, 855–856. [Google Scholar]
- Yang, C.; Yuan, C.S.; Han, Y.F.; Jia, Z.-J. Sesquiterpene lactone glycosides from Carpesium macrocephalum. Chin. Chem. Lett. 2002, 13, 247–248. [Google Scholar]
- Kim, M.-R.; Kim, C.-S.; Hwang, K.-H.; Park, I.-Y.; Hong, S.-S.; Son, J.-K.; Moon, D.-C. Isolation and structures of guaianolides from Carpesium macrocephalum. J. Nat. Prod. 2002, 65, 583–584. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shi, Y.-P.; Jia, Z.-J. Sesquiterpene lactone glycosides, eudesmanolides, and other constituents from Carpesium macrocephalum. Planta Med. 2002, 68, 626–630. [Google Scholar] [CrossRef]
- Yang, C.; Zhu, Y.; Jia, Z.-J. Sesquiterpene lactones and other constituents from the aerial parts of Carpesium macrocephalum. Aust. J. Chem. 2003, 56, 621–624. [Google Scholar] [CrossRef]
- Kim, M.-R.; Lee, S.-K.; Kim, C.-S.; Kim, K.-S.; Moon, D.-C. Phytochemical constituents of Carpesium macrocephalum Fr. et. Sav. Arch. Pharm. Res. 2004, 27, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-P.; Yang, Y.-X.; Liu, Q.-X.; Chen, L.-P.; Li, H.-L. Monoterpenoids from whole plants of Carpesium macrocephalum. Chin. Tradit. Herb. Drugs 2015, 46, 2985–2988. [Google Scholar] [CrossRef]
- Zhang, J.-P.; Xu, X.-K.; Ye, J.; Yang, Y.-X.; Gao, S.; Li, H.-L.; Zhang, W.-D. Three new sesquiterpene lactone dimers from Carpesium macrocephalum. Fitoterapia 2016, 110, 72–76. [Google Scholar] [CrossRef]
- Xie, C.; Sun, L.; Meng, L.; Wang, M.; Xu, J.; Bartlam, M.; Guo, Y. Sesquiterpenes from Carpesium macrocephalum inhibit Candida albicans biofilm formation and dimorphism. Bioorg. Med. Chem. Lett. 2015, 25, 5409–5411. [Google Scholar] [CrossRef]
- Wang, J.; Wang, N.; Yao, X.; Ishii, R.; Kitanaka, S. Inhibitory activity of Chinese herbal medicines toward histamine release from mast cells and nitric oxide production by macrophage-like cell line, RAW 264.7. J. Nat. Med. 2006, 60, 73–77. [Google Scholar] [CrossRef]
- Lee, H.J.; Lim, H.J.; Lee, D.Y.; Jung, H.; Kim, M.-R.; Moon, D.-C.; Kim, K.I.; Lee, M.-S.; Ryu, J.-H. Carabrol suppresses LPS-induced nitric oxide synthase expression by inactivation of p38 and JNK via inhibition of I-κBα degradation in RAW 264.7 cells. Biochem. Biophys. Res. Commun. 2010, 391, 1400–1404. [Google Scholar] [CrossRef]
- Kalemba, D.; Matla, M.; Smętek, A. Antimicrobial Activities of Essential Oils. In Dietary Phytochemicals and Microbes, 1st ed.; Patra, A.K., Ed.; Science + Business Media: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2012; Chapter 5; pp. 157–183. [Google Scholar] [CrossRef]
- Aponso, M.; Hearn, M.T.W.; Patti, A.F.; Bennett, L.E. Relaxation effects of essential oils are explained by their interactions with human brain neurotransmitter receptors and electroencephalography rhythms. ACS Chem. Neurosci. 2022, 13, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, M.; Wei, Y.; Li, H.; He, X.; Yang, Q.; Li, Z.; Duan, J.; Wu, Z.; Chen, Q.; et al. Inhalation aromatherapy via brain-targeted nasal delivery: Natural volatiles or essential oils on mood disorders. Front. Pharmacol. 2022, 13, 860043. [Google Scholar] [CrossRef]
- Sattayakhom, A.; Wichit, S.; Koomhin, P. The effects of essential oils on the nervous system: A scoping review. Molecules 2023, 28, 3771. [Google Scholar] [CrossRef]
- Ahn, C.; Lee, J.-H.; Park, M.-J.; Kim, J.-W.; Yang, J.; Yoo, Y.-M.; Jeung, E.-B. Cytostatic effects of plant essential oils on human skin and lung cells. Exp. Ther. Med. 2020, 19, 2008–2018. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhu, L.; Wang, S.; Gao, Y.; Jin, F. Molecular mechanism of the anti-inflammatory effects of plant essential oils: A systematic review. J. Ethnopharmacol. 2023, 301, 115829. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Malarz, J.; Stojakowska, A. Composition of essential oils from roots and aerial parts of Carpesium divaricatum, a traditional herbal medicine and wild edible plant from south-east Asia, grown in Poland. Molecules 2019, 24, 4418. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Malarz, J.; Szoka, Ł.; Kwiatkowski, P.; Stojakowska, A. Composition of essential oils from roots and aerial parts of Carpesium cernuum and their antibacterial and cytotoxic activities. Molecules 2021, 26, 1883. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Szoka, Ł.; Kwiatkowski, P.; Meena, S.N.; Stojakowska, A. Bioprospecting of the Telekia speciosa: Uncovering the composition and biological properties of its essential oils. Appl. Sci. 2023, 13, 5674. [Google Scholar] [CrossRef]
- Englund, M.; Pornpongrungrueng, P.; Geustafsson, M.H.G.; Anderberg, A.A. Phylogenetic relationships and generic delimitation in Inuleae subtribe Inulinae (Asteraceae) based on ITS and cpDNA sequence data. Cladistics 2009, 25, 319–352. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Larruscain, D.; Santos-Vicente, M.; Anderberg, A.A.; Rico, E.; Martínez-Ortega, M.M. Phylogeny of the Inula group (Asteraceae: Inuleae): Evidence from nuclear and plastid genomes and a recircumscription of Pentanema. Taxon 2018, 67, 149–164. [Google Scholar] [CrossRef]
- Lunz, K.; Stappen, I. Back to the roots—An overview of the chemical composition and bioactivity of selected root-essential oils. Molecules 2021, 26, 3155. [Google Scholar] [CrossRef] [PubMed]
- Radulović, N.; Blagojević, P.; Palić, R.; Zlatković, B. Volatiles of Telekia speciosa (Schreb.) Baumg. (Asteraceae) from Serbia. J. Essent. Oil Res. 2010, 22, 250–254. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Stojakowska, A.; Kalemba, D. Chemical composition of essential oils from a multiple shoot culture of Telekia speciosa and different plant organs. Nat. Prod. Commun. 2012, 7, 625–628. [Google Scholar] [CrossRef]
- Cilović Kozarević, E.; Šarić-Kundalić, B.; Ibišević, M.; Horozić, E.; Glamočlija, J.; Soković, M.; Arsenijević, J.; Maksimović, Z. GC/MS analysis and antimicrobial activity of essential oils of Telekia speciosa (Schreb.) Baumg. Lek. Sirovine 2021, 41, 35–40. [Google Scholar] [CrossRef]
- Kameoka, H.; Sagara, K.; Miyazawa, M. Components of essential oils of Kakushitsu (Daucus carota L. and Carpesium abrotanoides L.). Nippon. Nōgeikagaku Kaishi 1989, 63, 185–188. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, L.; Lin, L.; Zhang, R.; Du, Y.; Chen, H.; Huang, M.; Guo, K.; Yang, X. Essential oil from Carpesium abrotanoides L. induces apoptosis via activating mitochondrial pathway in hepatocellular carcinoma cells. Curr. Med. Sci. 2018, 38, 1045–1053. [Google Scholar] [CrossRef]
- Haris, A.; Azeem, M.; Binyameen, M. Mosquito repellent potential of Carpesium abrotanoides essential oil and its main components against a dengue vector, Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2022, 59, 801–809. [Google Scholar] [CrossRef]
- Bourrel, C.; Vilarem, G.; Perineau, F. Chemical analysis, bacteriostatic and fungistatic properties of the essential oil of elecampane (Inula helenium L.). J. Essent. Oil Res. 1993, 5, 411–417. [Google Scholar] [CrossRef]
- Boatto, G.; Pintore, G.; Palomba, M.; de Simone, F.; Ramundo, E.; Iodice, C. Composition and antibacterial activity of Inula helenium and Rosmarinus officinalis essential oils. Fitoterapia 1994, 65, 279–280. [Google Scholar]
- Deriu, A.; Zanetti, S.; Sechi, L.A.; Marongiu, B.; Piras, A.; Porcedda, S.; Tuveri, E. Antimicrobial activity of Inula helenium L. essential oil against Gram-positive and Gram-negative bacteria and Candida spp. Int. J. Antimicrob. Agents 2008, 31, 588–590. [Google Scholar] [CrossRef]
- Stojanović-Radić, Z.; Čomić, L.; Radulović, N.; Blagojević, P.; Denić, M.; Miltojević, A.; Rajković, J.; Mihajilov-Krstev, T. Antistaphylococcal activity of Inula helenium L. root essential oil: Eudesmane sesquiterpene lactones induce cell membrane damage. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1015–1025. [Google Scholar] [CrossRef]
- Stojanović-Radić, Z.; Dimitrijević, M.; Genčić, M.; Pejčić, M.; Radulović, N. Anticandidal activity of Inula helenium root essential oil: Synergistic potential, anti-virulence efficacy and mechanism of action. Ind. Crops Prod. 2020, 149, 112373. [Google Scholar] [CrossRef]
- Konishi, T.; Shimada, Y.; Nagao, T.; Okabe, H.; Konoshima, T. Antiproliferative sesquiterpene lactones from the roots of Inula helenium. Biol. Pharm. Bull. 2002, 25, 1370–1372. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J.; Chen, Y.; Zou, D.; Pu, Y.; Wei, M.; Huang, Y.; Li, Y.; Huang, Q.; Chen, J. Alantolactone inhibits melanoma cell culture viability and migration and promotes apoptosis by inhibiting Wnt/β-catenin signaling. Anticancer Agents Med. Chem. 2023, 23, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhao, Q.; Dai, X.; Wen, X.; Luo, X.; Duan, Y.; Yang, Z.; Dai, Q. Alantolactone enhances the sensitivity of melanoma to MAPK pathway inhibitors by targeting inhibition of STAT3 activation and down-regulating stem cell markers. Cancer Cell Int. 2024, 24, 191. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 5th ed.; Texensis Publishing: Gruver, TX, USA, 2017. [Google Scholar]
- Kalsi, P.S.; Goyal, R.; Talwar, K.K.; Chhabra, B.R. Stereostructures of two biologically active sesquiterpene lactones from Inula racemosa. Phytochemistry 1989, 28, 2093–2096. [Google Scholar] [CrossRef]
- Perez, A.L.C.; Nava, L.M.; Romo de Vivar, A. Absinthifolide, a sesquiterpene glycoside from Bahia absinthifolia var. absinthifolia. Phytochemistry 1987, 26, 765–767. [Google Scholar] [CrossRef]
- Klochkov, S.G.; Afanaseva, S.V.; Pushin, A.N. Acidic isomerization of alantolactone derivatives. Chem. Nat. Compd. 2006, 42, 400–406. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, Approved Standard, 9th ed.; CLSI document M07-A9; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; Di Lorenzo, A.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef]
- Kwiatkowski, P.; Łopusiewicz, Ł.; Pruss, A.; Kostek, M.; Sienkiewicz, M.; Bonikowski, R.; Wojciechowska-Koszko, I.; Dołęgowska, B. Antibacterial activity of selected essential oil compounds alone and in combination with β-lactam antibiotics against MRSA strains. Int. J. Mol. Sci. 2020, 21, 7106. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| No | Compound | APEO | REO | RI Exp. 1 | RI Lit. 2 | Identification Method |
|---|---|---|---|---|---|---|
| Amount [%] | ||||||
| 1 | 2-Pentylfuran | tr | 0.1 | 965 | 971 | RI, MS |
| 2 | Octanal | 0.5 | 0.1 | 975 | 982 | RI, MS |
| 3 | β-Myrcene | tr | 976 | 987 | RI, MS | |
| 4 | (E)-2-(2-Pentenyl)furan | 2.0 | 977 | 983 | RI, MS | |
| 5 | Benzeneacetaldehyde | tr | 1007 | 1009 | RI, MS | |
| 6 | p-Cymene | tr | 1008 | 1013 | RI, MS | |
| 7 | Limonene | 0.1 | tr | 1018 | 1025 | RI, MS |
| 8 | (Z)-β-Ocimene | tr | tr | 1036 | 1029 | RI, MS |
| 9 | (E)-Oct-2-en-1-al | tr | 1037 | 1034 | RI, MS | |
| 10 | (E)-β-Ocimene | tr | 1039 | 1041 | RI, MS | |
| 11 | (3E,5E)-3,5-Octadien-2-one | tr | tr | 1048 | 1057 | RI, MS |
| 12 | trans-Linalooloxide (furanoid) | 0.1 | tr | 1056 | 1058 | RI, MS |
| 13 | Non-1-en-3-ol | tr | tr | 1065 | 1068 | RI, MS |
| 14 | p-Cymenene | tr | 1074 | 1076 | RI, MS | |
| 15 | 6-Methyl-3E,5-heptadien-2-one | tr | 1076 | 1076 | RI, MS | |
| 16 | Terpinolene | 0.1 | tr | 1077 | 1078 | RI, MS |
| 17 | Nonanal | 0.2 | 1081 | 1084 | RI, MS | |
| 18 | Linalool | 9.3 | 0.1 | 1085 | 1081 | RI, MS |
| 19 | Limona ketone | tr | 1096 | 1105 | RI, MS | |
| 20 | Non-3-en-2-one | tr | tr | 1113 | 1114 | RI, MS |
| 21 | (2E,6Z)-Nonadienal | tr | 1122 | 1125 | RI, MS | |
| 22 | (E)-Non-2-enal | 0.1 | tr | 1135 | 1233 | RI, MS |
| 23 | Nerol oxide | 3.6 | 0.1 | 1137 | 1137 | RI, MS |
| 24 | p-Ethylbenzaldehyde | tr | 1142 | 1147 | RI, MS | |
| 25 | 4-(2-Methyl-1-cyclohex-1-yn)but-2E-enal | 0.2 | 2.2 | 1150 | 1149 | RI, MS |
| 26 | Cymen-9-ol | 0.1 | tr | 1161 | 1157 | RI, MS |
| 27 | Terpinen-4-ol | tr | tr | 1161 | 1161 | RI, MS |
| 28 | Caprylic acid | 0.1 | 1168 | 1167 | RI, MS | |
| 29 | α-Terpineol | 2.2 | 0.1 | 1172 | 1174 | RI, MS |
| 30 | p-Cumic aldehyde | 0.1 | 0.2 | 1194 | 1214 | RI, MS |
| 31 | β-Cyclocitral | tr | 1201 | 1197 | RI, MS | |
| 32 | Nerol | 1.0 | 1210 | 1215 | RI, MS | |
| 33 | Thymol methyl ether | 7.1 | 1214 | 1215 | RI, MS | |
| 32 + 33 | Nerol + Thymol methyl ether (0.9:0.1) | 26.0 | 1215 | 1215 | RI, MS | |
| 34 | Geraniol | 0.9 | 0.1 | 1234 | 1232 | RI, MS |
| 35 | Geranial | 0.4 | tr | 1242 | 1244 | RI, MS |
| 36 | p-tert-Butylphenol | tr | tr | 1246 | 1250 | RI, MS |
| 37 | α-Ionene | tr | tr | 1252 | 1255 | RI, MS |
| 38 | Pelargonic acid | tr | 0.2 | 1255 | 1260 | RI, MS |
| 39 | Isoprop-1-enyl-2-isopropylbenzene | 0.2 | 1138 | 1140 | RI, MS | |
| 40 | Thymol | 0.1 | 0.1 | 1267 | 1260 | RI, MS |
| 41 | Deca-2,4-dienal | tr | tr | 1272 | 1270 | RI, MS |
| 42 | Carvacrol | tr | 0.1 | 1308 | 1278 | RI, MS |
| 43 | 7α-Silphiperfol-5-ene | tr | 0.1 | 1326 | 1329 | RI, MS |
| 44 | Eugenol | tr | 1329 | 1331 | RI, MS | |
| 45 | Dehydro-ar-ionene | 0.1 | 1330 | 1332 | RI, MS | |
| 46 | Presilphiperfol-7-ene | 0.2 | 1336 | 1333 | RI, MS | |
| 47 | 1,1,4,5-Tetramethylindane | tr | 1341 | 1355 | RI, MS | |
| 48 | 7β-Silphiperfol-5-ene | 0.2 | 0.6 | 1345 | 1350 | RI, MS |
| 49 | α-Longipinene | tr | 0.1 | 1352 | 1358 | RI, MS |
| 50 | Silphiperfol-1-ene | 0.1 | 0.1 | 1360 | 1358 | RI, MS |
| 51 | (E)-β-Damascenone | tr | 1361 | 1361 | RI, MS | |
| 52 | Longicyclene | 0.1 | tr | 1371 | 1376 | RI, MS |
| 53 | α-Copaene | 0.3 | 1373 | 1377 | RI, MS | |
| 54 | Silphiperfol-6-ene | 0.3 | 1375 | 1378 | RI, MS | |
| 55 | Modheph-2-ene | 1.7 | 4.6 | 1381 | 1382 | RI, MS |
| 56 | β-Bourbonene | 0.3 | 1383 | 1386 | RI, MS | |
| 57 | α-Isocomene | 0.8 | 1.8 | 1387 | 1388 | RI, MS |
| 58 | 2,5-Dimethoxy-p-cymene (Thymohydroquinone dimethyl ether) | 7.9 | 6.6 | 1399 | 1399 | RI, MS |
| 59 | Italicene | 0.2 | tr | 1403 | 1404 | RI, MS |
| 60 | β-Isocomene | 0.7 | 1.9 | 1406 | 1407 | RI, MS |
| 61 | Isobornyl isobutyrate | 0.6 | 1410 | 1410 | RI, MS | |
| 62 | 7,8-Dihydro-β-ionone | tr | tr | 1412 | 1411 | RI, MS |
| 63 | (E)-β-Caryophyllene | 0.3 | 0.4 | 1417 | 1421 | RI, MS |
| 64 | Geranylacetone | 0.3 | 0.1 | 1427 | 1428 | RI, MS |
| 65 | trans-α-Bergamotene | 0.1 | 0.1 | 1432 | 1434 | RI, MS |
| 66 | epi-β-Santalene | 0.2 | 0.3 | 1442 | 1446 | RI, MS |
| 67 | α-Himachalene | tr | 0.1 | 1445 | 1447 | RI, MS |
| 68 | α-Humulene | tr | 0.1 | 1451 | 1455 | RI, MS |
| 69 | β-Santalene | 0.1 | tr | 1454 | 1459 | RI, MS |
| 70 | 8,9-Didehydrothymyl isobutyrate | tr | 0.3 | 1456 | 1458 | RI, MS |
| 71 | Thymyl isobutyrate | 0.4 | 1457 | 1462 | RI, MS | |
| 72 | allo-Aromadendrene | 0.6 | 1458 | 1462 | RI, MS | |
| 73 | (E)-β-Ionone | 0.4 | 0.1 | 1462 | 1468 | RI, MS |
| 74 | 6-Isopropenyl-4,8α-dimethyl-1,2,3,5,6,7,8,8α-octahydro-2-naphthalenyl acetate | 0.1 | tr | 1466 | MS | |
| 75 | cis-β-Guaiene | tr | 1468 | 1469 | RI, MS | |
| 76 | Neryl isobutyrate | 12.1 | 2.4 | 1471 | 1469 | RI, MS |
| 77 | γ-Himachalene | tr | tr | 1474 | 1479 | RI, MS |
| 78 | Thujopsadiene | 0.2 | tr | 1478 | 1470 | RI, MS |
| 79 | Patchoulene | 0.6 | 0.2 | 1482 | 1473 | RI, MS |
| 80 | 1,4-Dioxaspiro [4,6]undec-6-yl acetate | 0.1 | 0.8 | 1486 | MS | |
| 81 | Dibenzofuran | tr | tr | 1490 | 1504 | RI, MS |
| 82 | Bicyclogermacrene | 0.2 | 1491 | 1494 | RI, MS | |
| 83 | α-Selinene | 1.2 | tr | 1492 | 1495 | RI, MS |
| 84 | β-Bisabolene | 0.1 | 0.2 | 1500 | 1500 | RI, MS |
| 85 | Isoshyobunone | 0.9 | 5.6 | 1507 | 1518 | RI, MS |
| 86 | δ-Cadinene | 0.2 | 0.1 | 1514 | 1520 | RI, MS |
| 87 | cis/trans-Calamenene | tr | 0.2 | 1520 | 1523 | RI, MS |
| 88 | 10-epi-Italicene ether | 0.1 | tr | 1522 | 1531 | RI, MS |
| 89 | (E)-β-Caryophyllene oxide | tr | 0.1 | 1542 | 1546 | RI, MS |
| 90 | (Z)-Nerolidol | 0.1 | 0.2 | 1546 | 1546 | RI, MS |
| 91 | (E)-Nerolidol | 1.5 | 1.1 | 1556 | 1555 | RI, MS |
| 92 | Neryl isovalerate | 0.4 | 0.3 | 1562 | 1565 | RI, MS |
| 93 | Caryophyllene epoxide | 0.5 | 4.0 | 1570 | 1571 | RI, MS |
| 94 | Unidentified (MS: 148/133/91/187 M220) | 0.1 | 0.4 | 1579 | ||
| 95 | Isoaromadendrene epoxide | 0.2 | 1.1 | 1582 | 1592 | RI, MS |
| 96 | β-Himachalene epoxide | 0.1 | 1.3 | 1594 | 1594 | RI, MS |
| 97 | Unidentified (MS: 162/147/120/173 M206) | 0.4 | 1597 | |||
| 98 | Unidentified (MS: 162/147/120/173 M206) | 1.2 | 0.2 | 1600 | ||
| 99 | Zierone | 0.1 | 0.8 | 1611 | 1582 3 | RI, MS |
| 100 | Torryeol | tr | 1612 | 1607 | RI, MS | |
| 101 | 2,5-Ditert-butylbenzo-1,4-quinone | 0.2 | 0.2 | 1613 | 1613 | RI, MS |
| 102 | Cubenol | tr | 0.4 | 1619 | 1620 | RI, MS |
| 103 | Caryophylla-3(15),7(14)-dien-6-ol | 0.1 | 0.3 | 1625 | 1630 | RI, MS |
| 104 | T-Muurolol | 1.0 | 1.5 | 1631 | 1633 | RI, MS |
| 105 | β-Selinenol | 0.3 | 1.4 | 1635 | 1638 | RI, MS |
| 106 | α-Cadinol | 0.2 | 0.4 | 1638 | 1643 | RI, MS |
| 107 | Eudesm-4(15)-en-7-ol | tr | 0.7 | 1640 | 1643 | RI, MS |
| 108 | Unidentified (MS: 162/147/91/119/41) | tr | 0.1 | 1649 | ||
| 109 | 6-Methoxythymyl isobutyrate | 1.7 | 1.5 | 1656 | 1659 | RI, MS |
| 110 | α-Acorenol | 0.1 | 0.2 | 1667 | 1667 | RI, MS |
| 111 | 2,3-Dihydrofarnesol | 0.1 | 0.2 | 1670 | 1674 | RI, MS |
| 112 | 6-Methoxy-8,9-didehydrothymyl isobutyrate | tr | 0.1 | 1676 | 1676 | RI, MS |
| 113 | 10-Isobutyryloxy-8,9-didehydrothymol methyl ether | tr | tr | 1684 | 1684 | RI, MS |
| 114 | β-(E)-Santalol | tr | tr | 1684 | 1680 | RI, MS |
| 115 | n-Pentadecanal | 0.4 | 1692 | 1702 | RI, MS | |
| 116 | Phenantrene | 0.1 | 1747 | 1740 | RI, MS | |
| 117 | β-Santalol acetate | 0.2 | tr | 1801 | 1800 | RI, MS |
| 118 | Unidentified (MS: 71/119/134/43 M223) | 0.5 | 1818 | |||
| 119 | Diisobutyl phthalate (artifact) | 2.9 | 1.5 | 1826 | 1826 | RI, MS |
| 120 | Dihydroisoalantolactone | 0.2 | 1860 | 1875 | RI, MS | |
| 121 | Alantolactone | 0.1 | 29.3 | 1875 | 1873 | RI, MS |
| 122 | 9-Isobutyryloxythymyl isobutyrate | 0.1 | 1.1 | 1878 | 1884 | RI, MS |
| 123 | 10-Isobutyryloxy-8,9-didehydrothymyl isobutyrate | 0.1 | 0.8 | 1890 | 1887 | RI, MS |
| 124 | (5E,9E)-Farnesylacetone | tr | 0.4 | 1895 | 1885 | RI, MS |
| 125 | Isoalantolactone | 0.7 | 0.7 | 1900 | 1912 | RI, MS |
| 126 | Alloalantolactone (1-Deoxyivangustin/(+)-Diplophyllin) | 4.4 | 1914 | 1916 3 | RI, MS | |
| 127 | Dibutyl phthalate (artifact) | 0.4 | tr | 1920 | 1912 | RI, MS |
| 128 | Isoalantodiene (3-Dehydroalantolactone) | 0.1 | 1922 | MS | ||
| 129 | Unidentified (MS: 121/91/79/95/119/145/190/105/41 M232) | 0.6 | 1931 | |||
| 130 | Palmitic acid | 0.2 | 0.2 | 1940 | 1952 | RI, MS |
| 131 | 9-(2-Methylbutyryloxy)thymyl isobutyrate | 0.2 | 1964 | 1964 | RI, MS | |
| 132 | 10-Isobutyryloxy-8,9-epoxythymyl isobutyrate | tr | 2.2 | 1985 | 1972 | RI, MS |
| 133 | (E,E)-Geranyl linalool | 6.5 | tr | 2012 | 2020 | RI, MS |
| 134 | 10-(2-methylbutyryloxy)-8,9-epoxythymyl isobutyrate | 0.4 | 0.3 | 2068 | 2084 | RI, MS |
| 135 | Tricosane | 0.2 | 2293 | 2300 | RI, MS | |
| 136 | Tetracosane | tr | 2394 | 2400 | RI, MS | |
| 137 | Heptacosane | 0.2 | 0.1 | 2659 | 2700 | RI, MS |
| Sum of identified | 96.0 | 96.7 | ||||
| Yield of the essential oil [%] | 0.08 | 0.13 | ||||
| Tested Chemicals | Minimum Inhibitory Concentration (µg/mL) | |||
|---|---|---|---|---|
| Staphylococcus aureus ATCC 29213 (MSSA) | Staphylococcus aureus ATCC 43300 (MRSA) | Escherichia coli ATCC 25922 | Pseudomonas aeruginosa ATCC 27853 | |
| APEO | 2080.0 ± 0.0 | 2080.0 ± 0.0 | nd | nd |
| REO | 3380.0 ± 0.0 | 3380.0 ± 0.0 | nd | nd |
| Thymol a | 0.9 ± 0.0 | 0.9 ± 0.0 | 7.5 ± 0.0 | 7.5 ± 0.0 |
| Gentamycin sulphate | 0.31 ± 0.0 | 19.5 ±0.0 | 0.61 ± 0.0 | 1.22 ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wajs-Bonikowska, A.; Malarz, J.; Szoka, Ł.; Kwiatkowski, P.; Stojakowska, A. Composition and Activities of Carpesium macrocephalum Franch. & Sav. Essential Oils. Molecules 2024, 29, 4658. https://doi.org/10.3390/molecules29194658
Wajs-Bonikowska A, Malarz J, Szoka Ł, Kwiatkowski P, Stojakowska A. Composition and Activities of Carpesium macrocephalum Franch. & Sav. Essential Oils. Molecules. 2024; 29(19):4658. https://doi.org/10.3390/molecules29194658
Chicago/Turabian StyleWajs-Bonikowska, Anna, Janusz Malarz, Łukasz Szoka, Paweł Kwiatkowski, and Anna Stojakowska. 2024. "Composition and Activities of Carpesium macrocephalum Franch. & Sav. Essential Oils" Molecules 29, no. 19: 4658. https://doi.org/10.3390/molecules29194658
APA StyleWajs-Bonikowska, A., Malarz, J., Szoka, Ł., Kwiatkowski, P., & Stojakowska, A. (2024). Composition and Activities of Carpesium macrocephalum Franch. & Sav. Essential Oils. Molecules, 29(19), 4658. https://doi.org/10.3390/molecules29194658