
Phytotoxicity of Bisphenol A to Allium cepa Root Cells Is Mediated through Growth Hormone Gibberellic Acid and Reactive Oxygen Species




Abstract
1. Introduction
2. Results and Discussion
2.1. Impact of BPA on A. cepa Root Growth
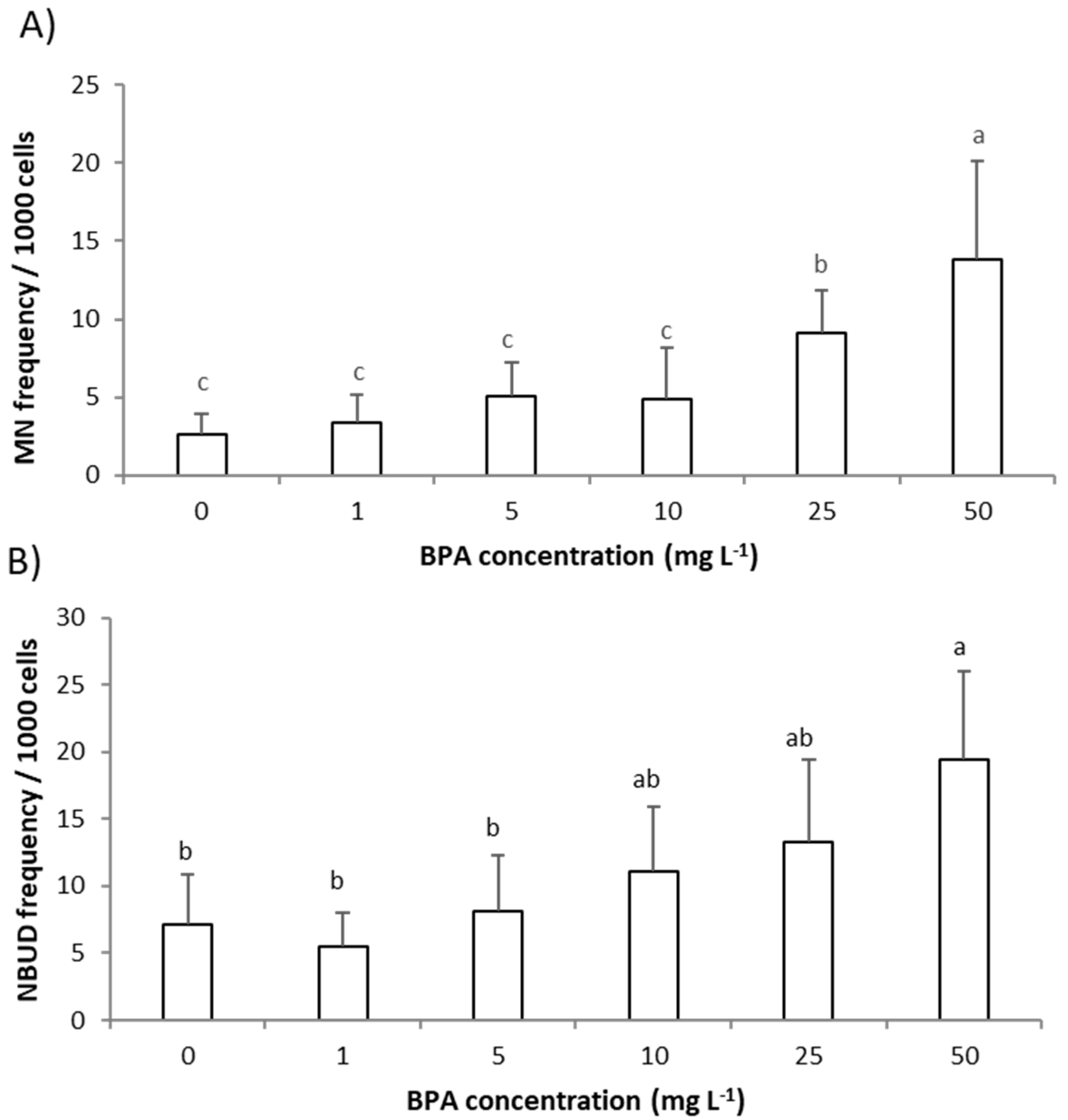
2.2. Genotoxic Effect of BPA to A. cepa Root Meristem Cells
2.3. Impact of BPA on the Level of ROS in A. cepa Root Cells
2.4. Impact of BPA on the Level of Oxidative Damage and Superoxide Dismutase (SOD) Activity in A. cepa Root Cells
2.5. Impact of BPA on the Level of Phytochemicals in A. cepa Root Cells
2.6. Principle Component Analysis (PCA)
3. Materials and Methods
3.1. Chemicals and Preparation of BPA Solution
3.2. Allium Test
3.3. Determination of Mitotic Index and Micronucleus
3.4. Determination of GA3 Content
3.5. Determination of ROS In Situ
3.6. Determination of Oxidative Stress Parameters
3.7. Determination of Phytochemicals
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Naomi, R.; Yazid, M.D.; Bahari, H.; Keong, Y.Y.; Rajandram, R.; Embong, H.; Teoh, S.H.; Halim, S.; Othman, F. Bisphenol A (BPA) Leading to Obesity and Cardiovascular Complications: A Compilation of Current In Vivo Study. Int. J. Mol. Sci. 2022, 23, 2969. [Google Scholar] [CrossRef]
- Vehar, A.; Kovačič, A.; Hvala, N.; Škufca, D.; Levstek, M.; Stražar, M.; Žgajnar Gotvajn, A.; Heath, E. An Assessment of Mass Flows, Removal and Environmental Emissions of Bisphenols in a Sequencing Batch Reactor Wastewater Treatment Plant. Molecules 2022, 27, 8634. [Google Scholar] [CrossRef]
- Cimmino, I.; Fiory, F.; Perruolo, G.; Miele, C.; Beguinot, F.; Formisano, P.; Oriente, F. Potential Mechanisms of Bisphenol A (BPA) Contributing to Human Disease. Int. J. Mol. Sci. 2020, 21, 5761. [Google Scholar] [CrossRef]
- Erler, C.; Novak, J. Bisphenol a Exposure: Human Risk and Health Policy. J. Pediatr. Nurs. 2010, 25, 400–407. [Google Scholar] [CrossRef]
- Ali, I.; Liu, B.; Farooq, M.A.; Islam, F.; Azizullah, A.; Yu, C.; Su, W.; Gan, Y. Toxicological Effects of Bisphenol A on Growth and Antioxidant Defense System in Oryza sativa as Revealed by Ultrastructure Analysis. Ecotoxicol. Environ. Saf. 2016, 124, 277–284. [Google Scholar] [CrossRef]
- Rotimi, O.A.; Olawole, T.D.; De Campos, O.C.; Adelani, I.B.; Rotimi, S.O. Bisphenol A in Africa: A Review of Environmental and Biological Levels. Sci. Total Environ. 2021, 764, 142854. [Google Scholar] [CrossRef]
- Malea, P.; Kokkinidi, D.; Kevrekidou, A.; Adamakis, I.-D.S. The Enzymatic and Non-Enzymatic Antioxidant System Response of the Seagrass Cymodocea nodosa to Bisphenol-A Toxicity. Int. J. Mol. Sci. 2022, 23, 1348. [Google Scholar] [CrossRef]
- Careghini, A.; Mastorgio, A.F.; Saponaro, S.; Sezenna, E. Bisphenol A, Nonylphenols, Benzophenones, and Benzotriazoles in Soils, Groundwater, Surface Water, Sediments, and Food: A Review. Environ. Sci. Pollut. Res. 2015, 22, 5711–5741. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, L.; Zhou, Q.; Huang, X. Hazards of Bisphenol A (BPA) Exposure: A Systematic Review of Plant Toxicology Studies. J. Hazard. Mater. 2020, 384, 121488. [Google Scholar] [CrossRef]
- European Commission; Joint Research Centre; Institute for Health and Consumer Protection. Updated European Union Risk Assessment Report: 4,4′-Isopropylidenediphenol (Bisphenol-A): Human Health Addendum of February 2008; Publications Office: Luxembourg, 2010. [Google Scholar]
- Chen, L.; Chen, J.; Zhou, G.; Wang, Y.; Xu, C.; Wang, X. Molecular Dynamics Simulations of the Permeation of Bisphenol A and Pore Formation in a Lipid Membrane. Sci. Rep. 2016, 6, 33399. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Wang, S.; Yang, Q.; Zhou, Q.; Huang, X. A Preliminary Analysis of the Effects of Bisphenol A on the Plant Root Growth via Changes in Endogenous Plant Hormones. Ecotoxicol. Environ. Saf. 2018, 150, 152–158. [Google Scholar] [CrossRef]
- Siddiqui, S.; Al Amri, S.A.M.; Al Ghamdy, H.A.; Alqahtani, W.S.S.; Alquyr, S.M.; Yassin, H.M. Impact of Bisphenol A on Seed Germination, Radicle Length and Cytogenetic Alterations in Pisum sativum L. Caryologia 2021, 74, 103–109. [Google Scholar] [CrossRef]
- Dogan, M.; Yumrutas, O.; Saygideger, S.; Korkunc, M.; Gulnaz, O.; Sokmen, A. Effects of Bisphenol A and Tetrabromobisphenol A on Chickpea Roots in Germination Stage. Am.-Eurasian J. Agric. Environ. Sci. 2010, 9, 186–192. [Google Scholar]
- Fu, W.; Chen, X.; Zheng, X.; Liu, A.; Wang, W.; Ji, J.; Wang, G.; Guan, C. Phytoremediation Potential, Antioxidant Response, Photosynthetic Behavior and Rhizosphere Bacterial Community Adaptation of Tobacco (Nicotiana tabacum L.) in a Bisphenol A-Contaminated Soil. Environ. Sci. Pollut. Res. 2022, 29, 84366–84382. [Google Scholar] [CrossRef]
- Kaźmierczak, A.; Kornaś, A.; Mościpan, M.; Łęcka, J. Influence of Bisphenol A on Growth and Metabolism of Vicia faba ssp. Minor Seedlings Depending on Lighting Conditions. Sci. Rep. 2022, 12, 20259. [Google Scholar] [CrossRef]
- Chhaya, J.T.U. Evaluation of Phytotoxic and Clastogenic Potential of Phenolic Pollutant Bisphenol A and Its Bioremediated Product in Triticum Aestivum, Cicer arietinum and Allium cepa. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 777–790. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Panteris, E.; Eleftheriou, E.P. Tubulin Acetylation Mediates Bisphenol A Effects on the Microtubule Arrays of Allium cepa and Triticum turgidum. Biomolecules 2019, 9, 185. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Zhou, L.; Wang, L.; Zhou, Q.; Huang, X. Analysis of Effects of a New Environmental Pollutant, Bisphenol A, on Antioxidant Systems in Soybean Roots at Different Growth Stages. Sci. Rep. 2016, 6, 23782. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Sperdouli, I.; Eleftheriou, E.P.; Moustakas, M. Hydrogen Peroxide Production by the Spot-like Mode Action of Bisphenol A. Front. Plant Sci. 2020, 11, 1196. [Google Scholar] [CrossRef]
- Fiskesjö, G. The Allium Test as a Standard in Environmental Monitoring. Hereditas 1985, 102, 99–112. [Google Scholar] [CrossRef]
- Leme, D.M.; Marin-Morales, M.A. Allium cepa Test in Environmental Monitoring: A Review on Its Application. Mutat. Res./Rev. Mutat. Res. 2009, 682, 71–81. [Google Scholar] [CrossRef]
- Firbas, P.; Amon, T. Chromosome Damage Studies in the Onion Plant Allium cepa L. Caryologia 2014, 67, 25–35. [Google Scholar] [CrossRef]
- Aksoy, Ö. Detection of Environmental Mutagens Through Plant Bioassays. In Plant Ecology—Traditional Approaches to Recent Trends; Yousaf, Z., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-3339-1. [Google Scholar]
- Mesi, A.D.; Kopliku, D. Cyto- and Genotoxic Activity of Pesticide Cypermex Plus 550 EC on Allium cepa L. J. Int. Environ. Appl. Sci. 2015, 10, 475–481. [Google Scholar]
- Garay-Arroyo, A.; De La Paz Sánchez, M.; García-Ponce, B.; Azpeitia, E.; Álvarez-Buylla, E.R. Hormone Symphony during Root Growth and Development. Dev. Dyn. 2012, 241, 1867–1885. [Google Scholar] [CrossRef]
- Štraser, A.; Filipič, M.; Novak, M.; Žegura, B. Double Strand Breaks and Cell-Cycle Arrest Induced by the Cyanobacterial Toxin Cylindrospermopsin in HepG2 Cells. Mar. Drugs 2013, 11, 3077–3090. [Google Scholar]
- Marnett, L.J. Oxy Radicals, Lipid Peroxidation and DNA Damage. Toxicology 2002, 181, 219–222. [Google Scholar] [CrossRef]
- Marnett, L.J. Lipid Peroxidation—DNA Damage by Malondialdehyde. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1999, 424, 83–95. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- D’Angelo, S.; Scafuro, M.; Meccariello, R. BPA and Nutraceuticals, Simultaneous Effects on Endocrine Functions. EMIDDT 2019, 19, 594–604. [Google Scholar] [CrossRef]
- Zahra, Z.; Khan, M.R.; Majid, M.; Maryam, S.; Sajid, M. Gonadoprotective Ability of Vincetoxicum arnottianum Extract against Bisphenol A-induced Testicular Toxicity and Hormonal Imbalance in Male Sprague Dawley Rats. Andrologia 2020, 52, e13590. [Google Scholar] [CrossRef]
- Rademacher, W. Growth Retardants: Effects on Gibberellin Biosynthesis and Other Metabolic Pathways. Annu. Rev. Plant Biol. 2000, 51, 501–531. [Google Scholar] [CrossRef]
- Fiskesjo, G. Allium Test for Screening Chemicals; Evaluation of Cytological Parameters. Plants Environ. Stud. 1997, 11, 307–333. [Google Scholar]
- Brkanac, S.R.; Gerić, M.; Gajski, G.; Vujčić, V.; Garaj-Vrhovac, V.; Kremer, D.; Domijan, A.-M. Toxicity and Antioxidant Capacity of Frangula Alnus Mill. Bark and Its Active Component Emodin. Regul. Toxicol. Pharmacol. 2015, 73, 923–929. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Levine, R.; Garland, D.; Oliver, C.; Amici, A.; Climent, I. Determination of Carbonyl Content in Oxidatively Modified Proteins. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Howard, L.R.; Clark, J.R.; Brownmiller, C. Antioxidant Capacity and Phenolic Content in Blueberries as Affected by Genotype and Growing Season. J. Sci. Food Agric. 2003, 83, 1238–1247. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPA (mg L−1) | Interphase | Prophase | Metaphase | Anaphase | Telophase |
|---|---|---|---|---|---|
| 0 | 92.48 ± 1.14 cA | 4.32 ± 1.06 aB | 2.00 ± 1.06 aC | 1.35 ± 0.68 aCD | 0.80 ± 0.71 aD |
| 1 | 94.28 ± 0.94 bA | 3.28 ± 0.97 bB | 1.27 ± 0.23 abC | 0.96 ± 0.74 abC | 0.75 ± 0.30 aC |
| 5 | 94.95 ± 2.04 abA | 2.30 ± 0.82 bcB | 1.05 ± 0.91 abBC | 0.93 ± 0.64 abC | 0.60 ± 0.49 aC |
| 10 | 95.32 ± 0.66 abA | 2.76 ± 0.38 bcB | 0.85 ± 0.53 bC | 0.48 ± 0.23 bCD | 0.40 ± 0.24 aD |
| 25 | 95.96 ± 0.84 aA | 2.40 ± 0.40 bcB | 0.80 ± 0.49 bC | 0.52 ± 0.18 bCD | 0.20 ± 0.20 aD |
| 50 | 96.16 ± 0.36 aA | 2.04 ± 0.57 cB | 0.75 ± 0.25 bC | 0.56 ± 0.43 bC | 0.40 ± 0.20 aC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujčić Bok, V.; Gerić, M.; Gajski, G.; Gagić, S.; Domijan, A.-M. Phytotoxicity of Bisphenol A to Allium cepa Root Cells Is Mediated through Growth Hormone Gibberellic Acid and Reactive Oxygen Species. Molecules 2023, 28, 2046. https://doi.org/10.3390/molecules28052046
Vujčić Bok V, Gerić M, Gajski G, Gagić S, Domijan A-M. Phytotoxicity of Bisphenol A to Allium cepa Root Cells Is Mediated through Growth Hormone Gibberellic Acid and Reactive Oxygen Species. Molecules. 2023; 28(5):2046. https://doi.org/10.3390/molecules28052046
Chicago/Turabian StyleVujčić Bok, Valerija, Marko Gerić, Goran Gajski, Sanja Gagić, and Ana-Marija Domijan. 2023. "Phytotoxicity of Bisphenol A to Allium cepa Root Cells Is Mediated through Growth Hormone Gibberellic Acid and Reactive Oxygen Species" Molecules 28, no. 5: 2046. https://doi.org/10.3390/molecules28052046
APA StyleVujčić Bok, V., Gerić, M., Gajski, G., Gagić, S., & Domijan, A.-M. (2023). Phytotoxicity of Bisphenol A to Allium cepa Root Cells Is Mediated through Growth Hormone Gibberellic Acid and Reactive Oxygen Species. Molecules, 28(5), 2046. https://doi.org/10.3390/molecules28052046