
Using Extracted Sugars from Spoiled Date Fruits as a Sustainable Feedstock for Ethanol Production by New Yeast Isolates




Abstract

1. Introduction
2. Results
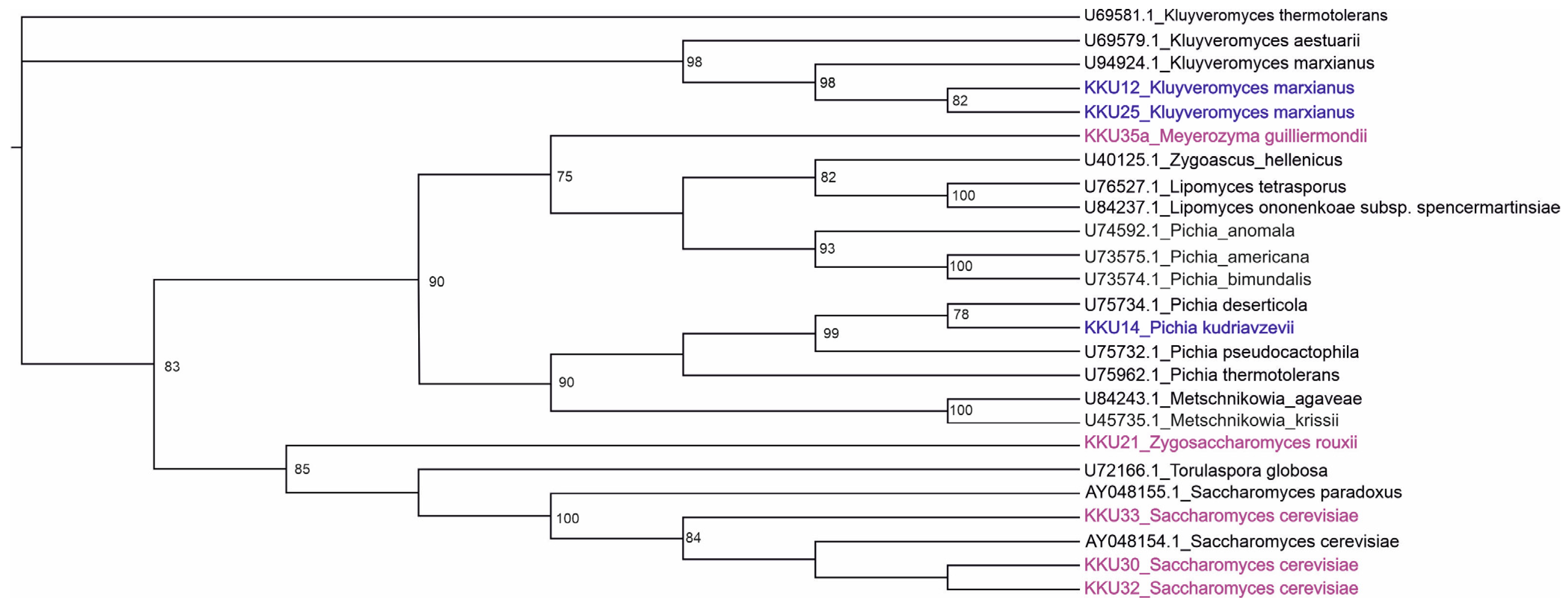
2.1. Identification of Yeast Strains
2.2. Extraction of Sugars from SDF
2.3. Ethanol Production from SDF Extract
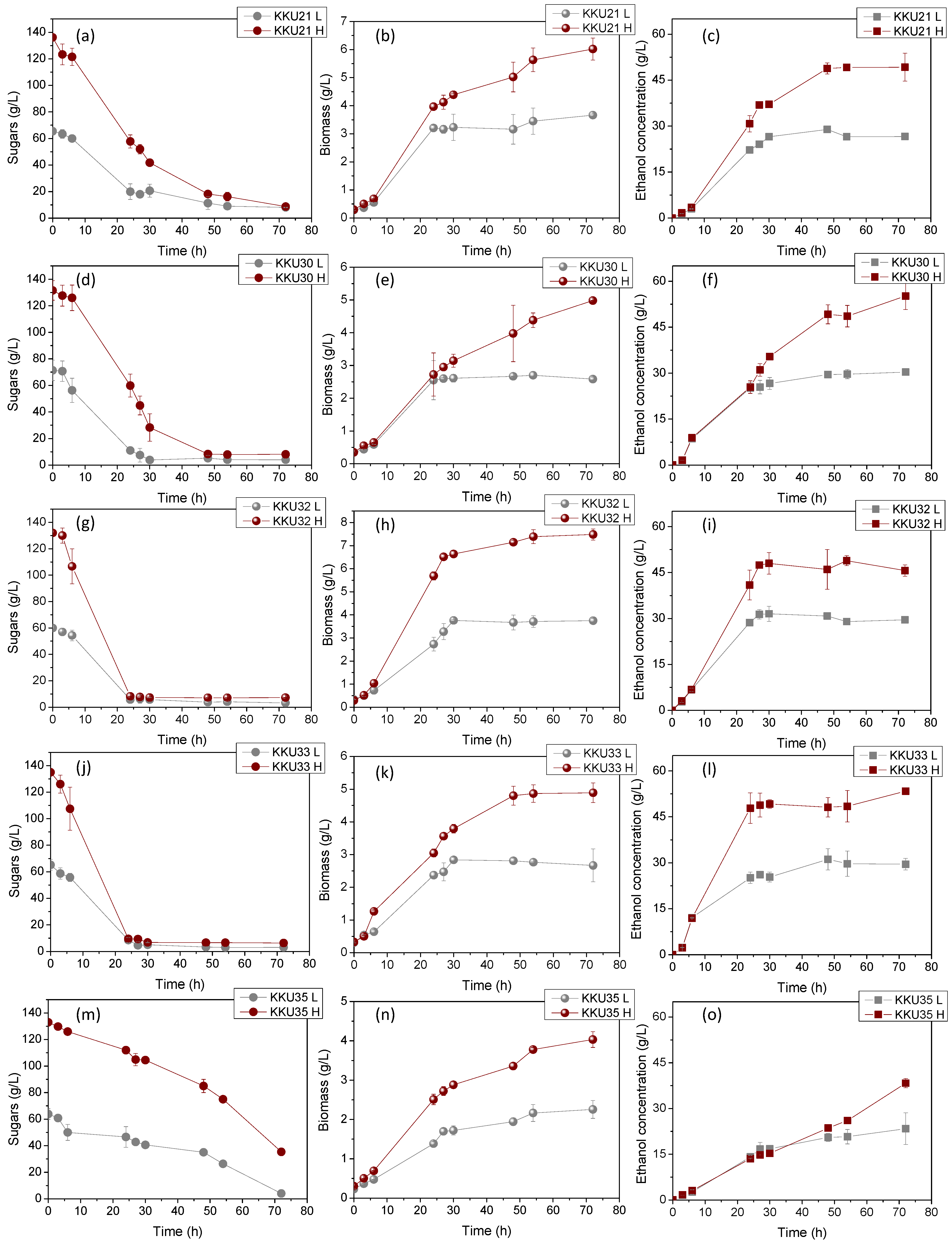
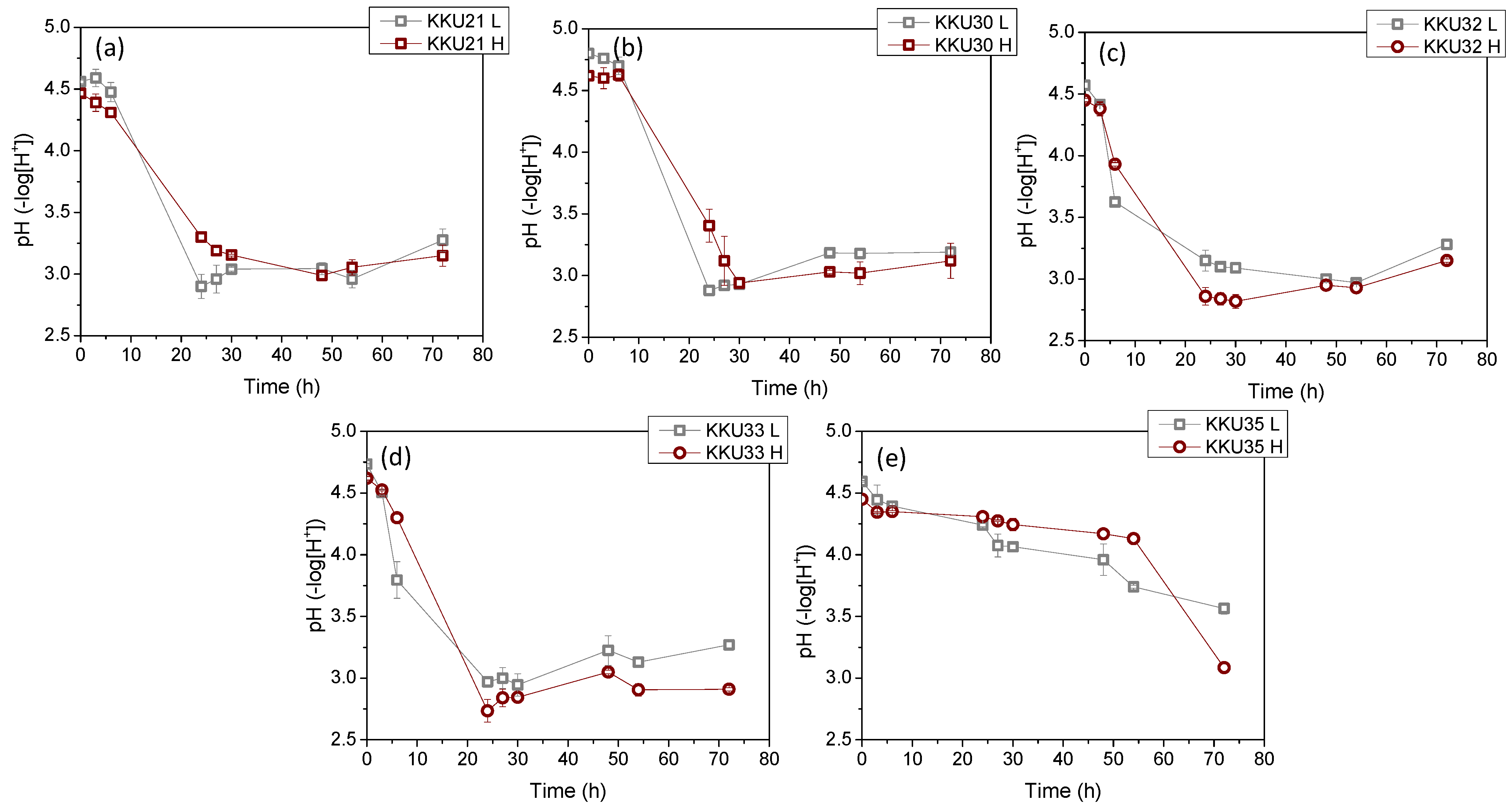
2.3.1. Batch Experiments with Mono-Cultures
2.3.2. Scaling Up with Co-Culture
3. Discussion
4. Materials and Methods
4.1. Feedstock
4.2. Extraction of Sugars from SDF
4.3. Experimental Design and Statistical Analysis
4.4. Identification of Microorganisms
4.5. Bioethanol Production
4.5.1. Isolates and Pre-Cultures
4.5.2. Fermentation Experiments
4.6. Analytical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barua, S.; Sahu, D.; Sultana, F.; Baruah, S.; Mahapatra, S. Bioethanol, internal combustion engines and the development of zero-waste biorefineries: An approach towards sustainable motor spirit. RSC Sustain. 2023, 1, 1065–1084. [Google Scholar] [CrossRef]
- Lahiri, A.; Daniel, S.; Kanthapazham, R.; Vanaraj, R.; Thambidurai, A.; Peter, L.S. A critical review on food waste management for the production of materials and biofuel. J. Hazard. Mater Adv. 2023, 10, 100266. [Google Scholar] [CrossRef]
- Castillo, A.B.; Cortes, D.J.D.; Sorino, C.F.; Soriño, C.K.P.; El-Naas, M.H.; Ahmed, T. Bioethanol Production from Waste and Nonsalable Date Palm (Phoenix dactylifera L.) Fruits: Potentials and Challenges. Sustainability 2023, 15, 2937. [Google Scholar] [CrossRef]
- Hashem, M.; Al-Qahtani, M.S.; Alamri, S.A.; Moustafa, Y.S.; Lyberatos, G.; Ntaikou, I. Valorizing food wastes: Assessment of novel yeast strains for enhanced production of single-cell protein from wasted date molasses. Biomass Convers. Biorefin. 2022, 12, 4491–4502. [Google Scholar] [CrossRef]
- Troncoso-Ortega, E.; Valenzuela, R.; Reyes-Contreras, P.; Castaño-Rivera, P.; Schiappacasse, L.-N.; Parra, C. Maximizing Bioethanol Production from Eucalyptus globulus Using Steam Explosion Pretreatment: A Multifactorial Design and Fermenter Development for High Solid Loads. Fermentation 2023, 9, 965. [Google Scholar] [CrossRef]
- Ntaikou, I.; Siankevich, S.; Lyberatos, G. Effect of thermo-chemical pretreatment on the saccharification and enzymatic digestibility of olive mill stones and their bioconversion towards alcohols. Environ. Sci. Pollut. Res. 2021, 28, 24570–24579. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, G.; Balbi, M.d.P.; Salierno, G.; Cassanello, M.; De Blasio, C.; Galvagno, M. A Cleaner Delignification of Urban Leaf Waste Biomass for Bioethanol Production, Optimised by Experimental Design. Processes 2022, 10, 943. [Google Scholar] [CrossRef]
- Antonopoulou, G.; Ntaikou, I.; Pastore, C.; di Bitonto, L.; Bebelis, S.; Lyberatos, G. An overall perspective for the energetic valorization of household food waste using microbial fuel cell technology of its extract, coupled with anaerobic digestion of the solid residue. Appl. Energy 2019, 238, 239–248. [Google Scholar] [CrossRef]
- Antonopoulou, G.; Alexandropoulou, M.; Ntaikou, I.; Lyberatos, G. From waste to fuel: Energy recovery from household food waste via its bioconversion to energy carriers based on microbiological processes. Sci. Total Environ. 2020, 732, 139230. [Google Scholar] [CrossRef]
- Lamidi, S.; Olaleye, N.; Bankole, Y.; Obalola, A.; Aribike, E.; Adigun, I. Applications of Response Surface Methodology (RSM) in Product Design, Development, and Process Optimization; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Szpisjak-Gulyas, N.; Al-Tayawi, A.N.; Horvath, Z.H.; Laszlo, Z.; Kertesz, S.; Hodur, C. Methods for experimental design, central composite design and the Box–Behnken design, to optimise operational parameters: A review. Acta Aliment. 2023, 52, 521–537. [Google Scholar] [CrossRef]
- Hashem, M.; Alamri, S.A.; Asseri, T.A.Y.; Mostafa, Y.S.; Lyberatos, G.; Ntaikou, I. On the Optimization of Fermentation Conditions for Enhanced Bioethanol Yields from Starchy Biowaste via Yeast Co-Cultures. Sustainability 2021, 13, 1890. [Google Scholar] [CrossRef]
- Ntaikou, I.; Antonopoulou, G.; Lyberatos, G. Sustainable Second-Generation Bioethanol Production from Enzymatically Hydrolyzed Domestic Food Waste Using Pichia anomala as Biocatalyst. Sustainability 2021, 13, 259. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef]
- Molinet, J.; Cubillos, F.A. Wild Yeast for the Future: Exploring the Use of Wild Strains for Wine and Beer Fermentation. Front. Genet. 2020, 11, 589350. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.; Serra, F.; Tárraga, J.; Medina, I.; Carbonell, J.; Pulido, L.; de María, A.; Capella-Gutíerrez, S.; Huerta-Cepas, J.; Gabaldón, T.; et al. Phylemon 2.0: A suite of web-tools for molecular evolution, phylogenetics, phylogenomics and hypotheses testing. Nucleic Acids Res. 2011, 39 (Suppl. S2), W470–W474. [Google Scholar] [CrossRef] [PubMed]
- Ntaikou, I.; Alexandropoulou, M.; Kamilari, M.; Alamri, S.; Moustafa, Y.; Hashem, M.; Antonopoulou, G.; Lyberatos, G. Saccharification of starchy food waste through thermochemical and enzymatic pretreatment, towards enhanced bioethanol production via newly isolated non-conventional yeast strains. Energy 2023, 281, 128259. [Google Scholar] [CrossRef]
- Ruchala, J.; Kurylenko, O.O.; Dmytruk, K.V.; Sibirny, A.A. Construction of advanced producers of first- and second-generation ethanol in Saccharomyces cerevisiae and selected species of non-conventional yeasts (Scheffersomyces stipitis, Ogataea polymorpha). J. Ind. Microbiol. Biotechnol. 2020, 47, 109–132. [Google Scholar] [CrossRef] [PubMed]
- Wingren, A.; Soderstrom, J.; Galbe, M.; Zacchi, G. Process considerations and economic evaluation of two-step steam pretreatment for production of fuel ethanol from softwood. Biotechnol. Prog. 2004, 20, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Farouk, W.M.; Abdullah, A.S.; Mohammed, S.A.; Alawee, W.H.; Omara, Z.M.; Essa, F.A. Modeling and optimization of working conditions of pyramid solar still with different nanoparticles using response surface methodology. Case Stud. Therm. Eng. 2022, 33, 101984. [Google Scholar] [CrossRef]
- Anwar, K.; Said, M.; Amin, M.A.M. Overview on the Response Surface Methodology (RSM)in Extraction Processes. J. Appl. Sci. Proc. Eng. 2005, 2, 8–17. [Google Scholar] [CrossRef]
- Messadi, N.; Mechmeche, M.; Setti, K.; Tizemmour, Z.; Hamdi, M.; Kachouri, F. Optimization of Extraction Parameters and Characterization of Tunisian Date Extract: A Scientific Approach Toward Their Utilization. Sugar Tech 2023, 25, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Ntaikou, I.; Menis, N.; Alexandropoulou, M.; Antonopoulou, G.; Lyberatos, G. Valorization of kitchen biowaste for ethanol production via simultaneous saccharification and fermentation using co-cultures of the yeasts Saccharomyces cerevisiae and Pichia stipitis. Bioresour. Technol. 2018, 263, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Dakal, T.C.; Solieri, L.; Giudici, P. Adaptive response and tolerance to sugar and salt stress in the food yeast Zygosaccharomyces rouxii. Int. J. Food Microbiol. 2014, 18, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Gao, H.; Qian, X.; Jiang, Y.; Zhou, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Biotechnological applications of the non-conventional yeast Meyerozyma guilliermondii. Biotechnol. Adv. 2021, 1, 107674. [Google Scholar] [CrossRef]
- Cui, R.; Liu, X.; Wang, Q.; Wang, S.; He, J.; Zhou, S.; Zhao, G.; Yao, Y. Effects of co-fermentation of high-yield ester Zygosaccharomyces rouxii and Saccharomyces cerevisiae on the quality of rice wine. JSFA Rep. 2024, 4, 78–91. [Google Scholar] [CrossRef]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. Whole-genome comparison reveals novel genetic elements that characterize the genome of industrial strains of Saccharomyces cerevisiae. PLoS Genet. 2011, 7, e1001287. [Google Scholar] [CrossRef]
- Sazegari, S.; Niazi, A.; Zinati, Z.; Eskandari, M.H. Mining transcriptomic data to identify Saccharomyces cerevisiae signatures related to improved and repressed ethanol production under fermentation. PLoS ONE 2022, 17, e0259476. [Google Scholar] [CrossRef]
- Kurtzman, C.P. Recognition of yeast species from gene sequence comparisons. Open Appl. Inform. J. 2011, 5, 20–29. [Google Scholar] [CrossRef]
- Tompson, J.D.; Higgins, D.J.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2020, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- APHA; AWWA; WPCF. Standard Methods for the Examination of Water and Wastewater; Franson, M.A., Ed.; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Quémeneur, M.; Trably, E.; Steyer, J.-P.; Carrère, H. Do furanic and phenolic compounds of lignocellulosic and algae biomass hydrolyzate inhibit anaerobic mixed cultures? A comprehensive review. Biotechnol. Adv. 2014, 32, 934–951. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Best Hit on GENBANK (Accession Number) | Similarity (%) | Genbank Accession Number |
|---|---|---|---|
| KKU21 | Zygosaccharomyces rouxii isolate D13 (MK341573.1) | 99.08 | OR245523 |
| KKU30 | Saccharomyces cerevisiae isolate WHY-5 (MG641150.1) | 99.3 | OR245524 |
| KKU32 | Saccharomyces cerevisiae strain GV5 03 (KP250857.1) | 99.17 | OR245525 |
| KKU33 | Saccharomyces cerevisiae strain GV5 03 (KP250857.1) | 99.65 | OR245526 |
| KKU35 | Meyerozyma guilliermondii strain ML4 (MK907983.1) | 98.62 | OR245527 |
| Sources | Sum of Squares | DF | Mean Square | F-Value | p-Value * |
|---|---|---|---|---|---|
| Linear Model | 9394.68 | 9 | 1043.85 | 7.92 | 0.0062 (s) |
| A—Temperature | 1192.31 | 1 | 1192.31 | 9.05 | 0.0197 |
| B—Organic loading | 6224.49 | 1 | 6224.49 | 47.24 | 0.0002 |
| C—Time | 1742.42 | 1 | 1742.42 | 13.22 | 00083 |
| AB | 5.38 | 1 | 5.38 | 0.0409 | 0.8456 |
| AC | 7.49 | 1 | 7.49 | 0.0569 | 0.8183 |
| BC | 20.25 | 1 | 20.25 | 0.1597 | 0.7067 |
| A2 | 11.09 | 1 | 11.09 | 0.0842 | 0.7801 |
| B2 | 163.46 | 1 | 163.46 | 1.24 | 0.3021 |
| C2 | 25.99 | 1 | 25.99 | 0.1972 | 0.6704 |
| Residual | 922.27 | 7 | 131.75 | ||
| Lack of Fit | 50.60 | 3 | 16.87 | 0.0774 | 0.9689 (ns) |
| Source | Sum of Squares | DF | Mean Square | F-Value | p-Value * |
|---|---|---|---|---|---|
| Linear Model | 0.0327 | 3 | 0.0109 | 10.65 | 0.0008 (s) |
| A—Temperature | 0.0153 | 1 | 0.0153 | 14.97 | 0.0019 |
| B—Organic loading | 0.0010 | 1 | 0.0010 | 0.9772 | 0.3409 |
| C—Time | 0.0164 | 1 | 0.0164 | 16.01 | 0.0015 |
| Residual | 0.0133 | 13 | 0.0010 | ||
| Lack of Fit | 0.0035 | 9 | 0.0004 | 0.1608 | 0.9890 (ns) |
| Response | Final Equations in Terms of Actual and Coded Factors | |
|---|---|---|
| Concentration (R1) | Coded | R1 = 100.453 + 12.2081 × A + 27.8938 × B + 14.7581 × C |
| Actual = | 49.3692 + 1.22081 × A + 2.78938 × B + 1.47581 × C | |
| Extraction Efficiency (R2) | Coded | R2 = 0.3398 + 0.0438 × A − 0.0112 × B + 0.0453 × C |
| Actual = | 0.151401 + 0.004379 × A − 0.00119 × B + 0.004528 × C | |
| Isolate | Extract Concentration | rS, g/L·h | rEtOH, g/L·h | YX/S, g Biomass/g Sugars Cons. | YEtOH/S, g Eth/g Sugars Cons. | Sugar Consumption, % |
|---|---|---|---|---|---|---|
| KKU21 | L | 1.05 ± 0.02 | 0.60 ± 0.08 | 0.056 ± 0.001 | 0.46 ± 0.01 | 89.1 ± 0.9 |
| H | 1.77 ± 0.11 | 0.91 ± 0.02 | 0.045 ± 0.001 | 0.39 ± 0.01 | 93.6 ± 0.3 | |
| KKU30 | L | 2.25 ± 0.04 | 0.89 ± 0.01 | 0.035 ± 0.001 | 0.45 ± 0.02 | 94.8 ± 0.5 |
| H | 2.29 ± 0.09 | 0.90 ± 0.14 | 0.038 ± 0.000 | 0.45 ± 0.02 | 94.1 ± 0.1 | |
| KKU32 | L | 2.87 ± 0.08 | 1.05 ± 0.01 | 0.062 ± 0.001 | 0.48 ± 0.01 | 94.3 ± 0.8 |
| H | 4.16 ± 0.25 | 1.34 ± 0.06 | 0.058 ± 0.002 | 0.36 ± 0.01 | 94.5 ± 0.1 | |
| KKU33 | L | 5.86 ± 0.05 | 1.30 ± 0.11 | 0.039 ± 0.007 | 0.47 ± 0.00 | 94.1 ± 0.2 |
| H | 4.27 ± 0.25 | 1.64 ± 0.29 | 0.035 ± 0.002 | 0.43 ± 0.01 | 93.9 ± 0.5 | |
| KKU35 | L | 0.83 ± 0.02 | 0.32 ± 0.01 | 0.034 ± 0.003 | 0.48 ± 0.01 | 92.2 ± 0.1 |
| H | 1.36 ± 0.05 | 0.53 ± 0.05 | 0.038 ± 0.013 | 0.39 ± 0.02 | 73.6 ± 0.2 |
| Run | A | B | C | Concentration, g/L | Yield, g Sugars/g SDF |
|---|---|---|---|---|---|
| 1 | 0 | 1 | 1 | 143.10 | 0.373 |
| 2 | 0 | 1 | −1 | 113.58 | 0.282 |
| 3 | 1 | −1 | 0 | 84.66 | 0.395 |
| 4 | −1 | 1 | 0 | 116.23 | 0.284 |
| 5 | −1 | 0 | 1 | 103.10 | 0.341 |
| 6 | 0 | 0 | 0 | 100.45 | 0.339 |
| 7 | 0 | 0 | 0 | 100.45 | 0.339 |
| 8 | 0 | 0 | 0 | 100.45 | 0.339 |
| 9 | −1 | −1 | 0 | 60.45 | 0.308 |
| 10 | 1 | 0 | −1 | 97.80 | 0.338 |
| 11 | 0 | 0 | 1 | 115.21 | 0.385 |
| 12 | 0 | 0 | 0 | 100.45 | 0.339 |
| 13 | 1 | 0 | 1 | 127.32 | 0.428 |
| 14 | 1 | 1 | 0 | 140.45 | 0.371 |
| 15 | 0 | −1 | 1 | 87.31 | 0.397 |
| 16 | 0 | −1 | −1 | 57.8 | 0.306 |
| 17 | −1 | 0 | −1 | 73.58 | 0.250 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonopoulou, G.; Kamilari, M.; Georgopoulou, D.; Ntaikou, I. Using Extracted Sugars from Spoiled Date Fruits as a Sustainable Feedstock for Ethanol Production by New Yeast Isolates. Molecules 2024, 29, 3816. https://doi.org/10.3390/molecules29163816
Antonopoulou G, Kamilari M, Georgopoulou D, Ntaikou I. Using Extracted Sugars from Spoiled Date Fruits as a Sustainable Feedstock for Ethanol Production by New Yeast Isolates. Molecules. 2024; 29(16):3816. https://doi.org/10.3390/molecules29163816
Chicago/Turabian StyleAntonopoulou, Georgia, Maria Kamilari, Dimitra Georgopoulou, and Ioanna Ntaikou. 2024. "Using Extracted Sugars from Spoiled Date Fruits as a Sustainable Feedstock for Ethanol Production by New Yeast Isolates" Molecules 29, no. 16: 3816. https://doi.org/10.3390/molecules29163816
APA StyleAntonopoulou, G., Kamilari, M., Georgopoulou, D., & Ntaikou, I. (2024). Using Extracted Sugars from Spoiled Date Fruits as a Sustainable Feedstock for Ethanol Production by New Yeast Isolates. Molecules, 29(16), 3816. https://doi.org/10.3390/molecules29163816