

Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk




Abstract
1. Introduction
2. Genetics and Biosynthesis of Milk Caseins
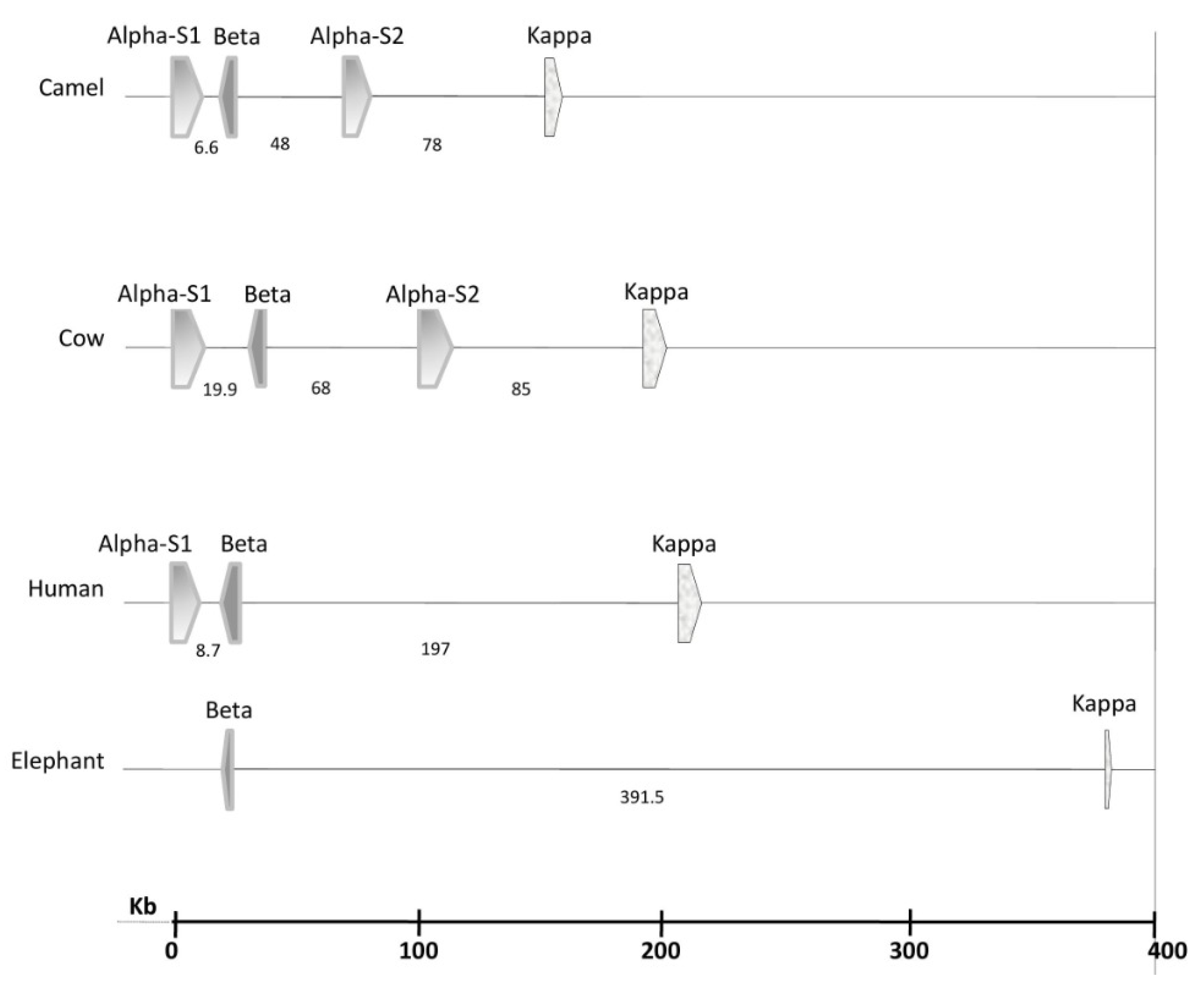
2.1. Genetics
2.2. Biosynthesis of Milk and Milk Caseins
3. Molecular Structures of Milk Caseins
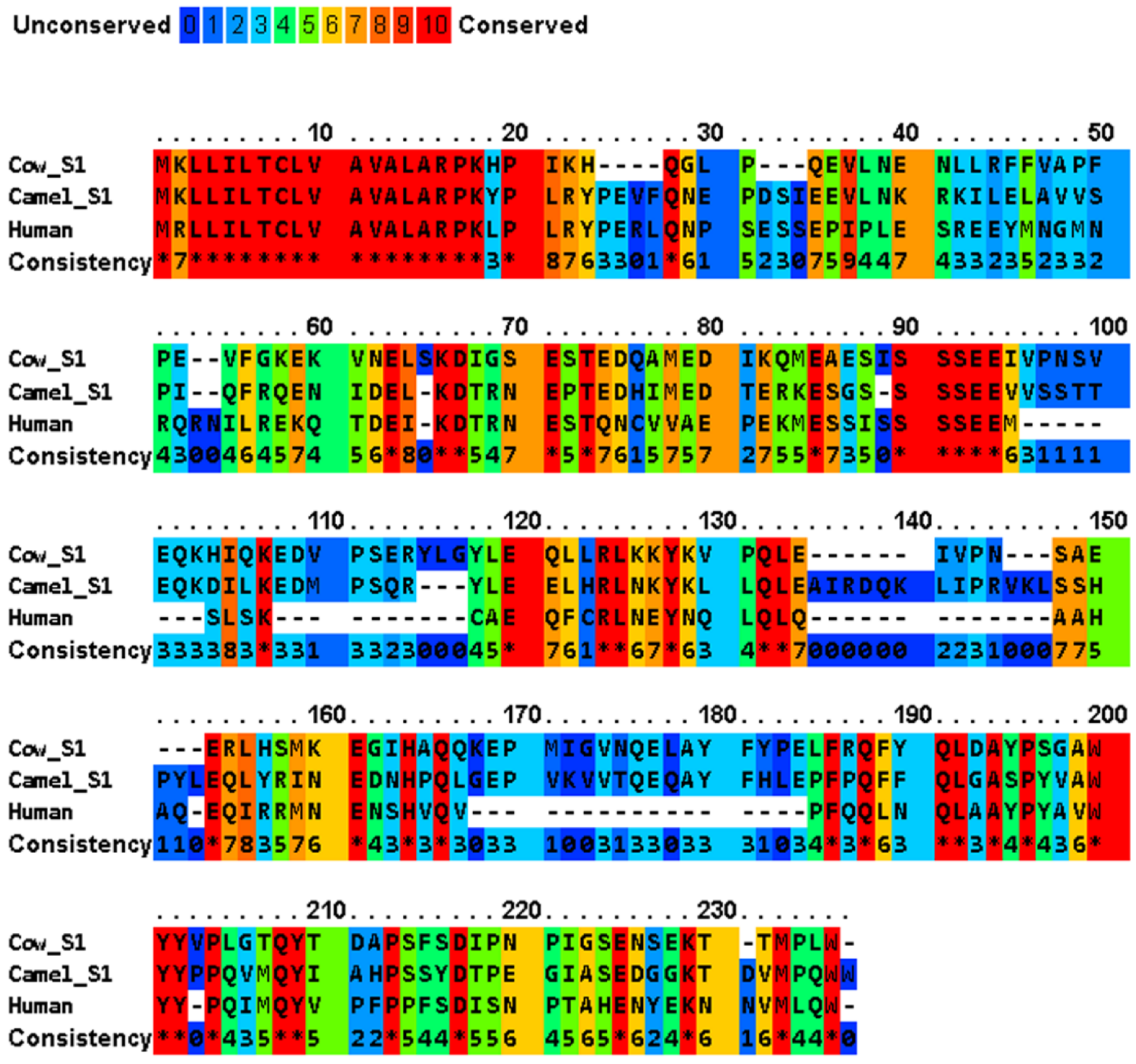
3.1. Amino Acid Sequence Identities, and Instability
3.2. Distribution of Uncharged Polar, Hydrophobic, Aromatic, Isoelectric Points and Polar Amino Acids in Caseins
3.3. Posttranslational Phosphorylation and Glycosylation of Milk Caseins
3.4. Predicted Secondary Structures of Milk Caseins in the Four Animals
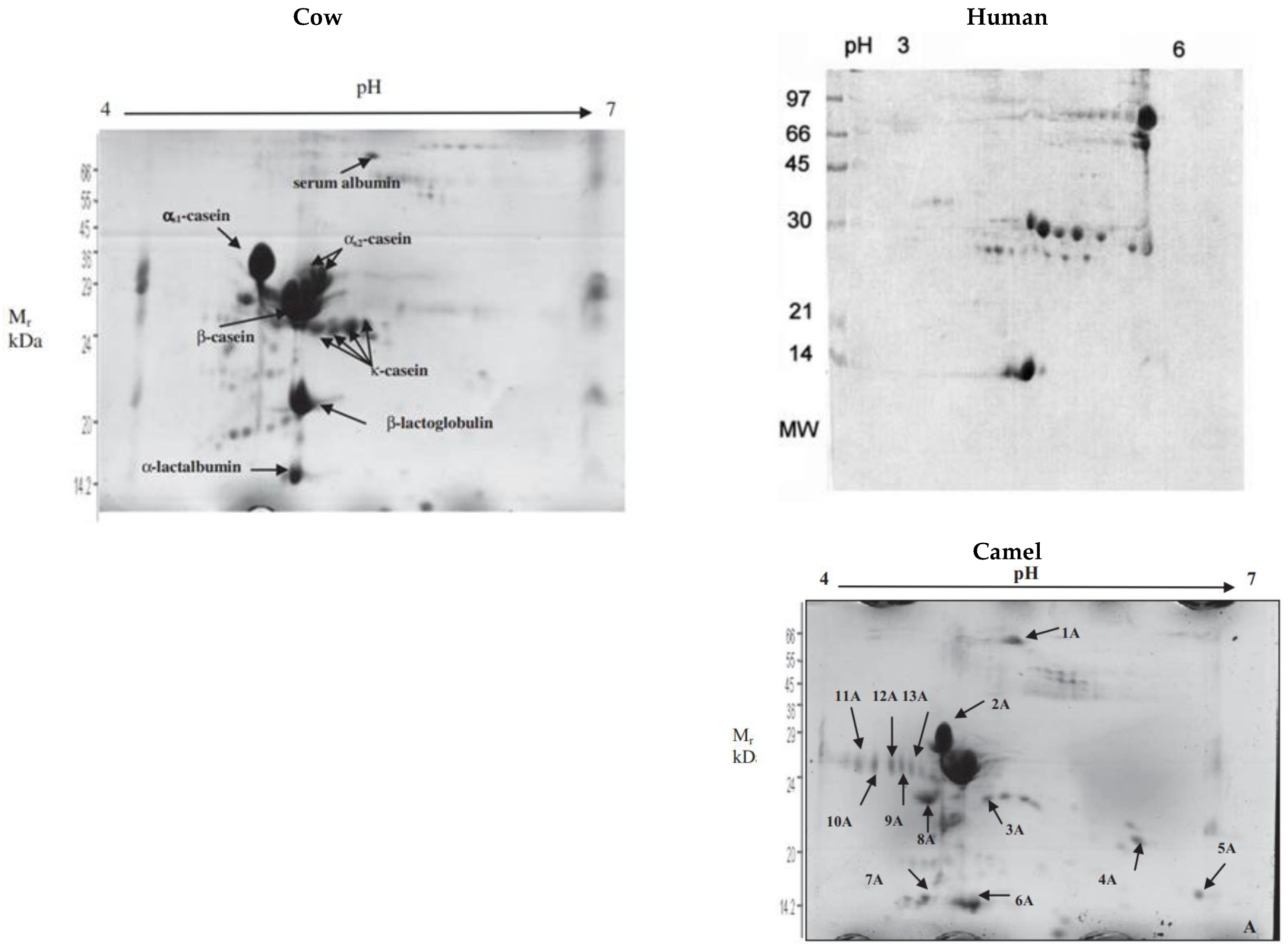
4. Casein Micelle Composition and Structure
5. Functional Properties of Milk Caseins
5.1. Milk Types
5.2. Coagulation Properties
5.3. Ethanol Stability of Milk
5.4. Heat Sensitivity
5.5. Proteolysis Sites and Products
5.6. Chaperone Activities
5.7. Nanoencapsulation Properties
6. Nutritional Properties and Applications
6.1. Diabetes
6.2. Allergy
6.3. Autism
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhat, M.Y.; Dar, T.A.; Singh, L.R. Casein Proteins: Structural and Functional Aspects. In Milk Proteins—From Structure to Biological Properties and Health Aspects; Intech Open: London, UK, 2016; Available online: https://www.intechopen.com/state.item.id (accessed on 20 December 2022).
- Roy, D.; Ye, A.; Moughan, P.J.; Singh, H. Composition, Structure, and Digestive Dynamics of Milk from Different Species—A Review. Front. Nutr. 2020, 7, 195. [Google Scholar] [CrossRef]
- Holland, J.W.; Boland, M.J. Post-translational modifications of caseins. In Milk Proteins, 2nd ed.; Singh, H., Boland, M., Thompson, A., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 141–168. [Google Scholar]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited review: Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Huppertz, T.; Urban, V.S.; Petukhov, A.V. Casein micelles and their internal structure. Adv. Colloid Interface Sci. 2012, 171, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, D.G.; Corredig, M. The structure of the casein micelle of milk and its changes during processing. Annu. Rev. Food Sci. Technol. 2012, 3, 449–467. [Google Scholar] [CrossRef] [PubMed]
- De Kruif, C.G.; Zhulina, E.B. κ-casein as a polyelectrolyte brush on the surface of casein micelles. Colloids Surf. A Physicochem. Eng. Asp. 1996, 117, 151–159. [Google Scholar] [CrossRef]
- Osthoff, G.; de Wit, M.; Hugo, A.; Kamara, B.I. Milk composition of three free-ranging African elephant (Loxodonta africana africana) cows during mid lactation. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 148, 1–5. [Google Scholar] [CrossRef]
- Madende, M.; Osthoff, G.; Patterton, H.G.; Patterton, H.E.; Martin, P.; Opperman, D.J. Characterization of casein and alpha lactalbumin of African elephant (Loxodonta africana) milk. J. Dairy Sci. 2015, 98, 8308–8318. [Google Scholar] [CrossRef]
- Himschoot, E.A.; Wenker, E.S.; Reed, E.G.; Sampson, J.; Power, M.L. Macronutrient composition of milk from two captive African elephant (Loxodonta africana) cows. Zoo Biol. 2021, 40, 192–200. [Google Scholar] [CrossRef]
- Takatsu, Z.; Tsuda, M.; Yamada, A.; Matsumoto, H.; Takai, A.; Takeda, Y.; Takase, M. Elephant’s breast milk contains large amounts of glucosamine. J. Vet. Med. Sci. 2017, 79, 524–533. [Google Scholar] [CrossRef]
- Sadiq, U.; Gill, H.; Chandrapala, J. Casein Micelles as an Emerging Delivery System for Bioactive Food Components. Foods 2021, 10, 1965. [Google Scholar] [CrossRef]
- Horne, D.S. Casein micelle structure: Models and muddles. Curr. Opin. Colloid Interface Sci. 2006, 11, 148–153. [Google Scholar] [CrossRef]
- Glantz, M.; Devold, T.G.; Vegarud, G.E.; Lindmark Månsson, H.; Stålhammar, H.; Paulsson, M. Importance of casein micelle size and milk composition for milk gelation. J. Dairy Sci. 2010, 93, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Casanova, M.; da Silva Pinto, A.F.; de Carvalho, A.F.; Gaucheron, F. Casein micelles: From the monomers to the supramolecular structure. Braz. J. Food Technol. 2019, 22. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Huppertz, T. Casein Micelles: Size distribution in milks from individual cows. J Agric Food Chem. 2012, 60, 4649–4655. [Google Scholar] [CrossRef] [PubMed]
- Pauciullo, A.; Shuiep, E.T.; Ogah, M.D.; Cosenza, G.; Di Stasio, L.; Erhardt, G. Casein gene cluster in camelids: Comparative genome analysis and new findings on haplotype variability and physical mapping. Front. Genet. 2019, 10, 748. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Lafont, A.G.; Sire, J.Y. The Evolution of Milk Casein Genes from Tooth Genes before the Origin of Mammals. Mol. Biol. Evol. 2011, 28, 2053–2061. [Google Scholar] [CrossRef]
- Rijnkels, M. Multispecies comparison of the casein gene loci and evolution of casein gene family. J Mammary Gland. Biol Neoplasia 2002, 7, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediat. Clin. North Am. 2013, 60, 49. [Google Scholar] [CrossRef]
- Walstra, P.; Walstra, P.; Wouters, J.T.M.; Geurts, T.J. Dairy Science and Technology; 2005. Available online: https://www.taylorfrancis.com/books/mono/10.1201/9781420028010/dairy-science-technology-walstra-pieter-walstra-jan-wouters-tom-geurts (accessed on 21 December 2022).
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 2016, 7, 1–22. [Google Scholar] [CrossRef]
- Cai, J.; Wang, D.; Zhao, F.Q.; Liang, S.; Liu, J. AMPK-mTOR pathway is involved in glucose-modulated amino acid sensing and utilization in the mammary glands of lactating goats. J. Anim. Sci. Biotechnol. 2020, 11, 32. [Google Scholar] [CrossRef]
- Gao, H.; Hu, H.; Zheng, N.; Wang, J. Leucine and histidine independently regulate milk protein synthesis in bovine mammary epithelial cells via mTOR signaling pathway. J. Zhejiang Univ. Sci. B 2015, 16, 560. [Google Scholar] [CrossRef]
- Wyszomierski, S.L.; Rosen, J.M. Cooperative effects of STAT5 (signal transducer and activator of transcription 5) and C/EBPbeta (CCAAT/enhancer-binding protein-beta) on beta-casein gene transcription are mediated by the glucocorticoid receptor. Mol. Endocrinol. 2001, 15, 228–240. [Google Scholar] [PubMed]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Chemistry of Milk Constituents. In Fundamentals of Cheese Science; Springer: Berlin/Heidelberg, German, 2017; pp. 71–104. Available online: https://link.springer.com/chapter/10.1007/978-1-4899-7681-9_4 (accessed on 21 December 2022).
- De Castro, R.J.S.; Domingues, M.A.F.; Ohara, A.; Okuro, P.K.; dos Santos, J.G.; Brexó, R.P.; Sato, H.H. Whey protein as a key component in food systems: Physicochemical properties, production technologies and applications. Food Struct. 2017, 14, 17–29. [Google Scholar] [CrossRef]
- Farrell, H.M.; Malin, E.L.; Brown, E.M.; Qi, P.X. Casein micelle structure: What can be learned from milk synthesis and structural biology? Curr. Opin. Colloid Interface Sci. 2006, 11, 135–147. [Google Scholar] [CrossRef]
- Shekar, P.C.; Goel, S.; Rani, S.D.S.; Sarathi, D.P.; Alex, J.L.; Singh, S.; Kumar, S. κ-Casein-deficient mice fail to lactate. Proc. Natl. Acad. Sci. USA 2006, 103, 8000. [Google Scholar] [CrossRef] [PubMed]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited review: Milk protein polymorphisms in cattle: Effect on animal breeding and human nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef]
- Manguy, J.; Shields, D.C. Implications of kappa-casein evolutionary diversity for the self-assembly and aggregation of casein micelles. R. Soc. Open Sci. 2019, 6, 190939. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Mahomud, M.S.; Haque, M.E. Heat-Induced Interaction of Milk Proteins: Impact on Yoghurt Structure. Int. J. Food Sci. 2021, 2021, 5569917. [Google Scholar] [CrossRef]
- Meier, S.; Korkuć, P.; Arends, D.; Brockmann, G.A. DNA Sequence Variants and Protein Haplotypes of Casein Genes in German Black Pied Cattle (DSN). Front. Genet. 2019, 10, 1129. [Google Scholar] [CrossRef]
- Lipman, D.J.; Souvorov, A.; Koonin, E.v.; Panchenko, A.R.; Tatusova, T.A. The relationship of protein conservation and sequence length. BMC Evol. Biol. 2023, 2, 1–10. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 2022, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Rokas, A.; Carroll, S.B. Frequent and widespread parallel evolution of protein sequences. Mol. Biol. Evol. 2023, 25, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Naylor, G.J.P.; Collins, T.M.; Brown, W.M. Hydrophobicity and phylogeny. Nature 1995, 373, 565–566. [Google Scholar] [CrossRef] [PubMed]
- Wölk, M.; Milkovska-Stamenova, S.; Hoffmann, R. Comprehensive Profiling of the Native and Modified Peptidomes of Raw Bovine Milk and Processed Milk Products. Foods 2020, 9, 1841. [Google Scholar] [CrossRef]
- Holland, J.W. Post-Translational Modifications of Caseins and whey. In Milk Proteins; Academic Press: Cambridge, MA, USA, 2008; pp. 107–132. [Google Scholar]
- Iukalo, A.V. New Approach for Isolation of Individual Caseins from Cow Milk by the Preparative Electrophoresis. Adv. Biol. Chem. 2014, 4, 382–387. [Google Scholar] [CrossRef]
- Baranyi, M.; Hiripi, L.; Szabó, L.; Catunda, A.P.; Harsányi, I.; Komáromy, P.; Bősze, Z. Isolation and some effects of functional, low-phenylalanine kappa-casein expressed in the milk of transgenic rabbits. J. Biotechnol. 2007, 128, 383–392. [Google Scholar] [CrossRef]
- Bahraminejad, E.; Paliwal, D.; Sunde, M.; Holt, C.; Carver, J.A.; Thorn, D.C. Amyloid fibril formation by αS1- and β-casein implies that fibril formation is a general property of casein proteins. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2022, 1870, 140854. [Google Scholar] [CrossRef]
- Holt, C.; Raynes, J.K.; Carver, J.A. Sequence characteristics responsible for protein-protein interactions in the intrinsically disordered regions of caseins, amelogenins, and small heat-shock proteins. Biopolymer 2019, 110, e23319. [Google Scholar] [CrossRef]
- Thorn, D.C.; Ecroyd, H.; Carver, J.A.; Holt, C.; Thorn, D.C. Casein structures in the context of unfolded proteins. Int. Dairy J. 2015, 46, 2–11. [Google Scholar] [CrossRef]
- Treweek, T.M.; Thorn, D.C.; Price, W.E.; Carver, J.A. The chaperone action of bovine milk αS1- and αS2-caseins and their associated form αS-casein. Arch. Biochem. Biophys. 2011, 510, 42–52. [Google Scholar] [CrossRef]
- Audain, E.; Ramos, Y.; Hermjakob, H.; Flower, D.R.; Perez-Riverol, Y. Accurate estimation of isoelectric point of protein and peptide based on amino acid sequences. Bioinform. 2016, 32, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Tokmakov, A.A.; Kurotani, A.; Sato, K.I. Protein pI and Intracellular Localization. Front. Mol. Biosci. 2021, 8, 1179. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, L.; Miotto, M.; Bò, L.; Ruocco, G.; Raimondo, D.; Milanetti, E. Characterizing Hydropathy of Amino Acid Side Chain in a Protein Environment by Investigating the Structural Changes of Water Molecules Network. Front. Mol. Biosci. 2021, 8, 626837. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cai, L.; Liao, B.; Fu, X.; Bing, P.; Yang, J. Prediction of Protein Subcellular Localization Based on Fusion of Multi-view Features. Molecules 2019, 24, 919. [Google Scholar] [CrossRef] [PubMed]
- Pánek, J.; Eidhammer, I.; Aasland, R. Using hydropathy features for function prediction of membrane proteins. Mol. Membr. Biol. 2009, 24, 304–312. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Rice, P.; Longden, L.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Ellouze, M.; Vial, C.; Attia, H.; Ayadi, M.A. Effect of pH and heat treatment on structure, surface characteristics and emulsifying properties of purified camel β-casein. Food Chem. 2021, 365, 130421. [Google Scholar] [CrossRef]
- Atamer, Z.; Post, A.E.; Schubert, T.; Holder, A.; Boom, R.M.; Hinrichs, J. Bovine β-casein: Isolation, properties and functionality. A review. Int. Dairy J. 2017, 66, 115–125. [Google Scholar] [CrossRef]
- Esmaili, M.; Ghaffari, S.M.; Moosavi-Movahedi, Z.; Atri, M.S.; Sharifizadeh, A.; Farhadi, M.; Yousefi, R.; Chobert, J.-M.; Haertlé, T.; Moosavi-Movahedi, A.A. Beta casein-micelle as a nano vehicle for solubility enhancement of curcumin; food industry application. Lebensm.-Wiss. Technol. 2011, 44, 2166–2172. [Google Scholar] [CrossRef]
- Li, M.; Auty, M.A.E.; Crowley, S.V.; Kelly, A.L.; O’Mahony, J.A.; Brodkorb, A. Self-association of bovine β-casein as influenced by calcium chloride, buffer type and temperature. Food Hydrocoll. 2019, 88, 190–198. [Google Scholar] [CrossRef]
- Nick Pace, C.; Martin Scholtz, J.; Grimsley, G.R. Forces stabilizing proteins. FEBS Lett. 2014, 588, 2177–2184. [Google Scholar] [CrossRef]
- Gamage, D.G.; Gunaratne, A.; Periyannan, G.R.; Russell, T.G. Applicability of Instability Index for In vitro Protein Stability Prediction. Protein Pept. Lett. 2019, 26, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Springer: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Guruprasad, K.; Reddy, B.V.B.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Lucey, J.A.; Otter, D.; Horne, D.S. A 100-Year Review: Progress on the chemistry of milk and its components. J. Dairy Sci. 2017, 100, 9916–9932. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.H.; Visker, M.H.P.W.; Miranda, G.; Delacroix-Buchet, A.; Bovenhuis, H.; Martin, P. The relationships among bovine αS-casein phosphorylation isoforms suggest different phosphorylation pathways. J. Dairy Sci. 2016, 99, 8168–8177. [Google Scholar] [CrossRef]
- Hinz, K.; O’Connor, P.M.; Huppertz, T.; Ross, R.P.; Kelly, A.L. Comparison of the principal proteins in bovine, caprine, buffalo, equine and camel milk. J. Dairy Res. 2012, 79, 185–191. [Google Scholar] [CrossRef]
- Ryskaliyeva, A.; Henry, C.; Miranda, G.; Faye, B.; Konuspayeva, G.; Martin, P. Combining different proteomic approaches to resolve complexity of the milk protein fraction of dromedary, Bactrian camels and hybrids, from different regions of Kazakhstan. PLoS ONE. 2018, 13, e0197026. [Google Scholar] [CrossRef]
- Bertino, E.; Gastaldi, D.; Monti, G.; Baro, C.; Fortunato, D.; Perono, G.L.; Coscia, A.; Fabris, C.; Mussap, M.; Conti, A. Detailed proteomic analysis on DM: Insight into its hypoallergenicity. Front. Biosci. 2010, 1, 526–536. [Google Scholar] [CrossRef]
- Dingess, K.; Gazi, I.; Toorn, H.V.D.; Mank, M.; Stahl, B.; Reiding, K.; Heck, A. Monitoring human milk β-casein phosphorylation and o-glycosylation over lactation reveals distinct differences between the proteome and endogenous peptidome. Int. J. Mol. Sci. 2021, 22, 8140. [Google Scholar] [CrossRef]
- Sørensen, E.S.; Møller, L.; Vinther, M.; Petersen, T.E.; Rasmussen, L.K. The phosphorylation pattern of human αs1-casein is markedly different from the ruminant species. Eur. J. Biochem. 2003, 270, 3651–3655. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.; Nwosu, C.C.; Strum, J.S.; Seipert, R.R.; An, H.J.; Zivkovic, A.M.; German, J.B.; Lebrilla, C.B. Site-specific protein glycosylation analysis with glycan isomer differentiation. Anal. Bioanal. Chem. 2012, 403, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Kim, B.J.; Koh, J.; Dallas, D.C. Analysis of Bovine Kappa-Casein Glycomacropeptide by Liquid Chromatography-Tandem Mass Spectrometry. Foods 2021, 10, 2028. [Google Scholar] [CrossRef]
- Minervini, F.; Algaron, F.; Rizzello, C.G.; Fox, P.F.; Monnet, V.; Gobbetti, M. Angiotensin I-Converting-Enzyme-Inhibitory and Antibacterial Peptides from Lactobacillus helveticus PR4 Proteinase-Hydrolyzed Caseins of Milk from Six Species. Appl. Environ. Microbiol. 2003, 69, 5297. [Google Scholar] [CrossRef]
- Goonatilleke, E.; Huang, J.; Xu, G.; Wu, L.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Human Milk Proteins and Their Glycosylation Exhibit Quantitative Dynamic Variations during Lactation. J. Nutr. 2019, 149, 1317. [Google Scholar] [CrossRef]
- Swelum, A.A.; El-Saadony, M.T.; Abdo, M.; Ombarak, R.A.; Hussein, E.O.; Suliman, G.; Alhimaidi, A.R.; Ammari, A.A.; Ba-Awadh, H.; Taha, A.E.; et al. Nutritional, antimicrobial and medicinal properties of Camel’s milk: A review. Saudi J. Biol. Sci. 2021, 28, 3126. [Google Scholar] [CrossRef]
- Leonhard, K.; Prausnitz, J.M.; Radke, C.J. Solvent–amino acid interaction energies in three-dimensional-lattice Monte Carlo simulations of a model 27-mer protein: Folding thermodynamics and kinetics. Protein Sci. 2004, 13, 358. [Google Scholar] [CrossRef] [PubMed]
- Khoury, G.A.; Smadbeck, J.; Kieslich, C.A.; Floudas, C.A. Protein folding and de novo protein design for biotechnological applications. Trends Biotechnol. 2014, 32, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Corredig, M.; Nair, P.K.; Li, Y.; Eshpari, H.; Zhao, Z. Invited review: Understanding the behavior of caseins in milk concentrates. J. Dairy Sci. 2019, 102, 4772–4782. [Google Scholar] [CrossRef]
- Martin, P.; Cebo, C.; Miranda, G. Interspecies comparison of milk proteins: Quantitative variability and molecular diversity. In Advanced Dairy Chemistry: Proteins: Basic Aspects, 4th ed.; Springer: Boston, MA, USA, 2013; Volume 1A, pp. 387–429. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Głąb, T.K.; Boratyński, J. Potential of Casein as a Carrier for Biologically Active Agents. Top. Curr. Chem. 2017, 375, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Broyard, C.; Gaucheron, F. Modifications of structures and functions of caseins: A scientific and technological challenge. Dairy Sci. Technol. 2015, 95, 831–862. [Google Scholar] [CrossRef]
- McMahon, D.J.; Oommen, B.S. Supramolecular Structure of the Casein Micelle. J. Dairy Sci. 2008, 91, 1709–1721. [Google Scholar] [CrossRef] [PubMed]
- Hettiarachchi, C.A.; Swulius, M.T.; Harte, F.M. Assessing constituent volumes and morphology of bovine casein micelles using cryo-electron tomography. J. Dairy Sci. 2020, 103, 3971–3979. [Google Scholar] [CrossRef]
- Hindmarsh, J.P.; Watkinson, P. Experimental evidence for previously unclassified calcium phosphate structures in the casein micelle. J. Dairy Sci. 2017, 100, 6938–6948. [Google Scholar] [CrossRef] [PubMed]
- Salvador, D.; Acosta, Y.; Zamora, A.; Castillo, M. Rennet-Induced Casein Micelle Aggregation Models: A Review. Foods 2022, 11, 1243. [Google Scholar] [CrossRef]
- Zhao, Z. Colloidal and Rennet Coagulation Properties of Concentrated Casein Micelles with Altered Mineral Equilibrium. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2015. [Google Scholar]
- Dalgleish, D.G. On the structural models of bovine casein micelles—Review and possible improvements. Soft Matter. 2011, 7, 2265–2272. [Google Scholar] [CrossRef]
- Qi, P.X. Studies of casein micelle structure: The past and the present. Lait 2007, 87, 363–383. [Google Scholar] [CrossRef]
- Holt, C. Casein and casein micelle structures, functions and diversity in 20 species. Int. Dairy J. 2016, 60, 2–13. [Google Scholar] [CrossRef]
- Horne, D.S. Casein micelle structure and stability. In Milk Proteins; Academic Press: Cambridge, MA, USA, 2020; pp. 213–250. [Google Scholar]
- Bonfatti, V.; Chiarot, G.; Carnier, P. Glycosylation of κ-casein: Genetic and nongenetic variation and effects on rennet coagulation properties of milk. J. Dairy Sci. 2014, 97, 1961–1969. [Google Scholar] [CrossRef]
- Bijl, E.; de Vries, R.; van Valenberg, H.; Huppertz, T.; van Hooijdonk, T. Factors influencing casein micelle size in milk of individual cows: Genetic variants and glycosylation of κ-casein. Int. Dairy J. 2014, 34, 135–141. [Google Scholar] [CrossRef]
- Le Parc, A.; Leonil, J.; Chanat, E. αS1-casein, which is essential for efficient ER-to-Golgi casein transport, is also present in a tightly membrane-associated form. BMC Cell Biol. 2010, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Kolb, A.F.; Huber, R.C.; Lillico, S.G.; Carlisle, A.; Robinson, C.J.; Neil, C.; Petrie, L.; Sorensen, D.B.; Olsson, I.A.S.; Whitelaw, C.B.A. Milk Lacking α-Casein Leads to Permanent Reduction in Body Size in Mice. PLoS ONE 2011, 6, 21775. [Google Scholar] [CrossRef] [PubMed]
- Persuy, M.A.; Printz, C.; Medrano, J.F.; Mercier, J.C. A single nucleotide deletion resulting in a premature stop codon is associated with marked reduction of transcripts from a goat beta-casein null allele. Anim. Genet. 1999, 30, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, T.; Hennebel, J.B.; Considine, T.; Shakeel-Ur-Rehman Kelly, A.L.; Fox, P.F. A method for the large-scale isolation of β-casein. Food Chem. 2006, 99, 45–50. [Google Scholar] [CrossRef]
- Yahimi Yazdi, S.; Corredig, M.; Dalgleish, D.G. Studying the structure of β-casein-depleted bovine casein micelles using electron microscopy and fluorescent polyphenols. Food Hydrocoll. 2014, 42, 171–177. [Google Scholar] [CrossRef]
- Sanders, H.M.; Jovcevski, B.; Carver, J.A.; Pukala, T.L. The molecular chaperone β-casein prevents amorphous and fibrillar aggregation of α-lactalbumin by stabilisation of dynamic disorder. Biochem. J. 2020, 477, 629. [Google Scholar] [CrossRef] [PubMed]
- Phadungath, C. Casein micelle structure: A concise review. Songklanakarin J. Sci. Technol. (SJST) 2005, 27, 201–212. [Google Scholar]
- Attia, H.; Kherouatou, N.; Nasri, M.; Khorchani, T. Characterization of the dromedary milk casein micelle and study of its changes during acidification. Lait 2000, 80, 503–515. [Google Scholar] [CrossRef]
- Kamal, M.; Foukani, M.; Karoui, R. Rheological and physical properties of camel and cow milk gels enriched with phosphate and calcium during acid-induced gelation. J. Food Sci. Technology. 2017, 54, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, M.; Mukesh, M.; Kataria, R.S.; Mishra, B.P.; Joshii, B.K. Milk proteins and human health: A1/A2 milk hypothesis. Indian J. Endocrinol. Metab. 2012, 16, 856. [Google Scholar] [CrossRef]
- Kaplan, M.; Baydemir, B.; Günar, B.B.; Arslan, A.; Duman, H.; Karav, S. Benefits of A2 Milk for Sports Nutrition, Health and Performance. Front. Nutr. 2022, 9, 1500. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, M.; Lamberti, C.; Cirrincione, S.; Giuffrida, M.G.; Cavallarin, L. A2 milk and BCM-7 peptide as emerging parameters of milk quality. Front. Nutrition. 2022, 9, 65. [Google Scholar] [CrossRef]
- Lucey, J.A. Acid coagulation of milk. In Advanced Dairy Chemistry; Springer: New York, NY, USA, 2016; pp. 309–328. [Google Scholar]
- Mbye, M.; Sobti, B.; Al Nuami, M.K.; Al Shamsi, Y.; Al Khateri, L.; Al Saedi, R.; Saeed, M.; Ramachandran, T.; Hamed, F.; Kamal-Eldin, A. Physicochemical properties, sensory quality, and coagulation behavior of camel versus bovine milk soft unripened cheeses. NFS J. 2020, 20, 28–36. [Google Scholar] [CrossRef]
- Hunter, T. Why nature chose phosphate to modify proteins. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2513–2516. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Johansson, M.; Lundh, Å.; Nagy, P.; Kamal-Eldin, A. Short communication: Caseins and α-lactalbumin content of camel milk (Camelus dromedarius) determined by capillary electrophoresis. J. Dairy Sci. 2020, 103, 11094–11099. [Google Scholar] [CrossRef]
- Huppertz, T.; Chia, L.W. Milk protein coagulation under gastric conditions: A review. Int. Dairy J. 2021, 113, 104882. [Google Scholar] [CrossRef]
- Dallas, D.C.; Guerrero, A.; Khaldi, N.; Borghese, R.; Bhandari, A.; Underwood, M.A.; Lebrilla, C.B.; German, J.B.; Barile, D. A Peptidomic Analysis of Human Milk Digestion in the Infant Stomach Reveals Protein-Specific Degradation Patterns. J. Nutr. 2014, 144, 815. [Google Scholar] [CrossRef]
- Zou, Z.; Duley, J.A.; Cowley, D.M.; Reed, S.; Arachchige, B.J.; Koorts, P.; Shaw, P.N.; Bansal, N. Digestibility of proteins in camel milk in comparison to bovine and human milk using an in vitro infant gastrointestinal digestion system. Food Chem. 2022, 374, 131704. [Google Scholar] [CrossRef]
- De Oliveira, S.C.; Bellanger, A.; Ménard, O.; Pladys, P.; Le Gouar, Y.; Dirson, E.; Kroell, F.; Dupont, D.; Deglaire, A.; Bourlieu, C. Impact of human milk pasteurization on gastric digestion in preterm infants: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 379–390. [Google Scholar] [CrossRef]
- Ye, A.; Cui, J.; Dalgleish, D.; Singh, H. Formation of a structured clot during the gastric digestion of milk: Impact on the rate of protein hydrolysis. Food Hydrocoll. 2016, 52, 478–486. [Google Scholar] [CrossRef]
- Moschopoulou, E. Characteristics of rennet and other enzymes from small ruminants used in cheese production. Small Rumin. Res. 2011, 101, 188–195. [Google Scholar] [CrossRef]
- Mbye, M.; Ayyash, M.; Abu-Jdayil, B.; Kamal-Eldin, A. The Texture of Camel Milk Cheese: Effects of Milk Composition, Coagulants, and Processing Conditions. Front. Nutr. 2022, 9, 868320. [Google Scholar] [CrossRef] [PubMed]
- De la Vara, J.Á.; Berruga, M.I.; Cappelli, J.; Landete-Castillejos, T.; Carmona, M.; Gallego, L.; Molina, A. Some aspects of the ethanol stability of red deer milk (Cervus elaphus hispanicus): A comparison with other dairy species. Int. Dairy J. 2018, 86, 103–109. [Google Scholar] [CrossRef]
- Alhaj, O.A.; Lajnaf, R.; Jrad, Z.; Alshuniaber, M.A.; Jahrami, H.A.; Serag El-Din, M.F. Comparison of Ethanol Stability and Chemical Composition of Camel Milk from Five Samples. Animals 2022, 12, 615. [Google Scholar] [CrossRef]
- Mohamed, H.; Ayyash, M.; Kamal-Eldin, A. Effect of heat treatments on camel milk proteins—A review. Int. Dairy J. 2022, 133, 105404. [Google Scholar] [CrossRef]
- Ho, T.M.; Zou, Z.; Bansal, N. Camel milk: A review of its nutritional value, heat stability, and potential food products. Food Res. Int. 2022, 153, 110870. [Google Scholar] [CrossRef] [PubMed]
- Farah, Z.; Atkins, D. Heat coagulation of camel milk. J. Dairy Res. 2017, 59, 229–231. [Google Scholar] [CrossRef]
- Alhaj, A.O.; Metwalli, A.; Ismail, E. Heat Stability of Camel Milk Proteins After Sterilisation Process. J. Camel Pract. Res. 2011, 18, 277–282. [Google Scholar]
- Dallas, D.C.; Murray, N.M.; Gan, J. Proteolytic Systems in Milk: Perspectives on the Evolutionary Function within the Mammary Gland and the Infant. J. Mammary Gland. Biol. Neoplasia 2015, 20, 133. [Google Scholar] [CrossRef]
- Ismail, B.; Choi, L.H.; Were, L.M.; Nielsen, S.S. Activity and nature of plasminogen activators associated with the casein micelle. J. Dairy Sci. 2006, 89, 3285–3295. [Google Scholar] [CrossRef] [PubMed]
- Ismail, B.; Nielsen, S.S. Invited review: Plasmin protease in milk: Current knowledge and relevance to dairy industry. J. Dairy Sci. 2010, 93, 4999–5009. [Google Scholar] [CrossRef] [PubMed]
- Crudden, A.; Patrick Fox, F.; Kelly, A.L. Factors affecting the hydrolytic action of plasmin in milk. Int. Dairy J. 2005, 15, 305–313. [Google Scholar] [CrossRef]
- Gaiaschi, A.; Beretta, B.; Poiesi, C.; Conti, A.; Giuffrida, M.; Galli, C.; Restani, P. Proteolysis of β-Casein as a Marker of Grana Padano Cheese Ripening. J. Dairy Sci. 2001, 84, 60–65. [Google Scholar] [CrossRef]
- Pihlanto-Leppala, A.; Pahkala, E.; Antila, V. Hydrolysis of κ-casein in solution by chymosin, plasmin, trypsin and Lactobacillus -proteinases. Agric. Food Sci. 1993, 2, 489–496. [Google Scholar] [CrossRef]
- Le Bars, D.; Gripon, J. Hydrolysis of αs1-casein by bovine plasmin Hydrolysis of uS1-casein by bovine plasmin. Lait 1993, 73, 337–344. [Google Scholar] [CrossRef]
- Roep, B.O.; Thomaidou, S.; van Tienhoven, R.; Zaldumbide, A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat. Rev. Endocrinol. 2021, 17, 150. [Google Scholar] [CrossRef]
- Sedaghati, M.; Ezzatpanah, H.; Boojar, M.M.A.; Ebrahimi, M.T.; Aminafshar, M. Plasmin digest of κ-casein as a source of antibacterial peptides. J. Dairy Res. 2014, 81, 245–251. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Salvesen, G. Handbook of proteolytic enzymes. Academic press: Cambridge, MA, USA, 2013; pp. 1–3. [Google Scholar]
- Bell, S.J.; Grochoski, G.T.; Clarke, A.J. Health implications of milk containing beta-casein with the A2 genetic variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Kamiński, S.; Cieślińska, A.; Kostyra, E. Polymorphism of bovine beta-casein and its potential effect on human health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef]
- Gobbetti, M.; Stepaniak, L.; de Angelis, M.; Corsetti, A.; di Cagno, R. Latent bioactive peptides in milk proteins: Proteolytic activation and significance in dairy processing. Crit. Rev. Food Sci. Nutr. 2002, 42, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Jinsmaa, Y.; Yoshikawa, M. Enzymatic release of neocasomorphin and beta-casomorphin from bovine beta-casein. Peptides 1999, 20, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Swaisgood, H.E. Review and update of casein chemistry. J. Dairy Sci. 1993, 76, 3054–3061. [Google Scholar] [CrossRef] [PubMed]
- Cade, R.; Privette, M.; Fregly, M.; Rowland, N.; Sun, Z.; Zele, V.; Wagemaker, H.; Edelstein, C. Autism and schizophrenia: Intestinal disorders. Nutr. Neurosci. 2000, 3, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, K.L.; Knivsberg, A.M. Can the pathophysiology of autism be explained by the nature of the discovered urine peptides? Nutr. Neurosci. 2003, 6, 19–28. [Google Scholar] [CrossRef]
- Lamb, M.M.; Miller, M.; Seifert, J.A.; Frederiksen, B.; Kroehl, M.; Rewers, M.; Norris, J.M. The Effect of Childhood Cow’s Milk Intake and HLA-DR Genotype on Risk of Islet Autoimmunity and Type 1 Diabetes: The Diabetes Autoimmunity Study in the Young (DAISY). Pediatr. Diabetes 2015, 16, 31. [Google Scholar] [CrossRef]
- Gottlieb, S. Early exposure to cows’ milk raises risk of diabetes in high risk children. BMJ Br. Med. J. 2000, 321, 1040. [Google Scholar]
- Nguyen, D.D.; Johnson, S.K.; Busetti, F.; Solah, V.A. Formation and Degradation of Beta-casomorphins in Dairy Processing. Crit. Rev. Food Sci. Nutr. 2015, 55, 1955. [Google Scholar] [CrossRef]
- Madureira, A.R.; Tavares, T.; Gomes, A.M.P.; Pintado, M.E.; Malcata, F.X. Invited review: Physiological properties of bioactive peptides obtained from whey proteins. J. Dairy Sci. 2010, 93, 437–455. [Google Scholar] [CrossRef]
- Silva, S.V.; Malcata, F.X. Caseins as source of bioactive peptides. Int Dairy J. 2005, 15, 1–15. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Shamsia, S.; Conte, A. Release of angiotensin converting enzyme-inhibitory peptides during in vitro gastro-intestinal digestion of camel milk. Int. Dairy J. 2016, 56, 119–128. [Google Scholar] [CrossRef]
- Mudgil, P.; Kamal, H.; Yuen, G.C.; Maqsood, S. Characterization and identification of novel antidiabetic and anti-obesity peptides from camel milk protein hydrolysates. Food Chem. 2018, 259, 46–54. [Google Scholar] [CrossRef]
- Thorn, D.C.; Ecroyd, H.; Sunde, M.; Poon, S.; Carver, J.A. Amyloid fibril formation by bovine milk alpha s2-casein occurs under physiological conditions yet is prevented by its natural counterpart, alpha s1-casein. Biochemistry 2022, 47, 3926–3936. [Google Scholar] [CrossRef] [PubMed]
- Ecroyd, H.; Thorn, D.C.; Liu, Y.; Carver, J.A. The dissociated form of kappa-casein is the precursor to its amyloid fibril formation. Biochem. J. 2010, 429, 251–260. [Google Scholar] [CrossRef]
- Wada, Y.; Loennerdal, B. Bioactive peptides derived from human milk proteins—Mechanisms of action. J. Nutr. biochemistry. 2014, 25, 503–514. [Google Scholar] [CrossRef]
- Koudelka, T.; Dehle, F.C.; Musgrave, I.F.; Hoffmann, P.; Carver, J.A. Methionine oxidation enhances κ-casein amyloid fibril formation. J. Agric. Food Chem. 2012, 60, 4144–4155. [Google Scholar] [CrossRef] [PubMed]
- Gai, N.; Uniacke-lowe, T.; O’regan, J.; Faulkner, H.; Kelly, A.L. Effect of Protein Genotypes on Physicochemical Properties and Protein Functionality of Bovine Milk: A Review. Foods 2021, 10, 2409. [Google Scholar] [CrossRef]
- Zhang, X.; Fu, X.; Zhang, H.; Liu, C.; Jiao, W.; Chang, Z. Chaperone-like activity of beta-casein. Int. J. Biochem. Cell Biol. 2005, 37, 1232–1240. [Google Scholar] [CrossRef]
- Koudelka, T.; Hoffmann, P.; Carver, J.A. Dephosphorylation of alpha(s)- and beta-caseins and its effect on chaperone activity: A structural and functional investigation. J. Agric. Food Chem. 2009, 57, 5956–5964. [Google Scholar] [CrossRef]
- Ghayour, N.; Hosseini, S.M.H.; Eskandari, M.H.; Esteghlal, S.; Nekoei, A.-R.; Gahruie, H.H.; Tatar, M.; Naghibalhossaini, F. Nanoencapsulation of quercetin and curcumin in casein-based delivery systems. Food Hydrocoll. 2019, 87, 394–403. [Google Scholar] [CrossRef]
- Zimet, P.; Rosenberg, D.; Livney, Y.D. Re-assembled casein micelles and casein nanoparticles as nano-vehicles for ω-3 polyunsaturated fatty acids. Food Hydrocoll. 2011, 25, 1270–1276. [Google Scholar] [CrossRef]
- Menéndez-Aguirre, O.; Kessler, A.; Stuetz, W.; Grune, T.; Weiss, J.; Hinrichs, J. Increased loading of vitamin D2 in reassembled casein micelles with temperature-modulated high pressure treatment. Food Res. Int. 2014, 64, 74–80. [Google Scholar] [CrossRef]
- Jarunglumlert, T.; Nakagawa, K.; Adachi, S. Influence of aggregate structure of casein on the encapsulation efficiency of β-carotene entrapped via hydrophobic interaction. Food Struct. 2015, 5, 42–50. [Google Scholar] [CrossRef]
- Malekhosseini, P.; Alami, M.; Khomeiri, M.; Esteghlal, S.; Nekoei, A.R.; Hosseini, S.M.H. Development of casein-based nanoencapsulation systems for delivery of epigallocatechin gallate and folic acid. Food Sci. Nutr. 2019, 7, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Shapira, A.; Assaraf, Y.G.; Livney, Y.D. Beta-casein nanovehicles for oral delivery of chemotherapeutic drugs. Nanomedicine 2010, 6, 119–126. [Google Scholar] [CrossRef]
- Ketto, I.A.; Knutsen, T.M.; Øyaas, J.; Heringstad, B.; Ådnøy, T.; Devold, T.G.; Skeie, S.B. Effects of milk protein polymorphism and composition, casein micelle size and salt distribution on the milk coagulation properties in Norwegian Red cattle. Int. Dairy J. 2017, 70, 55–64. [Google Scholar] [CrossRef]
- Semwal, R.; Joshi, S.K.; Semwal, R.B.; Sodhi, M.; Upadhyaya, K.; Semwal, D.K. Effects of A1 and A2 variants of β-casein on human health—Is β-casomorphin-7 really a harmful peptide in cow milk? Nutrire 2022, 47, 8. [Google Scholar] [CrossRef]
- Cieślińska, A.; Fiedorowicz, E.; Rozmus, D.; Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Kamiński, S. Does a Little Difference Make a Big Difference? Bovine β-Casein A1 and A2 Variants and Human Health—An Update. Int. J. Mol. Sci. 2022, 23, 15637. [Google Scholar] [PubMed]
- IDF Diabetes Atlas 10th Edition. Available online: www.diabetesatlas.org (accessed on 25 December 2022).
- Rewers, M.; Ludvigsson, J. Environmental risk factors for type 1 diabetes. Lancet 2016, 387, 2340–2348. [Google Scholar] [CrossRef]
- Pociot, F.; Lernmark, Å. Genetic risk factors for type 1 diabetes. Lancet 2016, 387, 2331–2339. [Google Scholar] [CrossRef]
- Chia, J.S.J.; McRae, J.L.; Kukuljan, S.; Woodford, K.; Elliott, R.B.; Swinburn, B.; Dwyer, K.M. A1 beta-casein milk protein and other environmental pre-disposing factors for type 1 diabetes. Nutr. Diabetes 2017, 7, e274. [Google Scholar] [CrossRef] [PubMed]
- Harrison, L.C.; Honeyman, M.C. Cow’s milk and type 1 diabetes: The real debate is about mucosal immune function. Diabetes 1999, 48, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Regnell, S.E.; Lernmark, Å. Early prediction of autoimmune (type 1) diabetes. Diabetologia 2017, 60, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Navale, A.M.; Paranjape, A.N. Glucose transporters: Physiological and pathological roles. Biophys. Rev. 2016, 8, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Poppitt, S.D. Cow’s Milk and Dairy Consumption: Is There Now Consensus for Cardiometabolic Health? Front. Nutr. 2020, 7, 277. [Google Scholar] [CrossRef]
- Bhavadharini, B.; Dehghan, M.; Mente, A.; Rangarajan, S.; Sheridan, P.; Mohan, V.; Iqbal, R.; Gupta, R.; Lear, S.; Wentzel-Viljoen, E.; et al. Association of dairy consumption with metabolic syndrome, hypertension and diabetes in 147,812 individuals from 21 countries. BMJ Open Diabetes Res. Care 2020, 8, e000826. [Google Scholar] [CrossRef]
- Buziau, A.M.; Soedamah-Muthu, S.S.; Geleijnse, J.M.; Mishra, G.D. Total Fermented Dairy Food Intake Is Inversely Associated with Cardiovascular Disease Risk in Women. J. Nutr. 2019, 149, 1797–1804. [Google Scholar] [CrossRef]
- Awwad, S.F.; Abdalla, A.; Howarth, F.C.; Stojanovska, L.; Kamal-Eldin, A.; Ayyash, M.M. Invited review: Potential effects of short- and long-term intake of fermented dairy products on prevention and control of type 2 diabetes mellitus. J. Dairy Sci. 2022, 105, 4722–4733. [Google Scholar] [CrossRef]
- Mohamad, R.H.; Zekry, Z.K.; Al-Mehdar, H.A.; Salama, O.; El-Shaieb, S.E.; El-Basmy, A.A.; Al-Said, M.G.A.M.; Sharawy, S.M. Camel milk as an adjuvant therapy for the treatment of type 1 diabetes: Verification of a traditional ethnomedical practice. J. Med. Food 2009, 12, 461–465. [Google Scholar] [CrossRef]
- Korish, A.A. The antidiabetic action of camel milk in experimental type 2 diabetes mellitus: An overview on the changes in incretin hormones, insulin resistance, and inflammatory cytokines. Horm. Metab. Res. 2014, 46, 404–411. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Budania, S.; Sharma, P.; Gupta, R.; Kochar, D.K.; Panwar, R.B.; Sahani, S.M. Zero prevalence of diabetes in camel milk consuming Raica community of north-west Rajasthan, India. Diabetes Res. Clin. Pract. 2007, 76, 290–296. [Google Scholar] [CrossRef]
- Bhat, D.K.; Kanga, U.; Kumar, N.; Agrawal, R.P.; Mourya, M.; Kalaivani, M.; Kaur, T.; Mehra, N.K. The Raikas—A unique combination of high prevalence of type 1 diabetes susceptibility genes and near zero incidence of the disease. Hum. Immunol. 2014, 75, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Mirmiran, P.; Ejtahed, H.S.; Angoorani, P.; Eslami, F.; Azizi, F. Camel Milk Has Beneficial Effects on Diabetes Mellitus: A Systematic Review. Int. J. Endocrinol. Metab. 2017, 15, 42150. [Google Scholar] [CrossRef] [PubMed]
- Mihic, T.; Rainkie, D.; Wilby, K.J.; Pawluk, S.A. The Therapeutic Effects of Camel Milk: A Systematic Review of Animal and Human Trials. J. Evid. Based Complement. Altern. Med. 2016, 21, 110–126. [Google Scholar] [CrossRef]
- Mansour, A.A.; Nassan, M.A.; Saleh, O.M.; Soliman, M.M. Protective effect of camel milk as anti-diabetic supplement: Biochemical, molecular and immunohistochemical study. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Fallah, Z.; Feizi, A.; Hashemipour, M.; Kelishadi, R. Effect of fermented camel milk on glucose metabolism, insulin resistance, and inflammatory biomarkers of adolescents with metabolic syndrome: A double-blind, randomized, crossover trial. J. Res. Med. Sci. 2018, 23, 32. [Google Scholar]
- Ayoub, M.A.; Palakkott, A.R.; Ashraf, A.; Iratni, R. The molecular basis of the anti-diabetic properties of camel milk. Diabetes Res. Clin. Pract. 2018, 146, 305–312. [Google Scholar] [CrossRef]
- Shori, A.B. Camel milk as a potential therapy for controlling diabetes and its complications: A review of in vivo studies. J. Food Drug Anal. 2015, 23, 609–618. [Google Scholar] [CrossRef]
- Malik, A.; Al-Senaidy, A.; Skrzypczak-Jankun, E.; Jankun, J. A study of the anti-diabetic agents of camel milk. Int. J. Mol. Med. 2012, 30, 585–592. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Cadamuro, C.; le Gouic, A.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl peptidase IV (DPP-IV) inhibitory properties of a camel whey protein enriched hydrolysate preparation. Food Chem. 2019, 279, 70–79. [Google Scholar] [CrossRef]
- Brandelli, A.; Daroit, D.J.; Corrêa, A.P.F. Whey as a source of peptides with remarkable biological activities. Food Res. Int. 2015, 73, 149–161. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Isolation and characterization of peptides with dipeptidyl peptidase-IV inhibitory activity from pepsin-treated bovine whey proteins. Peptides 2014, 54, 39–48. [Google Scholar] [CrossRef]
- Giannetti, A.; Toschi Vespasiani, G.; Ricci, G.; Miniaci, A.; di Palmo, E.; Pession, A. Cow’s Milk Protein Allergy as a Model of Food Allergies. Nutrients 2021, 13, 1525. [Google Scholar] [CrossRef]
- Polidori, P.; Vincenzetti, S. Use of Donkey Milk in Children with Cow’s Milk Protein Allergy. Foods 2013, 2, 151–159. [Google Scholar] [CrossRef]
- Flom, J.D.; Sicherer, S.H. Epidemiology of Cow’s Milk Allergy. Nutrients 2019, 11, 1051. [Google Scholar] [CrossRef]
- Geiselhart, S.; Podzhilkova, A.; Hoffmann-Sommergruber, K. Cow’s Milk Processing—Friend or Foe in Food Allergy? Foods 2021, 10, 572. [Google Scholar] [CrossRef]
- Bernard, H.; Meisel, H.; Creminon, C.; Wal, J.M. Post-translational phosphorylation affects the IgE binding capacity of caseins. FEBS Lett. 2000, 467, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Schlimme, E.; Meisel, H. Bioactive peptides derived from milk proteins. Structural, physiological and analytical aspects. Nahrung 1995, 39, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Caira, S.; Pizzano, R.; Picariello, G.; Pinto, G.; Cuollo, M.; Chianese, L. Allergenicity of Milk Proteins. Milk Protein. 2012, 12, 2012. [Google Scholar]
- Jenkins, J.A.; Breiteneder, H.; Mills, E.N.C. Evolutionary distance from human homologs reflects allergenicity of animal food proteins. J. Allergy Clin. Immunol. 2007, 120, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Katz, Y.; Goldberg, M.R.; Zadik-Mnuhin, G.; Leshno, M.; Heyman, E. Cross-sensitization between milk proteins: Reactivity to a “kosher” epitope? Isr. Med. Assoc. J. 2008, 10, 85–88. [Google Scholar] [PubMed]
- Monti, G.; Bertino, E.; Muratore, M.C.; Coscia, A.; Cresi, F.; Silvestro, L.; Fabris, C.; Fortunato, D.; Giuffrida, M.G.; Conti, A. Efficacy of donkey’s milk in treating highly problematic cow’s milk allergic children: An in vivo and in vitro study. Pediatr. Allergy Immunol. 2007, 18, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Ehlayel, M.S.; Hazeima, K.A.; Al-Mesaifri, F.; Bener, A. Camel milk: An alternative for cow’s milk allergy in children. Allergy Asthma Proc. 2011, 32, 255–258. [Google Scholar] [CrossRef]
- Di Francesco, L.; Di Girolamo, F.; Mennini, M.; Masotti, A.; Salvatori, G.; Rigon, G.; Signore, F.; Pietrantoni, E.; Scapaticci, M.; Lante, I.; et al. A MALDI-TOF MS Approach for Mammalian, Human, and Formula Milks’ Profiling. Nutrients 2018, 10, 1238. [Google Scholar] [CrossRef] [PubMed]
- Maryniak, N.Z.; Hansen, E.B.; Ballegaard, A.S.R.; Sancho, A.I.; Bøgh, K.L. Comparison of the Allergenicity and Immunogenicity of Camel and Cow’s Milk—A Study in Brown Norway Rats. Nutrients 2018, 10, 1903. [Google Scholar] [CrossRef]
- Vojdani, A.; Turnpaugh, C.; Vojdani, E. Immune reactivity against a variety of mammalian milks and plant-based milk substitutes. J. Dairy Res. 2018, 85, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, C.; Martin, P.; Candalh, C.; Fauquant, J.; Piot, M.; Roucayrol, A.M.; Pilla, F.; Heyman, M. Goats’ milk of defective alpha(s1)-casein genotype decreases intestinal and systemic sensitization to beta-lactoglobulin in guinea pigs. J. Dairy Res. 2001, 68, 217–227. [Google Scholar] [CrossRef]
- Hodges, H.; Fealko, C.; Soares, N. Autism spectrum disorder: Definition, epidemiology, causes, and clinical evaluation. Transl. Pediatr. 2020, 9 (Suppl. 1), S55–S65. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.J.; Brugha, T.S.; Erskine, H.E.; Scheurer, R.W.; Vos, T.; Scott, J.G. The epidemiology and global burden of autism spectrum disorders. Psychol. Med. 2015, 45, 601–613. [Google Scholar] [CrossRef]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Rosenberg, C.R.; White, T.; et al. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. MMWR Surveill. Summ. 2018, 67, 1–23. [Google Scholar] [CrossRef]
- Tailford, K.A.; Berry, C.L.; Thomas, A.C.; Campbell, J.H. A casein variant in cow’s milk is atherogenic. Atherosclerosis 2003, 170, 13–19. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, C.N.S. Beta-casein A1, ischaemic heart disease mortality, and other illnesses. Med. Hypotheses 2001, 56, 262–272. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Total Casein Concentration (g/L Milk) | Relative Casein Composition (%) | Mean Micelle Diameter (nm) | ||||
|---|---|---|---|---|---|---|---|
| αS1 | αS2 | β | κ | αS1 + β | |||
| African elephant | Unknown | - | - | 89 | 11 | 89 | 350–700 |
| Buffalo | 32–40 | 40 | 9 | 35 | 12 | 75 | 190 |
| Cow | 24.6–28 | 38 | 10 | 40 | 12 | 78 | 150–182 |
| Camel | 22.1–26.0 | 22 | 9 | 65.5 | 3.5 | 87 | 380 |
| Goat | 23.3–46.3 | 20 | 16 | 41 | 17 | 61 | 180–300 |
| Horse | 9.4–13.6 | 17.7 | 1.5 | 79 | 1.8 | 96 | 255 |
| Human | 2.4–4.2 | 3 | - | 70 | 27 | 73 | 64–80 |
| Sheep | 41.8–52.6 | 50 | - | 40 | 10 | 90 | 180–210 |
| Range | - | 3–55 | 9–28 | 26–89 | 3.5–27 | 61–96 | - |
| Alpha-s1 | Cow | Camel | Alpha-s2 | Cow | |||
|---|---|---|---|---|---|---|---|
| Camel | 50.24 | Camel | 63.16 | ||||
| Human | 39.66 | 47.75 | |||||
| beta- | Cow | Camel | Human | kappa- | Cow | Camel | Human |
| Camel | 66.82 | Camel | 60.22 | ||||
| Human | 56.07 | 62.9 | Human | 54.14 | 59.34 | ||
| Elephant | 54.76 | 55.3 | 62.61 | Elephant | 48.31 | 56.25 | 53.98 |
| Animal | Accession Number | Whole Protein | Mature Protein * | Signal Peptide (retrieved from the Uniprot Database (The UniProt Consortium, 2017) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sequence Length | Molecular Weight (KDa) | Theoretical pI | Instability Index | Sequence Length | Molecular Weight (KDa) | Theoretical pI | Instability Index | ||||
| αS1-Casein | |||||||||||
| 1 | Cow | P02662 | 214 | 24.5289 | 4.98 | 56.03 | 199 | 22.9748 | 4.91 | 57.99 | MKLLILTCLVAVALA |
| 2 | Camel | O97943 | 230 | 26.8614 | 4.96 | 64.07 | 215 | 25.3073 | 4.89 | 66.44 | MKLLILTCLVAVALA |
| 3 | Human | P47710 | 185 | 21.6710 | 5.32 | 70.90 | 170 | 20.0894 | 5.17 | 74.75 | MRLLILTCLVAVALA |
| αS2-Casein | |||||||||||
| 4 | Cow | P02663 | 222 | 26.0187 | 8.55 | 44.68 | 207 | 24.3485 | 8.34 | 46.27 | MKFFIFTCLLAVALA |
| 5 | Camel | O97944 | 193 | 22.9641 | 6.00 | 58.11 | 178 | 21.2659 | 5.80 | 61.08 | MKFFIFTCLLAVVLA |
| β-Casein | |||||||||||
| 6 | Cow | XP_010804480.2 | 259 | 29.22125 | 6.17 | 90.11 | 209 | 23.6924 | 5.38 | 97.56 | MKVLILACRVALALA |
| 7 | Camel | Q9TVD0 | 232 | 26.2178 | 5.62 | 96.58 | 217 | 24.6507 | 5.27 | 100.04 | MKVLILACRVALALA |
| 8 | Human | P05814 | 226 | 25.3810 | 5.52 | 73.18 | 211 | 23.8578 | 5.33 | 74.17 | MKVLILACLVALALA |
| 9 | Elephant | G3TDS4 | 223 | 25.2770 | 6.21 | 72.14 | 208 | 23.6995 | 5.97 | 73.06 | MKVFILACLVAFALG |
| κ-Casein | |||||||||||
| 10 | Cow | P02668 | 190 | 21.2693 | 6.29 | 54.21 | 169 | 18.9744 | 5.93 | 57.46 | MMKSFFLVVTILALTLPFLGA |
| 11 | Camel | P79139 | 182 | 20.4176 | 8.55 | 44.72 | 162 | 18.2538 | 8.03 | 46.49 | MKSFFLVVTILALTLPFLGA |
| 12 | Human | P07498 | 182 | 20.3010 | 8.97 | 56.48 | 162 | 18.1628 | 8.68 | 59.84 | MKSFLLVVNALALTLPFLAV |
| 13 | Elephant | XP_023408995.1 | 181 | 20.5520 | 8.45 | 48.56 | 161 | 18.3880 | 7.98 | 46.37 | MKGFLLVVNILLLPLPFLAA |
| Animal | Accession Number | Percentage Conformation (%) | Disorder (%) | |||
|---|---|---|---|---|---|---|
| α-Helixes/Exposed | β-Sheets/Medium | β-Turns/Buried | ||||
| αS1-Casein | ||||||
| 1 | Cow | P02662 | 38/64 | 1/14 | 59/21 | 68 |
| 2 | Camel | O97943 | 36/58 | 3/17 | 59/23 | 61 |
| 3 | Human | P47710 | 46/66 | 3/12 | 49/20 | 69 |
| αS2-Casein | ||||||
| 4 | Cow | P02663 | 48/70 | 0/14 | 50/14 | 57 |
| 5 | Camel | O97944 | 43/58 | 0/19 | 56/22 | 58 |
| β-Casein | ||||||
| 6 | Cow | XP_010804480.2 | 29/64 | 2/20 | 68/15 | 46 |
| 7 | Camel | Q9TVD0 | 26/64 | 1/22 | 71/13 | 46 |
| 8 | Human | P05814 | 23/57 | 1/27 | 75/14 | 38 |
| 9 | Elephant | G3TDS4 | 19/60 | 1/23 | 78/15 | 41 |
| κ-Casein | ||||||
| 10 | Cow | P02668 | 8/58 | 5/19 | 86/21 | 53 |
| 11 | Camel | P79139 | 6/52 | 7/19 | 86/28 | 37 |
| 12 | Human | P07498 | 6/48 | 6/24 | 86/26 | 26 |
| 13 | Elephant | XP_023408995.1 | 14/56 | 7/20 | 77/23 | 30 |
| Animal | Opioid Peptide | Amino Acid Sequence |
|---|---|---|
| Cow | β-casomorphin-4 | Tyr60-Pro-Phe-Pro63 (YPFP) |
| β-casomorphin-5 | Tyr60-Pro-Phe-Pro-Gly64 (YPFPG) | |
| β-casomorphin-6 | Tyr60-Pro-Phe-Pro-Gly-Pro65 (YPFPGP) | |
| β-casomorphin-7 | Tyr60-Pro-Phe-Pro-Gly-Pro-Ile66 (YPFPGPI) | |
| β-casomorphin-8 | Tyr60-Pro-Phe-Pro-Gly-Pro-Ile-Pro/His67 (YPFPGPIP/H) | |
| β-casomorphin-11 | Tyr60-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asn-Ser-Leu70 (YPFPGPIPNSL) | |
| k-casein-casoxin-A | Tyr35-Pro-Ser-Tyr-Gly-Leu-Asn-Tyr42 (YPSYGLNY) | |
| k-casein-casoxin-B | Tyr58-Pro-Tyr-Tyr61 (YPYY) | |
| k-casein-casoxin-C | Tyr25-Ile-Pro-Ile-Gln-Tyr-Val-Leu-Ser-Arg34 (YIPIQYVLSR) | |
| αS1-casein-exorphin | Arg90-Tyr-Leu-Gly-Tyr-Leu-Glu96 (RYLGYLE) | |
| Human | β-casorphin-4 | Tyr41-Pro-Ser-Phe444 (YPSF) |
| β-casomorphin-4 | Tyr51-Pro-Phe-Val54 (YPFV) | |
| β-casomorphin-5 | Tyr51-Pro-Phe-Val-Glu555 (YPFVE) | |
| β-casomorphin-7 | Tyr51-Pro-Phe-Val-Glu-Pro-Ile57 (YPFVEPI) | |
| β-casomorphin-8 | Tyr51-Pro-Phe-Val-Glu-Pro-Ile-Pro58 (YPFVEPIP) | |
| k-casein-casoxin-D | Tyr158-Val-Pro-Phe-Pro-Pro-Phe164 (YVPFPPF) |
| Casein | Peptide | Bioactivity |
|---|---|---|
| κ-Casein | 103LSFMAIPPK111 | Antithrombotic |
| 108IPP110 | ACE-inhibitory | |
| 106MAIPPKK112 | Antithrombotic | |
| 106MAIPPKKNQDK116 | Antithrombotic | |
| 113NQDK116 | Antithrombotic | |
| β-Casein | 1RELEELNVPGEIVESLSSSEESITR25 | Calcium-binding |
| 1RELEELNVPGEIVESLSSSEESITRINK28 | Immunomodulatory, calcium-binding | |
| 2ELEELNVPGEIVESLSSSEESITRINK28 | Calcium-binding | |
| 63PGPIPN68 | Immunomodulatory | |
| 191LLY193 | Immunomodulatory | |
| 193LLYQEPVLGPVRGPFPIIV209 | Immunomodulatory | |
| 74IPP76 | ACE-inhibitory | |
| 84VPP86 | ACE-inhibitory | |
| 108EMPFPK113 | ACE-inhibitory | |
| 177AVPYPQR183 | ACE-inhibitory | |
| 193YQEPVL198 | ACE-inhibitory | |
| 193YQEPVLGPVRGPFPI202 | ACE-inhibitory | |
| 193YQEPVLGPVRGPFPIIV209 | Antimicrobail | |
| 199GPVRGPFPIIV204 | ACE-inhibitory | |
| αS1-Casein | 1RPKHPIKHQGLPQEVLNENLLRF23 | Immunomodulatory, antimicrobial |
| 194TTMPLW199 | Immunomodulatory | |
| 23FF24 | ACE-inhibitory | |
| 23FFVAP27 | ACE-inhibitory | |
| 43DIGSESTEDQAMEDIK58 | Calcium-binding | |
| 45GSESTEDQAME55 | Calcium-binding | |
| 59QMEAESISSSEEIVPNSVEQK79 | Calcium-binding | |
| 66SSSEEIVPN74 | Calcium-binding | |
| 102KKYKVPQ109 | ACE-inhibitory | |
| 106VPQLEIVPNSAEER119 | Calcium-binding | |
| 142LAYFYP147 | ACE-inhibitory | |
| 157DAYPSGAW164 | ACE-inhibitory | |
| 194TTMPLW199 | ACE-inhibitory | |
| αS2-Casein | 1KNTMEHVSSSEESIISQETYKQEKNMAINPSK32 | Immunomodulatory |
| 2NTMEHVSSSEESIISQETYK21 | Calcium-binding | |
| 46NANEEEYSIGSSSEESAEVATEEVK70 | Calcium-binding | |
| 55GSSSEESAEVATEEVKITVDD75 | Calcium-binding | |
| 126EQLSTSEENSK136 | Calcium-binding | |
| 138TVDMESTEVFTK149 | Calcium-binding | |
| 164LKKISQRYQKFALPQY179 | Antimicrobial | |
| 165KKISQRYQKFALPQYLKTVYQHQKAMKPWIQPKTKVIPY203 | Antimicrobial | |
| 174FALPQY179 | ACE-inhibitory | |
| 174FALPQYLK181 | ACE-inhibitory | |
| 183VYQHQKAMKPWIQPKTKVIPYVRYL207 | Antimicrobial | |
| 189AMKPW193 | ACE-inhibitory | |
| 189AMKPWIQPK197 | ACE-inhibitory | |
| 190MKPWIQPK197 | ACE-inhibitory | |
| 198TKVIP202 | ACE-inhibitory | |
| Camel αs1-CN | 4DNLMPQFM8 | DPP-IV binding |
| 1WNWGWLLWQL9 | DPP-IV binding | |
| 1TF2 | ACE-inhibitory | |
| 1LxV3 | Glycose intake | |
| Human αs1-CN | 1YPER18 | ACE-inhibitory |
| 136YYPQIMQY143 | ACE-inhibitory | |
| 164NNVMLQW170 | ACE-inhibitory | |
| Human β-CN | 169VPYPQ173 | Antioxidant |
| 154WSVPQPK160 | Antioxidant | |
| 54VEPIPY59 | Immunostimulatin | |
| Human κ-CN | 31YPNSYP36 | Antioxidant |
| 53NPYVPR58 | Antioxidant |
| Milk Proteins | Identified Allergic Epitope Peptide Fractions |
|---|---|
| Casein proteins (Bos d 8) | The whole casein fraction |
| αS1-casein (Bos d 9) | 16–35, 17–36, 19–30, 39–48, 69–78, 93–98, 93–102, 109–120, 123–132, 139–154, 141–150, 159–174, and 173–194. The whole molecule and the larger fractions have the highest effect |
| αS2-casein (Bos d 10) | 31–44, 43–56, 83–100, 93–108, 105–114, 117–128, 143–158, 157–172, 165–188, and 191–200 |
| β-casein (Bos d 11) | 1–16, 45–54, 55–70, 83–92, 107–120, 135–144, 149–164, 167–184, and 185–208 |
| κ-casein (Bos d 12) | 15–24, 37–46, 55–80, 83–92, and 105–116 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Runthala, A.; Mbye, M.; Ayyash, M.; Xu, Y.; Kamal-Eldin, A. Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk. Molecules 2023, 28, 2023. https://doi.org/10.3390/molecules28052023
Runthala A, Mbye M, Ayyash M, Xu Y, Kamal-Eldin A. Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk. Molecules. 2023; 28(5):2023. https://doi.org/10.3390/molecules28052023
Chicago/Turabian StyleRunthala, Ashish, Mustapha Mbye, Mutamed Ayyash, Yajun Xu, and Afaf Kamal-Eldin. 2023. "Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk" Molecules 28, no. 5: 2023. https://doi.org/10.3390/molecules28052023
APA StyleRunthala, A., Mbye, M., Ayyash, M., Xu, Y., & Kamal-Eldin, A. (2023). Caseins: Versatility of Their Micellar Organization in Relation to the Functional and Nutritional Properties of Milk. Molecules, 28(5), 2023. https://doi.org/10.3390/molecules28052023